
Circulatory Endothelin 1-Regulating RNAs Panel: Promising Biomarkers for Non-Invasive NAFLD/NASH Diagnosis and Stratification: Clinical and Molecular Pilot Study

 , and
, and
Abstract
:1. Introduction
2. Results
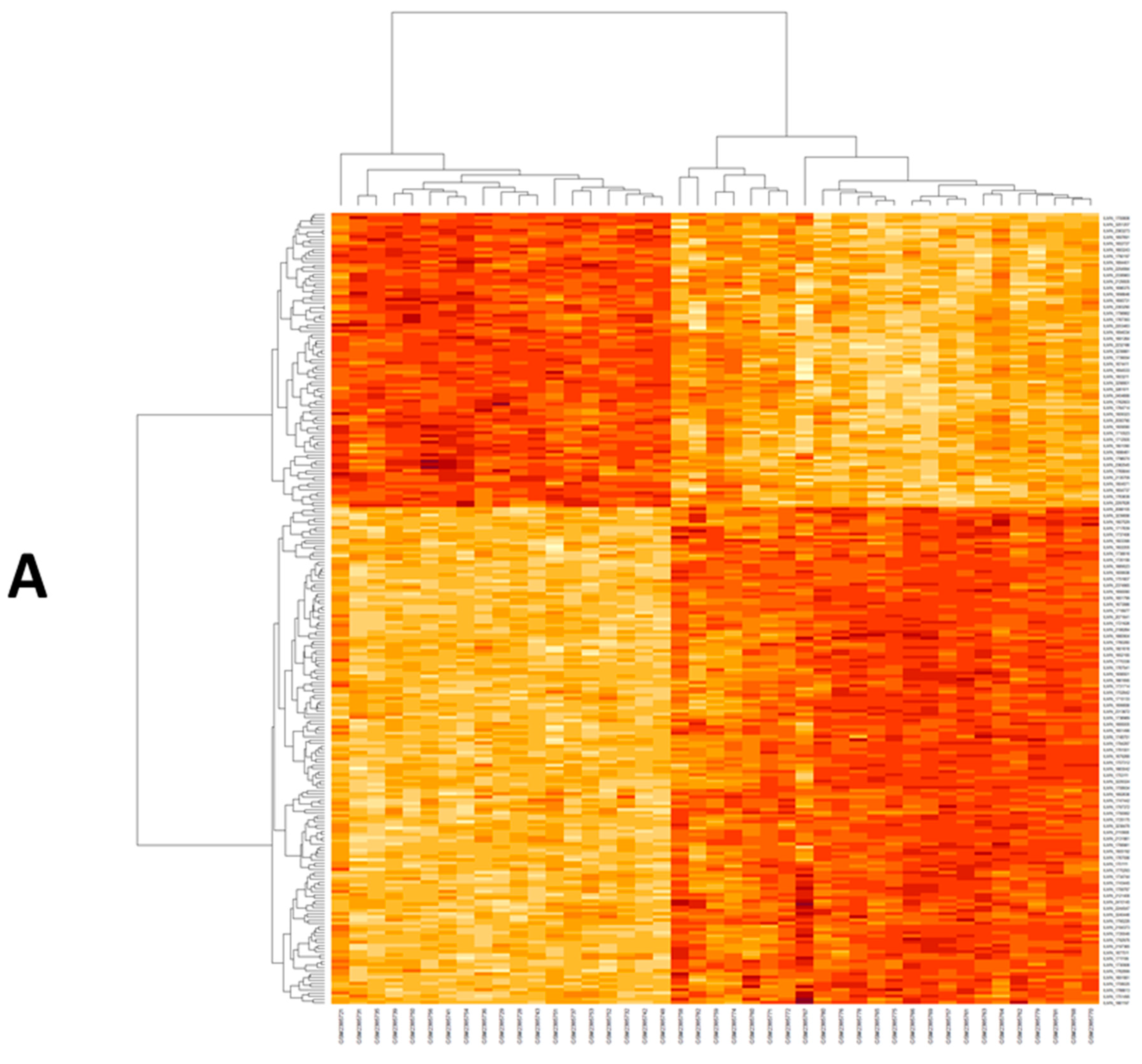
2.1. Retrieval of Differentially Expressed mRNAs (DEG) from GEO Data Set
2.2. Analysis of Biochemical and Clinical Parameters in NAFLD/NASH
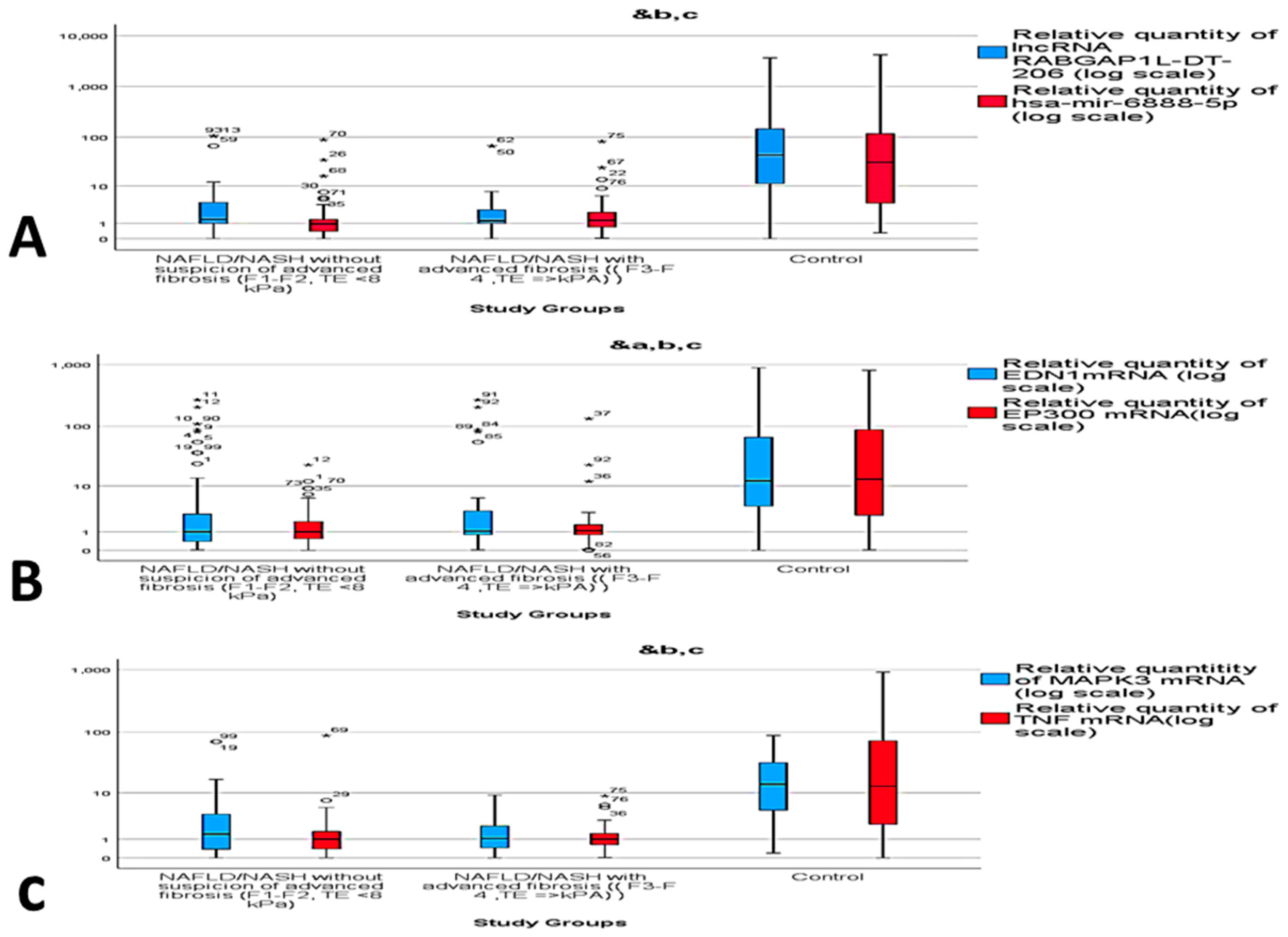
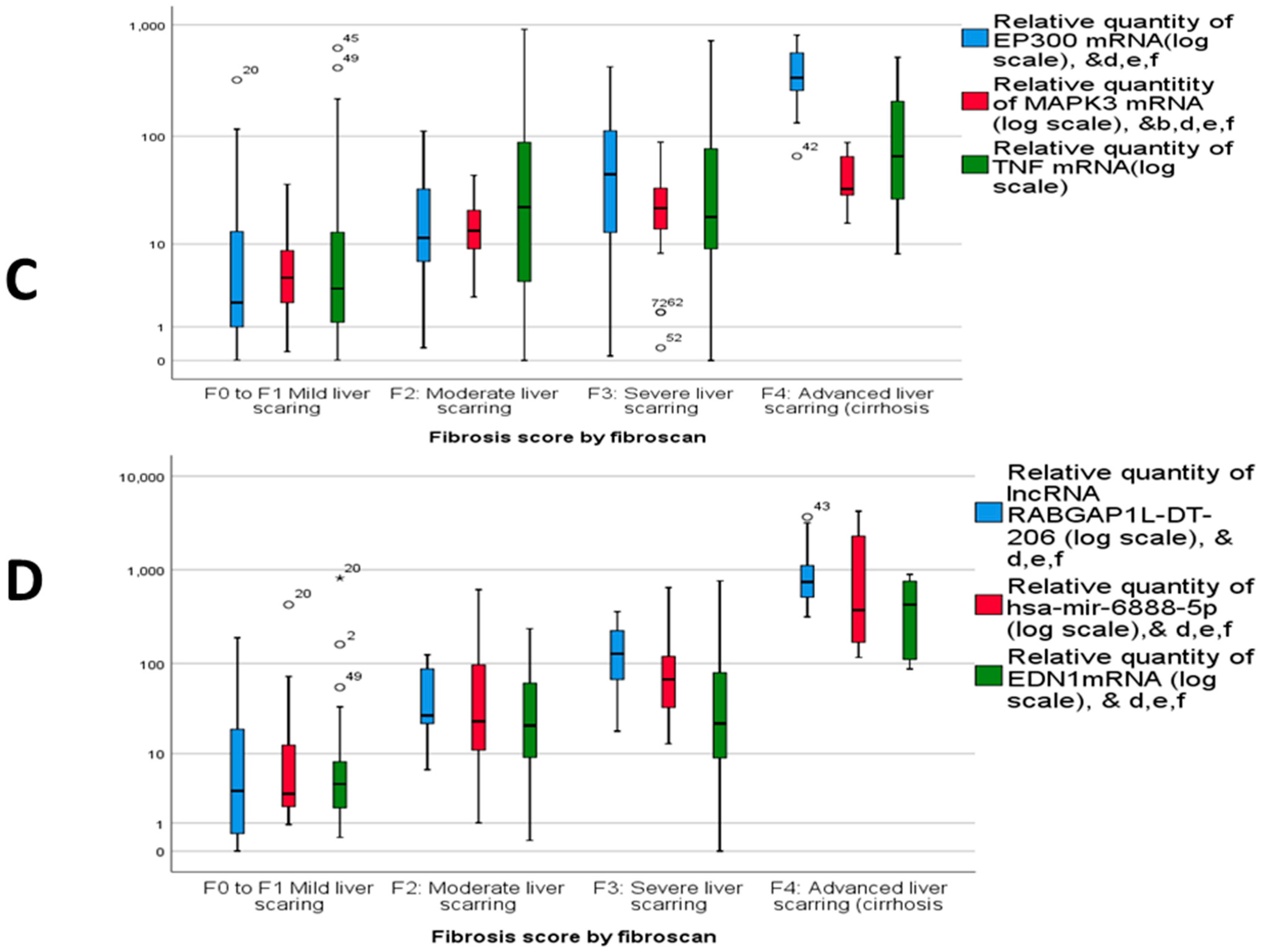
2.3. Dysregulated mRNA/miRNA/lncRNA Axis Expression in NAFLD/NASH
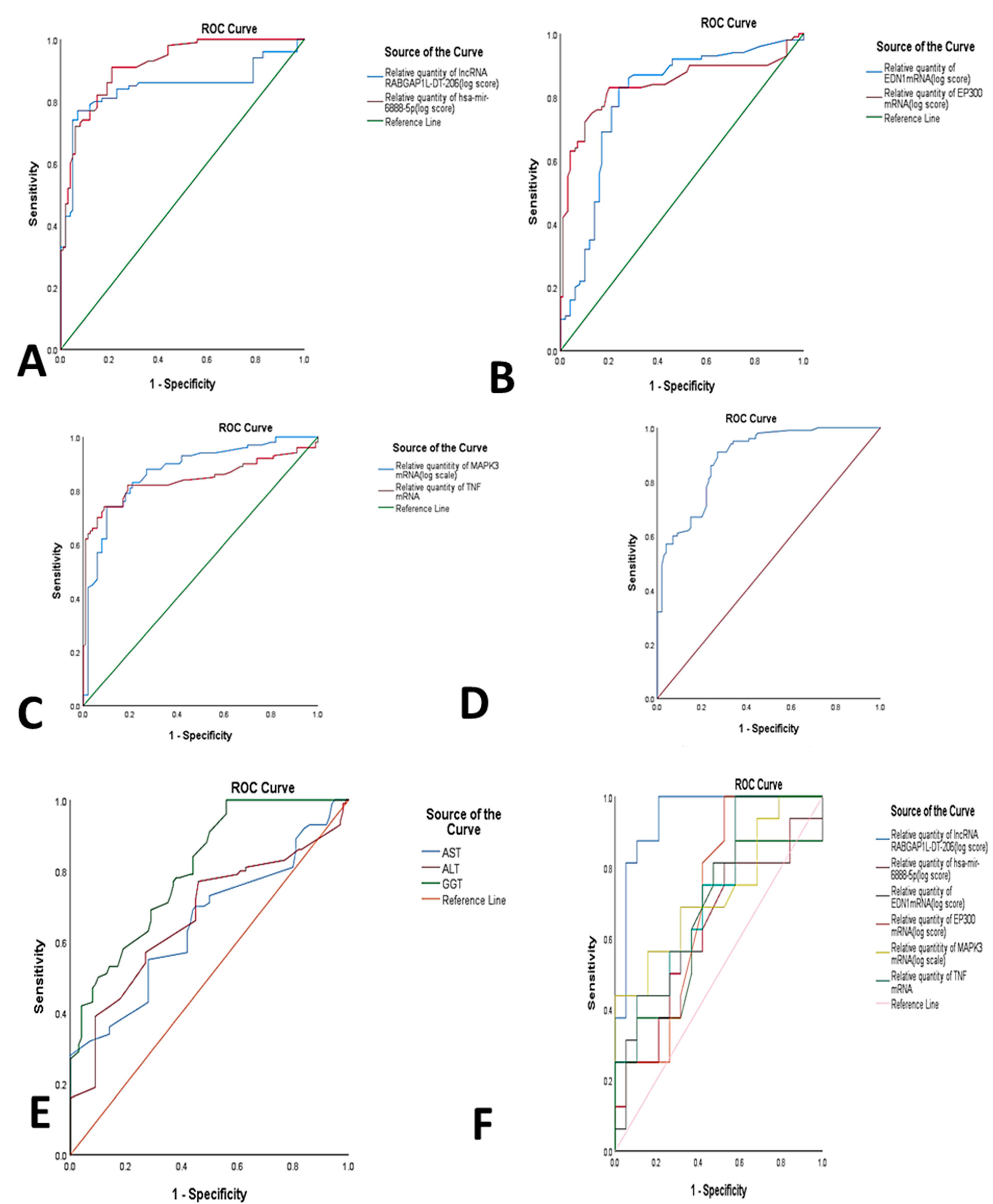
2.4. Diagnostic Performance of RNAs Panel in NASH
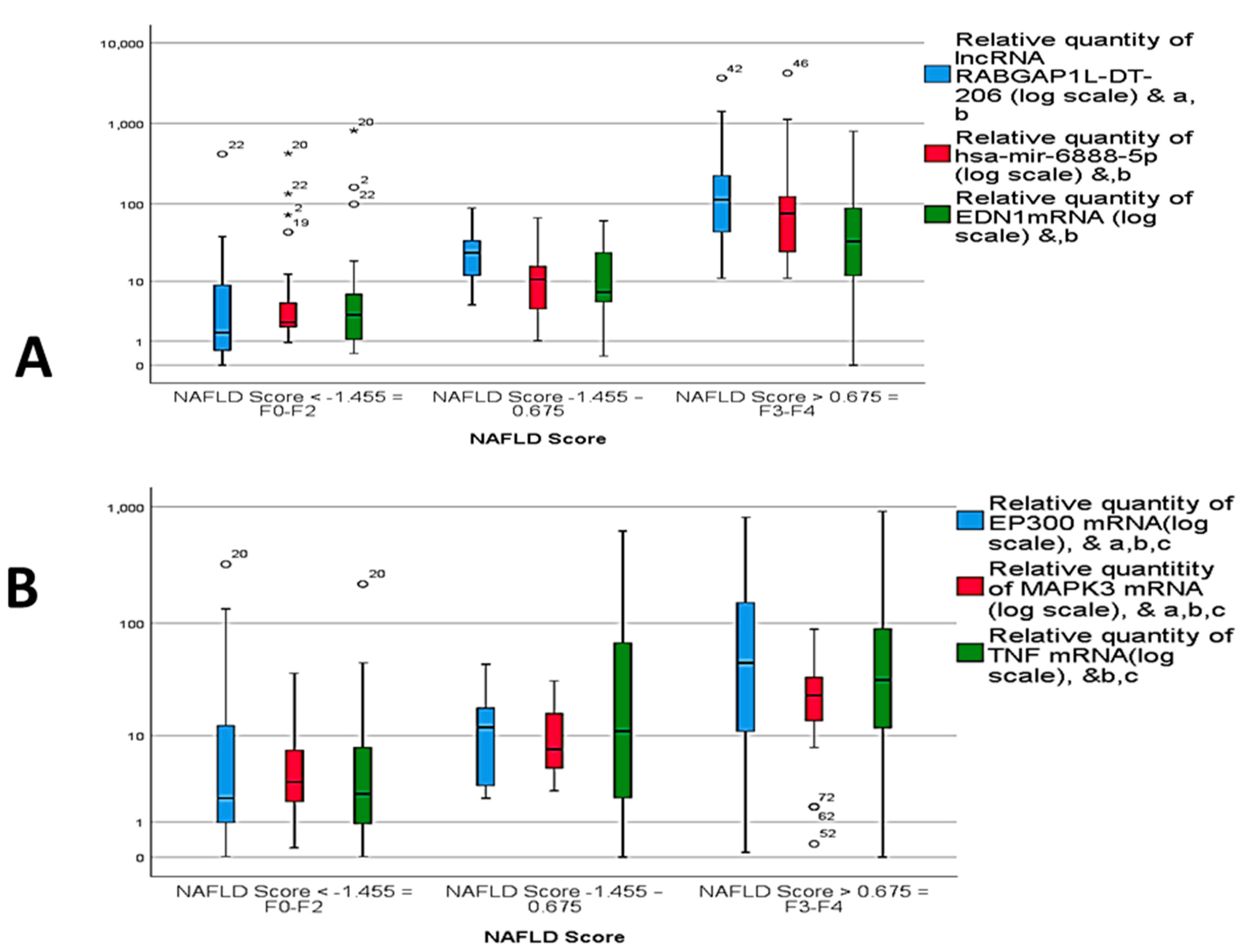
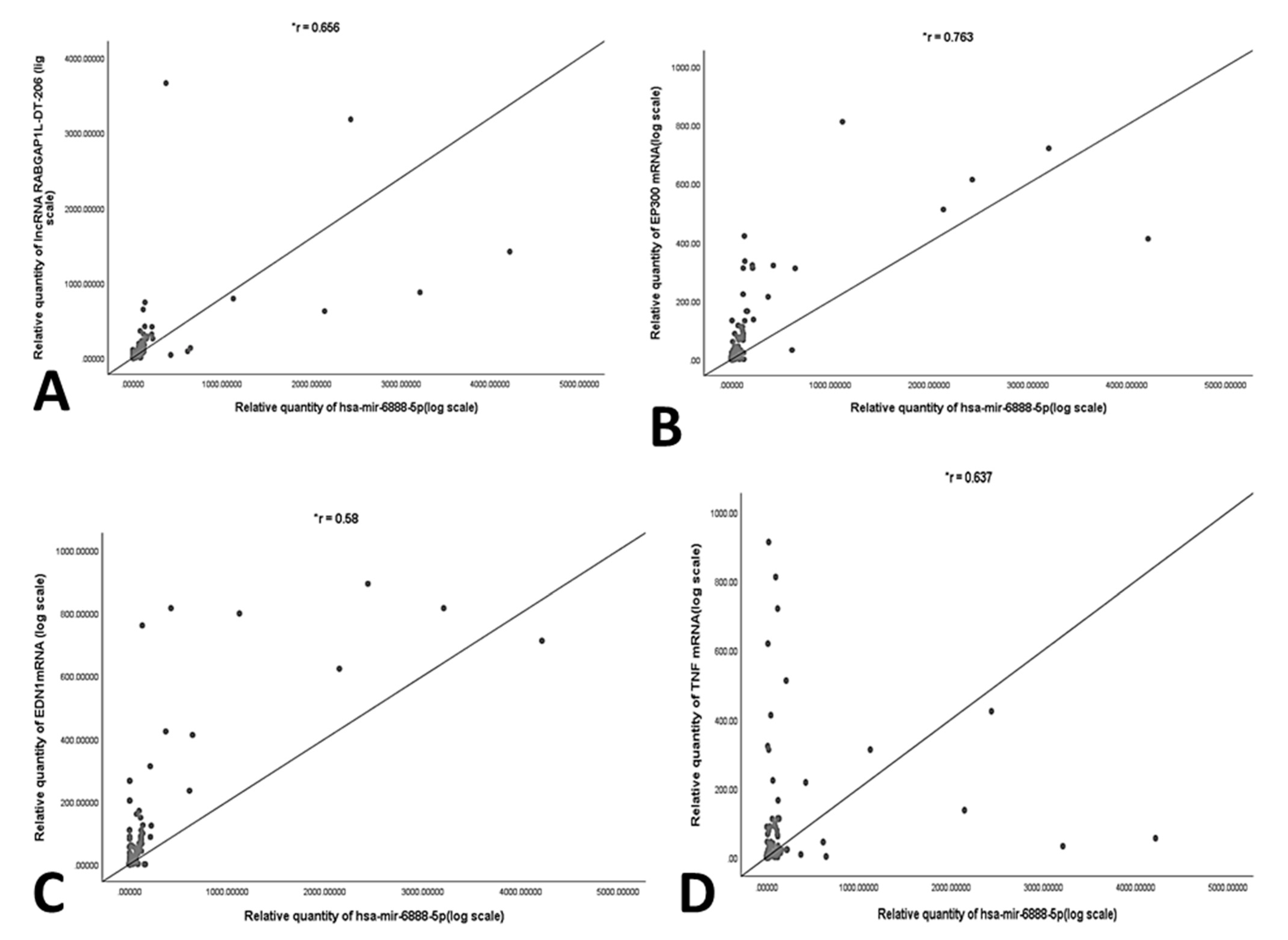
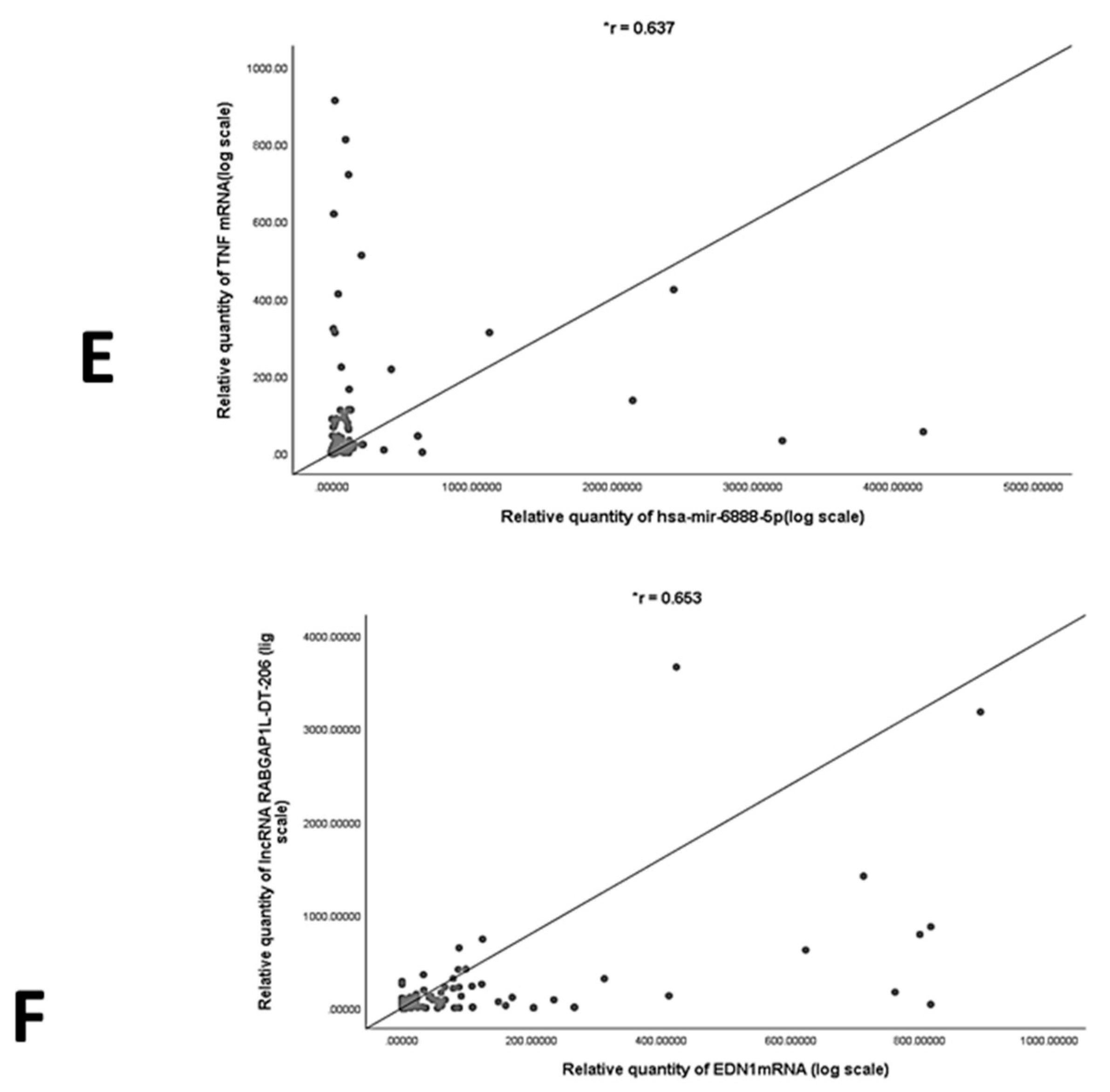
2.5. Correlation Analysis and Multivariate Regression Analysis of NASH Predictors
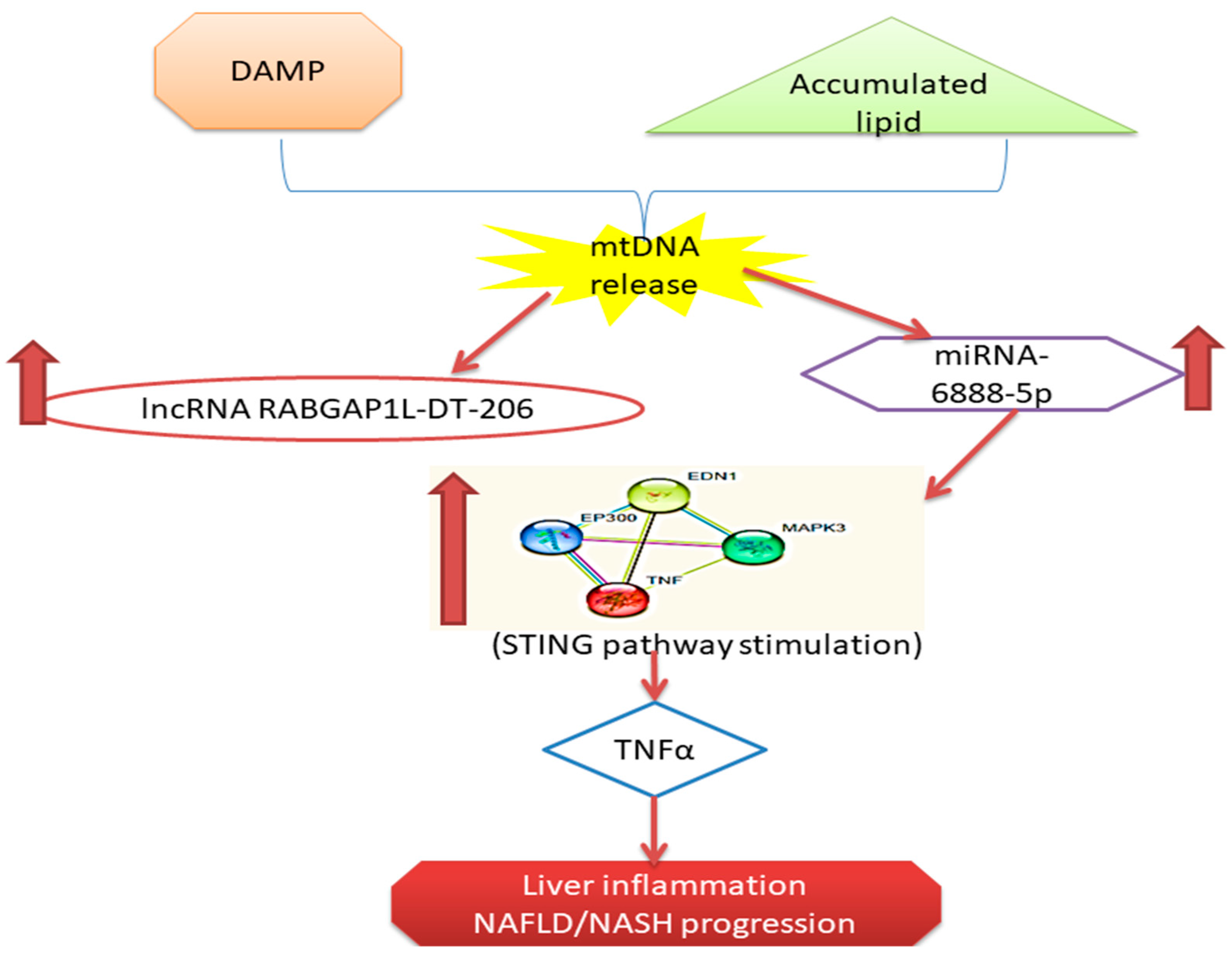
3. Discussion
4. Materials and Methods
4.1. Biomarker Filtration of mRNA-miRNA-lncRNA Panel from Public Microarray Database
4.2. Study Subjects
4.3. Total RNA Extraction and Quantitative Real Time PCR (RT-qPCR)
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dongwei, X.; Yizhu, T.; Qiang, X.; Bibo, K. The cGAS-STING Pathway: Novel Perspectives in Liver Diseases. Front. Immunol. 2021, 12, 1569. [Google Scholar]
- Bessone, F.; Razori, M.V.; Roma, M.G. Molecular pathways of nonalcoholic fatty liver disease development and progression. Cell. Mol. Life Sci. 2019, 76, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Piazzolla, V.A.; Mangia, A. Noninvasive Diagnosis of NAFLD and NASH. Cells 2020, 9, 1005. [Google Scholar] [CrossRef] [PubMed]
- Calzadilla Bertot, L.; Adams, L.A. The Natural Course of Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2016, 17, 774. [Google Scholar] [CrossRef] [Green Version]
- Kazankov, K.; Jørgensen, S.M.D.; Thomsen, K.L.; Møller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Grønbæk, H. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 145–159. [Google Scholar] [CrossRef]
- Mao, Y.; Luo, W.; Zhang, L.; Wu, W.; Yuan, L.; Xu, H.; Song, J.; Fujiwara, K.; Abe, J.-I.; Lemaire, S.A.; et al. STING–IRF3 Triggers Endothelial Inflammation in Response to Free Fatty Acid-Induced Mitochondrial Damage in Diet-Induced Obesity. Arter. Thromb. Vasc. Biol. 2017, 37, 920–929. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.; Cui, C.; Qing, L.; Wang, L.; He, T.; Yan, F.; Liu, F.; Shen, Y.; Hou, X.; Chen, L. Activation of the STING-IRF3 pathway promotes hepatocyte inflammation, apoptosis and induces metabolic disorders in nonalcoholic fatty liver disease. Metabolism 2018, 81, 13–24. [Google Scholar] [CrossRef]
- Luo, X.; Li, H.; Ma, L.; Zhou, J.; Guo, X.; Woo, S.-L.; Pei, Y.; Knight, L.R.; Deveau, M.; Chen, Y.; et al. Expression of STING Is Increased in Liver Tissues From Patients With NAFLD and Promotes Macrophage-Mediated Hepatic Inflammation and Fibrosis in Mice. Gastroenterology 2018, 155, 1971–1984e4. [Google Scholar] [CrossRef] [Green Version]
- Ganz, M.; Szabo, G. Immune and inflammatory pathways in NASH. Hepatol. Int. 2013, 7, S771–S781. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Scott, M.J.; Fan, J.; Billiar, T.R. Location is the key to function: HMGB1 in sepsis and trauma-induced inflammation. J. Leukoc. Biol. 2019, 106, 161–169. [Google Scholar] [CrossRef]
- Li, L.; Chen, L.; Hu, L.; Liu, Y.; Sun, H.-Y.; Tang, J.; Hou, Y.-J.; Chang, Y.-X.; Tu, Q.-Q.; Feng, G.-S.; et al. Nuclear factor high-mobility group box1 mediating the activation of toll-like receptor 4 signaling in hepatocytes in the early stage of nonalcoholic fatty liver disease in mice. Hepatology 2011, 54, 1620–1630. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Liu, Z.; Xu, Z.; Liu, J.; Zhang, J. High mobility group box 1 (HMGB1): A pivotal regulator of hematopoietic malignancies. J. Hematol. Oncol. 2020, 13, 91. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Hou, W.; Zhang, Q.; Kang, R.; Fan, X.-G.; Tang, D. Emerging Role of High-Mobility Group Box 1 (HMGB1) in Liver Diseases. Mol. Med. 2013, 19, 357–366. [Google Scholar] [CrossRef]
- Chandrashekaran, V.; Seth, R.; Dattaroy, D.; Alhasson, F.; Ziolenka, J.; Carson, J.; Berger, F.G.; Kalyanaraman, B.; Diehl, A.M.; Chatterjee, S. HMGB1-RAGE pathway drives peroxynitrite signaling-induced IBD-like inflammation in murine nonalcoholic fatty liver disease. Redox Biol. 2017, 13, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, I.; Santoro, N.; Chen, Y.; Hoque, R.; Ouyang, X.; Caprio, S.; Shlomchik, M.J.; Coffman, R.L.; Candia, A.; Mehal, W.Z. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of TLR. J. Clin. Investig. 2016, 126, 859–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Liu, Y.; An, W.; Song, J.; Zhang, Y.; Zhao, X. STING-mediated inflammation in Kupffer cells contributes to progression of nonalcoholic steatohepatitis. J. Clin. Investig. 2018, 129, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Baffy, G. MicroRNAs in Nonalcoholic Fatty Liver Disease. J. Clin. Med. 2015, 4, 1977–1988. [Google Scholar] [CrossRef] [Green Version]
- Gerhard, G.S.; Di Stefano, J.K. Micro RNAs in the development of non-alcoholic fatty liver disease. World J. Hepatol. 2015. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Hu, C.; Jia, W. miRNAs in non-alcoholic fatty liver disease. Front. Med. 2016, 10, 389–396. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, H.; Xu, C.; Yu, C.; Li, Y. Long Non-Coding RNA Profiling in a Non-Alcoholic Fatty Liver Disease Rodent Model: New Insight into Pathogenesis. Int. J. Mol. Sci. 2017, 18, 21. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, B.; Bellet, M.M.; Katada, S.; Astarita, G.; Hirayama, J.; Amin, R.H.; Granneman, J.G.; Piomelli, D.; Leff, T.; Sassone-Corsi, P. PER2 Controls Lipid Metabolism by Direct Regulation of PPARγ. Cell Metab. 2010, 12, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Chi, X.; Qu, N.; Wang, C. Long noncoding RNA lncARSR promotes hepatic lipogenesis via Akt/SREBP-1c pathway and contributes to the pathogenesis of nonalcoholic steatohepatitis. Biochem. Biophys. Res. Commun. 2018, 499, 66–70. [Google Scholar] [CrossRef]
- Wruck, W.; Graffmann, N.; Kawala, M.-A.; Adjaye, J. Concise Review: Current Status and Future Directions on Research Related to Nonalcoholic Fatty Liver Disease. Stem Cells 2016, 35, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Barber, G.N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef]
- Petrasek, J.; Iracheta-Vellve, A.; Csak, T.; Satishchandran, A.; Kodys, K.; Kurt-Jones, E.A.; Fitzgerald, K.; Szabo, G. STING-IRF3 pathway links endoplasmic reticulum stress with hepatocyte apoptosis in early alcoholic liver disease. Proc. Natl. Acad. Sci. USA 2013, 110, 16544–16549. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Cervantes, C.; Liu, J.; He, S.; Zhou, H.; Zhang, B.; Cai, H.; Yin, D.; Hu, D.; Li, Z.; et al. DsbA-L prevents obesi-ty-induced inflammation and insulin resistance by suppressing the mtDNA release-activated cGAS-cGAMP-STING path-way. Proc. Natl. Acad. Sci. USA 2017, 114, 12196–12201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerger, A.; El-Khoueiry, A.; Zhang, W.; Yang, D.; Singh, H.; Bohanes, P.; Ning, Y.; Winder, T.; LaBonte, M.J.; Wilson, P.M.; et al. Phar-macogenetic angiogenesis profiling for first-line Bevacizumab plus oxaliplatin-based chemotherapy in patients with meta-static colorectal cancer. Clin. Cancer Res. 2011, 17, 5783–5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davenport, A.P.; Hyndman, K.A.; Dhaun, N.; Southan, C.; Kohan, D.E.; Pollock, J.S.; Pollock, D.M.; Webb, D.J.; Maguire, J.J.; Barker, E.L.; et al. Endothelin. Pharmacol. Rev. 2016, 68, 357–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitts, K.R. Endothelin receptor antagonism in portal hypertension. Expert Opin. Investig. Drugs 2009, 18, 135–142. [Google Scholar] [CrossRef]
- Farina, G.; York, M.; Collins, C.; Lafyatis, R. dsRNA activation of endothelin-1 and markers of vascular activation in endothelial cells and fibroblasts. Ann. Rheum. Dis. 2010, 70, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Miren, B.; Raurell, I.; Hide, D.; Fernández-Iglesias, A.; Gil, M.; Barberá, A.; Salcedo, M.T.; Augustin, S.; Genescà, J.; Martell, M. Restoration of liver sinusoidal cell phenotypes by statins improves por-tal hypertension and histology in rats with NASH. Sci. Rep. 2019, 9, 1–12. [Google Scholar]
- Bulent, D.; Ozenirler, S.; Elbeg, S.; Akyol, G. The Serum Endothelın-1 Level in Steatosıs and NASH, and Its Relatıon wıth Severıty of Lıver Fıbrosıs. Dig. Dis. Sci. 2007, 52, 2622–2628. [Google Scholar]
- Chan, H.M.; La Thangue, N.B. p300/CBP proteins: HATs for transcriptional bridges and scaffolds. J. Cell Sci. 2001, 114, 2363–2373. [Google Scholar] [CrossRef] [PubMed]
- Dancy, B.M.; Cole, P.A. Protein lysine acetylation by p300/CBP. Chem. Rev. 2015, 115, 2419–2452. [Google Scholar] [CrossRef]
- Gee, C.T.; Koleski, E.J.; Pomerantz, W.C.K. Fragment Screening and Druggability Assessment for the CBP/p300 KIX Domain through Protein-Observed19F NMR Spectroscopy. Angew. Chem. Int. Ed. 2015, 54, 3735–3739. [Google Scholar] [CrossRef] [Green Version]
- Martín-Antonio, B.; Álvarez-Laderas, I.; Cardesa, R.; Márquez-Malaver, F.; Baez, A.; Carmona, M.; Falantes, J.; Suarez-Lledo, M.; Fernández-Avilés, F.; Martínez, C.; et al. A constitutional variant in the transcription factor EP300 strongly influences the clinical outcome of patients submitted to allo-SCT. Bone Marrow Transplant. 2012, 47, 1206–1211. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Ansari, M.A.; Kumar, B.; Dutta, D.; Roy, A.; Chikoti, L.; Pisano, G.; Dutta, S.; Vahedi, S.; Veettil, M.V.; et al. Histone H2B-IFI16 Recognition of Nuclear Herpesviral Genome Induces Cytoplasmic Interferon-β Responses. PLOS Pathog. 2016, 12, e1005967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takegoshi, K.; Honda, M.; Okada, H.; Takabatake, R.; Nagata, N.; Campbell, J.S.; Nishikawa, M.; Shimakami, T.; Shirasaki, T.; Sakai, Y.; et al. Branched-chain amino acids prevent hepatic fibrosis and development of hepatocellular carcinoma in a non-alcoholic steatohepatitis mouse model. Oncotarget 2017, 8, 18191–18205. [Google Scholar] [CrossRef]
- Jin, E.; Han, S.; Son, M.; Kim, S.W. Cordyceps bassiana inhibits smooth muscle cell proliferation via the ERK1/2 MAPK signaling pathway. Cell. Mol. Biol. Lett. 2016, 21, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, L.; Jin, J.; Luan, Y.; Chen, C.; Li, Y.; Chu, H.; Wang, X.; Liao, G.; Yu, Y.; et al. p38 inhibition provides anti–DNA virus immunity by regulation of USP21 phosphorylation and STING activation. J. Exp. Med. 2017, 214, 991–1010. [Google Scholar] [CrossRef]
- Liang, D.; Xiao-Feng, H.; Guan-Jun, D.; Er-Ling, H.; Sheng, C.; Ting-Ting, W.; Qin-Gang, H.; Yan-Hong, N.; Ya-Yi, H. Activated STING enhances Tregs infiltration in the HPV-related carcinogenesis of tongue squamous cells via the c-jun/CCL22 signal. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2015, 1852, 2494–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ande, S.R.; Nguyen, K.H.; Nyomba, B.L.G.; Mishra, S. Prohibitin-induced, obesity-associated insulin resistance and accompanying low-grade inflammation causes NASH and HCC. Sci. Rep. 2016, 6, 23608. [Google Scholar] [CrossRef] [PubMed]
- Afrin, M.R.; Arumugam, S.; Pitchaimani, V.; Karuppagounder, V.; Thandavarayan, R.A.; Harima, M.; Hossain, C.F.; Suzuki, K.; Sone, H.; Matsubayashi, Y.; et al. Le Carbone prevents liver damage in non-alcoholic steatohepatitis-hepatocellular carcinoma mouse model via AMPKα-SIRT1 signaling pathway activation. Heliyon 2021, 7, e05888. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.G.; Symons, J.A.; McDowell, T.L.; McDevitt, H.O.; Duff, G.W. Effects of a polymor-phism in the human tumor necrosis factor α promoter on transcriptional activation. Natl. Acad. Sci. 1997, 94, 3195–3199. [Google Scholar] [CrossRef] [Green Version]
- Vassalli, P. The pathophysiology of tumor necrosis factors. Annu. Rev. Immunol. 1992, 10, 411–452. [Google Scholar] [CrossRef]
- Kim, J.Y.; Garcia-Carbonell, R.; Yamachika, S.; Zhao, P.; Dhar, D.; Loomba, R.; Kaufman, R.J.; Saltiel, A.R.; Karin, M. ER Stress Drives Lipogenesis and Steatohepatitis via Caspase-2 Activation of S1P. Cell 2018, 175, 133–145.e15. [Google Scholar] [CrossRef] [Green Version]
- Todoric, J.; Di Caro, G.; Reibe, S.; Henstridge, D.C.; Green, C.R.; Vrbanac, A.; Ceteci, F.; Conche, C.; McNulty, R.; Shalapour, S.; et al. Fructose stimulated de novo lipogenesis is promoted by inflammation. Nat. Metab. 2020, 2, 1034–1045. [Google Scholar] [CrossRef]
- Nakagawa, H.; Umemura, A.; Taniguchi, K.; Font-Burgada, J.; Dhar, D.; Ogata, H.; Zhong, Z.; Valasek, M.A.; Seki, E.; Hidalgo, J.; et al. ER Stress Cooperates with Hypernutrition to Trigger TNF-Dependent Spontaneous HCC Development. Cancer Cell 2014, 26, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, S.; Panera, N.; Gnani, D.; Nobili, V. Dual Role of MicroRNAs in NAFLD. Int. J. Mol. Sci. 2013, 14, 8437–8455. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Durán, R.; Romero-Gómez, M. Epigenetic mechanisms in non-alcoholic fatty liver disease: An emerging field. World J. Hepatol. 2015, 7, 2497. [Google Scholar] [CrossRef]
- Chang, J.; Nicolas, E.; Marks, D.; Sander, C.; Lerro, A.; Buendia, M.A.; Xu, C.; Mason, W.S.; Moloshok, T.; Bort, R.; et al. miR-122, a Mammalian Liver-Specific microRNA, is Processed from hcr mRNA and MayDownregulate the High Affinity Cationic Amino Acid Transporter CAT-1. RNA Biol. 2004, 1, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, O.; Puri, P.; Eicken, C.; Contos, M.J.; Mirshahi, F.; Maher, J.W.; Kellum, J.M.; Min, H.; Luketic, V.A.; Sanyal, A.J. Nonalcoholic steatohepatitis is associated with altered hepatic MicroRNA expression. Hepatology 2008, 48, 1810–1820. [Google Scholar] [CrossRef] [Green Version]
- Vaschetto, L.M. miRNA activation is an endogenous gene expression pathway. RNA Biol. 2018, 15, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Ramchandran, R.; Chaluvally-Raghavan, P. miRNA-Mediated RNA Activation in Mammalian Cells. Adv. Exp. Med. Biol. 2017, 983, 81–89. [Google Scholar] [CrossRef]
- Huang, R.; Duan, X.; Fan, J.; Li, G.; Wang, B. Role of Noncoding RNA in Development of Nonalcoholic Fatty Liver Disease. BioMed Res. Int. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Lu, Z.; Cai, J.; Huang, K.; Chen, B.; Li, G.; Dong, P.; Zheng, J. MALAT1 functions as a competing endogenous RNA to mediate Rac1 expression by sequestering miR-101b in liver fibrosis. Cell Cycle 2015, 14, 3885–3896. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Chen, J.; Chen, N. Long noncoding RNA MALAT1 promotes hepatic steatosis and insulin re-sistance by increasing nuclear SREBP-1c protein stability. Sci. Rep. 2016, 6, 1–11. [Google Scholar]
- Zheng, J.; Yuan, Z.; Dong, P.; Wu, L.; Zhang, Y.; Hu, Y.; Zheng, L. Long non-coding RNA PVT1 activates hepatic stellate cells through competitively binding microRNA-152. Oncotarget 2016, 7, 62886–62897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Arendt, B.M.; Comelli, E.M.; Ma, D.W.; Lou, W.; Teterina, A.; Kim, T.; Fung, S.K.; Wong, D.K.; McGilvray, I.; Fischer, S.E.; et al. Altered hepatic gene expression in nonalcoholic fatty liver disease is associated with lower hepatic n-3 and n-6 polyunsaturated fatty acids. Hepatology 2015, 61, 1565–1578. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado, A.L.F.D.A.; De Carvalho, L.; Oliveira, A.C.; dos Santos, V.N.; Vieira, J.; Parise, E.R. Insulin resistance index (HOMA-IR) in the differentiation of patients with non-alcoholic fatty liver disease and healthy individuals. Arq. De Gastroenterol. 2010, 47, 165–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Group A NAFLD/NASH without Suspicion of Advanced Fibrosis (F1–F2, TE < 8 kPa), n = 60 | Group B NAFLD/NASH with Suspicion of Advanced Fibrosis (F3–F4, TE ≥ 8 kPa), n = 40 | Group 3 Control N = 100 | p Value |
|---|---|---|---|---|
| Sex | ||||
| male | 39 (65%) | 30 (75%) | 64 (64%) | 0.441 |
| female | 21 (35%) | 10 (25%) | 36 (36%) | |
| History of diabetes mellitus | 0.00 ** | |||
| positive | 49 (81.7%) | 34 (85%) | 42 (42%) | |
| negative | 11 (18.3%) | 6 (15%) | 58 (58%) | |
| Body mass index (kg/m2) BMI | 35.5 ± 5.1 | 33.7 ± 6.7 | 25.9 ± 3.3 | a 0.00 ** b 0.00 ** c 0.134 |
| Total cholesterol (mg/dL) | 298.18 ± 59.6 | 289.4 ± 60.1 | 189 ± 85.9 | a 0.00 ** b 0.00 ** c 0.545 |
| LDLc (mg/dL) | 209.9 ± 49.5 | 199 ± 60.4 | 136.18 ± 66.3 | a 0.001 ** b 0.00 ** c 0.386 |
| HDLc (mg/dL) | 30.8.5 ± 9.09 | 27.6 ± 6.5 | 50.43 ± 20.8 | a 0.00 ** b 0.00 ** c 0.328 |
| Total triglycerides (mg/dL) | 270.3 ± 77.6 | 298.15 ± 58.4 | 179.7 ± 90.7 | a 0.00 ** b 0.00 ** c 0.106 |
| albumin creatinine ratio | 25.07 ± 4.2 | 23.5 ± 5.01 | 20.2 ± 6.9 | a 0.00 ** b 0.00 ** c 0.207 |
| AST (IU/L) | 71.2 ± 36.9 | 70.6 ± 41.2 | 51 ± 19.7 | a 0.00 ** b 0.002 ** c 0.993 |
| ALT (IU/L) | 46.3 ± 25.2 | 59.7 ± 44.8 | 34.3 ± 16.4 | a 0.00 ** b 0.007 ** c 0.012 * |
| Total bilirubin(mg/dL) | 2.6 ± 0.9 | 3 ± 0.8 | 1.5 ± 1.2 | a 0.00 ** b 0.00 ** c 0.057 |
| Direct bilirubin (mg/dL) | 1.5 ± 0.66 | 1.7 ± 0.69 | 0.88 ± 0.39 | a 0.00 ** b 0.00 ** c 0.022 * |
| Albumin(g/dL) | 2.5 ± 0.5 | 2.4 ± 039 | 3.23 ± 0.3 | a 0.00 ** b 0.00 ** c 0.065 |
| Gamma glutammyl transferase (IU/L) | 57.8 ± 39.9 | 65.6 ± 31.3 | 22.3 ± 21.7 | a 0.00 ** b 0.00 ** c 0.243 |
| Alpha fetoprotein | 180.5.9 ± 439 | 359 ± 433 | 18.0 ± 31.27 | a 0.004 ** b 0.00 ** c 0.012 * |
| Fasting blood glucose(mg/dL) | 207.5 ± 83.3 | 179.3 ± 83.5 | 151.0 ± 87 | a 0.000 * b 0.106 c 0.194 |
| Glycated hemoglobin HbA1c (%) | 7.07 ± 1.09 | 7.8 ± 2.01 | 6.5 ± 2.7 | a 0.27 b 0.008 ** c 0.000 * |
| HOMA IR | 12.66 ± 7.9 | 19.3 ± 6.8 | 5.0 ± 6.1 | a 0.000 ** b 0.00 ** c 0.000 * |
| NAFLD Score | --- | -- | ||
| NAFLD Score < −1.455 = F0–F2 | 27 (45%) | 0 (0%) | ||
| NAFLD Score −1.455 − 0.675 | 33 (65%) | 2 (5%) | ||
| NAFLD Score > 0.675 = F3–F4 | 0 (0%) | 38 (95%) | ||
| Fibrosis score | --- | --- | ||
| F0 to F1 Mild liver scaring | 34 (56.7%) | 0 (0%) | ||
| F2: Moderate liver scarring | 26 (46.3%) | 0 (0%) | ||
| F3: Severe liver scarring | 0 (0%) | 29 (72.5%) | ||
| F4: Advanced liver scarring (cirrhosis) | 0 (0%) | 11 (27.5%) | ||
| steatosis grading | ||||
| S1 mild steatosis | 15 (25%) | 0 (0%) | ||
| S2 moderate steatosis | 21 (35%) | 4 (10%) | ||
| S3 severe steatosis | 5 (8.3%) | 36 (90%) | ||
| S4 non steatosis | 19 (31.7%) | 0 (0%) | ||
| Test Result Variable(s) | Area | Std. Error | Asymptotic Sig. | Asymptotic 95% Confidence Interval | Cutoff | Sensitivity | Specificity | |
|---|---|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | |||||||
| NAFLD/NASH vs. Control | ||||||||
| lncRNA RABGAP1L-DT-206 | 0.844 | 0.031 | 0.000 | 0.82 | 0.905 | 4.8 | 81% | 83% |
| has-miR-mir-6888-5p | 0.916 | 0.019 | 0.000 | 0.879 | 0.953 | 1.97 | 91% | 77% |
| EDN1mRNA | 0.797 | 0.033 | 0.000 | 0.731 | 0.862 | 1.85 | 87% | 70% |
| EP300 mRNA | 0.839 | 0.031 | 0.000 | 0.779 | 0.900 | 2.15 | 83% | 80% |
| MAPK3 mRNA | 0.871 | 0.026 | 0.000 | 0.820 | 0.921 | 2.65 | 88% | 73% |
| TNF mRNA | 0.841 | 0.031 | 0.000 | 0.781 | 0.901 | 2.05 | 82% | 81% |
| Combined RNAs | 0.888 | 0.022 | 0.000 | 0.923 | 0.845 | 3.25 | 91% | 73% |
| AST | 0.653 | 0.039 | 0.000 | 57 | 0.577 | 0.729 | 55% | 72% |
| ALT | 0.669 | 0.039 | 0.000 | 27 | 0.593 | 0.745 | 57% | 73% |
| GGT | 0.806 | 0.030 | 0.000 | 39.5 | 0.748 | 0.864 | 66% | 71% |
| Group A vs. Group B | ||||||||
| lncRNA RABGAP1L-DT-206 | 0.944 | 0.038 | 0.000 | 3.4 | 0.869 | 1 | 100% | 79% |
| hsa-mir-6888-5p | 0.628 | 0.097 | 0.197 | 3.6 | 0.438 | 0.819 | 50.7% | 69.6% |
| EDN1mRNA | 0.648 | 0.097 | 0.136 | 4.1 | 0.457 | 0.839 | 62.5% | 64.5% |
| EP300 mRNA | 0.707 | 0.089 | 0.037 | 2.3 | 0.533 | 0.832 | 81.3% | 58.9% |
| MAPK3 mRNA | 0.729 | 0.088 | 0.021 | 4.05 | 0.556 | 0.901 | 68.8% | 58.9% |
| TNF mRNA | 0.727 | 0.089 | 0.022 | 2.3 | 0.561 | 0.893 | 75.3% | 58.9% |
| B | S.E. | Sig. | Exp(B) | 95% C.I. for EXP(B) | ||
|---|---|---|---|---|---|---|
| Lower | Upper | |||||
| Age | 0.007 | 0.029 | 0.803 | 1.007 | 0.952 | 1.066 |
| lncRNA RABGAP1L-DT-206 | −0.08 | 0.010 | 0.05 | 0.982 | 0.963 | 1.002 |
| has-miR-mir-6888-5p | −0.021 | 0.015 | 0.174 | 0.979 | 0.951 | 1.009 |
| EDN1mRNA | 0.009 | 0.008 | 0.255 | 1.010 | 0.993 | 1.026 |
| EP300 mRNA | −0.024 | 0.018 | 0.192 | 0.976 | 0.942 | 1.012 |
| MAPK3 mRNA | −0.042 | 0.020 | 0.034 | 0.959 | 0.922 | 0.997 |
| TNF mRNA | −0.045 | 0.020 | 0.025 | 0.956 | 0.918 | 0.994 |
| ALT | −0.025 | 0.010 | 0.011 | 0.976 | 0.957 | 0.994 |
| Constant | 2.108 | 1.615 | 0.192 | 8.233 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albadawy, R.; Agwa, S.H.A.; Khairy, E.; Saad, M.; El Touchy, N.; Othman, M.; El Kassas, M.; Matboli, M. Circulatory Endothelin 1-Regulating RNAs Panel: Promising Biomarkers for Non-Invasive NAFLD/NASH Diagnosis and Stratification: Clinical and Molecular Pilot Study. Genes 2021, 12, 1813. https://doi.org/10.3390/genes12111813
Albadawy R, Agwa SHA, Khairy E, Saad M, El Touchy N, Othman M, El Kassas M, Matboli M. Circulatory Endothelin 1-Regulating RNAs Panel: Promising Biomarkers for Non-Invasive NAFLD/NASH Diagnosis and Stratification: Clinical and Molecular Pilot Study. Genes. 2021; 12(11):1813. https://doi.org/10.3390/genes12111813
Chicago/Turabian StyleAlbadawy, Reda, Sara H. A. Agwa, Eman Khairy, Maha Saad, Naglaa El Touchy, Mohamed Othman, Mohamed El Kassas, and Marwa Matboli. 2021. "Circulatory Endothelin 1-Regulating RNAs Panel: Promising Biomarkers for Non-Invasive NAFLD/NASH Diagnosis and Stratification: Clinical and Molecular Pilot Study" Genes 12, no. 11: 1813. https://doi.org/10.3390/genes12111813