
A Common CDH13 Variant Is Associated with Low Agreeableness and Neural Responses to Working Memory Tasks in ADHD

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Case-Control Cohort
2.2. Genotyping
2.3. Measurement of Personality Traits
2.4. Functional Electrophysiology
2.5. Statistical Analysis
3. Results
3.1. Case-Control Association Study
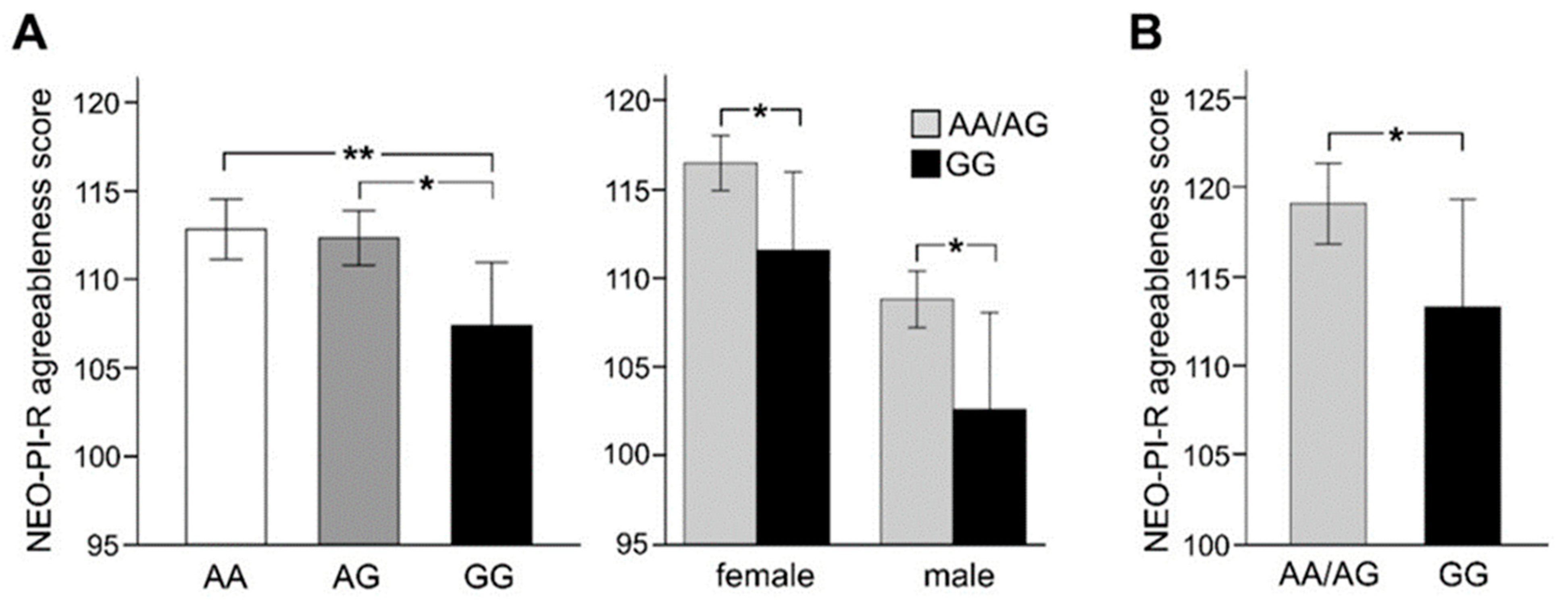
3.2. Association of rs2199430 with Personality Traits
3.3. Functional Electrophysiology
3.3.1. N-Back (Working Memory)—Behavioral Data
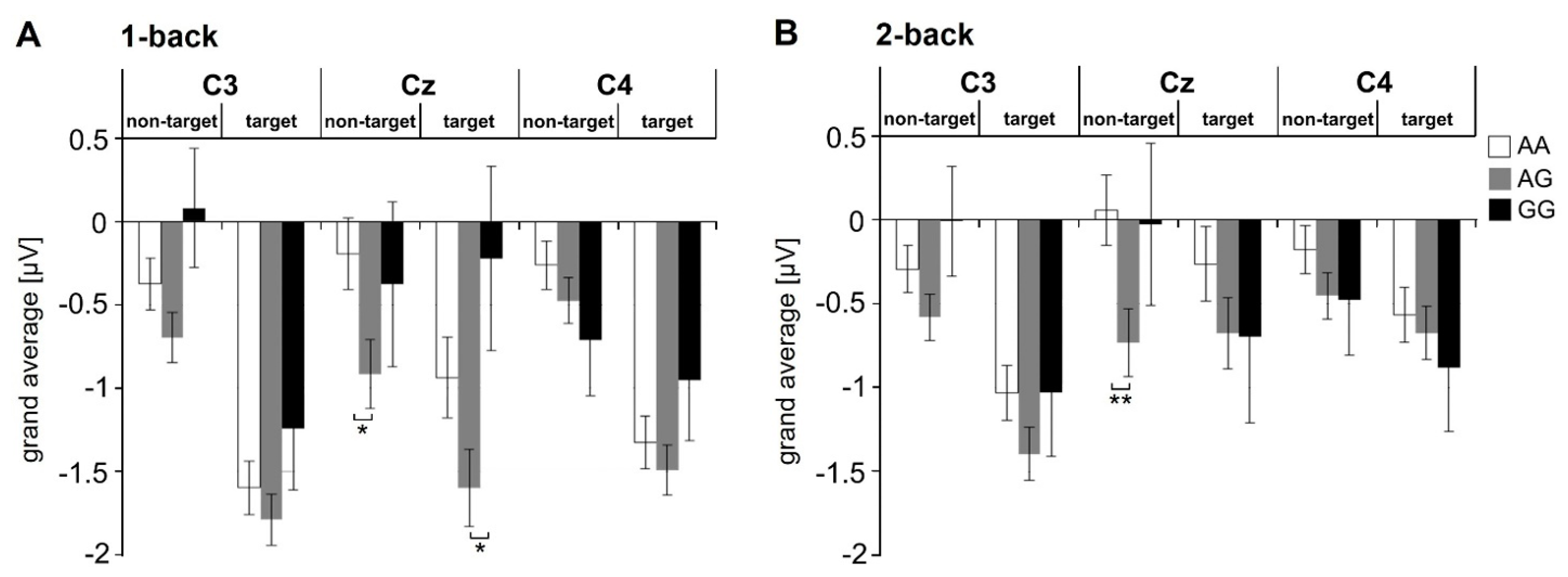
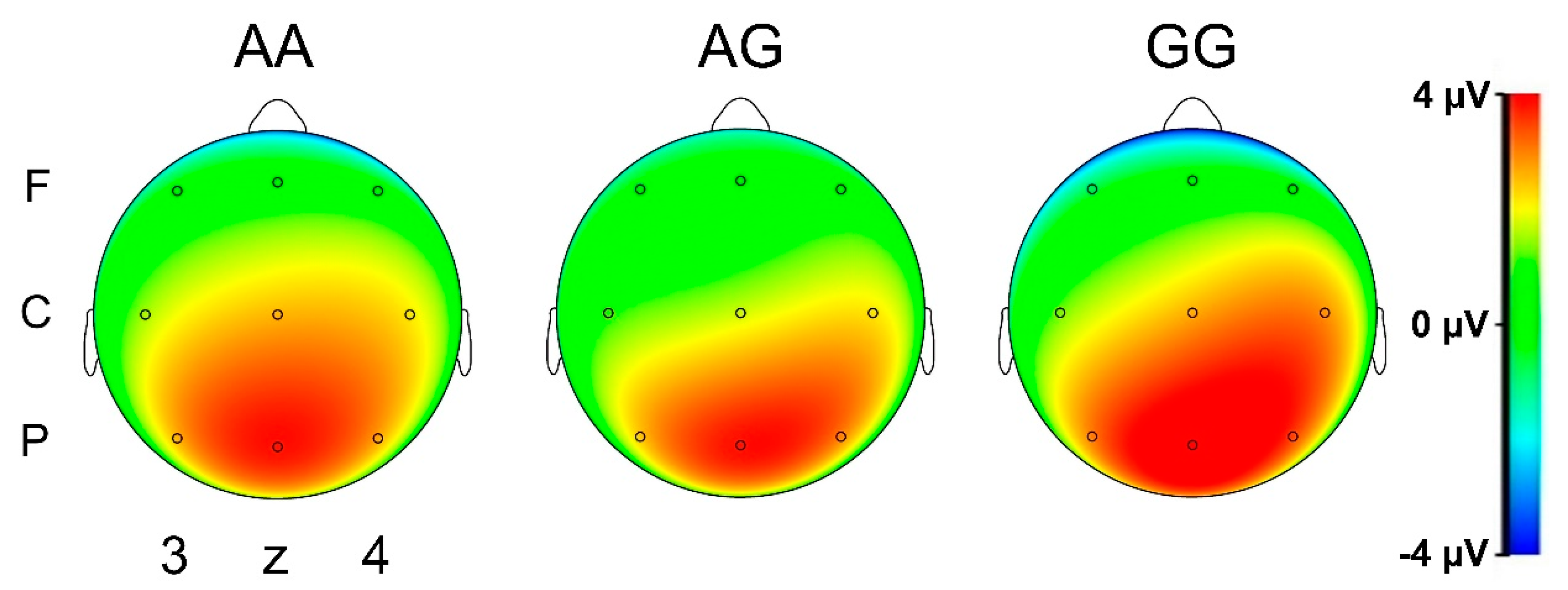
3.3.2. N-Back (Working Memory)—ERP Data
N200 Amplitude
P300 Amplitude
3.3.3. CPT—Behavioral Data
3.3.4. CPT—Neurophysiological/ERP Data
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lasky-Su, J.; Neale, B.M.; Franke, B.; Anney, R.J.; Zhou, K.; Maller, J.B.; Vasquez, A.A.; Chen, W.; Asherson, P.; Buitelaar, J.; et al. Genome-wide association scan of quantitative traits for attention deficit hyperactivity disorder identifies novel associations and confirms candidate gene associations. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 147, 1345–1354. [Google Scholar] [CrossRef]
- Lesch, K.P.; Timmesfeld, N.; Renner, T.J.; Halperin, R.; Roser, C.; Nguyen, T.T.; Craig, D.W.; Romanos, J.; Heine, M.; Meyer, J.; et al. Molecular genetics of adult ADHD: Converging evidence from genome-wide association and extended pedigree linkage studies. J. Neural. Transm. 2008, 115, 1573–1585. [Google Scholar] [CrossRef]
- Neale, B.M.; Medland, S.; Ripke, S.; Anney, R.J.; Asherson, P.; Buitelaar, J.; Franke, B.; Gill, M.; Kent, L.; Holmans, P.; et al. Case-control genome-wide association study of attention-deficit/hyperactivity disorder. J. Am. Acad. Child. Adolesc. Psychiatry 2010, 49, 906–920. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Dempfle, A.; Arcos-Burgos, M.; Bakker, S.C.; Banaschewski, T.; Biederman, J.; Buitelaar, J.; Castellanos, F.X.; Doyle, A.; Ebstein, R.P.; et al. Meta-analysis of genome-wide linkage scans of attention deficit hyperactivity disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 147, 1392–1398. [Google Scholar] [CrossRef] [Green Version]
- Kushima, I.; Aleksic, B.; Nakatochi, M.; Shimamura, T.; Okada, T.; Uno, Y.; Morikawa, M.; Ishizuka, K.; Shiino, T.; Kimura, H.; et al. Comparative Analyses of Copy-Number Variation in Autism Spectrum Disorder and Schizophrenia Reveal Etiological Overlap and Biological Insights. Cell Rep. 2018, 24, 2838–2856. [Google Scholar] [CrossRef] [Green Version]
- Sanders, S.J.; He, X.; Willsey, A.J.; Ercan-Sencicek, A.G.; Samocha, K.E.; Cicek, A.E.; Murtha, M.T.; Bal, V.H.; Bishop, S.L.; Dong, S.; et al. Insights into Autism Spectrum Disorder Genomic Architecture and Biology from 71 Risk Loci. Neuron 2015, 87, 1215–1233. [Google Scholar] [CrossRef] [Green Version]
- Edwards, A.C.; Aliev, F.; Bierut, L.J.; Bucholz, K.K.; Edenberg, H.; Hesselbrock, V.; Kramer, J.; Kuperman, S.; Nurnberger, J.I., Jr.; Schuckit, M.A.; et al. Genome-wide association study of comorbid depressive syndrome and alcohol dependence. Psychiatr. Genet. 2012, 22, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Howard, D.M.; Adams, M.J.; Clarke, T.K.; Hafferty, J.D.; Gibson, J.; Shirali, M.; Coleman, J.R.I.; Hagenaars, S.P.; Ward, J.; Wigmore, E.M.; et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat. Neurosci. 2019, 22, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Cohen-Woods, S.; Chen, Q.; Noor, A.; Knight, J.; Hosang, G.; Parikh, S.V.; De Luca, V.; Tozzi, F.; Muglia, P.; et al. Genome-wide association study of bipolar disorder in Canadian and UK populations corroborates disease loci including SYNE1 and CSMD1. BMC Med. Genet. 2014, 15, 2. [Google Scholar] [CrossRef] [Green Version]
- Uhl, G.R.; Drgon, T.; Liu, Q.R.; Johnson, C.; Walther, D.; Komiyama, T.; Harano, M.; Sekine, Y.; Inada, T.; Ozaki, N.; et al. Genome-wide association for methamphetamine dependence: Convergent results from 2 samples. Arch. Gen. Psychiatry 2008, 65, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Treutlein, J.; Cichon, S.; Ridinger, M.; Wodarz, N.; Soyka, M.; Zill, P.; Maier, W.; Moessner, R.; Gaebel, W.; Dahmen, N.; et al. Genome-wide association study of alcohol dependence. Arch. Gen. Psychiatry 2009, 66, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.B.; Engelhardt, B.E.; Wardle, M.C.; Sokoloff, G.; Stephens, M.; de Wit, H.; Palmer, A.A. Genome-wide association study of d-amphetamine response in healthy volunteers identifies putative associations, including cadherin 13 (CDH13). PLoS ONE 2012, 7, e42646. [Google Scholar] [CrossRef]
- Drgonova, J.; Walther, D.; Hartstein, G.L.; Bukhari, M.O.; Baumann, M.H.; Katz, J.; Hall, F.S.; Arnold, E.R.; Flax, S.; Riley, A.; et al. Cadherin 13: Human cis-regulation and selectively-altered addiction phenotypes and cerebral cortical dopamine in knockout mice. Mol. Med. 2016, 22, 537–547. [Google Scholar] [CrossRef]
- Kohannim, O.; Hibar, D.P.; Jahanshad, N.; Stein, J.L.; Hua, X.; Toga, A.W.; Jack, C.R., Jr.; Weiner, M.W.; Thompson, P.M. The Alzheimer’s Disease Neuroimaging Initiative. Predicting Temporal Lobe Volume on Mri from Genotypes Using L(1)-L(2) Regularized Regression. Proc. IEEE Int. Symp. Biomed. Imaging 2012, 1160–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otowa, T.; Yoshida, E.; Sugaya, N.; Yasuda, S.; Nishimura, Y.; Inoue, K.; Tochigi, M.; Umekage, T.; Miyagawa, T.; Nishida, N.; et al. Genome-wide association study of panic disorder in the Japanese population. J. Hum. Genet. 2009, 54, 122–126. [Google Scholar] [CrossRef]
- Rivero, O.; Sich, S.; Popp, S.; Schmitt, A.; Franke, B.; Lesch, K.P. Impact of the ADHD-susceptibility gene CDH13 on development and function of brain networks. Eur. Neuropsychopharmacol. 2013, 23, 492–507. [Google Scholar] [CrossRef]
- Fredette, B.J.; Miller, J.; Ranscht, B. Inhibition of motor axon growth by T-cadherin substrata. Development 1996, 122, 3163–3171. [Google Scholar] [CrossRef]
- Forero, A.; Rivero, O.; Waldchen, S.; Ku, H.P.; Kiser, D.P.; Gartner, Y.; Pennington, L.S.; Waider, J.; Gaspar, P.; Jansch, C.; et al. Cadherin-13 Deficiency Increases Dorsal Raphe 5-HT Neuron Density and Prefrontal Cortex Innervation in the Mouse Brain. Front. Cell Neurosci. 2017, 11, 307. [Google Scholar] [CrossRef] [Green Version]
- Rivero, O.; Selten, M.M.; Sich, S.; Popp, S.; Bacmeister, L.; Amendola, E.; Negwer, M.; Schubert, D.; Proft, F.; Kiser, D.; et al. Cadherin-13, a risk gene for ADHD and comorbid disorders, impacts GABAergic function in hippocampus and cognition. Transl. Psychiatry 2015, 5, e655. [Google Scholar] [CrossRef] [Green Version]
- Kiser, D.P.; Popp, S.; Schmitt-Bohrer, A.G.; Strekalova, T.; van den Hove, D.L.; Lesch, K.P.; Rivero, O. Early-life stress impairs developmental programming in Cadherin 13 (CDH13)-deficient mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 158–168. [Google Scholar] [CrossRef]
- Takeuchi, T.; Misaki, A.; Liang, S.B.; Tachibana, A.; Hayashi, N.; Sonobe, H.; Ohtsuki, Y. Expression of T-cadherin (CDH13, H-Cadherin) in human brain and its characteristics as a negative growth regulator of epidermal growth factor in neuroblastoma cells. J. Neurochem. 2000, 74, 1489–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiihonen, J.; Rautiainen, M.R.; Ollila, H.M.; Repo-Tiihonen, E.; Virkkunen, M.; Palotie, A.; Pietilainen, O.; Kristiansson, K.; Joukamaa, M.; Lauerma, H.; et al. Genetic background of extreme violent behavior. Mol. Psychiatry 2015, 20, 786–792. [Google Scholar] [CrossRef] [Green Version]
- Terracciano, A.; Sanna, S.; Uda, M.; Deiana, B.; Usala, G.; Busonero, F.; Maschio, A.; Scally, M.; Patriciu, N.; Chen, W.M.; et al. Genome-wide association scan for five major dimensions of personality. Mol. Psychiatry 2010, 15, 647–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anckarsäter, H.; Stahlberg, O.; Larson, T.; Hakansson, C.; Jutblad, S.B.; Niklasson, L.; Nyden, A.; Wentz, E.; Westergren, S.; Cloninger, C.R.; et al. The impact of ADHD and autism spectrum disorders on temperament, character, and personality development. Am. J. Psychiatry 2006, 163, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- August, G.J.; Realmuto, G.M.; MacDonald, A.W., 3rd; Nugent, S.M.; Crosby, R. Prevalence of ADHD and comorbid disorders among elementary school children screened for disruptive behavior. J. Abnorm. Child. Psychol. 1996, 24, 571–595. [Google Scholar] [CrossRef] [PubMed]
- Arias-Vasquez, A.; Altink, M.E.; Rommelse, N.N.; Slaats-Willemse, D.I.; Buschgens, C.J.; Fliers, E.A.; Faraone, S.V.; Sergeant, J.A.; Oosterlaan, J.; Franke, B.; et al. CDH13 is associated with working memory performance in attention deficit/hyperactivity disorder. Genes Brain Behav. 2011, 10, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Franke, B.; Vasquez, A.A.; Johansson, S.; Hoogman, M.; Romanos, J.; Boreatti-Hummer, A.; Heine, M.; Jacob, C.P.; Lesch, K.P.; Casas, M.; et al. Multicenter analysis of the SLC6A3/DAT1 VNTR haplotype in persistent ADHD suggests differential involvement of the gene in childhood and persistent ADHD. Neuropsychopharmacology 2010, 35, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.P.; Romanos, J.; Dempfle, A.; Heine, M.; Windemuth-Kieselbach, C.; Kruse, A.; Reif, A.; Walitza, S.; Romanos, M.; Strobel, A.; et al. Co-morbidity of adult attention-deficit/hyperactivity disorder with focus on personality traits and related disorders in a tertiary referral center. Eur. Arch. Psychiatry Clin. Neurosci. 2007, 257, 309–317. [Google Scholar] [CrossRef]
- Jacob, C.P.; Gross-Lesch, S.; Reichert, S.; Geissler, J.; Jans, T.; Kittel-Schneider, S.; Nguyen, T.T.; Romanos, M.; Reif, A.; Dempfle, A.; et al. Sex- and Subtype-Related Differences of Personality Disorders (Axis II) and Personality Traits in Persistent ADHD. J. Atten. Disord. 2016, 20, 1056–1065. [Google Scholar] [CrossRef]
- Schiele, M.A.; Ziegler, C.; Holitschke, K.; Schartner, C.; Schmidt, B.; Weber, H.; Reif, A.; Romanos, M.; Pauli, P.; Zwanzger, P.; et al. Influence of 5-HTT variation, childhood trauma and self-efficacy on anxiety traits: A gene-environment-coping interaction study. J. Neural Transm. 2016, 123, 895–904. [Google Scholar] [CrossRef]
- Schiele, M.A.; Herzog, K.; Kollert, L.; Schartner, C.; Leehr, E.J.; Bohnlein, J.; Repple, J.; Rosenkranz, K.; Lonsdorf, T.B.; Dannlowski, U.; et al. Extending the vulnerability-stress model of mental disorders: Three-dimensional NPSR1 x environment x coping interaction study in anxiety. Br. J. Psychiatry 2020, 217, 645–650. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic. Acids. Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.T.; McCrae, R.R. Revised NEO Personality Inventory (NEO-PI-R) and NEO Five Factor Inventory: Professional Manual; Psychological Assessment Resources: Odessa, FL, USA, 1992. [Google Scholar]
- Owen, A.M.; McMillan, K.M.; Laird, A.R.; Bullmore, E. N-back working memory paradigm: A meta-analysis of normative functional neuroimaging studies. Hum. Brain Mapp. 2005, 25, 46–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccio, C.A.; Reynolds, C.R.; Lowe, P.; Moore, J.J. The continuous performance test: A window on the neural substrates for attention? Arch. Clin. Neuropsychol. 2002, 17, 235–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezdjian, S.; Baker, L.A.; Lozano, D.I.; Raine, A. Assessing inattention and impulsivity in children during the Go/NoGo task. Br. J. Dev. Psychol. 2009, 27, 365–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missonnier, P.; Hasler, R.; Perroud, N.; Herrmann, F.R.; Millet, P.; Richiardi, J.; Malafosse, A.; Giannakopoulos, P.; Baud, P. EEG anomalies in adult ADHD subjects performing a working memory task. Neuroscience 2013, 241, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Fallgatter, A.J.; Ehlis, A.C.; Rosler, M.; Strik, W.K.; Blocher, D.; Herrmann, M.J. Diminished prefrontal brain function in adults with psychopathology in childhood related to attention deficit hyperactivity disorder. Psychiatry Res. 2005, 138, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Merker, S.; Reif, A.; Ziegler, G.C.; Weber, H.; Mayer, U.; Ehlis, A.C.; Conzelmann, A.; Johansson, S.; Muller-Reible, C.; Nanda, I.; et al. SLC2A3 single-nucleotide polymorphism and duplication influence cognitive processing and population-specific risk for attention-deficit/hyperactivity disorder. J. Child. Psychol. Psychiatry 2017, 58, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, G.C.; Roser, C.; Renner, T.; Hahn, T.; Ehlis, A.C.; Weber, H.; Dempfle, A.; Walitza, S.; Jacob, C.; Romanos, M.; et al. KCNJ6 variants modulate reward-related brain processes and impact executive functions in attention-deficit/hyperactivity disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2020, 183, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Fallgatter, A.J.; Strik, W.K. The NoGo-anteriorization as a neurophysiological standard-index for cognitive response control. Int. J. Psychophysiol. 1999, 32, 233–238. [Google Scholar] [CrossRef]
- Costa, P.T.; Terracciano, A.; McCrae, R.R. Gender differences in personality traits across cultures: Robust and surprising findings. J. Pers. Soc. Psychol. 2001, 81, 322–331. [Google Scholar] [CrossRef]
- Fallgatter, A.J.; Brandeis, D.; Strik, W.K. A robust assessment of the NoGo-anteriorisation of P300 microstates in a cued Continuous Performance Test. Brain Topogr. 1997, 9, 295–302. [Google Scholar] [CrossRef]
- Fallgatter, A.J.; Ehlis, A.C.; Seifert, J.; Strik, W.K.; Scheuerpflug, P.; Zillessen, K.E.; Herrmann, M.J.; Warnke, A. Altered response control and anterior cingulate function in attention-deficit/hyperactivity disorder boys. Clin. Neurophysiol. 2004, 115, 973–981. [Google Scholar] [CrossRef]
- Gottesman, I.I.; Gould, T.D. The endophenotype concept in psychiatry: Etymology and strategic intentions. Am. J. Psychiatry 2003, 160, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Lencz, T.; Szeszko, P.R.; DeRosse, P.; Burdick, K.E.; Bromet, E.J.; Bilder, R.M.; Malhotra, A.K. A schizophrenia risk gene, ZNF804A, influences neuroanatomical and neurocognitive phenotypes. Neuropsychopharmacology 2010, 35, 2284–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostendorf, F.; Angleitner, A. NEO-Persönlichkeitsinventar Nach Costa Und McCrae [NEO Personality Inventory According to Costa & McCrae]: NEO-PI-R. In Manual Revidierte Fassung; Hogrefe: Göttingen, Germany, 2004. [Google Scholar]
- Nigg, J.T.; John, O.P.; Blaskey, L.G.; Huang-Pollock, C.L.; Willcutt, E.G.; Hinshaw, S.P.; Pennington, B. Big five dimensions and ADHD symptoms: Links between personality traits and clinical symptoms. J. Personal. Soc. Psychol. 2002, 83, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Gleason, K.A.; Jensen-Campbell, L.A.; Richardson, D.S. Agreeableness as a predictor of aggression in adolescence. Aggress. Behav. 2004, 30, 43–61. [Google Scholar] [CrossRef]
- Laursen, B.; Pulkkinen, L.; Adams, R. The antecedents and correlates of agreeableness in adulthood. Dev. Psychol. 2002, 38, 591–603. [Google Scholar] [CrossRef]
- Pailing, A.; Boon, J.; Egan, V. Personality, the Dark Triad and violence. Personal. Individ. Differ. 2014, 67, 81–86. [Google Scholar] [CrossRef]
- Seo, D.; Patrick, C.J.; Kennealy, P.J. Role of Serotonin and Dopamine System Interactions in the Neurobiology of Impulsive Aggression and its Comorbidity with other Clinical Disorders. Aggress. Violent. Behav. 2008, 13, 383–395. [Google Scholar] [CrossRef] [Green Version]
- Duke, A.A.; Begue, L.; Bell, R.; Eisenlohr-Moul, T. Revisiting the serotonin-aggression relation in humans: A meta-analysis. Psychol. Bull. 2013, 139, 1148–1172. [Google Scholar] [CrossRef] [Green Version]
- Graziano, W.G.; Habashi, M.M.; Sheese, B.E.; Tobin, R.M. Agreeableness, empathy, and helping: A person × situation perspective. J. Personal. Soc. Psychol. 2007, 93, 583–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habashi, M.M.; Graziano, W.G.; Hoover, A.E. Searching for the Prosocial Personality: A Big Five Approach to Linking Personality and Prosocial Behavior. Personal. Soc. Psychol. Bull. 2016, 42, 1177–1192. [Google Scholar] [CrossRef] [Green Version]
- Tantra, M.; Guo, L.; Kim, J.; Zainolabidin, N.; Eulenburg, V.; Augustine, G.J.; Chen, A.I. Conditional deletion of Cadherin 13 perturbs Golgi cells and disrupts social and cognitive behaviors. Genes Brain Behav. 2018, 17, e12466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schriber, R.A.; Robins, R.W.; Solomon, M. Personality and self-insight in individuals with autism spectrum disorder. J. Pers. Soc. Psychol. 2014, 106, 112–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodi-Smith, J.; Rodgers, J.D.; Cunningham, S.A.; Lopata, C.; Thomeer, M.L. Meta-analysis of Big Five personality traits in autism spectrum disorder. Autism 2019, 23, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Antshel, K.M.; Zhang-James, Y.; Faraone, S.V. The comorbidity of ADHD and autism spectrum disorder. Expert Rev. Neurother 2013, 13, 1117–1128. [Google Scholar] [CrossRef]
- Zou, Y.F.; Ye, D.Q.; Feng, X.L.; Su, H.; Pan, F.M.; Liao, F.F. Meta-analysis of BDNF Val66Met polymorphism association with treatment response in patients with major depressive disorder. Eur. Neuropsychopharmacol. 2010, 20, 535–544. [Google Scholar] [CrossRef]
- Verhagen, M.; van der Meij, A.; van Deurzen, P.A.; Janzing, J.G.; Arias-Vasquez, A.; Buitelaar, J.K.; Franke, B. Meta-analysis of the BDNF Val66Met polymorphism in major depressive disorder: Effects of gender and ethnicity. Mol. Psychiatry 2010, 15, 260–271. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Wang, L.; Kuang, W.; Xu, J.; Li, S.; Chen, J.; Yang, Y. Brain-derived neurotrophic factor Val66Met polymorphism association with antidepressant efficacy: A systematic review and meta-analysis. Asia Pac. Psychiatry 2014, 6, 241–251. [Google Scholar] [CrossRef]
- Comings, D.E.; MacMurray, J.P. Molecular heterosis: A review. Mol. Genet. Metab. 2000, 71, 19–31. [Google Scholar] [CrossRef]
- Enriquez-Geppert, S.; Konrad, C.; Pantev, C.; Huster, R.J. Conflict and inhibition differentially affect the N200/P300 complex in a combined go/nogo and stop-signal task. Neuroimage 2010, 51, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Folstein, J.R.; Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology 2008, 45, 152–170. [Google Scholar] [CrossRef] [Green Version]
- McEvoy, L.K.; Smith, M.E.; Gevins, A. Dynamic cortical networks of verbal and spatial working memory: Effects of memory load and task practice. Cereb. Cortex 1998, 8, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.H.; Azzam, P.N. Characterization of N200 and P300: Selected studies of the Event-Related Potential. Int. J. Med. Sci. 2005, 2, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Courtney, S.M.; Petit, L.; Maisog, J.M.; Ungerleider, L.G.; Haxby, J.V. An area specialized for spatial working memory in human frontal cortex. Science 1998, 279, 1347–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubillo, A.; Halari, R.; Smith, A.; Taylor, E.; Rubia, K. A review of fronto-striatal and fronto-cortical brain abnormalities in children and adults with Attention Deficit Hyperactivity Disorder (ADHD) and new evidence for dysfunction in adults with ADHD during motivation and attention. Cortex 2012, 48, 194–215. [Google Scholar] [CrossRef]
- Forero, A.; Ku, H.P.; Malpartida, A.B.; Waldchen, S.; Alhama-Riba, J.; Kulka, C.; Aboagye, B.; Norton, W.H.J.; Young, A.M.J.; Ding, Y.Q.; et al. Serotonin (5-HT) neuron-specific inactivation of Cadherin-13 impacts 5-HT system formation and cognitive function. Neuropharmacology 2020, 168, 108018. [Google Scholar] [CrossRef]
- Vitale, M.R.; Zoller, J.E.M.; Jansch, C.; Janz, A.; Edenhofer, F.; Klopocki, E.; van den Hove, D.; Vanmierlo, T.; Rivero, O.; Nadif Kasri, N.; et al. Generation of induced pluripotent stem cell (iPSC) lines carrying a heterozygous (UKWMPi002-A-1) and null mutant knockout (UKWMPi002-A-2) of Cadherin 13 associated with neurodevelopmental disorders using CRISPR/Cas9. Stem. Cell Res. 2021, 51, 102169. [Google Scholar] [CrossRef]
- Mossink, B.; van Rhijn, J.R.; Wang, S.; Linda, K.; Vitale, M.R.; Zoller, J.E.M.; van Hugte, E.J.H.; Bak, J.; Verboven, A.H.A.; Selten, M.; et al. Cadherin-13 is a critical regulator of GABAergic modulation in human stem-cell-derived neuronal networks. Mol. Psychiatry 2021. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| rs2199430 Genotype | n | NEO-PI-R Agreeableness | t-Statistic | p-Value, Corr. | |
|---|---|---|---|---|---|
| Inattentive Type | AA/AG | 209 | M = 116.7 | t229 = 2.9 | 0.012 |
| SD = 15.5 | |||||
| GG | 22 | M = 106.7 | |||
| SD = 13.2 | |||||
| Combined Type | AA/AG | 516 | M = 111.6 | t571 = 2.1 | 0.107 |
| SD = 16.0 | |||||
| GG | 57 | M = 106.8 | |||
| SD = 18.2 | |||||
| Hyperactive Type | AA/AG | 59 | M = 107.5 | t64 = −1.1 | 0.825 |
| SD = 15.9 | |||||
| GG | 7 | M = 114.4 | |||
| SD = 14.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziegler, G.C.; Ehlis, A.-C.; Weber, H.; Vitale, M.R.; Zöller, J.E.M.; Ku, H.-P.; Schiele, M.A.; Kürbitz, L.I.; Romanos, M.; Pauli, P.; et al. A Common CDH13 Variant Is Associated with Low Agreeableness and Neural Responses to Working Memory Tasks in ADHD. Genes 2021, 12, 1356. https://doi.org/10.3390/genes12091356
Ziegler GC, Ehlis A-C, Weber H, Vitale MR, Zöller JEM, Ku H-P, Schiele MA, Kürbitz LI, Romanos M, Pauli P, et al. A Common CDH13 Variant Is Associated with Low Agreeableness and Neural Responses to Working Memory Tasks in ADHD. Genes. 2021; 12(9):1356. https://doi.org/10.3390/genes12091356
Chicago/Turabian StyleZiegler, Georg C., Ann-Christine Ehlis, Heike Weber, Maria Rosaria Vitale, Johanna E. M. Zöller, Hsing-Ping Ku, Miriam A. Schiele, Laura I. Kürbitz, Marcel Romanos, Paul Pauli, and et al. 2021. "A Common CDH13 Variant Is Associated with Low Agreeableness and Neural Responses to Working Memory Tasks in ADHD" Genes 12, no. 9: 1356. https://doi.org/10.3390/genes12091356