
Mating-Type Locus Organization and Mating-Type Chromosome Differentiation in the Bipolar Edible Button Mushroom Agaricus bisporus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Strains, Genomes and Progenies
2.2. Analyses of Protein Products
2.3. Orthology Reconstruction and Synonymous Divergence
2.4. Detection of Centromeres
2.5. Linkage Map of Mating-Type Locus of the Variety burnettii
3. Results
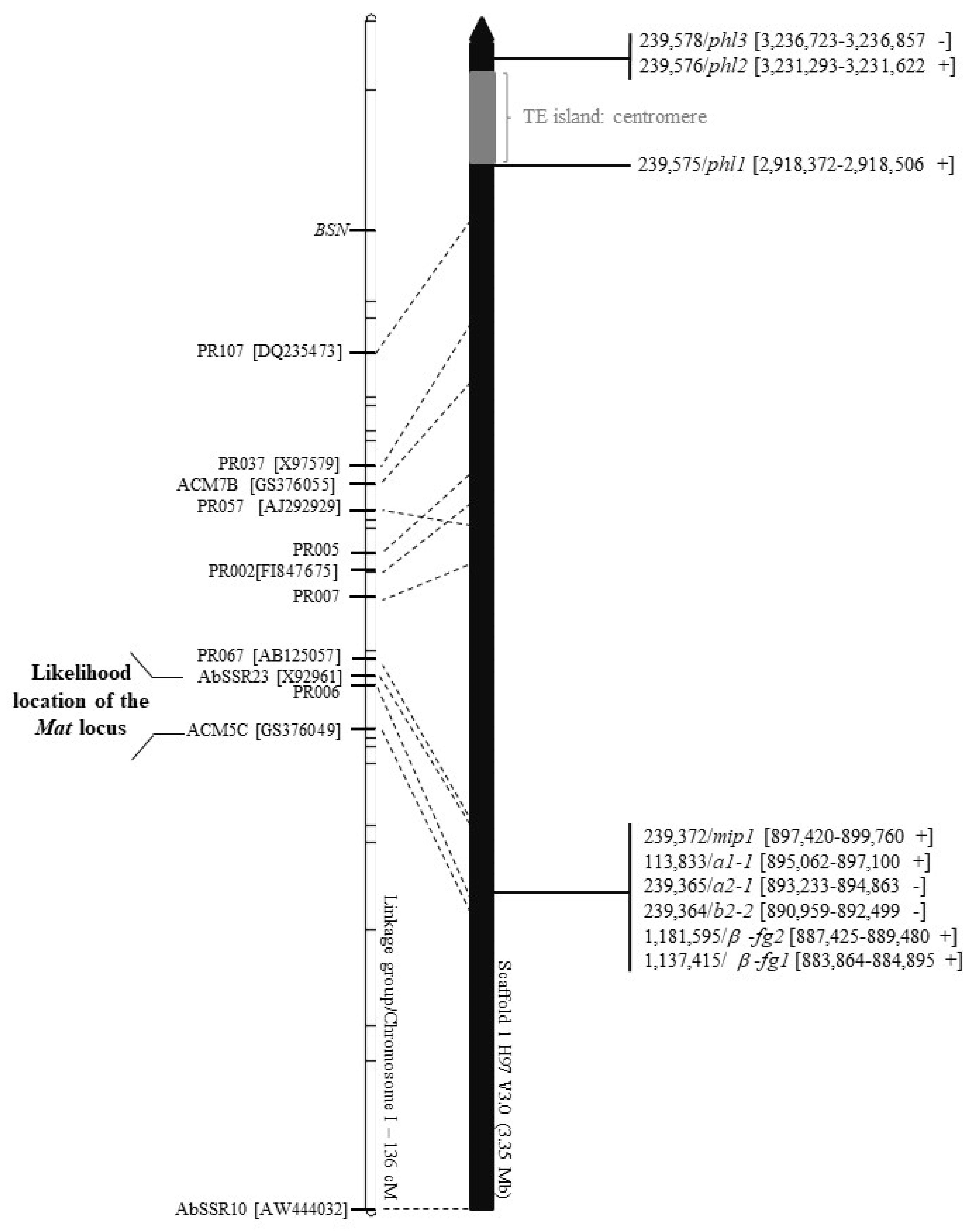
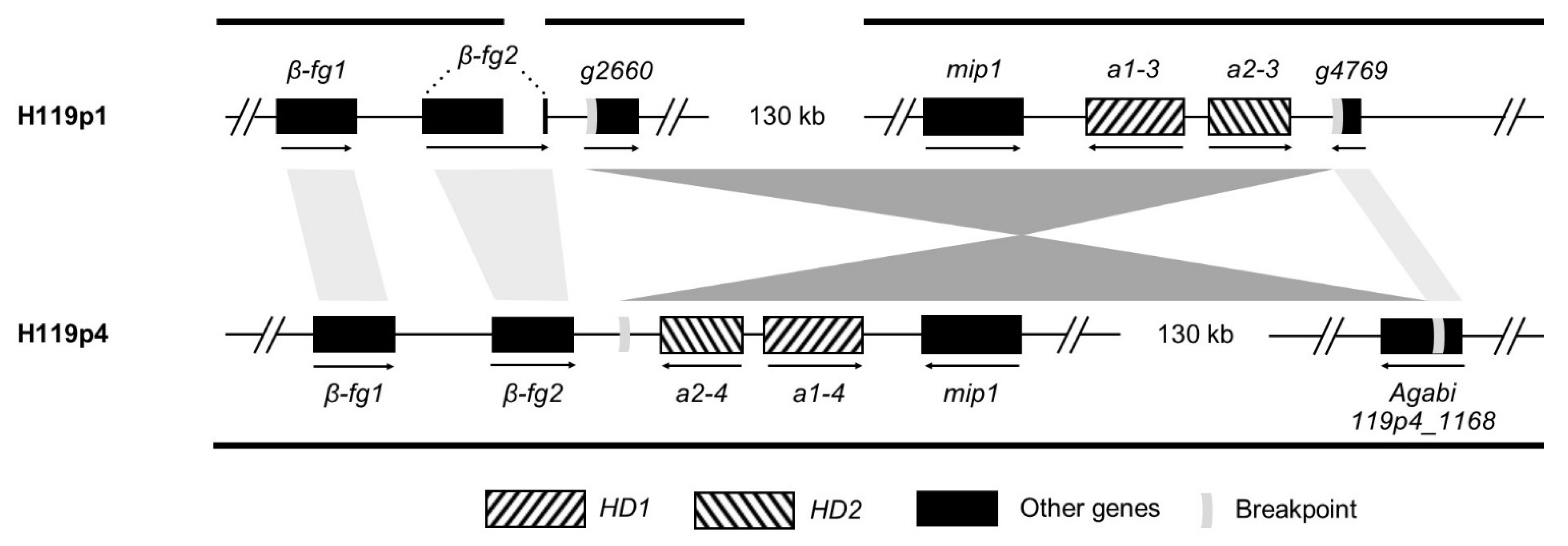
3.1. Defining the Exact Location of the MAT Locus on Chromosome 1
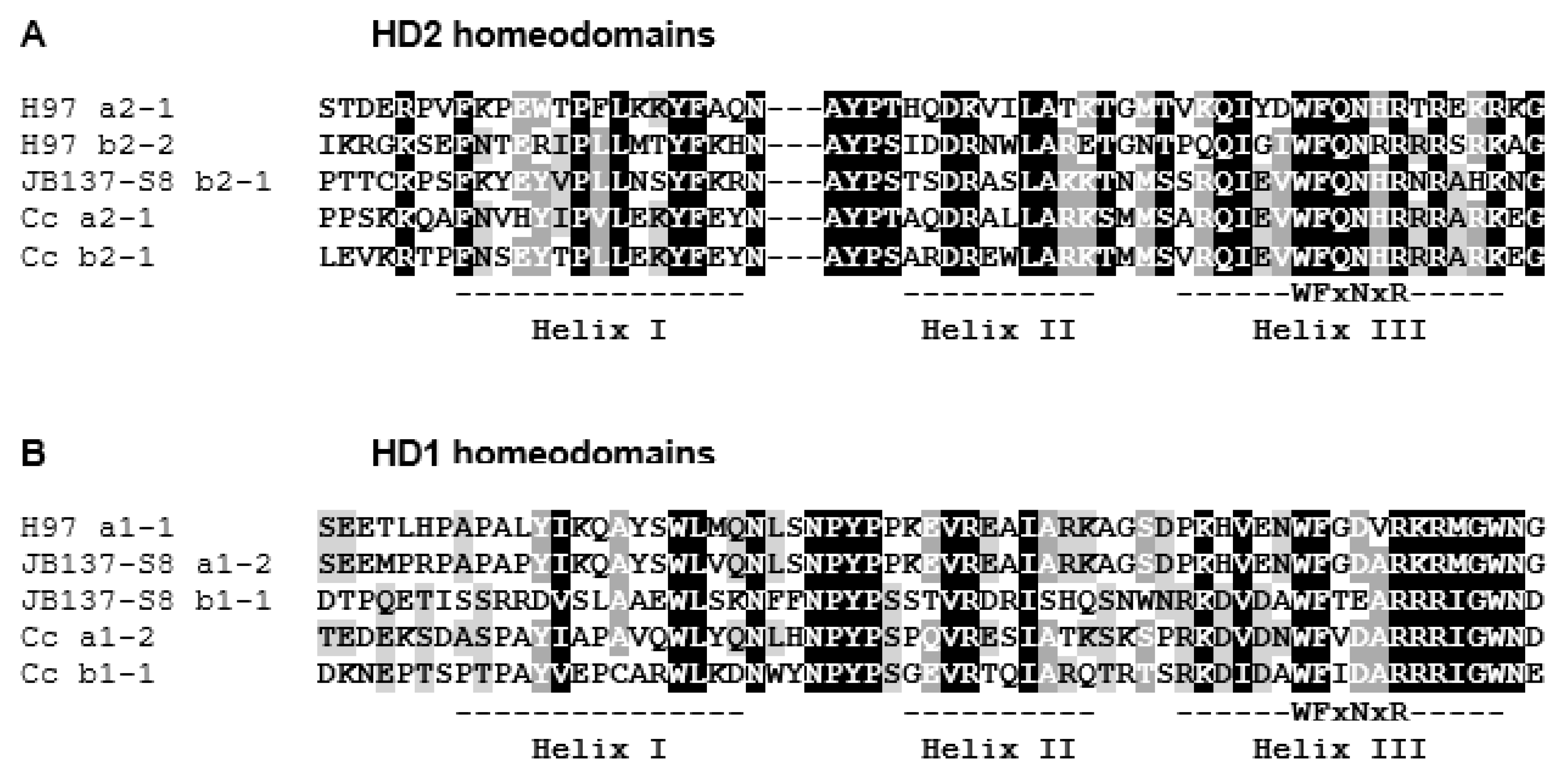
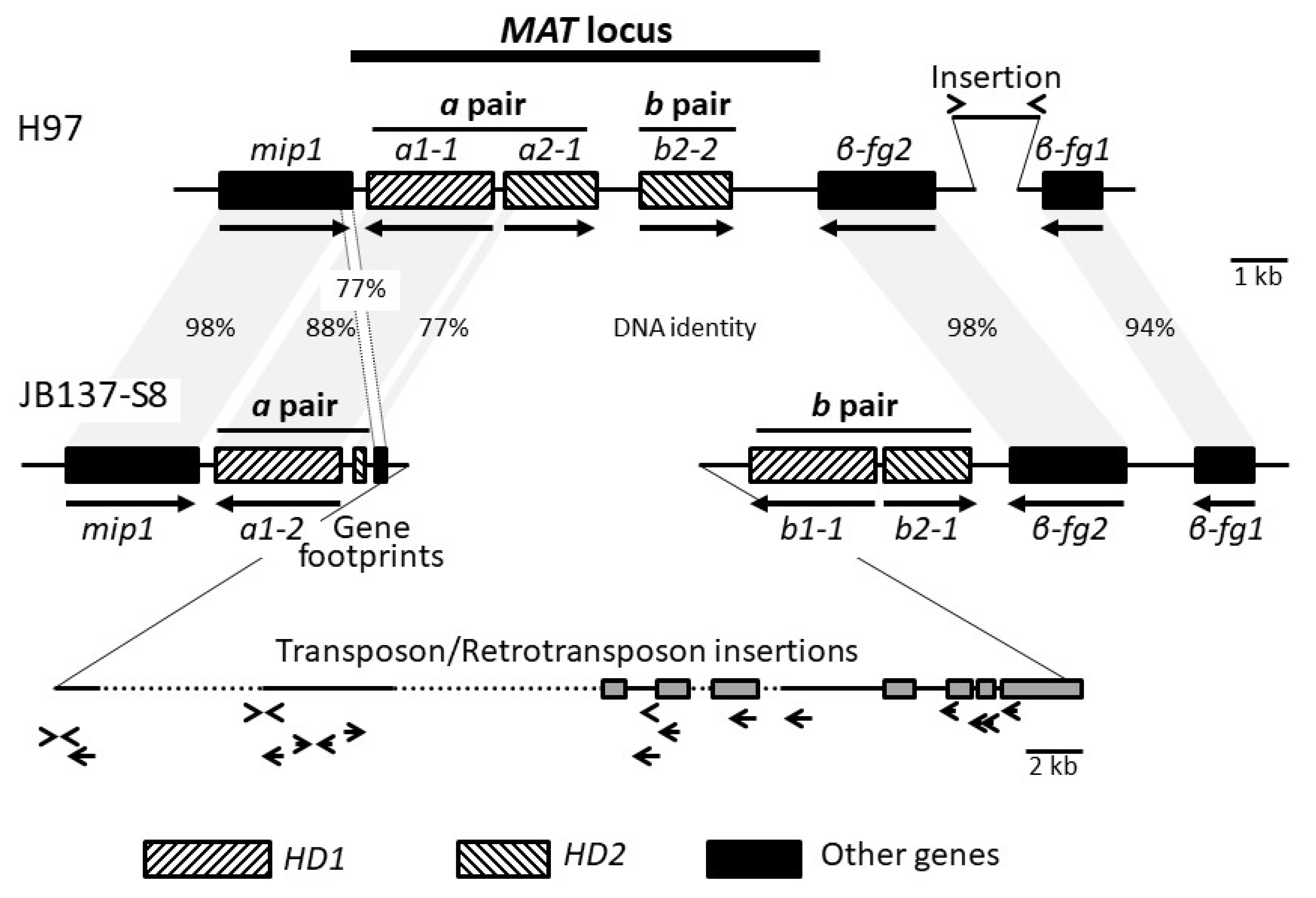
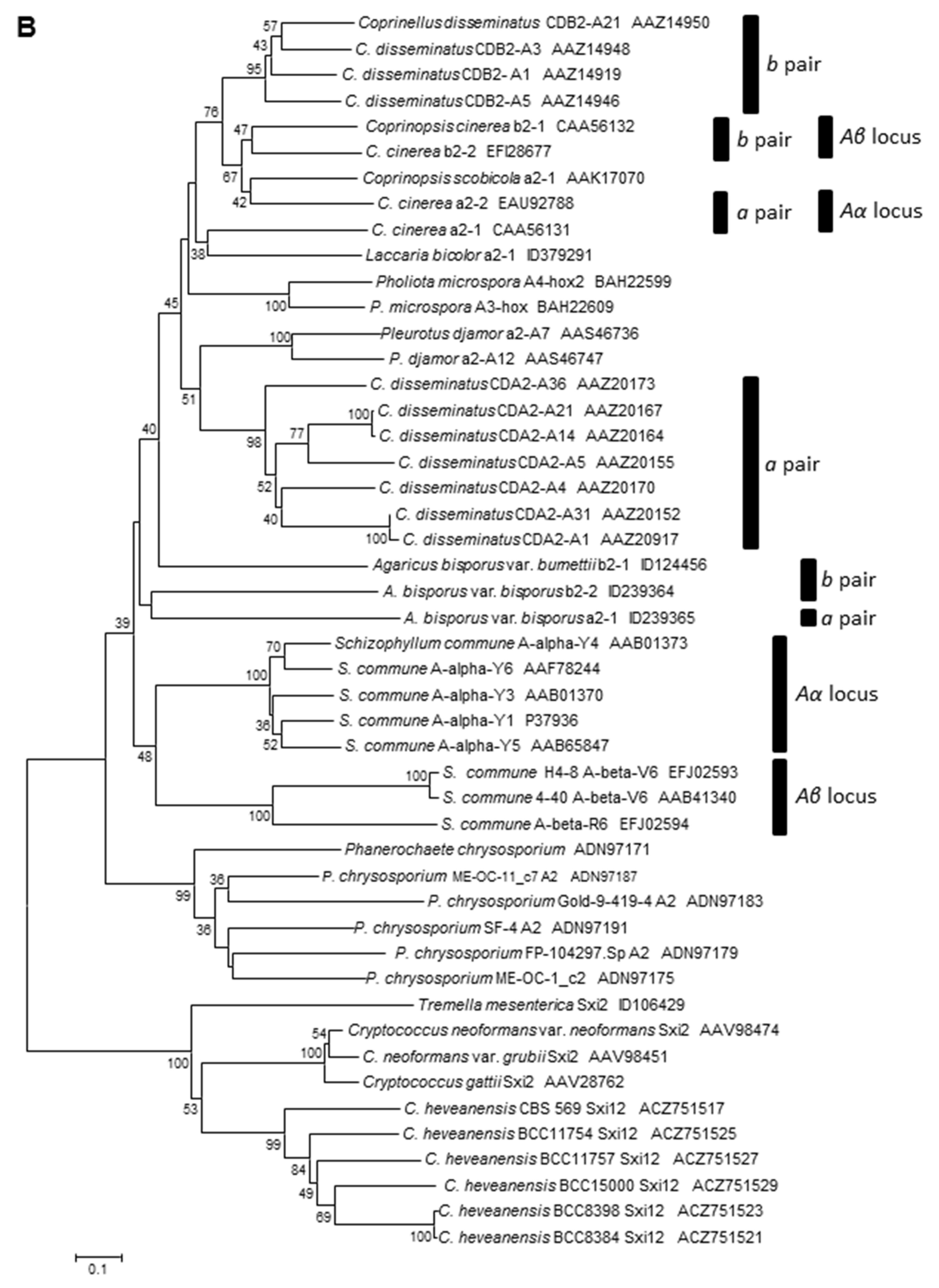
3.2. Identification and Structure of the Mating-Type Homeodomain Loci
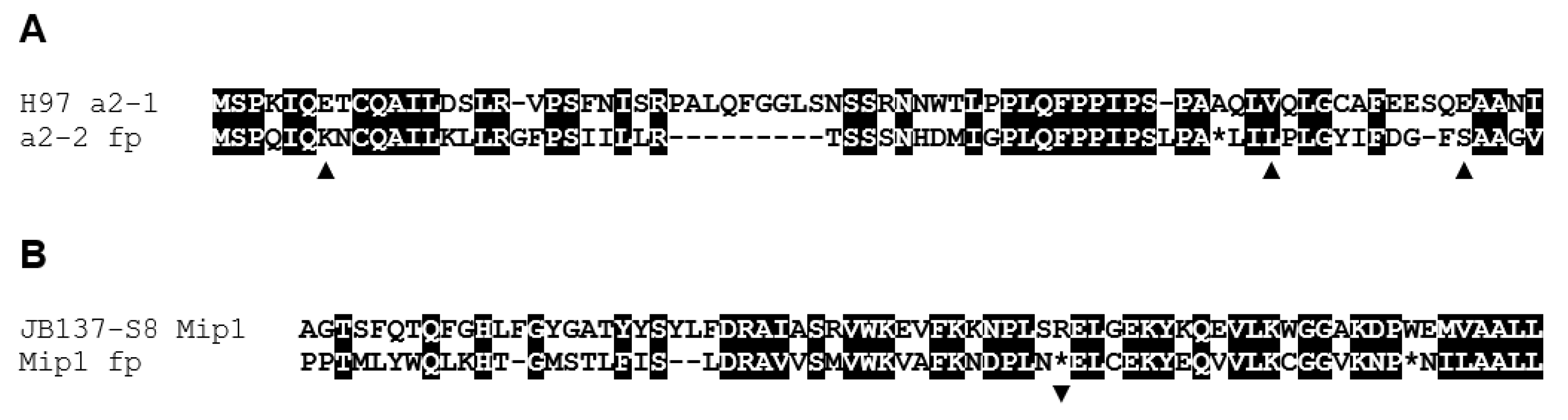
3.3. Orthologous Mating-Type Genes in the A. bisporus var. burnettii JB137-S8 Genome
3.4. MAT Genes and Their Products
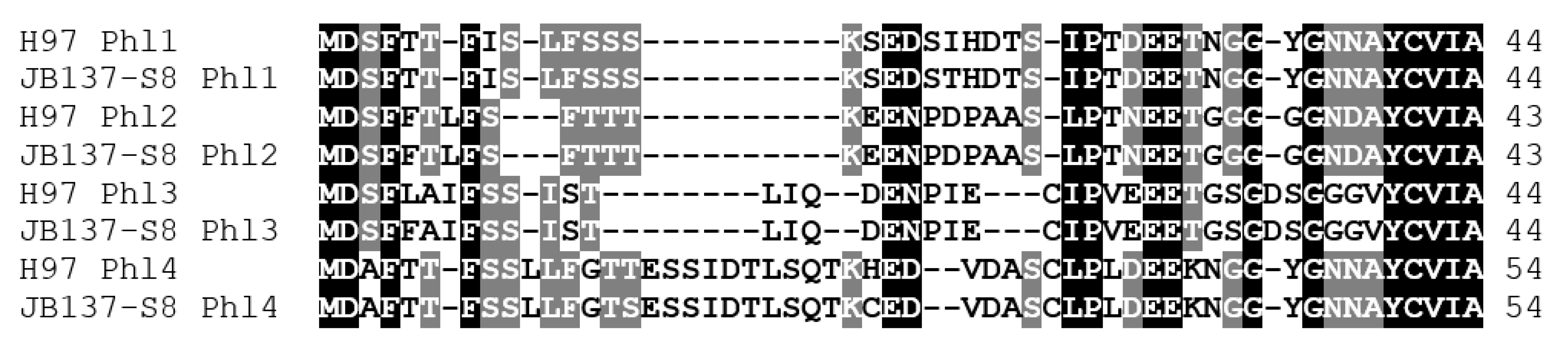
3.5. Genes for Pheromone-Like Peptide Precursors
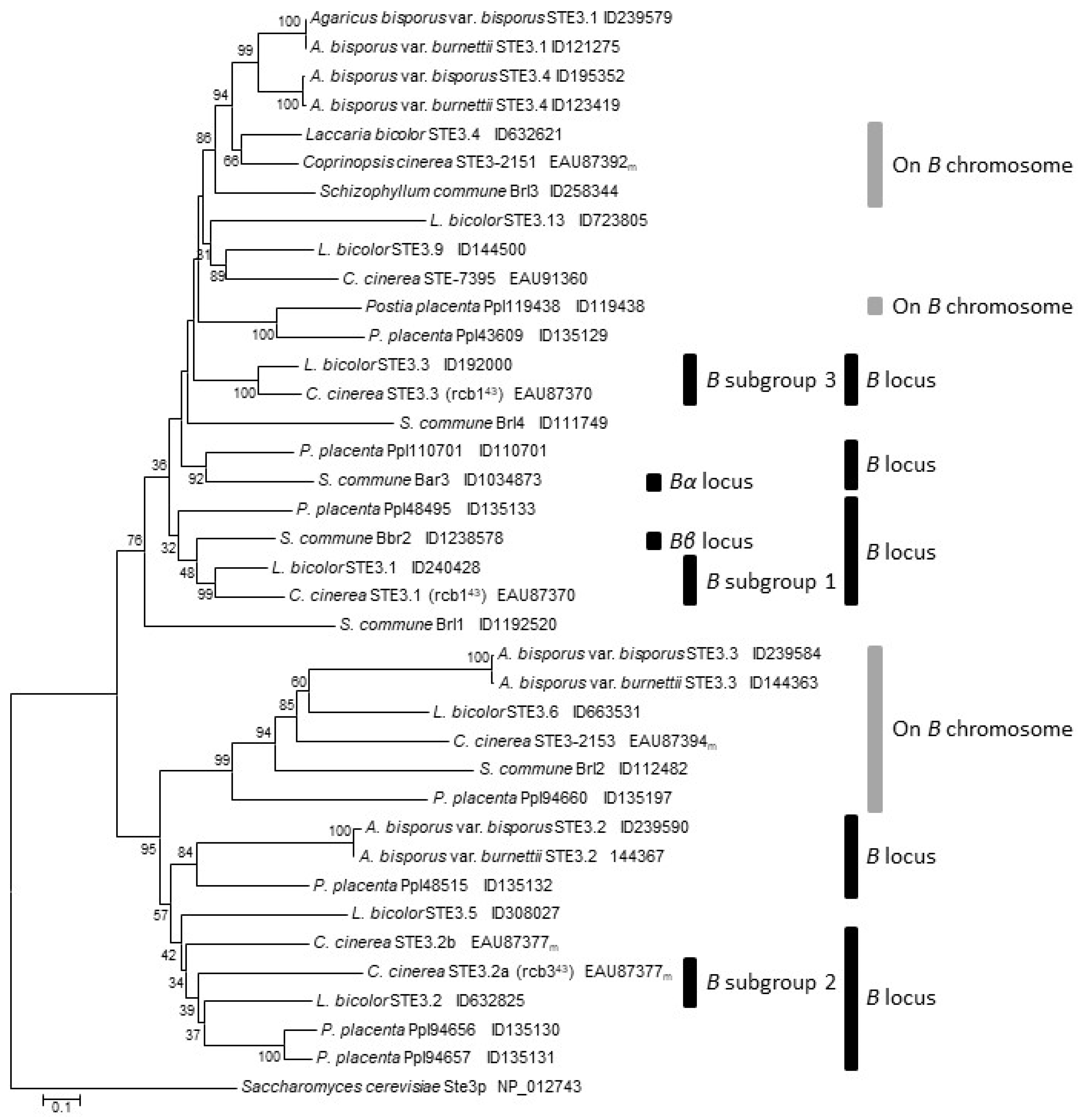
3.6. Genes for Pheromone Receptors
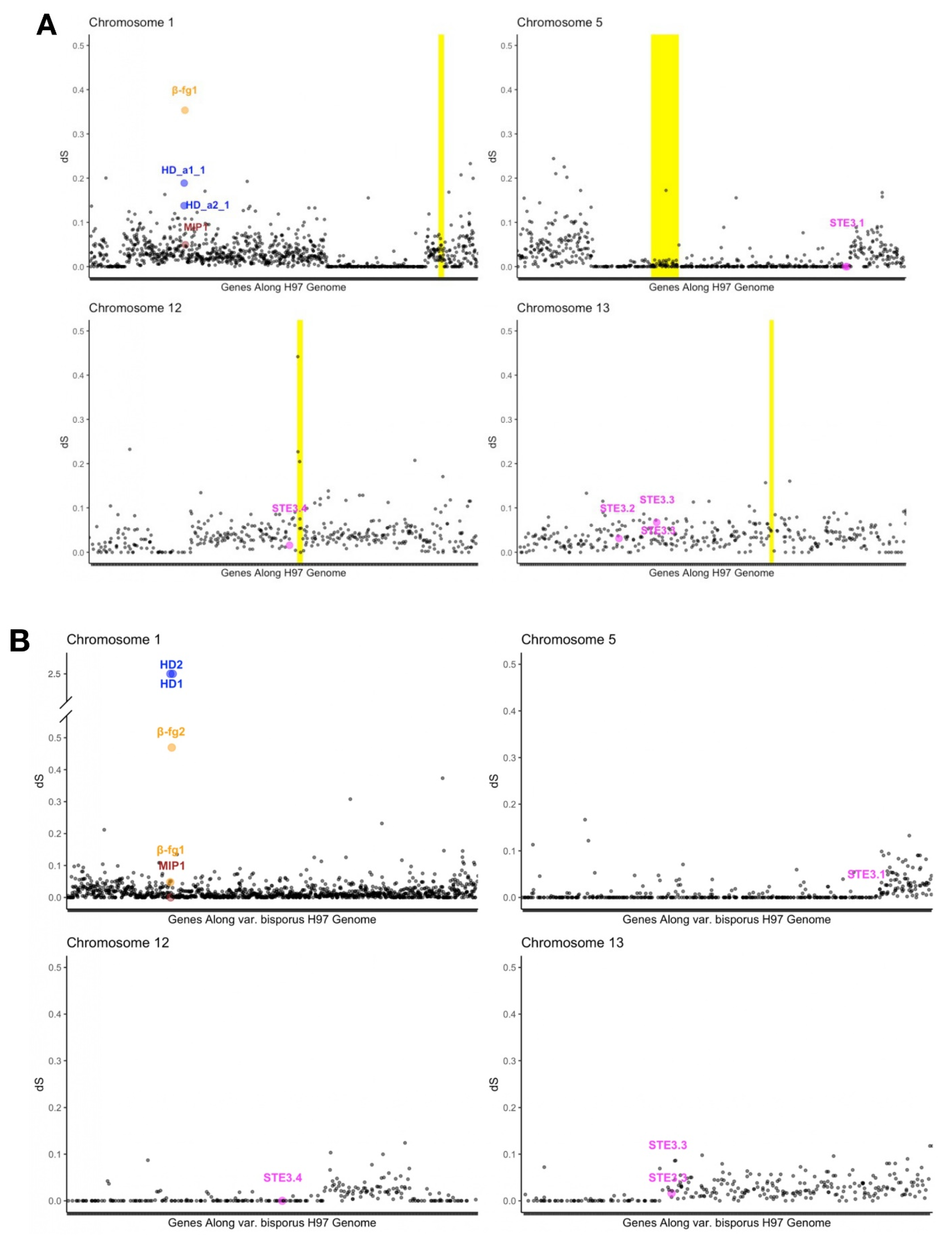
3.7. Differentiation along Mating-Type Chromosomes
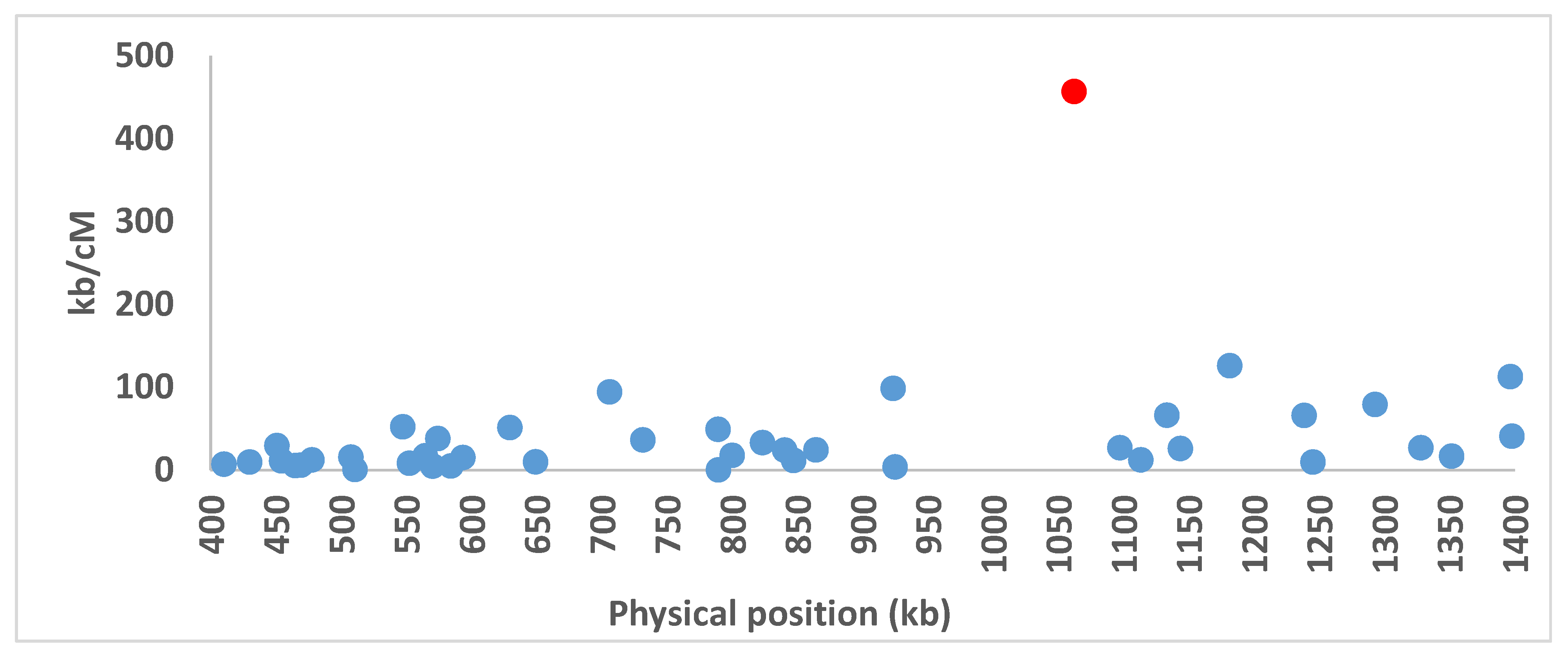
3.8. Recombination Suppression at the Mating-Type Locus in A. bisporus var. burnettii
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lande, R.; Schemske, D.W. The evolution of self-fertilisation and inbreeding depression in plants. 1. Genetic models. Evolution 1985, 39, 24–40. [Google Scholar] [CrossRef]
- Charlesworth, D.; Charlesworth, B. Inbreeding depression and its evolutionary consequences. Annu. Rev. Ecol. Syst. 1987, 18, 237–268. [Google Scholar] [CrossRef]
- Charlesworth, D.; Morgan, M.T.; Charlesworth, B. Inbreeding depression, genetic load, and the evolution of outcrossing rates in a multilocus system with no linkage. Evolution 1990, 44, 1469–1489. [Google Scholar] [CrossRef] [PubMed]
- Hereford, J. Does selfing or outcrossing promote local adaptation? Am. J. Bot 2010, 97, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Igic, B.; Lande, R.; Kohn, J. Loss of self-incompatibility and its evolutionary consequences. Int. J. Plant Sci. 2008, 169, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Lande, R.; Porcher, E. Maintenance of quantitative genetic variance under partial self-fertilization, with implications for evolution of selfing. Genetics 2015, 200, 891–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vekemans, X.; Poux, C.; Goubet, P.M.; Castric, V. The evolution of selfing from outcrossing ancestors in Brassicaceae: What have we learned from variation at the S-locus? J. Evol. Biol. 2014, 27, 1372–1385. [Google Scholar] [CrossRef]
- Kües, U.; James, T.; Heitman, J. The mycota XIV: Mating type in basidiomycetes: Unipolar, bipolar, and tetrapolar paterns of sexuality. In Evolution of Fungi and Fungal-Like Organisms; Pöggeler, S., Wöstemeyer, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 79–106. [Google Scholar]
- Nieuwenhuis, B.P.S.; Billiard, S.; Vuilleumier, S.; Petit, E.; Hood, M.E.; Giraud, T. Evolution of uni- and bifactorial sexual compatibility systems in fungi. Heredity 2013, 111, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Branco, S.; Carpentier, F.; Rodríguez de la Vega, R.; Badouin, H.; Snirc, A.; Le Prieur, S.; Coelho, M.; de Vienne, D.; Hartmann, F.; Begerow, D.; et al. Multiple convergent supergene evolution events in mating-type chromosomes. Nat. Commun. 2018, 9, 2000. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, F.E.; Duhamel, M.; Carpentier, F.; Hood, M.E.; Foulongne-Oriol, M.; Silar, P.; Malagnac, F.; Grognet, P.; Giraud, T. Recombination suppression and evolutionary strata around mating-type loci in fungi: Documenting patterns and understanding evolutionary and mechanistic causes. New Phytol. 2021, 229, 2470–2491. [Google Scholar] [CrossRef]
- Sun, S.; Coelho, M.; Heitman, J.; Nowrousian, M. Convergent evolution of linked mating-type loci in basidiomycete fungi. PLoS Genet. 2019, 15, e1008365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, T.Y. Why mushrooms have evolved to be so promiscuous: Insights from evolutionary and ecological patterns. Fungal Biol. Rev. 2015, 29, 167–178. [Google Scholar] [CrossRef]
- Branco, S.; Badouin, H.; Rodríguez de la Vega, R.; Gouzy, J.; Carpentier, F.; Aguileta, G.; Siguenza, S.; Brandenburg, J.; Coelho, M.; Hood, M.; et al. Evolutionary strata on young mating-type chromosomes despite lack of sexual antagonism. Proc. Natl. Acad. Sci USA 2017, 114, 7067–7072. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, F.E.; Ament-Velásquez, S.L.; Vogan, A.A.; Gautier, V.; Le Prieur, S.; Berramdane, M.; Snirc, A.; Johannesson, H.; Grognet, P.; Malagnac, F.; et al. Size variation of the nonrecombining region on the mating-type chromosomes in the fungal Podospora anserina species complex. Mol. Biol. Evol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.B.; Petsche, D.M.; Herr, F.B.; Horgen, P.A. Breeding relationships among several species of Agaricus. Can. J. Bot. Rev. Can. Bot. 1984, 62, 1884–1889. [Google Scholar] [CrossRef]
- Callac, P.; Billette, C.; Imbernon, M.; Kerrigan, R.W. A new tetrasporic variety of Agaricus bisporus occurs below sea level in the Sonoran desert of California. Mycologia 1993, 85, 835–851. [Google Scholar] [CrossRef]
- Callac, P.; de Haut, I.J.; Imbernon, M.; Guinberteau, J.; Desmerger, C.; Theochari, L. A novel homothallic variety of Agaricus bisporus comprises rare tetrasporic isolates from Europe. Mycologia 2003, 95, 222–231. [Google Scholar] [CrossRef]
- Calvo-Bado, L.; Noble, R.; Challen, M.; Dobrovin-Pennington, A.; Elliott, T. Sexuality and genetic identity in the Agaricus section Arvenses. Appl. Environ. Microbiol. 2000, 66, 728–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, T.J. Comparative sexuality in Agaricus species. J. Gen. Microbiol. 1978, 107, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Raper, C.A.; Kaye, C. Sexual and other relationships in the genus Agaricus. J. Gen. Microbiol. 1978, 105, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Raper, C.A.; Raper, J.R.; Miller, R.E. Genetic analysis of the life cycle of Agaricus bisporus. Mycologia 1972, 64, 1088–1117. [Google Scholar] [CrossRef]
- Xu, J.; Kerrigan, R.W.; Horgen, P.A.; Anderson, J.B. Localization of the mating-type gene in Agaricus bisporus. Appl. Environ. Microbiol. 1993, 59, 3044–3049. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.J.; Moore, D.; Elliott, T.J. Persistent meiotic arrest in basidia of Agaricus bisporus. Mycol. Res. 1992, 96, 125–127. [Google Scholar] [CrossRef]
- Elliott, T.J. Sex and the single spore. Mush. Sci. 1972, 8, 11–18. [Google Scholar]
- Kamzolkina, O.V.; Volkova, V.N.; Kozlova, M.V.; Pancheva, E.V.; Dyakov, Y.T.; Callac, P. Karyological evidence for meiosis in the three different types of life cycles existing in Agaricus bisporus. Mycologia 2006, 98, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A.S.M.; Gao, W.; Lavrijssen, B.; Hendrickx, P.; Sedaghat-Tellgerd, N.; Foulongne-Oriol, M.; Kong, W.S.; Schijlen, E.; Baars, J.J.P.; Visser, R.G.F. A detailed analysis of the recombination landscape of the button mushroom Agaricus bisporus var. bisporus. Fungal Genet. Biol. 2016, 93, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Sonnenberg, A.S.M.; Baars, J.J.P.; Hendrickx, P.-M.; Lavrijssen, B.; Gao, W.; Wejjn, A.; Mes, J.J. Breeding and strain protection in the button mushroom Agaricus bisporus. In Proceedings of the Mushroom Biology and Mushroom Products. In Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Products (ICMBMP7) INRA Bordeaux, Arcachon, France, 4–7 October 2011; pp. 7–15. [Google Scholar]
- Whittle, C.A.; Nygren, K.; Johannesson, H. Consequences of reproductive mode on genome evolution in fungi. Fungal Genet. Biol. 2011, 48, 661–667. [Google Scholar] [CrossRef]
- Hood, M.E.; Antonovics, J.; Koskella, B. Shared forces of sex chromosome evolution in haploids and diploids. Genetics 2004, 168, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Sonnenberg, A.S.M.; Sedaghat-Telgerd, N.; Lavrijssen, B.; Ohm, R.A.; Hendrickx, P.M.; Scholtmeijer, K.; Baars, J.J.P.; van Peer, A. Telomere-to-telomere assembled and centromere annotated genomes of the two main subspecies of the button mushroom Agaricus bisporus reveal especially polymorphic chromosome ends. Sci. Rep. 2020, 10, 14653. [Google Scholar] [CrossRef]
- Kerrigan, R.W.; Royer, J.C.; Baller, L.M.; Kohli, Y.; Horgen, P.A.; Anderson, J.B. Meiotic behavior and linkage relationships in the secondarily homothallic fungus Agaricus bisporus. Genetics 1993, 133, 225–236. [Google Scholar] [CrossRef]
- Xu, J. Analysis of inbreeding depression in Agaricus bisporus. Genetics 1995, 141, 137–145. [Google Scholar] [CrossRef]
- Hou, H.H.; Wu, L.C. Nuclear behaviour of cultivated mushroom. Bot. Bull. Acad. Sin. 1972, 13, 82–91. [Google Scholar]
- Miller, R. Evidence of sexuality in the cultivated mushroom Agaricus bisporus. Mycologia 1971, 63, 630–634. [Google Scholar] [CrossRef]
- Song, S.F.; Hu, K.J.; Hsieh, Y.L. Observations on the spored basidium in the cultivated mushroom (Agaricus bisporus). Mushroom Sci. 1972, 8, 295–303. [Google Scholar]
- Kerrigan, R.W.; Imbernon, M.; Callac, P.; Billette, C.; Olivier, J.M. The heterothallic life cycle of Agaricus bisporus var. burnettii, and the inheritance of its tetrasporic trait. Exp. Mycol. 1994, 18, 193–210. [Google Scholar]
- Foulongne-Oriol, M.; Spataro, C.; Cathalot, V.; Monllor, S.; Savoie, J.M. An expanded genetic linkage map of an intervarietal Agaricus bisporus var. bisporus x A. bisporus var. burnettii hybrid based on AFLP, SSR and CAPS markers sheds light on the recombination behaviour of the species. Fungal Genet. Biol. 2010, 47, 226–236. [Google Scholar] [CrossRef]
- Imbernon, M.; Callac, P.; Granit, S.; Pirobe, L. Allelic polymorphism at the mating type locus in Agaricus bisporus var. burnettii and confirmation of the dominance of its tetrasporic trait. Mush. Sci. 1995, 14, 1–19. [Google Scholar]
- Callac, P.; Hocquart, S.; Imbernon, M.; Desmerger, C.; Oivier, J.-M. Bsn-t alleles from French field strains of Agaricus bisporus. Appl. Environ. Microbiol. 1998, 64, 2105–2110. [Google Scholar] [CrossRef] [Green Version]
- Imbernon, M.; Callac, P.; Gasqui, P.; Kerrigan, R.W.; Velcko, A.J. BSN, the primary determinant of basidial spore number and reproductive mode in Agaricus bisporus, maps to chromosome I. Mycologia 1996, 88, 749–761. [Google Scholar] [CrossRef]
- Kües, U.; Tymon, A.M.; Richardson, W.V.J.; May, G.; Gieser, P.T.; Casselton, L.A. A mating-type factors of Coprinus cinereus have variable numbers of specificity genes encoding 2 classes of homeodomain proteins. Mol. Gen. Genet. 1994, 245, 45–52. [Google Scholar] [CrossRef]
- Casselton, L.A.; Kües, U. The origin of multiple mating types in the model mushrooms Coprinopsis cinerea and Schizophyllum commune. In Sex in Fungi; Heitman, J., Kronstad, J., Taylor, J., Casselton, L., Eds.; ASM Press: Washington, DC, USA, 2007; Volume 88, pp. 283–300. [Google Scholar]
- Findley, K.; Sun, S.; Fraser, J.A.; Hsueh, Y.P.; Averette, A.F.; Li, W.J.; Dietrich, F.S.; Heitman, J. Discovery of a modified tetrapolar sexual cycle in Cryptococcus amylolentus and the evolution of MAT in the Cryptococcus species complex. PLoS Genet. 2012, 8, e1002528. [Google Scholar] [CrossRef] [Green Version]
- Hibbett, D.S.; Donoghue, M.J. Analysis of character correlations among wood decay mechanisms, mating systems, and substrate randes in homobasidiomycetes. Syst. Biol. 2001, 50, 215–242. [Google Scholar] [CrossRef]
- Aimi, T.; Yoshida, R.; Ishikawa, M.; Bao, D.P.; Kitamoto, Y. Identification and linkage mapping of the genes for the putative homeodomain protein (hox1) and the putative pheromone receptor protein homologue (rcb1) in a bipolar basidiomycete, Pholiota nameko. Curr. Genet. 2005, 48, 184–194. [Google Scholar] [CrossRef]
- James, T.Y.; Lee, M.; van Diepen, L.T.A. A single mating-type locus composed of homeodomain genes promotes nuclear migration and heterokaryosis in the white-rot fungus Phanerochaete chrysosporium. Eukaryot. Cell 2011, 10, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, T.Y.; Srivilai, P.; Kües, U.; Vilgalys, R. Evolution of the bipolar mating system of the mushroom Coprinellus disseminatus from its tetrapolar ancestors involves loss of mating-type-specific pheromone receptor function. Genetics 2006, 172, 1877–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, A.; Aerts, A.; Asiegbu, F.; Belbahri, L.; Bouzid, O.; Broberg, A.; Canbäckja, B.; Coutinho, P.M.; Cullen, D.; Dalman, K.; et al. Insight into trade-off between wood decay and parasitism from the genome of a fungal forest pathogen. New Phytol. 2012, 194, 1001–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, R.; Tachikawa, T.; Ishikawa, M.; Mukaiyama, H.; Bao, D.; Aimi, T. Genomic structure of the A mating-type locus in a bipolar basidiomycete, Pholiota nameko. Mycol. Res. 2009, 113, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Morin, E.; Kohler, A.; Baker, A.R.; Foulongne-Oriol, M.; Lombard, V.; Nagy, L.G.; Ohm, R.A.; Patyshakuliyeva, A.; Brun, A.; Aerts, A.L.; et al. Genome sequence of the button mushroom Agaricus bisporus reveals mechanisms governing adaptation to a humic-rich ecological niche. Proc. Natl. Acad. Sci. USA 2012, 109, 17501–17506. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, E.; McGowan, J.; McCarthy, C.G.P.; Amini, A.; Grogan, H.; Fitzpatrick, D.A. Whole genome sequence of the commercially relevant mushroom strain Agaricus bisporus var. bisporus ARP23. G3 2019, 9, 3057–3066. [Google Scholar] [CrossRef] [Green Version]
- Sonnenberg, A.S.; de Groot, P.W.; Schaap, P.J.; Baars, J.J.; Visser, J.; Van Griensven, L.J. Isolation of expressed sequence tags of Agaricus bisporus and their assignment to chromosomes. Appl. Environ. Microbiol. 1996, 62, 4542–4547. [Google Scholar] [CrossRef] [Green Version]
- Foulongne-Oriol, M.; Dufourcq, R.; Spataro, C.; Devesse, C.; Broly, A.; Rodier, A.; Savoie, J.M. Comparative linkage mapping in the white button mushroom Agaricus bisporus provides foundation for breeding management. Curr. Genet. 2011, 57, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newburg, L. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; McGettigan, P.; McWilliam, H.; Valentin, F.; Wallace, I.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, K.R.; Nicholas, K.B.; Nicholas, K.; Nicholas, H.G.; Nicholas, K.; Nicho-Las, H.B.; Deerfield, D.; Nicholas, H.B.J.; Nicholas, H.; Nicholas, H.B.J.; et al. Genedoc: A tool for editing and annoting multiple sequence alignments. Embnet. News 1997. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Ashok Kumar, T. CFSSP: Chou and Fasman Secondary Structure Prediction server. Wide Spectrum Res. J. 2013, 1, 15–19. [Google Scholar]
- Hofmann, K.; Stoffel, W. TMbase—A database of membrane spanning proteins segments. Biol. Chem. Hoppe-Seyler 1993, 374, 166. [Google Scholar]
- Sonnhammer, E.L.L.; von Heijne, G.; Krogh, A. A hidden Markov model for predicting transmembrane helices in protein sequences. In Proceedings of the Sixth International Conference on Intelligent Systems for Molecular Biology, Montreal, QC, Canada, 28 June–1 July 1998; pp. 175–182. [Google Scholar]
- Juretic, D.; Zoranic, L.; Zucic, D. Basic charge clusters and predictions of membrane protein topology. J. Chem. Inf. Comput. Sci. 2002, 42, 620–632. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol 2013, 20, 714–737. [Google Scholar] [CrossRef] [Green Version]
- Hoff, K.J.; Lange, S.; Lomsadze, A.; Borodovsky, M.; Stanke, M. BRAKER1: Unsupervised RNA-seq-based genome annotation with GeneMark-ET and AUGUSTUS. Bioinformatics 2016, 32, 767–769. [Google Scholar] [CrossRef]
- Hoff, K.J.; Lomsadze, A.; Borodovsky, M.; Stanke, M. Whole-genome annotation with BRAKER. Methods Mol. Biol. 2019, 1962, 65–95. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Diekhans, M.; Baertsch, R.; Häussler, D. Using native and syntenically mapped cDNA alignments to improve de novo gene finding. Bioinformatics 2008, 24, 637–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanke, M.; Keller, O.; Gunduz, I.; Hayes, A.; Waack, S.; Morgenstern, B. AUGUSTUS: Ab initio prediction of alternative transcripts. Nucleic Acids Res. 2006, 34, W435–W439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekseth, O.K.; Kuiper, M.; Mironov, V. orthAgogue: An agile tool for the rapid prediction of orthology relations. Bioinformatics 2014, 30, 734–736. [Google Scholar] [CrossRef] [Green Version]
- Ranwez, V.; Harispe, S.; Delsuc, F.; Douzery, E. MACSE: Multiple alignment of coding sequences accounting for frameshifts and stop codons. PLoS ONE 2011, 6, e22594. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Nielsen, R. Estimating synonymous and nonsynonymous substitution rates under realistic evolutionary models. Mol. Biol. Evol. 2000, 17, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team, R.C. Others R: A Language and Environment for Statistical Computing; GBIF: Copenhagen, Denmark, 2013. [Google Scholar]
- Foulongne-Oriol, M.; Murat, C.; Castanera, R.; Ramirez, L.; Sonnenberg, A.S.M. Genome-wide survey of repetitive DNA elements in the button mushroom Agaricus bisporus. Fungal Genet. Biol. 2013, 55, 6–21. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Bewick, A.J.; Hofmeister, B.T.; Powers, R.A.; Mondo, S.J.; Grigoriev, I.V. Diversity of cytosine methylation across the fungal tree of life. Nat. Ecol. Evol. 2019, 3, 479–490. [Google Scholar] [CrossRef]
- Krueger, F.; Galore, T. A wrapper tool around Cutadapt and FastQC to consistently apply quality and adapter trimming to FastQ files. Github 2015, 516, 517. [Google Scholar]
- Krueger, F.; Andrews, S.R. Bismark: A flexible aligner and methylation caller for Bisulfite-Seq applications. Bioinformatics 2011, 27, 1571–1572. [Google Scholar] [CrossRef]
- Collard, B.; Mace, E.; McPhail, M.; Wenzl, P.; Cakir, M.; Fox, G.; Poulsen, D.; Jordan, D. How accurate are the marker orders in crop linkage maps generated from large marker datasets? Crop. Pasture Sci. 2009, 60. [Google Scholar] [CrossRef]
- Duboule, D. Guidebook to the Homeobox Genes; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Bürglin, T.R. Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domain conserved between plants and animals. Nucleic Acids Res. 1997, 25, 4173–4180. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.J.; Casselton, L.A. Mating in mushrooms: Increasing the chances but prolonging the affair. Trends Genet. 2001, 17, 393–400. [Google Scholar] [CrossRef]
- Kües, U.; Casselton, L.A. The origin of multiple mating types in mushrooms. J. Cell Sci. 1993, 104, 227–230. [Google Scholar] [CrossRef]
- Kües, U.; Casselton, L.A. Homedomains and regulation of sexual development in Basidiomycetes. Trends Genet. 1992, 8, 154–155. [Google Scholar] [CrossRef]
- Kües, U.; Asante-Owusu, R.N.; Mutasa, E.S.; Tymon, A.M.; Pardo, E.H.; O’Sbhea, S.F.; Göttgens, B.; Casselton, L.A. Two classes of homeodomain proteins specify the multiple A mating types of the mushroom Coprinus cinereus. Plant. Cell 1994, 6, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Niculita-Hirzel, H.; Labbe, J.; Kohler, A.; le Tacon, F.; Martin, F.; Sanders, I.R.; Kües, U. Gene organization of the mating type regions in the ectomycorrhizal fungus Laccaria bicolor reveals distinct evolution between the two mating type loci. New Phytol. 2008, 180, 329–342. [Google Scholar] [CrossRef]
- Stajich, J.E.; Wilke, S.K.; Ahren, D.; Au, C.H.; Birren, B.W.; Borodovsky, M.; Burns, C.; Canbäck, B.; Casselton, L.A.; Cheng, C.K.; et al. Insights into evolution of multicellular fungi from the assembled chromosomes of the mushroom Coprinopsis cinerea (Coprinus cinereus). Proc. Natl. Acad. Sci. USA 2010, 107, 11889–11894. [Google Scholar] [CrossRef] [Green Version]
- Yi, R.; Mukaiyama, H.; Tachikawa, T.; Shimomura, N.; Aimi, T. A-mating -type gene expression can drive clamp formation in the bipolar mushroom Pholiota microspora (Pholiota nameko). Eukaryot. Cell 2010, 9, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Martinez, D.; Challacombe, J.; Morgenstern, I.; Hibbett, D.; Schmoll, M.; Kubicek, C.P.; Ferreira, P.; Ruiz-Duenas, F.J.; Martinez, A.T.; Kersten, P.; et al. Genome, transcriptome, and secretome analysis of wood decay fungus Postia placenta supports unique mechanisms of lignocellulose conversion. Proc. Natl. Acad. Sci. USA 2009, 106, 1954–1959. [Google Scholar] [CrossRef] [Green Version]
- James, T.Y.; Liou, S.R.; Vilgalys, R. The genetic structure and diversity of the A and B mating-type genes from the tropical oyster mushroom, Pleurotus djamor. Fungal Genet. Biol. 2004, 41, 813–825. [Google Scholar] [CrossRef]
- Kües, U.; James, T.Y.; Vilgalys, R.; Challen, M.P. The chromosomal region containing pab-1, mip, and the A mating type locus of the secondarily homothallic homobasidiomycete Coprinus bilanatus. Curr. Genet. 2001, 39, 16–24. [Google Scholar] [CrossRef]
- Ohm, R.A.; de Jong, J.F.; Lugones, L.G.; Aerts, A.; Kothe, E.; Stajich, J.E.; de Vries, R.P.; Record, E.; Levasseur, A.; Baker, S.E.; et al. Genome sequence of the model mushroom Schizophyllum commune. Nat. Biotechnol. 2010, 28, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Casselton, L.A.; Olesnicky, N.S. Molecular genetics of mating recognition in basidiomycete fungi. Microbiol. Mol. Biol. Rev. 1998, 62, 55–77. [Google Scholar] [CrossRef] [Green Version]
- Pardo, E.H.; Oshea, S.F.; Casselton, L.A. Multiple versions of the a mating type locus of Coprinus cinereus are generated by three paralogous pairs of multiallelic homeobox genes. Genetics 1996, 144, 87–94. [Google Scholar] [CrossRef]
- Specht, C.A.; Stankis, M.M.; Novotny, C.P.; Ullrich, R.C. Mapping the heterologous DNA region that determines the 9 Aα mating type specificities of Schizophyllum commune. Genetics 1994, 137, 709–714. [Google Scholar] [CrossRef]
- Spit, A.; Mellor, E.J.C.; Casselton, L.A. A role for heterodimerization in nuclear localization of a homeodomain protein. Proc. Natl. Acad. Sci USA 1998, 95, 6228–6233. [Google Scholar] [CrossRef] [Green Version]
- Banham, A.H.; Asante-Owusu, R.N.; Göttgens, B.; Thompson, S.A.J.; Kingsnorth, C.S.; Mellor, E.J.C.; Casselton, L.A. An N-terminal dimerization domain permits homeodomain proteins to choose compatible partners and initiate sexual development in the mushroom Coprinus cinereus. Plant. Cell 1995, 7, 773–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, C.L.; Osier, M.; Novotny, C.P.; Ulrich, R.C. The specificity determinant of the Y mating-type proteins of Schizophyllum commune is also essential for Y-Z protein binding. Genetics 1997, 145, 253–260. [Google Scholar] [CrossRef]
- Caldwell, G.A.; Naider, F.; Becker, J.M. Fungal lipopeptide mating pheromones—A model system for the study of protein prenylation. Microbiol. Rev. 1995, 59, 406–422. [Google Scholar] [CrossRef] [PubMed]
- Kües, U. From two to many: Multiple mating types in Basidiomycetes. Fungal Biol. Rev. 2015, 29, 126–166. [Google Scholar] [CrossRef]
- Halsall, J.R.; Milner, M.J.; Casselton, L.A. Three subfamilies of pheromone and receptor genes generate multiple B mating specificities in the mushroom Coprinus cinereus. Genetics 2000, 154, 1115–1123. [Google Scholar] [CrossRef]
- Riquelme, M.; Challen, M.P.; Casselton, L.A.; Brown, A.J. The origin of multiple B mating specificities in Coprinus cinereus. Genetics 2005, 170, 1105–1119. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, S.F.; Chaure, P.T.; Halsall, J.R.; Olesnicky, N.S.; Leibbrandt, A.; Connerton, I.F.; Casselton, L.A. A large pheromone and receptor gene complex determines multiple B mating type specificities in Coprinus cinereus. Genetics 1998, 148, 1081–1090. [Google Scholar] [CrossRef]
- van Peer, A.F.; Park, S.Y.; Shin, P.G.; Jang, K.Y.; Yoo, Y.B.; Park, Y.J.; Lee, B.M.; Sung, G.H.; James, T.Y.; Kong, W.S. Comparative genomics of the mating-type loci of the mushroom Flammulina velutipes reveals widespread synteny and recent inversions. PLoS ONE 2011, 6, e0022249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kües, U.; Navarro-González, M. Mating-type orthologous genes in the primarily homothallic Moniliophthora perniciosa, the causal agent of Witches’ Broom Disease in cacao. J. Basic Microbiol. 2010, 50, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Coelho, M.A.; David-Palma, M.; Priest, S.J.; Heitman, J. The evolution of sexual reproduction and the mating-type locus: Links to pathogenesis of Cryptococcus human pathogenic fungi. Annu. Rev. Genet. 2019, 53, 417–444. [Google Scholar] [CrossRef]
- James, T.Y.; Sun, S.; Li, W.J.; Heitman, J.; Kuo, H.C.; Lee, Y.H.; Asiegbu, F.O.; Olson, A. Polyporales genomes reveal the genetic architecture underlying tetrapolar and bipolar mating systems. Mycologia 2013, 105, 1374–1390. [Google Scholar] [CrossRef]
- Coelho, M.A.; Sampaio, J.P.; Gonçalves, P. A deviation from the bipolar-tetrapolar mating paradigm in an early diverged Basidiomycete. PLoS Genet. 2010, 6, e1001052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, M.A.; Bakkeren, G.; Sun, S.; Hood, M.E.; Giraud, T. Fungal sex: The basidiomycota. In The Fungal Kingdom, Section 2: Life of Fungi; Gow, N., Heitman, J., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 147–175. [Google Scholar]
- Hsueh, Y.P.; Fraser, J.A.; Heitman, J. Transitions in sexuality: Recapitulation of an ancestral tri- and tetrapolar mating system in Cryptococcus neoformans. Eukaryot. Cell 2008, 7, 1847–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachtrog, D.; Mahajan, S.; Bracewell, R. Massive gene amplification on a recently formed Drosophila Y chromosome. Nat. Ecol. Evol. 2019, 3, 1587. [Google Scholar] [CrossRef]
- Isaya, G.; Sakati, W.; Rollins, R.; Shen, G.; Hanson, L.; Ullrich, R.; Novotny, C. Mammalian mitochondrial intermediate peptidase: Structure, fonction analysis of a new homolog from Schizophyllum commune and relationship to thimet oligopeptidase. Genomics 1995, 28, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Menkis, A.; Jacobson, D.J.; Gustafsson, T.; Johannesson, H. The mating-type chromosome in the filamentous ascomycete Neurospora tetrasperma represents a model for early evolution of sex chromosomes. PLoS Genet. 2008, 4, e1000030. [Google Scholar] [CrossRef] [Green Version]
- Badouin, H.; Hood, M.E.; Gouzy, J.; Aguileta, G.; Siguenza, S.; Perlin, M.H.; Cuomo, C.A.; Fairhead, C.; Branca, A.; Giraud, T. Chaos of rearrangements in the mating-type chromosomes of the anther-smut fungus Microbotryum lychnidis-dioicae. Genetics 2015, 200, 1275–1284. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Weijn, A.; Baars, J.J.P.; Mes, J.J.; Visser, R.G.F.; Sonnenberg, A.S.M. Quantitative trait locus mapping for bruising sensitivity and cap color of Agaricus bisporus (button mushrooms). Fungal Genet. Biol. 2015, 77, 69–81. [Google Scholar] [CrossRef]
- Bergero, R.; Charlesworth, D. The evolution of restricted recombination in sex chromosomes. Trends Ecol. Evol. 2009, 24, 94–102. [Google Scholar] [CrossRef]
- Jay, P.; Tezenas, E.; Giraud, T. A deleterious mutation-sheltering theory for the evolution of sex chromosomes and supergenes. BioRxiv 2021. [Google Scholar] [CrossRef]
- Devier, B.; Aguileta, G.; Hood, M.; Giraud, T. Ancient trans-specific polymorphism at pheromone receptor genes in basidiomycetes. Genetics 2009, 181, 209–223. [Google Scholar] [CrossRef] [Green Version]
- van Diepen, L.T.A.; Olson, A.; Ihrmark, K.; Stenlid, J.; James, T.Y. Extensive trans-specific polymorphism at the mating type locus of the root decay fungus Heterobasidion. Mol. Biol. Evol. 2013, 30, 2286–2301. [Google Scholar] [CrossRef] [Green Version]
- Olesnicky, N.S.; Brown, A.J.; Dowell, S.J.; Casselton, L.A. A constitutively active G-protein-coupled receptor causes mating self-compatibility in the mushroom Coprinus. Embo J. 1999, 18, 2756–2763. [Google Scholar] [CrossRef] [Green Version]
- Olesnicky, N.S.; Brown, A.J.; Honda, Y.; Dyos, S.L.; Dowell, S.J.; Casselton, L.A. Self-compatible B mutants in Coprinus with altered pheromone-receptor specificities. Genetics 2000, 156, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Fowler, T.J.; Mitton, M.F.; Vaillancourt, L.J.; Raper, C.A. Changes in mate recognition through alterations of pheromones and receptors in the multisexual mushroom fungus Schizophyllum commune. Genetics 2001, 158, 1491–1503. [Google Scholar] [CrossRef]
- Wirth, S.; Freihorst, D.; Krause, K.; Kothe, E. What role might non-mating receptors play in Schizophyllum Commune? J. Fungi 2021, 7, 399. [Google Scholar] [CrossRef] [PubMed]
- Kües, U.; Walser, P.J.; Klaus, M.J.; Aebi, M. Influence of activated A and B mating-type pathways on developmental processes in the basidiomycete Coprinus cinereus. Mol. Genet. Genom. 2002, 268, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Dubovoy, C. Class of genes affecting B factor regulated development in Schizophyllum commune. Genetics 1976, 82, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Vollmeister, E.; Schipper, K.; Baumann, S.; Haag, C.; Pohlmann, T.; Stock, J.; Feldbrügge, M. Fungal development of the plant pathogen Ustilago maydis. FEMS Microbiol. Rev. 2012, 36, 59–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, P.R. The structure of the A mating type locus in Coprinus lagopus. Genetics 1960, 45, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Schuurs, T.A.; Dalstra, H.J.P.; Scheer, J.M.J.; Wessels, J.G.H. Positioning of nuclei in the secondary mycelium of Schizophyllum commune in relation to differential gene expression. Fungal Genet. Biol. 1998, 23, 150–161. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foulongne-Oriol, M.; Taskent, O.; Kües, U.; Sonnenberg, A.S.M.; van Peer, A.F.; Giraud, T. Mating-Type Locus Organization and Mating-Type Chromosome Differentiation in the Bipolar Edible Button Mushroom Agaricus bisporus. Genes 2021, 12, 1079. https://doi.org/10.3390/genes12071079
Foulongne-Oriol M, Taskent O, Kües U, Sonnenberg ASM, van Peer AF, Giraud T. Mating-Type Locus Organization and Mating-Type Chromosome Differentiation in the Bipolar Edible Button Mushroom Agaricus bisporus. Genes. 2021; 12(7):1079. https://doi.org/10.3390/genes12071079
Chicago/Turabian StyleFoulongne-Oriol, Marie, Ozgur Taskent, Ursula Kües, Anton S. M. Sonnenberg, Arend F. van Peer, and Tatiana Giraud. 2021. "Mating-Type Locus Organization and Mating-Type Chromosome Differentiation in the Bipolar Edible Button Mushroom Agaricus bisporus" Genes 12, no. 7: 1079. https://doi.org/10.3390/genes12071079