Atypical Genotypes for Canine Agouti Signaling Protein Suggest Novel Chromosomal Rearrangement

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Datasets
2.2. Sample Collection and Genotyping
3. Results
3.1. Detected Frequencies of A3+
3.2. Phenotypic Impact of A3+
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dreger, D.L.; Hooser, B.N.; Hughes, A.M.; Ganesan, B.; Donner, J.; Anderson, H.; Holtvoigt, L.; Ekenstedt, K.J. True Colors: Commercially-acquired morphological genotypes reveal hidden allele variation among dog breeds, informing both trait ancestry and breed potential. PLoS ONE 2019, 14, e0223995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berryere, T.G.; Kerns, J.A.; Barsh, G.S.; Schmutz, S.M. Association of an Agouti allele with fawn or sable coat color in domestic dogs. Mamm. Genome 2005, 16, 262–272. [Google Scholar] [CrossRef]
- Dreger, D.L.; Schmutz, S.M. A SINE insertion causes the black-and-tan and saddle tan phenotypes in domestic dogs. J. Hered. 2011, 102, S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Kerns, J.A.; Newton, J.; Berryere, T.G.; Rubin, E.M.; Cheng, J.F.; Schmutz, S.M.; Barsh, G.S. Characterization of the dog Agouti gene and a nonagouti mutation in German Shepherd Dogs. Mamm. Genome 2004, 15, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Donner, J.; Kaukonen, M.; Anderson, H.; Möller, F.; Kyöstilä, K.; Sankari, S.; Hytönen, M.; Giger, U.; Lohi, H. Genetic panel screening of nearly 100 mutations reveals new insights into the breed distribution of risk variants for canine hereditary disorders. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef]
- Donner, J.; Anderson, H.; Davison, S.; Hughes, A.M.; Bouirmane, J.; Lindqvist, J.; Lytle, K.M.; Ganesan, B.; Kaukonen, M.; Forman, O.P.; et al. Frequency and distribution of 152 genetic disease variants in over 100,000 mixed breed and purebred dogs. PLoS Genet. 2018, 14, 1–20. [Google Scholar] [CrossRef]
- Everts, R.E.; Rothuizen, J.; Van Oost, B.A. Identification of a premature stop codon in the melanocyte-stimulating hormone receptor gene (MC1R) in Labrador and Golden retrievers with yellow coat colour. Anim. Genet. 2000, 31, 194–199. [Google Scholar] [CrossRef]
- Newton, J.M.; Wilkie, A.L.; He, L.; Jordan, S.A.; Metallinos, D.L.; Holmes, N.G.; Jackson, I.J.; Barsh, G.S. Melanocortin 1 receptor variation in the domestic dog. Mamm. Genome 2000, 11, 24–30. [Google Scholar] [CrossRef]
- Kerns, J.A.; Cargill, E.J.; Clark, L.A.; Candille, S.I.; Berryere, T.G.; Olivier, M.; Lust, G.; Todhunter, R.J.; Schmutz, S.M.; Murphy, K.E.; et al. Linkage and segregation analysis of black and brindle coat color in domestic dogs. Genetics 2007, 176, 1679–1689. [Google Scholar] [CrossRef] [Green Version]
- Candille, S.I.; Kaelin, C.B.; Cattanach, B.M.; Yu, B.; Thompson, D.A.; Nix, M.A.; Kerns, J.A.; Schmutz, S.M.; Millhauser, G.L.; Barsh, G.S. A β-defensin mutation causes black coat color in domestic dogs. Science 2007, 318, 1418–1423. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.G.; Dreger, D.L.; Rimbault, M.; Davis, B.W.; Mullen, A.B.; Carpintero-Ramirez, G.; Ostrander, E.A. Genomic analyses reveal the influence of geographic origin, migration, and hybridization on modern dog breed development. Cell Rep. 2017, 19, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Talenti, A.; Dreger, D.L.; Frattini, S.; Polli, M.; Marelli, S.; Harris, A.C.; Liotta, L.; Cocco, R.; Hogan, A.N.; Bigi, D.; et al. Studies of modern Italian dog populations reveal multiple patterns for domestic breed evolution. Ecol. Evol. 2017, 8, 2911–2925. [Google Scholar] [CrossRef] [PubMed]
- Dumont, B.L.; Payseur, B.A. Evolution of the genomic rate of recombination in mammals. Evolution 2008, 62, 276–294. [Google Scholar] [CrossRef] [PubMed]
- Han, J.L.; Yang, M.; Yue, Y.J.; Guo, T.T.; Liu, J.B.; Niu, C.E.; Yang, B.H. Analysis of agouti signaling protein (ASIP) gene polymorphisms and association with coat color in Tibetan sheep (Ovis aries). Genet. Mol. Res. 2015, 14, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Rochus, C.M.; Westberg Sunesson, K.; Jonas, E.; Mikko, S.; Johansson, A.M. Mutations in ASIP and MC1R: Dominant black and recessive black alleles segregate in native Swedish sheep populations. Anim. Genet. 2019, 50, 712–717. [Google Scholar] [CrossRef]
- Henkel, J.; Saif, R.; Jagannathan, V.; Schmocker, C.; Zeindler, F.; Bangerter, E.; Herren, U.; Posantzis, D.; Bulut, Z.; Ammann, P.; et al. Selection signatures in goats reveal copy number variants underlying breed-defining coat color phenotypes. PLoS Genet. 2019, 15, e1008536. [Google Scholar] [CrossRef]
- Liu, M.; Zhou, Y.; Rosen, B.D.; Van Tassell, C.P.; Stella, A.; Tosser-Klopp, G.; Rupp, R.; Paljiere, I.; Colli, L.; Sayre, B.; et al. Diversity of copy number variation in the worldwide goat population. Heredity 2019, 122, 636–646. [Google Scholar] [CrossRef]
- Fontanesi, L.; Beretti, F.; Riggio, V.; Gomez Gonzalez, E.; Dall’Olio, S.; Davoli, R.; Russo, V.; Portolano, B. Copy number variation and missense mutations of the agouti signaling protein (ASIP) gene in goat breeds with different coat colors. Cytogenet. Genome Res. 2009, 126, 333–347. [Google Scholar] [CrossRef]
- Norris, B.J.; Whan, V.A. A gene duplication affecting expression of the ovine ASIP gene is responsible for white and black sheep. Genome Res. 2008, 18, 1282–1293. [Google Scholar] [CrossRef] [Green Version]
- Ciampolini, R.; Cecchi, F.; Spaterna, A.; Bramante, A.; Bardet, S.M.; Oulmouden, A. Characterization of different 5′-untranslated exons of the ASIP gene in black-and-tan Doberman Pinscher and brindle Boxer dogs. Anim. Genet. 2013, 44, 114–117. [Google Scholar] [CrossRef]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guerin, G. Mutations in the agouti (ASIP), the extension (MC1R), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Eizirik, E.; Yuhki, N.; Johnson, W.E.; Menotti-Raymond, M.; Hannah, S.S.; O’Brien, S.J. Molecular genetics and evolution of melanism in the cat family. Curr. Biol. 2003, 13, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Hiragaki, T.; Inoue-Murayama, M.; Miwa, M.; Fujiwara, A.; Mizutani, M.; Minvielle, F.; Ito, S. Recessive black is allelic to the yellow plumage locus in Japanese quail and associated with a frameshift deletion in the ASIP gene. Genetics 2008, 178, 771–775. [Google Scholar] [CrossRef] [Green Version]
- Chandramohan, B.; Renieri, C.; La Manna, V.; La Terza, A. The alpaca agouti gene: Genomic locus, transcripts and causative mutations of eumelanic and phaeomelanic coat color. Gene 2013, 521, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Oey, H.; Isbel, L.; Hickey, P.; Ebaid, B.; Whitelaw, E. Genetic and epigenetic variation among inbred mouse littermates: Identification of inter-individual differentially methylated regions. Epigenetics Chromatin 2015, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Waterland, R.A.; Jirtle, R.L. Transposable elements: Targets for early ntritional effects on epigenetic gene regulation. Mol. Cell Biol. 2003, 23, 5293–5300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blewitt, M.E.; Vickaryous, N.K.; Paldi, A.; Koseki, H.; Whitelaw, E. Dynamic reprogramming of DNA methylation at an epigenetically sensitive allele in mice. PLoS Genet 2006, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ekram, M.B.; Kang, K.; Kim, H.; Kim, J. Retrotransposons as a major source of epigenetic variations in the mammalian genome. Epigenetics 2012, 7, 370–382. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
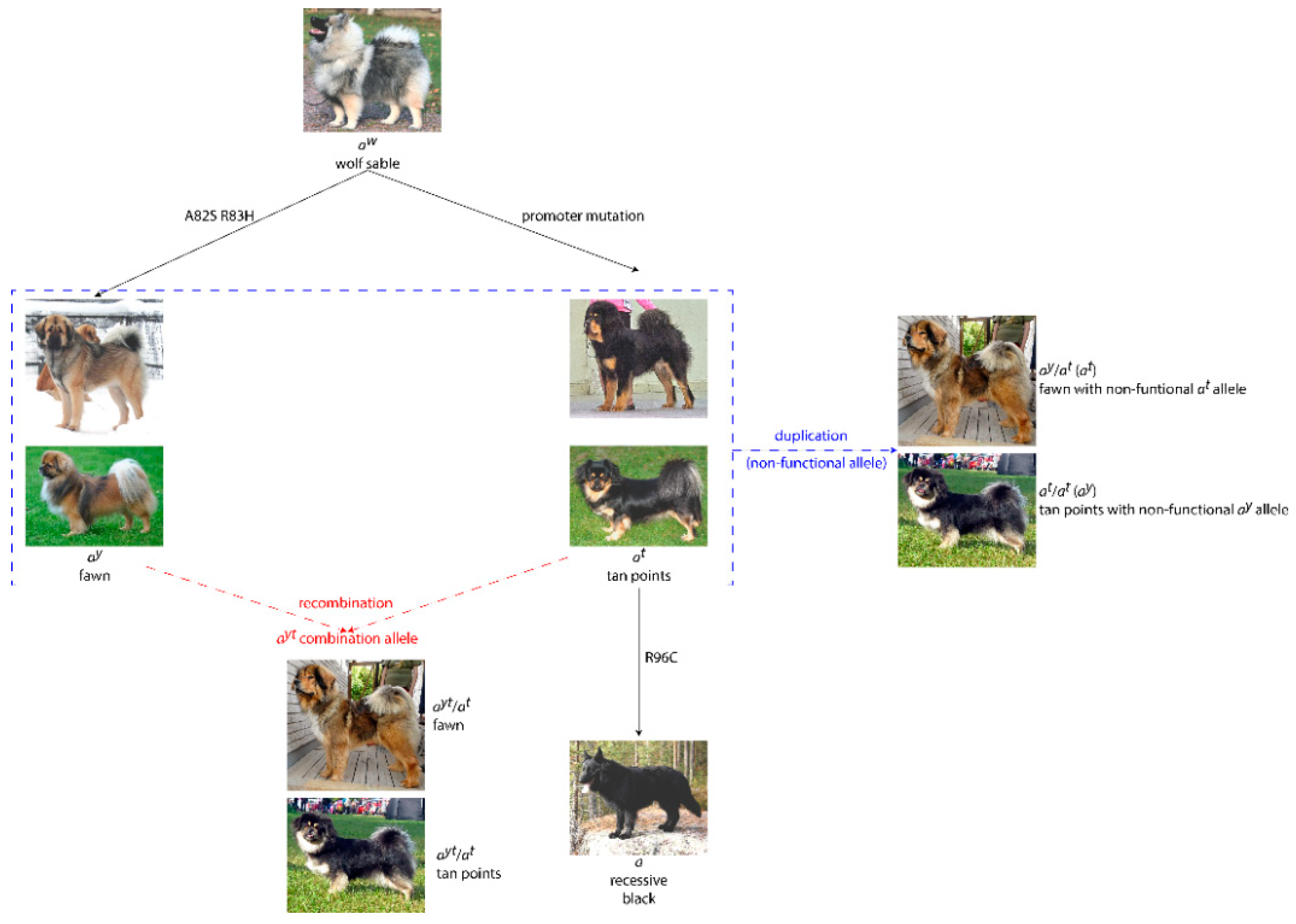{kind=link}
| Breed | WHTC | WHAE | MLAS | |||||
|---|---|---|---|---|---|---|---|---|
| N | A3+ | % | n | A3+ | % | n | A3+ | |
| Anatolian Shepherd | 22 | 1 | 4.55% | 18 | 1 | 5.56% | - | - |
| Basenji | - | - | - | 74 | 1 | 1.35% | - | - |
| Berger Picard | 9 | 1 | 11.11% | 7 | 0 | 0.00% | - | - |
| Boston Terrier | 64 | 4 | 6.25% | 94 | 0 | 0.00% | 3 | 3 |
| Brussels Griffon | 44 | 1 | 2.27% | 44 | 0 | 0.00% | - | - |
| Dingo | 12 | 2 | 16.67% | - | - | - | - | - |
| Dogo Argentino | 9 | 6 | 66.67% | 11 | 7 | 63.64% | - | - |
| East-Siberian Laika | - | - | - | 109 | 15 | 13.76% | - | - |
| Great Dane | 79 | 1 | 1.27% | 163 | 0 | 0.00% | - | - |
| Great Pyrenees | 51 | 8 | 15.69% | 70 | 9 | 12.86% | - | - |
| Irish Terrier | 33 | 1 | 3.03% | 55 | 0 | 0.00% | - | - |
| Kai Ken | - | - | - | 19 | 1 | 5.26% | - | - |
| Lagotto Romagnolo | 116 | 8 | 6.90% | 1840 | 80 | 4.35% | - | - |
| Maremma Sheepdog | 18 | 3 | 16.67% | 24 | 5 | 20.83% | - | - |
| Spanish Greyhound | 25 | 1 | 4.00% | 72 | 1 | 1.39% | - | - |
| Tibetan Mastiff | 83 | 6 | 7.23% | 108 | 14 | 12.96% | 1 | 1 |
| Tibetan Spaniel | 46 | 1 | 2.17% | 177 | 20 | 11.30% | 115 | 15 |
| Whippet | 68 | 1 | 1.47% | 304 | 4 | 1.32% | - | - |
| Total | 679 | 45 | 3189 | 158 | 119 | 19 | ||
| No. of Non-Wild-Type Variants a | ay [2] | at [3] | a [4] | Interpreted Genotype b | No. of Observations (Out of 3189 Dogs) | Proportion of Genotypes |
|---|---|---|---|---|---|---|
| p.A82S | g.23365298ins239 | p.R96C | ||||
| 2 | 2/2 | 1/1 | 1/1 | ay/ay | 909 | 28.50% |
| 1 | 1/2 | 1/1 | 1/1 | ay/aw | 29 | 0.91% |
| 2 | 1/2 | 1/2 | 1/1 | ay/at | 376 | 11.79% |
| 2 | 1/2 | 1/2 | 1/2 | ay/a | 44 | 1.38% |
| 0 | 1/1 | 1/1 | 1/1 | aw/aw | 13 | 0.41% |
| 1 | 1/1 | 1/2 | 1/1 | aw/at | 62 | 1.94% |
| 1 | 1/1 | 1/2 | 1/2 | aw/a | 0 | 0.00% |
| 2 | 1/1 | 2/2 | 1/1 | at/at | 1303 | 40.86% |
| 2 | 1/1 | 2/2 | 1/2 | at/a | 265 | 8.31% |
| 2 | 1/1 | 2/2 | 2/2 | a/a | 30 | 0.94% |
| 3 | 2/2 | 1/2 | 1/1 | ay/ay/at | 41 | 1.29% |
| 3 | 1/2 | 2/2 | 1/1 | ay/at/at | 92 | 2.88% |
| 3 | 1/2 | 2/2 | 1/2 | ay/at/a | 18 | 0.56% |
| 4 | 2/2 | 2/2 | 1/1 | ay/ay/at/at | 7 | 0.22% |
| Alleles | Breed | n | Color | Phenotype Allele |
|---|---|---|---|---|
| ay/ay/at | Boston Terrier | 3 | fawn (with brindle) | ay |
| East-Siberian Laika | 1 | fawn | ay | |
| Great Pyrenees | 1 | fawn (with white) | ay | |
| Tibetan Mastiff | 1 | fawn | ay | |
| Tibetan Spaniel | 20 | fawn | ay | |
| Whippet | 3 | fawn (with brindle) | ay | |
| ay/at/at | East-Siberian Laika | 1 | wolf sable | aw |
| 2 | dark fawn or wolf sable | ay or aw | ||
| 2 | fawn | ay | ||
| 1 | tan points | at | ||
| Tibetan Mastiff | 3 | fawn | ay | |
| Tibetan Spaniel | 12 | tan points | at | |
| ay/at/a | East-Siberian Laika | 2 | wolf sable | aw |
| ay/ay/at/at | East-Siberian Laika | 1 | wolf sable | aw |
| Whippet | 1 | fawn (with brindle) | ay |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dreger, D.L.; Anderson, H.; Donner, J.; Clark, J.A.; Dykstra, A.; Hughes, A.M.; Ekenstedt, K.J. Atypical Genotypes for Canine Agouti Signaling Protein Suggest Novel Chromosomal Rearrangement. Genes 2020, 11, 739. https://doi.org/10.3390/genes11070739
Dreger DL, Anderson H, Donner J, Clark JA, Dykstra A, Hughes AM, Ekenstedt KJ. Atypical Genotypes for Canine Agouti Signaling Protein Suggest Novel Chromosomal Rearrangement. Genes. 2020; 11(7):739. https://doi.org/10.3390/genes11070739
Chicago/Turabian StyleDreger, Dayna L., Heidi Anderson, Jonas Donner, Jessica A. Clark, Arlene Dykstra, Angela M. Hughes, and Kari J. Ekenstedt. 2020. "Atypical Genotypes for Canine Agouti Signaling Protein Suggest Novel Chromosomal Rearrangement" Genes 11, no. 7: 739. https://doi.org/10.3390/genes11070739