
Gene Conversion amongst Alu SINE Elements


1
Institute of Experimental Pathology, ZMBE, University of Münster, 48149 Münster, Germany
2
EvoPAD-RTG, University of Münster, 48149 Münster, Germany
*
Authors to whom correspondence should be addressed.
Genes 2021, 12(6), 905; https://doi.org/10.3390/genes12060905
Submission received: 9 May 2021
/
Revised: 30 May 2021
/
Accepted: 8 June 2021
/
Published: 11 June 2021
(This article belongs to the Special Issue Nucleotide Sequences and Genome Organization)
Abstract
:The process of non-allelic gene conversion acts on homologous sequences during recombination, replacing parts of one with the other to make them uniform. Such concerted evolution is best described as paralogous ribosomal RNA gene unification that serves to preserve the essential house-keeping functions of the converted genes. Transposed elements (TE), especially Alu short interspersed elements (SINE) that have more than a million copies in primate genomes, are a significant source of homologous units and a verified target of gene conversion. The consequences of such a recombination-based process are diverse, including multiplications of functional TE internal binding domains and, for evolutionists, confusing divergent annotations of orthologous transposable elements in related species. We systematically extracted and compared 68,097 Alu insertions in various primates looking for potential events of TE gene conversion and discovered 98 clear cases of Alu–Alu gene conversion, including 64 cases for which the direction of conversion was identified (e.g., AluS conversion to AluY). Gene conversion also does not necessarily affect the entire homologous sequence, and we detected 69 cases of partial gene conversion that resulted in virtual hybrids of two elements. Phylogenetic screening of gene-converted Alus revealed three clear hotspots of the process in the ancestors of Catarrhini, Hominoidea, and gibbons. In general, our systematic screening of orthologous primate loci for gene-converted TEs provides a new strategy and view of a post-integrative process that changes the identities of such elements.
1. Introduction
Genomes of most eukaryotic organisms contain a large number of repetitive sequences, a notable portion of which is composed of transposable elements (TE). For example, TEs occupy up to 69% of human genomes [1]. Despite the large numbers of TEs, only a few “master-copies” can actively propagate [2,3]. Accumulating changes in master copies leads to new subfamilies and types of TEs that commonly differ by several diagnostic sites and spread in limiting activity waves through the genome [4].
Due to their repetitive nature, high similarity, and large quantities in the genome, TEs present a significant substrate for non-allelic gene conversion. Gene conversion is a process whereby the genetic material of a donor sequence unidirectionally replaces that of a homologous acceptor sequence via recombination after a double-strand DNA break. Thus, gene conversion can proliferate mutations among TEs independently of the activity of the master-copy, leading to TE homogenization, a phenomenon known as concerted evolution [5]. Earlier retrotransposon studies reported a few cases of gene conversion between TE copies. For example, Kass and colleagues [6] described a case of gene conversion that changed a younger human Alu SINE to an older element. Roy et al. [7] suggested that gene conversion is responsible for ~10–20% of the variation in the young AluYa5 subfamily. A whole-genome gene conversion analysis among Alus in humans [8] focused on non-diagnostic mutations in Alu sequences revealed significant levels of gene conversion, especially among neighboring Alus. The authors found that gene conversion acts on Alus within a range of about 10 kb, inversely proportional to their distance from one another. Most studies of gene conversion between TEs focused on Alu SINEs in primates. However, similar effects were also reported for LTRs in other mammals [9,10] and in plants [11].
As the vast majority of genomic TEs are neutral to the effects of natural selection, gene conversion does not usually have a crucial impact on the organism. However, TE gene conversion can have an adaptive effect in rewiring regulatory networks (reviewed in [12]). For example, gene conversion among ISX TEs might be responsible for optimizing binding sites for the dosage compensation complex in Drosophila [13].
Gene conversion may also directly influence the evolution of TEs [12,14]. Transferring mutations to master copies may increase or reduce their activity, as proposed for example for the AluYh3a3 subfamily [15]. Moreover, gene conversion might lead to the formation of new TE families or help maintain them in endosymbiont genomes by preventing their degradation and loss [16].
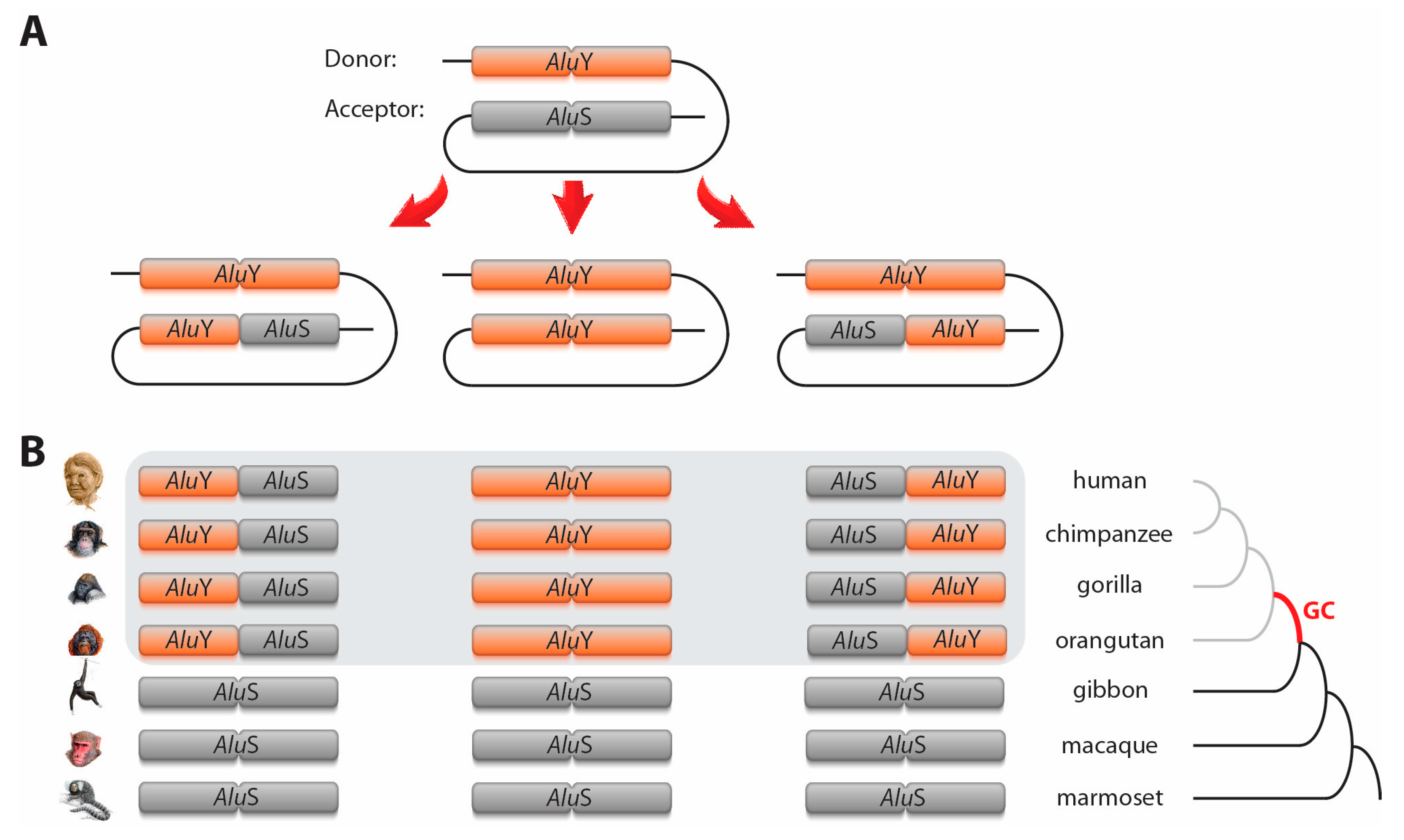
The extent of sequence similarity between donor and acceptor loci positively influences the frequency of gene conversion, and reaches an optimum at 89%–100% [17,18]. Therefore, a substantial number of gene conversion events involves young TEs of the same subfamily. Alu elements are the most abundant TEs in primate genomes and have served as a model group for TE-based gene conversion studies e.g., [7,8]. Alus evolved from 7 SL RNA around 65 million years ago in the ancestral lineage of primates and consist of dimeric sequences of about 300 nt (merged 5′- and 3′-monomers [19]). They diverged into three subfamilies/types—the oldest AluJ, the AluS, and the youngest AluY. More than a million Alu copies are distributed across the human genome, occupying about 11% of genomic space [20]. Because gene conversion also acts on relatively short sequences (beginning with 10 nt [17]), not only are entire Alu sequences substituted, but also partial Alu–Alu gene conversion occurs, resulting in hybrid elements (e.g., hybrids with 5′-AluS and 3′-AluY [21,22], Figure 1).
Changing the TE type via gene conversion might impact the global genome architecture and, for genome scientists, may also lead to faulty genome annotations and obstruct TE-based phylogenetic reconstructions. The phylogenetic presence of identical orthologous TE elements in several species indicates their close relationships, the identification of which can be compromised if gene conversion results in altered element types. We previously showed that parallel insertions and precise deletions of Alus are rare in primates, confirming their usefulness as virtually homoplasy-free markers in phylogenetic studies [23]. However, no study has yet evaluated gene conversion as an additional possible source of confounding TE presence/absence patterns. Replacing one Alu type with another in a monophyletic species group can lead to an incorrect conclusion about their phylogenomic relationship. Therefore, to determine the extent of possible homoplasy caused by gene conversion and the frequency of gene conversion in TEs of different ages, we performed a systematic screening for gene conversion among Alu elements belonging to clearly different primate Alu subfamilies and types (AluY/AluS and AluY/AluYc).
2. Materials and Methods
To identify potential incidences of Alu SINE gene conversion, we initially surveyed a published human genome RepeatMasker file to find all Alu SINE insertions. Because recurrent Alu clusters inhibit finding orthologous loci, we used fastCOEX to extract (nearly) complete human Alu elements with flanks largely free of additional TE sequences. The selected loci were then submitted to the 2-n-way computer suite to find all orthologous sequences of related primates. A subsequent RepeatMasker analysis of all individual hominoid TE regions revealed potential gene converted cases.
2.1. Hg38 Targeting AluY/Yc and AluS Sequences
We extracted target Alu element genomic coordinates from the human genome RepeatMasker outfile (December 2013, GRCh38/hg38) from the Genome Browser, University of California Santa Cruz (UCSC) (https://hgdownload.soe.ucsc.edu/goldenPath/hg38/bigZips/, (hg38.fa.out.gz), accessed on 9 June 2021). Using the fastCOEX program (http://retrogenomics.uni-muenster.de/tools/fastCOEX, accessed on 9 June 2021) [24], we selected coordinates of complete and nearly complete Alu elements (≤20 nt 5′-/3′-truncations) located in regions largely free from additional repeats (≥70% of 700 nt flanks repeat-free). Due to the high divergence of older AluJ elements from their consensus sequences, we restricted our analyses to younger AluY/Yc and AluS elements.
2.2. Screening for Orthologous AluS/Y and AluY/Yc in Hominoidea
To screen for orthologous Alu elements in primates we used the 2-n-way suite [25]. We first generated the following 2-way whole-genome alignments (lastz algorithm): human/chimpanzee (hg38/Clint_PTRv2), human/gorilla (hg38/gorGor4), human/orangutan (hg38/Susie_PABv2), and human/gibbon (hg38/Nleu3.0). The 2-ways were then uploaded to the n-way module to perform human hg38 coordinate-based screenings for full-length AluS, AluY, and AluYc elements and orthologous regions in other hominoids. N-way generated presence/absence tables, indicating the presence or absence of Alus for all investigated primate species. We extracted sequences of loci in which (1) the targeted elements were present in all Hominoidea; (2) the targeted elements were present in all Hominidae except gibbon; and (3) the elements were present in human, chimpanzee, and gorilla but absent in orangutan and gibbon.
2.3. Detecting and Processing Cases of Alu Gene Conversion
Using a local version of the RepeatMasker (https://www.repeatmasker.org/RepeatMasker, accessed on 9 June 2021), we identified the family affiliations of AluS, AluY, and AluYc hg38 orthologs for all investigated non-human primate species. For orthologous primate loci in which different Alu element subfamilies/types were detected (e.g., AluY in human vs. AluS in chimpanzee), we performed an extensive alignment analysis using the Phylogenetic Data Editor (PhyDE, version 0.9971, Muenster, Germany). To help reconstruct the evolution of gene-converted loci we supplemented the analyses with orthologous sequences of bonobo and a second gorilla individual (gorGor5). To identify the ancestral states of the Alu insertions, we also analyzed the sequences of at least one representative Old World (Macaca mulatta) and New World (Callithrix jacchus) monkey outgroup. The macaque and marmoset sequences were retrieved via genome blast for potentially gene converted loci and added to the alignments. All alignments were supplemented with respective Alu consensus sequences.
We accepted specific Alu loci as gene conversion cases if orthologous elements were assigned to different element subfamilies/types based on at least three coherent diagnostic nucleotides for AluS/Y or diagnostic indels for AluY/Yc (Supplementary File S1). For AluS/Y gene conversion, we accepted cases where the Alus belonged to one of the AluS and AluY subfamilies (including AluYc), respectively. For AluY/Yc gene conversion, we considered all AluYs (without 12-nt diagnostic deletion at the 5′-end of the consensus) and all AluYs with the specific deletion (AluYc, Yc3, Yc5, Yd2, Yd3, Yd8), respectively. Furthermore, we accepted as gene converted only those loci with deviating Alu types from species that were phylogenetically nested within their related primates (Figure 1). These decisions were based on the concepts that orthologous Alus of differing element subfamilies and types in various related primates cannot be explained solely by single events of independent insertions and must instead be attributed to gene conversion. For example, to explain the presence of an AluY in humans and an AluS in all other hominoids by orthologous independent insertions or precise deletions, we would require five independent insertions (AluY in the human lineage and four independent insertions of AluS in the different hominoid lineages) or, alternatively, an AluS insertion in the ancestral lineage of Hominoidea with a subsequent precise deletion and precise insertion of AluY in the human lineage. Another conceivable scenario includes the specific accumulation of mutations in the diagnostic positions leading by chance to a change of Alu types. However, we accepted only cases in which at least three specific neighboring mutations were necessary to change the Alu subfamily or type. Thus, for all these cases, a single gene conversion event represents the most parsimonious explanation.
2.4. Screening for Hybrid Elements in Human
To search for hybrid Alus in the human genome, we conducted additional analyses. Alu elements were extracted as described in Section 2.1 and the sequences then split into 5′- and 3′-monomers. We ran a local repeat masking for both datasets independently and compared the Alu subfamily/type affiliations of 5′- and 3′-monomer pairs. If they were assigned to different subfamilies/types by at least three coherent diagnostic changes, we performed a manual analysis including other primate species (see Section 2.3) and reconstructed the evolutionary history of gene conversion.
2.5. Screening for Potential Polymorphic Gene Conversion in the Human Population
We supplemented the loci with gene conversion in the human lineage (after the human-chimpanzee split) with genomic information of additional human individuals. We retrieved 35 human genomes available in NCBI for which the ethnical origin was described.
2.6. Counting of Gene Converted Alu TEs
We applied two quality controls: (1) at least two sources of data (assemblies, individuals, species) were used to verify gene conversion, (2) restricting comparisons to secured orthologs by omitting “low complexity” concatenated Alu element regions. To evaluate the “lowest verifiable level” of homoplasy caused by Alu–Alu gene conversion, we followed the calculation published in Doronina et al. for homoplasious deletions and insertions [23]. We applied the human-chimpanzee-rhesus macaque model group and focused on Alus inserted before the Catarrhini diversification. Accordingly, we calculated the ratio of gene converted Alus in the human and chimpanzee lineages compared to the total number of Alu insertions in the common ancestor of Catarrhini.
3. Results and Discussion
Here we present for the first time a systematic, genome-wide screening of primate genomes for clear Alu–Alu element type change via gene conversions. Two recently developed tools were combined to find 98 specific cases of gene conversion. fastCOEX derived Alu loci with almost TE-free flanks, and 2-n-way extracted their orthologous sequences in various primate species. Gene conversion is identifiable when different Alu subfamilies or types recombine (e.g., AluS change to AluY or vice versa). From a RepeatMasker report of the human genome using fastCOEX [24], we extracted human coordinates of 55,408 AluS and 12,689 AluY/Yc full-length elements with flanking regions largely free of other repetitive sequences. However, restricting our screening to these most reliable cases of Alu TEs reduces the total dataset of human Alus (~800.000 for AluS plus AluY) by about a tenth. We used the 2-n-way computer suite to retrieve 46,285 targeted AluS and 8099 AluY/Yc elements orthologous loci for a set of hominoid species (see Section 2.2 under Methods). We then applied a local RepeatMasker analysis to both annotate each hominoid insertion at orthologous positions, and, in a search for human hybrid elements, to compare the element subtypes of 5′- and 3′-Alu monomers for each human Alu. After manual inspection to verify orthology, we identified 98 cases in which some primate species or species groups contained different Alu elements or hybrid Alus compared to the others in the group (Table 1, Supplementary Table S1, and Supplementary File S2). It has to be mentioned that this number underestimates the actual extent of Alu gene converted loci. The more similar elements are, the more probable they involve in gene conversion. However, gene conversion of identical elements is difficult to trace. About half of the identified gene converted Alu elements were located in gene regions (introns or UTRs). The other half was found in the intergenic areas of the genome (Supplementary Table S1). However, because of restricting our survey to Alus free from flanking TEs, we underestimate the portion of Alu–Alu gene conversion in intergenic regions.
For 64 of the 98 gene conversion loci, we were able to reconstruct the original ancestral Alu element type and to determine the direction of gene conversion. For the AluS to AluY conversions (40 loci; Table 1, first three lines), the older AluS elements (AluSs ceased their main activity before the diversification of Catarrhini) were replaced by younger, potentially active AluY elements (AluYs exhibited their main activity starting with the divergence of Catarrhini) [26]. This suggested that young, actively transcribed DNA regions were the preferred donors for gene conversion [27]. However, we also observed incidences in which the reverse process occurred (12 cases; Table 1, line 4–6), providing evidence that old inactive elements might replace young active elements via gene conversion resulting in a sort of “life after death” spreading throughout the genome after silencing. For 14 of the reconstructed 52 loci involving both AluS and AluY elements, we detected gene conversion of the complete acceptor element, whereas in the remaining 38 loci only partial gene conversion occurred, leading to “mosaic” or hybrid elements (e.g., a hybrid of AluS 5′-monomer and AluY 3′-monomer). It should be mentioned that AluY/AluS gene conversion events resulted in hybrids of AluY 5′-monomer and AluS 3′-monomer (12 cases) can also be potentially AluSc8/AluS gene conversion because the 5′-monomer of AluSc8 shares the diagnostic mutations of AluY and the 3′-monomer of AluS. Furthermore, we detected an additional 31 cases of hybrid elements, in which we were unable to assign the pre-conversional state of the Alu elements (Table 1, line 9). We were unable to categorize AluY/AluS hybrids for cases of unidentified ancestral origins because they were indistinguishable from AluSc8 elements. We also observed 12 incidences of gene conversion among AluY and AluYc elements and 3 cases, in which the Alu loci underwent more than one gene conversion event during primate evolution (Table 1).
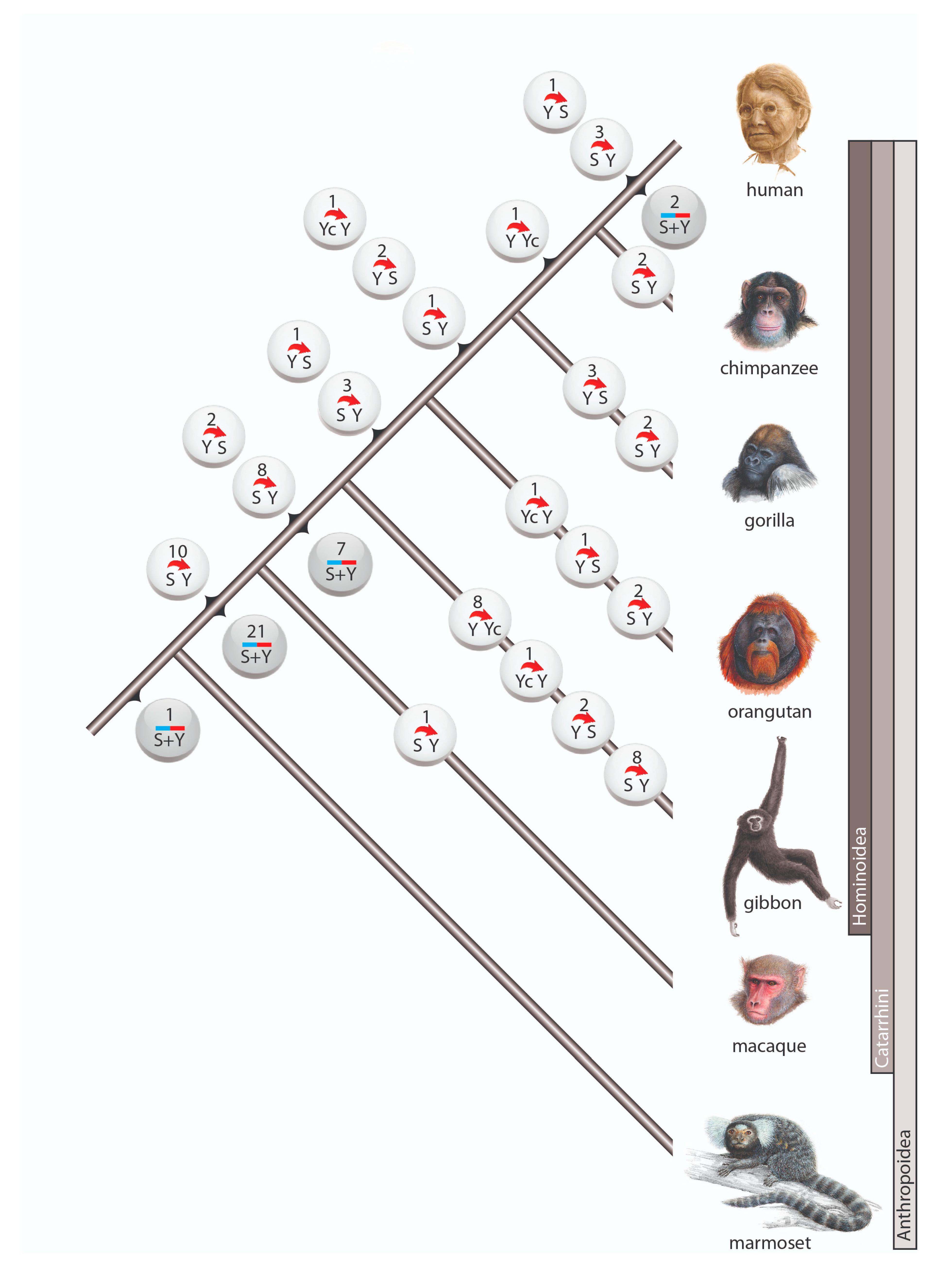
Among the 98 cases of gene conversion, 64 occurred on the lineage leading to humans (including 6 instances after human split from chimpanzee), whereas 31 gene conversions occurred on the terminal branches of other investigated primates (Figure 2). Within Anthropoidea we distinguished three waves of high gene conversion events: (1) on the ancestral branch of Catarrhini (31 conversions), (2) on the ancestral branch of hominoids (17 conversions), and (3) in the gibbon lineage (19 conversions). The first two of these higher incidences might be explained by the longer lengths of the ancestral internodes leading to Catarrhini and hominoids, both leaving substantial times for the occurrence and fixation of gene conversion events. The increased gene conversion events in gibbons might be partially explained by the more highly active gibbon-specific AluY elements (AluYd3a1_gib [28]), which contain the same diagnostic deletion as the AluYc element.
Another gene conversion-rich branch was that leading to gorilla. In our initial analysis, we screened the gorGor4 genome assembly (gorilla Kamilah, UCSC https://genome-euro.ucsc.edu/cgi-bin/hgGateway, accessed on 9 June 2021) and found 12 gene conversions (Supplementary File S3). A previous examination of interlocus gene conversion in gorGor4 [29], also observed a more frequent occurrence of gene conversion in gorilla than in other great apes. However, our expanded analysis of another gorilla genome (gorilla Susie, gorGor5) revealed only 5 gene conversion events (Figure 2, Supplementary Table S1), suggesting that the difference between gorGor4 and gorGor5 is an individual variation or a genomic artifact of the gorGor4 assembly. The gorGor6 assembly (August 2019, assembly Kamilah_GGO_v0/gorGor6) that recently became available carries none of the previously detected cases of gene conversions found solely in gorGor4, suggesting there might be assembly errors in gorGor4. Learning from gorilla, we compared gene conversion patterns for at least two related species or independent assemblies in cases when gene conversion occurred on a terminal branch to avoid such assembly issues.
We conducted a population analysis of human-specific gene conversions (6 cases), including 35 human individual genomes from Africa, Asia, America, and Europe (Supplementary Table S2). We found a consistent gene conversion pattern in all investigated genomes for the 5 loci containing AluS to AluY conversions. For the remaining one locus (AluY converted to AluS), gene conversion was only detected in some human individuals. Contrary to our expectations, we could not find a phylogenetic pattern of the gene conversion distribution among 35 individuals. Orthologous Alu gene conversion was found in 2 of 9 African individuals, 5 of 13 Asian, 1 of 4 American, 1 of 1 Puerto Rican, and 5 of 8 European individuals. We suggest that such a mosaic of gene conversion events might result from duplication of Alu loci in the human genome with the subsequent gene conversion in one of the copies. Alternatively, multiple independent conversions could have occurred.
In the present study, we examined gene conversion events leading to changes in the Alu subfamily or type affiliations in selected hominoids. It should be noted that because of sequence similarity, gene conversion occurs most frequently among identical or closely related elements, and is then unrecognizable. Here we showed that AluS/Y/Yc gene conversion occurred in all hominoid lineages. We suggest that the observed patterns of Alu–Alu gene conversion in hominoids are also representative of other primate species and TE types.
Parallel insertion, exact deletion, or gene conversion might lead to apparently conflicting presence/absence patterns at orthologous loci. Doronina et al. [23] showed there to be a negligibly low frequency of conflicting phylogenetic signals amongst Alu elements in primates. However, they did not examine gene conversion. Although Aleshin et al. [8] found a notable quantity of potential Alu–Alu gene conversions, their screening method (ignoring diagnostic Alu positions) does not evaluate the contribution of gene conversion to homoplasy. Similar to the data in Doronina et al. [23], we estimate the frequency of gene conversion-related homoplasy in the human-chimpanzee-rhesus macaque model group to be 0.0006% in human (3/544,034 × 100%) and 0.0004% in chimpanzee (2/544,034 × 100%), where 544,034 is the number of Alu insertions present in the Catarrhini ancestral lineage. Thus, we provide evidence for the existence of homoplasy caused by gene conversion, but show that the frequency is even lower than parallel insertions or precise deletions.
It should be mentioned that the classical, distance-based (the divergence of a TE sequence from a consensus sequence) calculations of the ages of TEs used in evolutionary studies might be distorted by gene conversion [9,11,30]. Our results suggest that transposition-in-transposition-based analyses [26] that take into account element types rather than accumulated mutations in TE sequences may provide a more reliable alternative. Indeed, we detected relatively few gene conversion events per lineage affecting diagnostic positions that resulted in TE subfamily or type changes, whereas the sharing of non-diagnostic mutations among Alus via gene conversion was shown to be a frequent phenomenon [8].
In summary, the footprints of gene conversion are directly detectable by genome-wide comparisons of deviating annotations of orthologous TEs in different species (e.g., orthologous Alu SINEs with different subfamily or type affiliations in primates). Many potential incidences of partial gene conversion were detected that resulted in hybrid elements. Incidences of gene conversion in TEs are frequent enough to visualize by genome-level screenings but rare enough that they do not challenge large-scale phylogenetic TE presence/absence studies.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/genes12060905/s1, Table S1: List of 98 Alu–Alu gene conversion loci and their genomic and species context, Table S2: Information of 35 additional human individuals including in the analyses for the six human-specific Alu–Alu gene conversion, Supplementary File S1: Pairwise alignment of subfamily or type AluY/AluSz and AluY/AluYc sequences, Supplementary File S2: Alignments of 98 Alu–Alu gene conversion loci. Supplementary File S3: Coordinates of Alu elements in human, which were identified as cases of Alu–Alu gene conversion based on gorilla assembly gorGor4.
Author Contributions
Conceptualization, L.D. and J.S., data collection and analyses, L.D. and O.R., supervision, L.D., writing L.D. and J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are presented in Supplementary Files.
Acknowledgments
We thank Marsha Bundman for editorial assistance, and Jon Baldur Hlidberg ([email protected]) for the paintings of primates. We acknowledge support from the Open Access Publication Fund of the University of Muenster.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Koning, A.P.; Gu, W.; Castoe, T.A.; Batzer, M.A.; Pollock, D.D. Repetitive elements may comprise over two-thirds of the human genome. PLoS Genet. 2011, 7, e1002384. [Google Scholar] [CrossRef] [Green Version]
- Brookfield, J.F.; Johnson, L.J. The evolution of mobile DNAs: When will transposons create phylogenies that look as if there is a master gene? Genetics 2006, 173, 1115–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, A.; Rozhdestvensky, T.S.; Kuryshev, V.Y.; Schmitz, J.; Brosius, J. An unusual primate locus that attracted two independent Alu insertions and facilitates their transcription. J. Mol. Biol. 2005, 350, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Kapitonov, V.; Jurka, J. The age of Alu subfamilies. J. Mol. Evol. 1996, 42, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.R.; Turner, B.J. Concerted evolution of repetitive DNA sequences in eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Kass, D.H.; Batzer, M.A.; Deininger, P.L. Gene conversion as a secondary mechanism of short interspersed element (SINE) evolution. Mol. Cell. Biol. 1995, 15, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.M.; Carroll, M.L.; Nguyen, S.V.; Salem, A.H.; Oldridge, M.; Wilkie, A.O.M.; Batzer, M.A.; Deininger, P.L. Potential gene conversion and source genes for recently integrated Alu elements. Genome Res. 2000, 10, 1485–1495. [Google Scholar] [CrossRef] [Green Version]
- Aleshin, A.; Zhi, D. Recombination-associated sequence homogenization of neighboring Alu elements: Signature of nonallelic gene conversion. Mol. Biol. Evol. 2010, 27, 2300–2311. [Google Scholar] [CrossRef] [Green Version]
- Kijima, T.E.; Innan, H. On the estimation of the insertion time of LTR retrotransposable elements. Mol. Biol. Evol. 2010, 27, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Trombetta, B.; Fantini, G.; D’Atanasio, E.; Sellitto, D.; Cruciani, F. Evidence of extensive non-allelic gene conversion among LTR elements in the human genome. Sci. Rep. 2016, 6, 28710. [Google Scholar] [CrossRef]
- Cossu, R.M.; Casola, C.; Giacomello, S.; Vidalis, A.; Scofield, D.G.; Zuccolo, A. LTR retrotransposons show low levels of unequal recombination and high rates of intraelement gene conversion in large plant genomes. Genome Biol. Evol. 2017, 9, 3449–3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawcett, J.A.; Innan, H. The role of gene conversion between transposable elements in rewiring regulatory networks. Genome Biol. Evol. 2019, 11, 1723–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, C.E.; Bachtrog, D. Non-allelic gene conversion enables rapid evolutionary change at multiple regulatory sites encoded by transposable elements. eLife 2015, 4, e05899. [Google Scholar] [CrossRef]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Styles, P.; Brookfield, J.F. Source gene composition and gene conversion of the AluYh and AluYi lineages of retrotransposons. BMC Evol. Biol. 2009, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Cordaux, R. Gene conversion maintains nonfunctional transposable elements in an obligate mutualistic endosymbiont. Mol. Biol. Evol. 2009, 26, 1679–1682. [Google Scholar] [CrossRef] [Green Version]
- Benovoy, D.; Drouin, G. Ectopic gene conversions in the human genome. Genomics 2009, 93, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Cooper, D.N.; Chuzhanova, N.; Férec, C.; Patrinos, G.P. Gene conversion: Mechanisms, evolution and human disease. Nat. Rev. Genet. 2007, 8, 762–775. [Google Scholar] [CrossRef]
- Kriegs, J.O.; Matzke, A.; Churakov, G.; Kuritzin, A.; Mayr, G.; Brosius, J.; Schmitz, J. Waves of genomic hitchhikers shed light on the evolution of gamebirds (Aves: Galliformes). BMC Evol. Biol. 2007, 7, 190. [Google Scholar] [CrossRef] [Green Version]
- Deininger, P. Alu elements: Know the SINEs. Genome Biol. 2011, 12, 236. [Google Scholar] [CrossRef] [Green Version]
- Batzer, M.A.; Rubin, C.M.; Hellmann-Blumberg, U.; Alegria-Hartman, M.; Leeflang, E.P.; Stern, J.D.; Bazan, H.A.; Shaikh, T.H.; Deininger, P.L.; Schmid, C.W. Dispersion and insertion polymorphism in two small subfamilies of recently amplified human Alu Repeats. J. Mol. Biol. 1995, 247, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Salem, A.-H.; Ray, D.A.; Hedges, D.J.; Jurka, J.; Batzer, M.A. Analysis of the human Alu Ye lineage. BMC Evol. Biol. 2005, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doronina, L.; Reising, O.; Clawson, H.; Ray, D.A.; Schmitz, J. True homoplasy of retrotransposon insertions in primates. Syst. Biol. 2019, 68, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Doronina, L.; Matzke, A.; Churakov, G.; Stoll, M.; Huge, A.; Schmitz, J. The beaver’s phylogenetic lineage illuminated by retroposon reads. Sci. Rep. 2017, 7, 43562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churakov, G.; Zhang, F.; Grundmann, N.; Makalowski, W.; Noll, A.; Doronina, L.; Schmitz, J. The multicomparative 2-n-way genome suite. Genome Res. 2020, 30, 1508–1516. [Google Scholar] [CrossRef]
- Churakov, G.; Grundmann, N.; Kuritzin, A.; Brosius, J.; Makalowski, W.; Schmitz, J. A novel web-based TinT application and the chronology of the Primate Alu retroposon activity. BMC Evol. Biol. 2010, 10, 376. [Google Scholar] [CrossRef] [Green Version]
- Schildkraut, E.; Miller, C.A.; Nickoloff, J.A. Transcription of a donor enhances its use during double-strand break-induced gene conversion in human cells. Mol. Cell. Biol. 2006, 26, 3098–3105. [Google Scholar] [CrossRef] [Green Version]
- Meyer, T.J.; McLain, A.T.; Oldenburg, J.M.; Faulk, C.; Bourgeois, M.G.; Conlin, E.M.; Mootnick, A.R.; De Jong, P.J.; Roos, C.; Carbone, L.; et al. An Alu-based phylogeny of gibbons (Hylobatidae). Mol. Biol. Evol. 2012, 29, 3441–3450. [Google Scholar] [CrossRef] [Green Version]
- Wacholder, A.; Pollock, D.D. PRDM9 and an epidemic of gene conversion and non-homologous recombination among Alu elements in ancestral gorillas. bioRxiv 2017, 241356. [Google Scholar]
- Jedlicka, P.; Lexa, M.; Kejnovsky, E. What can long terminal repeats tell us about the age of LTR retrotransposons, gene conversion and ectopic recombination? Front. Plant Sci. 2020, 11, 644. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of Alu–Alu gene conversion. (A) shows three different scenarios of gene conversion: in which a 5′-end Alu monomer, a complete Alu, or a 3′-end Alu monomer were converted. (B) Representative gene conversion at orthologous AluS loci leading to 5′-end AluY, AluY, and 3′-end AluY in the common ancestor of great apes (grey area; e.g., loci AluS_plus_7691_7692, HGib37,f and H_AluS6 in Supplementary Table S1 and Supplementary File S2, ignoring some variation in marmoset). The phylogenetic time-point of gene conversions is labelled GC (red branch of the tree).
Figure 1.
Schematic representation of Alu–Alu gene conversion. (A) shows three different scenarios of gene conversion: in which a 5′-end Alu monomer, a complete Alu, or a 3′-end Alu monomer were converted. (B) Representative gene conversion at orthologous AluS loci leading to 5′-end AluY, AluY, and 3′-end AluY in the common ancestor of great apes (grey area; e.g., loci AluS_plus_7691_7692, HGib37,f and H_AluS6 in Supplementary Table S1 and Supplementary File S2, ignoring some variation in marmoset). The phylogenetic time-point of gene conversions is labelled GC (red branch of the tree).

Figure 2.
Gene conversion in primates. Circles represent incidences of gene conversion including the number of such occurrences and their direction. White circles are incidences in which the ancestral Alu element and the direction of conversion were reconstructed. Gray circles represent incidences with unidentifiable conversion direction. The 3 cases with complex scenarios are not shown.
Figure 2.
Gene conversion in primates. Circles represent incidences of gene conversion including the number of such occurrences and their direction. White circles are incidences in which the ancestral Alu element and the direction of conversion were reconstructed. Gray circles represent incidences with unidentifiable conversion direction. The 3 cases with complex scenarios are not shown.


{kind=link}
{kind=link}
Table 1.
Gene conversion cases among Alu elements.
| Donor | Acceptor | Replaced Part of Alu | Number of Loci |
|---|---|---|---|
| Gene conversion with identified direction | |||
| AluY | AluS | Complete Alu | 11 |
| AluY | AluS | 3′-Alu unit (S-Y hybrid) | 23 |
| AluY | AluS | 5′-Alu unit (Y-S hybrid) | 6 |
| AluS | AluY | Complete Alu | 3 |
| AluS | AluY | 3′-Alu unit (Y-S hybrid) | 6 |
| AluS | AluY | 5′-Alu unit (S-Y hybrid) | 3 |
| AluYc | AluY | Diagnostic indel | 9 |
| AluY | AluYc | Diagnostic indel | 3 |
| Gene conversion with unidentified direction | |||
| Unidentified | Unidentified | AluS–AluY hybrid | 31 |
| Complex scenario | Complex scenario | Multiple gene conversion | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Doronina, L.; Reising, O.; Schmitz, J. Gene Conversion amongst Alu SINE Elements. Genes 2021, 12, 905. https://doi.org/10.3390/genes12060905
AMA Style
Doronina L, Reising O, Schmitz J. Gene Conversion amongst Alu SINE Elements. Genes. 2021; 12(6):905. https://doi.org/10.3390/genes12060905
Chicago/Turabian StyleDoronina, Liliya, Olga Reising, and Jürgen Schmitz. 2021. "Gene Conversion amongst Alu SINE Elements" Genes 12, no. 6: 905. https://doi.org/10.3390/genes12060905
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.