Myogenic Response to Increasing Concentrations of Ammonia Differs between Mammalian, Avian, and Fish Species: Cell Differentiation and Genetic Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture Media
2.2. Differentiation and Treatment Media
2.3. Myotube Diameter
2.4. RNA Extraction and Real-Time qPCR
2.5. Statistical Analysis
3. Results
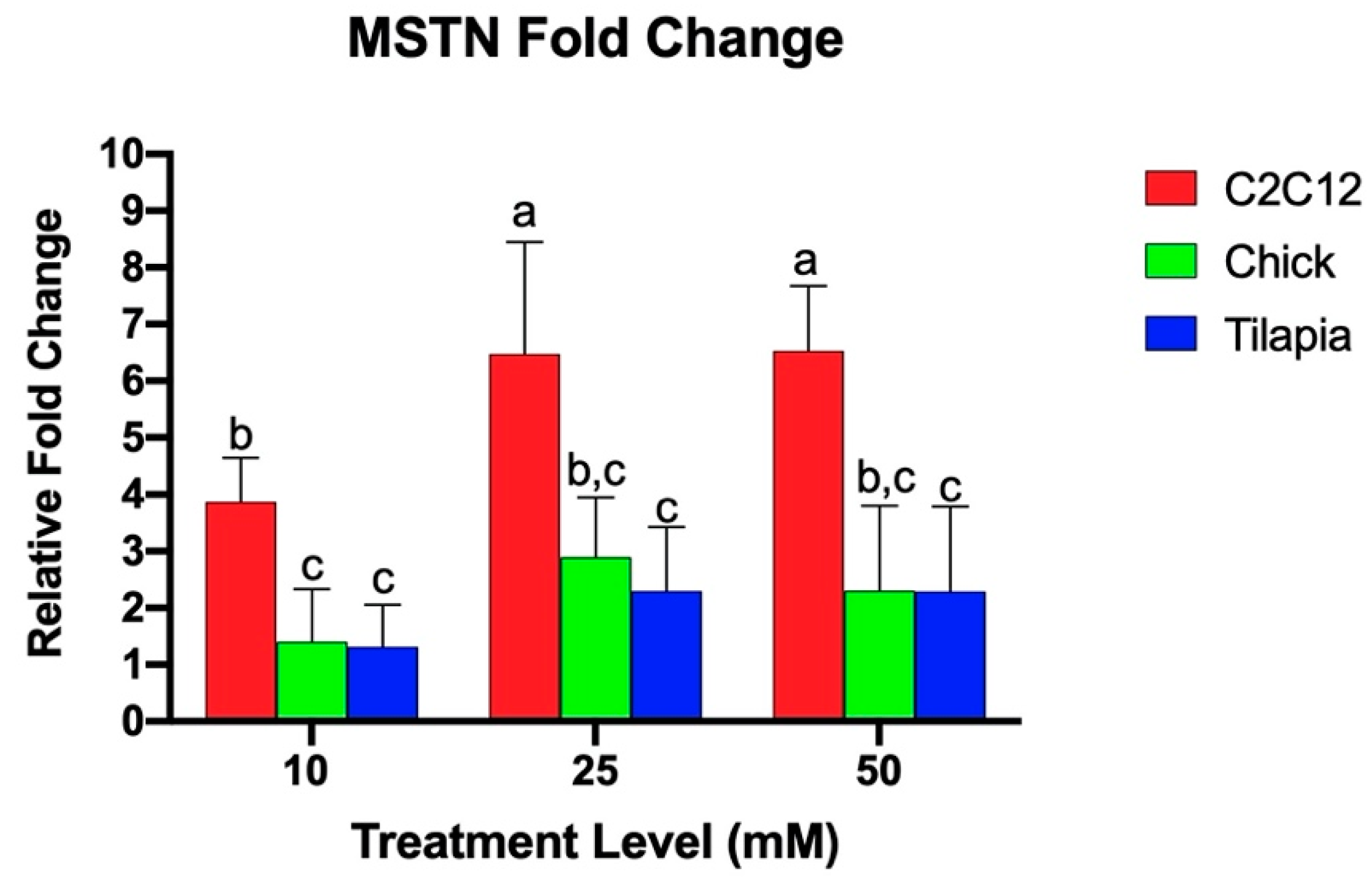
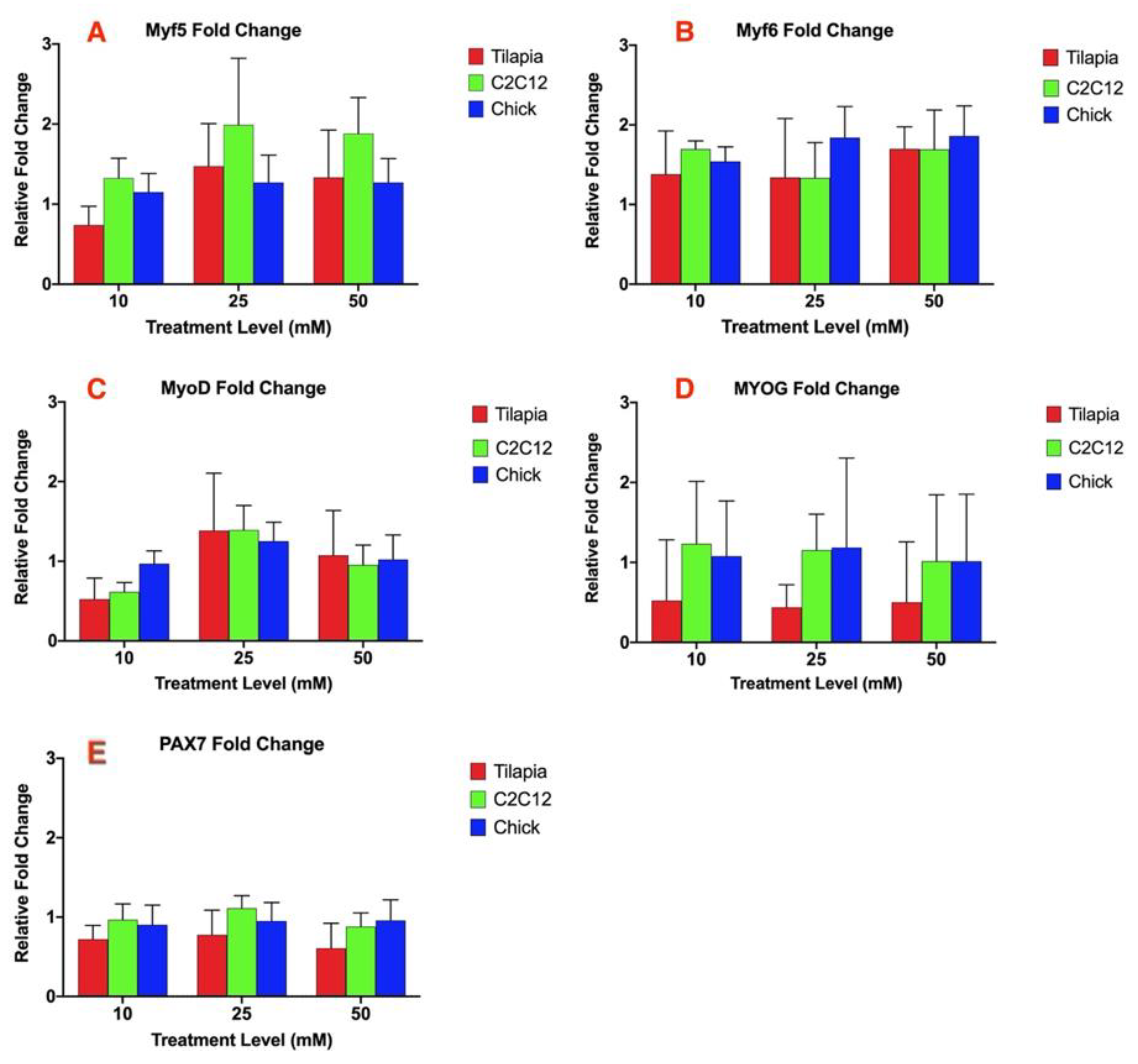
3.1. Myostatin Response to Ammonium Acetate Differs Between Species
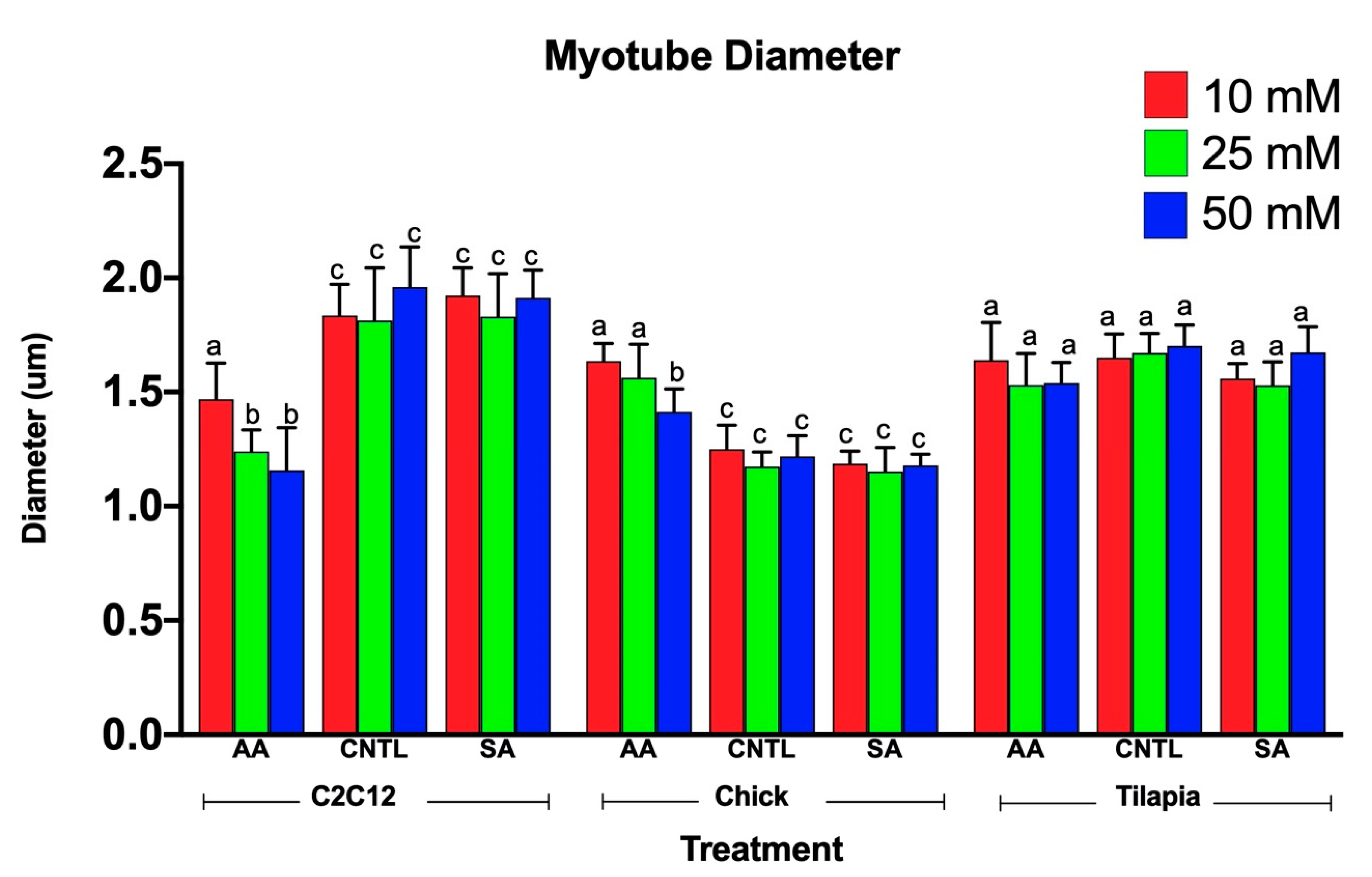
3.2. Myotube Diameter Changed in Response to Myostatin Expression for Mammalian and Avian Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lee, S.J.; McPherron, A.C. Myostatin and the Control of Skeletal Muscle Mass. Curr. Opin. Genet. Dev. 1999, 9, 604–607. [Google Scholar] [CrossRef]
- Amthor, H.; Nicholas, G.; McKinnell, I.; Kemp, C.F.; Sharma, M.; Kambadur, R.; Patel, K. Follistatin Complexes Myostatin and Antagonises Myostatin-Mediated Inhibition of Myogenesis. Dev. Biol. 2004, 270, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Langley, B.; Berry, C.; Sharma, M.; Kirk, S.; Bass, J.; Kambadur, R. Myostatin, a Negative Regulator of Muscle Growth, Functions by Inhibiting Myoblast Proliferation. J. Biol. Chem. 2000, 275, 40235–40243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Topouzis, S.; Liang, L.F.; Stotish, R.L. Myostatin Signaling through Smad2, Smad3 and Smad4 Is Regulated by the Inhibitory Smad7 by a Negative Feedback Mechanism. Cytokine 2004, 26, 262–272. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of Skeletal Muscle Mass in Mice by a New TGF-β Superfamily Member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Kambadur, R.; Sharma, M.; Smith, T.P.L.; Bass, J.J. Mutations in Myostatin (GDF8) in Double-Muscled Belgian Blue and Piedmontese Cattle. Genome Res. 1997, 7, 910–915. [Google Scholar] [CrossRef] [Green Version]
- Acosta, J.; Carpio, Y.; Borroto, I.; González, O.; Estrada, M.P. Myostatin Gene Silenced by RNAi Show a Zebrafish Giant Phenotype. J. Biotechnol. 2005, 119, 324–331. [Google Scholar] [CrossRef]
- Lee, S.B.; Kim, Y.S.; Oh, M.Y.; Jeong, I.H.; Seong, K.B.; Jin, H.J. Improving Rainbow Trout (Oncorhynchus mykiss) Growth by Treatment with a Fish (Paralichthys olivaceus) Myostatin Prodomain Expressed in Soluble Forms in E. coli. Aquaculture 2010, 302, 270–278. [Google Scholar] [CrossRef]
- Dasarathy, S.; Dodig, M.; Muc, S.M.; Kalhan, S.C.; McCullough, A.J. Skeletal Muscle Atrophy Is Associated with an Increased Expression of Myostatin and Impaired Satellite Cell Function in the Portacaval Anastamosis Rat. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, 1124–1130. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, E.N.; Pino, K.; Navarro, C.; Delgado, I.; Valdés, J.A.; Molina, A. Transient Inactivation of Myostatin Induces Muscle Hypertrophy and Overcompensatory Growth in Zebrafish via Inactivation of the SMAD Signaling Pathway. J. Biotechnol. 2013, 168, 295–302. [Google Scholar] [CrossRef]
- Sawatari, E.; Seki, R.; Adachi, T.; Hashimoto, H.; Uji, S.; Wakamatsu, Y.; Nakata, T.; Kinoshita, M. Overexpression of the Dominant-Negative form of Myostatin Results in Doubling of Muscle-Fiber Number in Transgenic Medaka (Oryzias latipes). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 155, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Maccatrozzo, L.; Bargelloni, L.; Cardazzo, B.; Rizzo, G.; Patarnello, T. A Novel Second Myostatin Gene Is Present in Teleost Fish. FEBS Lett. 2001, 509, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Radaelli, G.; Rowlerson, A.; Mascarello, F.; Patruno, M.; Funkenstein, B. Myostatin Precursor Is Present in Several Tissues in Teleost Fish: A Comparative Immunolocalization Study. Cell Tissue Res. 2003, 311, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Rescan, P.Y.; Jutel, I.; Rallière, C. Two Myostatin Genes Are Differentially Expressed in Myotomal Muscles of the Trout (Oncorhynchus mykiss). J. Exp. Biol. 2001, 204, 3523–3529. [Google Scholar] [PubMed]
- Wang, C.; Chen, Y.L.; Bian, W.P.; Xie, S.L.; Qi, G.L.; Liu, L.; Strauss, P.R.; Zou, J.X.; Pei, D.S. Deletion of Mstna and Mstnb Impairs the Immune System and Affects Growth Performance in Zebrafish. Fish Shellfish Immunol. 2018, 72, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.B.; Goetz, F.W. Differential Skeletal Muscle Expression of Myostatin across Teleost Species, and the Isolation of Multiple Myostatin Isoforms. FEBS Lett. 2001, 491, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Tsien, C.; Thapalaya, S.; Narayanan, A.; Weihl, C.C.; Ching, J.K.; Eghtesad, B.; Singh, K.; Fu, X.; Dubyak, G.; et al. Hyperammonemia-Mediated Autophagy in Skeletal Muscle Contributes to Sarcopenia of Cirrhosis. Am. J. Physiol. Endocrinol. Metab. 2012, 303, 983–993. [Google Scholar] [CrossRef] [Green Version]
- Stern, R.A.; Dasarathy, S.; Mozdziak, P.E. Ammonia Elicits a Different Myogenic Response in Avian and Murine Myotubes. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 99–110. [Google Scholar] [CrossRef]
- Stern, R.A.; Ashwell, C.M.; Dasarathy, S.; Mozdziak, P.E. The Effect of Hyperammonemia on Myostatin and Myogenic Regulatory Factor Gene Expression in Broiler Embryos. Animal 2015, 9, 992–999. [Google Scholar] [CrossRef] [Green Version]
- Dosdat, A.; Ruyet, J.P.-L.; Covès, D.; Dutto, G.; Gasset, E.; Roux, A.L.; Lemarié, G. Effect of Chronic Exposure to Ammonia on Growth, Food Utilisation and Metabolism of the European Sea Bass (Dicentrarchus labrax). Aquat. Living Resour. 2003, 16, 509–520. [Google Scholar] [CrossRef]
- Meijer, A.; Lamers, W.; Chamuleau, R. Nitrogen Metabolism and Ornithine Cycle Function. Physiol. Rev. 1990, 70, 701–748. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.W.; Vorhaben, J.E. Avian Mitochondrial Glutamine Metabolism. J. Biol. Chem. 1976, 251, 781–786. [Google Scholar] [PubMed]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The Multifunctional Fish Gill: Dominant Site of Gas Exchange, Osmoregulation, Acid-Base Regulation, and Excretion of Nitrogenous Waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.D.; Wilkie, M.P.; Edwards, S.L. Rh Glycoprotein Immunoreactivity in the Skin and Its Role in Extrabranchial Ammonia Excretion by the Sea Lamprey (Petromyzon marinus) in Fresh Water. Can. J. Zool. 2017, 95, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Randall, D.J.; Wood, C.M.; Perry, S.F.; Bergman, H.; Maloiy, G.M.O.; Mommsen, T.P.; Wright, P.A. Urea Excretion as a Strategy for Survival in a Fish Living in a Very Alkaline Environment. Nature 1989, 337, 165–166. [Google Scholar] [CrossRef]
- Walsh, P.J.; Danulat, E.; Mommsen, T.P. Variation in Urea Excretion in the Gulf Toadfish Opsanus beta. Mar. Biol. 1990, 106, 323–328. [Google Scholar] [CrossRef]
- Walsh, P.J.; Milligan, C.L. Effects of Feeding and Confinement on Nitrogen Metabolism and Excretion in the Gulf Toadfish Opsanus beta. J. Exp. Biol. 1995, 198, 1559–1566. [Google Scholar]
- Davuluri, G.; Allawy, A.; Thapaliya, S.; Rennison, J.H.; Singh, D.; Kumar, A.; Sandlers, Y.; Van Wagoner, D.R.; Flask, C.A.; Hoppel, C.; et al. Hyperammonaemia-induced skeletal muscle mitochondrial dysfunction results in cataplerosis and oxidative stress. J. Physiol. 2016, 594, 7341–7360. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Davuluri, G.; Welch, N.; Kim, A.; Gangadhariah, M.; Allawy, A.; Priyadarshini, A.; McMullen, M.R.; Sandlers, Y.; Willard, B.; et al. Oxidative stress mediates ethanol-induced skeletal muscle mitochondrial dysfunction and dysregulated protein synthesis and autophagy. Free Radic. Biol. Med. 2019, 145, 284–299. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]
- Qiu, J.; Thapaliya, S.; Runkana, A.; Yang, Y.; Tsien, C.; Mohan, M.L.; Narayanan, A.; Eghtesad, B.; Mozdziak, P.E.; McDonald, C.; et al. Hyperammonemia in Cirrhosis Induces Transcriptional Regulation of Myostatin by an NF-ΚB-Mediated Mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 18162–18167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trendelenburg, A.; Meyer, A.; Rohner, D.; Boyle, J.; Hatakeyama, S.; Glass, D.J. Myostatin Reduces Akt/TORC1/P70S6K Signaling, Inhibiting Myoblast Differentiation and Myotube Size. Am. J. Physiol. Cell Physiol. 2009, 296, 1258–1270. [Google Scholar] [CrossRef] [Green Version]
- Dasarathy, S.; Hatzoglou, M. Hyperammoniemia and Proteostasis in Cirrhosis. Physiol. Behav. 2018, 21, 30–36. [Google Scholar] [CrossRef]
- Dasarathy, S. Consilience in Sarcopenia of Cirrhosis. J. Cachexia Sarcopenia Muscle 2012, 3, 225–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterworth, R.F. Pathophysiology of Brain Dysfunction in Hyperammonemic Syndromes: The Many Faces of Glutamine. Mol. Genet. Metab. 2014. [Google Scholar] [CrossRef]
- Takahashi, H.; Koehler, R.C.; Brusilow, S.W.; Traystman, R.J. Inhibition of Brain Glutamine Accumulation Prevents Cerebral Edema in Hyperammonemic Rats. Am. J. Physiol. Heart Circ. Physiol. 1991, 261, 825–829. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M. Tilapia Culture in Salt Water: Environmental Requirements, Nutritional Implications and Economic Potentials. Av. Nutr. Acuícola 2006, 95–106. [Google Scholar]
- Taylor, P.; Redner, B.D.; Stickney, R.R. Acclimation to Ammonia by Tilapia Aurea. Trans. Am. Fish. Soc. 1979, 108, 383–388. [Google Scholar] [CrossRef]
- Iwata, K.; Deguchi, M. Metabolic Fate and Distribution of 15 N-Ammonia in an Ammonotelic Amphibious Fish, Periophthalmus Modestus, Following Immersion in 15 N-Ammonium Sulfate: A Long Term Experiment. Zool. Sci. 1995, 12, 175–184. [Google Scholar] [CrossRef]
- Singh, R.A.; Singh, S.N. Tissue Glutamine Synthetase Associated with Ammonia Detoxification and Nitrogen Metabolism in Clarias Batrachus. Arch. Int. Physiol. 1989, 97, 145–152. [Google Scholar]
- Stern, R.A.; Mozdziak, P.E. Glutamine Synthetase in Avian Muscle Contributes to a Positive Myogenic Response to Ammonia Compared with Mammalian Muscle. Am. J. Physiol. Integr. Comp. Physiol. 2019, 317, R214–R221. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.D.; Campbell, J.W. Distribution of Glutamine Synthetase and Carbamoyl-Phosphate Synthetase I in Vertebrate Liver. Evolution 1988, 85, 160–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonetto, A.; Penna, F.; Minero, V.G.; Reffo, P.; Costamagna, D.; Bonelli, G.; Baccino, F.M.; Costelli, P. Glutamine Prevents Myostatin Hyperexpression and Protein Hypercatabolism Induced in C2C12 Myotubes by Tumor Necrosis Factor-α. Amino Acids 2011, 40, 585–594. [Google Scholar] [CrossRef]
- Oudelaar, A.M.; Beagrie, R.A.; Gosden, M.; de Ornellas, S.; Georgiades, E.; Kerry, J.; Hidalgo, D.; Carrelha, J.; Shivalingam, A.; El-Sagheer, A.H.; et al. Dynamics of the 4D genome during in vivo lineage specification and differentiation. Nat. Commun. 2020, 11, 2722. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
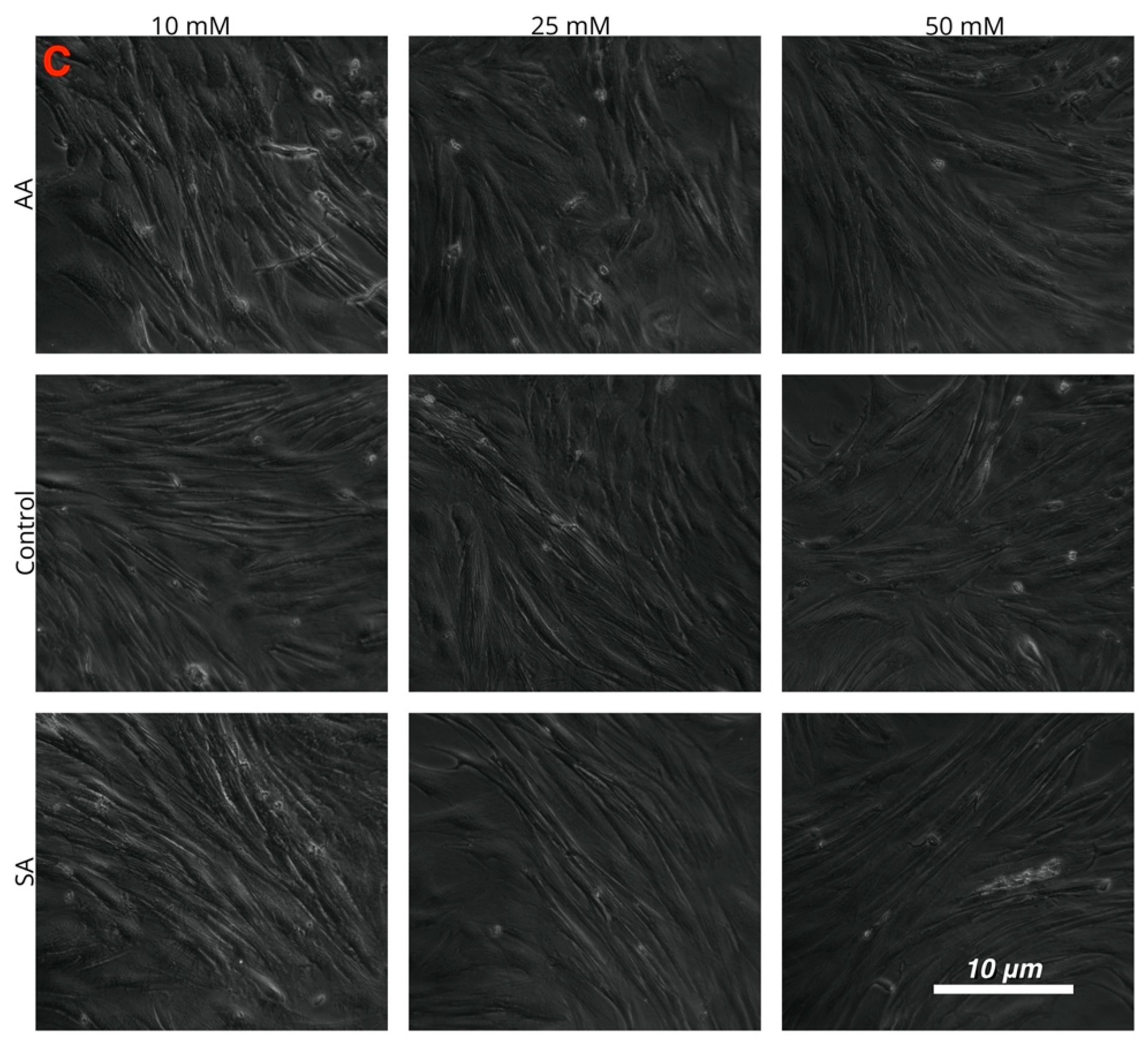{kind=link}
| Species | Primer 1 | Sequence | Bp Size |
|---|---|---|---|
| Tilapia | MSTN | F 5′-GCTGCGAATGAAGGAAGCTC-3′ R 5′-CGGTGGTCACTTCTTCCGAT-3′ | 317 |
| Tilapia | MyoD | F 5′-ACGGCATGACGGATTTTAACG-3′ R 5′-CTTGGTAAATCAGGTTGGGGTC-3′ | 315 |
| Tilapia | Myf5 | F 5′-AATGCAAACTACAGCAACGGC-3′ R 5′-GACAGGCGGTCCACGATACT-3′ | 110 |
| Tilapia | MYOG | F 5′-CAGCAGGGTTTGCTCTACCG-3′ R 5′-CTGAACTGGGCTCGCTTGAC-3′ | 102 |
| Tilapia | Myf6 | F 5′-CCCAAGCGGGTCACGATAAT-3′ R 5′-GCCTTACGTCTATCCGTGGG-3′ | 160 |
| Tilapia | PAX7 | F 5′-GACAGGCGGTCCACGATACT-3′ R 5′-TGCGCCTCTGCTTCCTTTTA-3′ | 194 |
| Tilapia | β-Actin | F 5′-TGGTGGGTATGGGTCAGAAAG-3′ R 5′-CTGTTGGCTTTGGGGTTCA-3′ | 217 |
| Murine | MSTN | F 5′-TCACGCTACCACGGAAACAA-3′ R 5′-AGGAGTCTTGACGGGTCTGA-3′ | 166 |
| Murine | MyoD | F 5′-GCTCTGATGGCATGATGGATT-3′ R 5′-CTATGCTGGACAGGCAGTCG-3′ | 150 |
| Murine | Myf5 | F 5′-AACTATTACAGCCTGCCGGG-3′ R 5′-GCTGGACAAGCAATCCAAGC-3′ | 198 |
| Murine | MYOG | F 5′-GTGCCCAGTGAATGCAACTC-3′ R 5′-CGAGCAAATGATCTCCTGGGT-3′ | 94 |
| Murine | Myf6 | F 5′-AGAAATTCTTGAGGGTGCGG-3′ R 5′-GCCCCTGGAATGATCCGAAA-3′ | 76 |
| Murine | PAX7 | F 5′-AGTTCGATTAGCCGAGTGCT-3′ R 5′-CATCCAGACGGTTCCCTTTGT-3′ | 142 |
| Murine | β-Actin | F 5′-AGATCAAGATCATTGCTCCTCC-3′ R 5′-AGCTCAGTAACAGTCCGCCTA-3′ | 170 |
| Avian | MSTN | F 5′-CGGAGAATGCGAATTTGTGTTTC-3′ R 5′-GGGACATCTTGGTGGGTGTG-3′ | 110 |
| Avian | MyoD | F 5′-CGCAGGAGAAACAGCTACGA-3′ R 5′-ATGCTTGAGAGGCAGTCGAG-3′ | 104 |
| Avian | Myf5 | F 5′-TGAGGGAACAGGTGGAGAACT-3′ R 5′-ACTCTGCTCCGTCGCGTA-3′ | 185 |
| Avian | MYOG | F 5′-CAGCCTCAACCAGCAGGAG-3′ R 5′-ACTGCTCAGGAGGTGATCTG-3′ | 166 |
| Avian | Myf6 | F 5′-AGGCTGGATCAGCAGGACAAAA-3′ R 5′-CGCGGGAATGGTCGGAAG-3′ | 139 |
| Avian | PAX7 | F 5′-GAAGGCCTTTGAGAGGACCC-3′ R 5′-GGTTGAATGCTGCGAGTTGG-3′ | 158 |
| Avian | β-Actin | F 5′-GTCCACCTTCCAGCAGATGT-3′ R 5′-TAAAGCCATGCCAATCTCG-3′ | 168 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miramontes, E.; Kempisty, B.; Petitte, J.; Dasarathy, S.; Kulus, M.; Wieczorkiewicz, M.; Mozdziak, P. Myogenic Response to Increasing Concentrations of Ammonia Differs between Mammalian, Avian, and Fish Species: Cell Differentiation and Genetic Study. Genes 2020, 11, 840. https://doi.org/10.3390/genes11080840
Miramontes E, Kempisty B, Petitte J, Dasarathy S, Kulus M, Wieczorkiewicz M, Mozdziak P. Myogenic Response to Increasing Concentrations of Ammonia Differs between Mammalian, Avian, and Fish Species: Cell Differentiation and Genetic Study. Genes. 2020; 11(8):840. https://doi.org/10.3390/genes11080840
Chicago/Turabian StyleMiramontes, Emily, Bartosz Kempisty, James Petitte, Srinivasan Dasarathy, Magdalena Kulus, Maria Wieczorkiewicz, and Paul Mozdziak. 2020. "Myogenic Response to Increasing Concentrations of Ammonia Differs between Mammalian, Avian, and Fish Species: Cell Differentiation and Genetic Study" Genes 11, no. 8: 840. https://doi.org/10.3390/genes11080840