Telomere and Centromere Staining Followed by M-FISH Improves Diagnosis of Chromosomal Instability and Its Clinical Utility

 , ,
, , 
 , ,
, ,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Preparation of Metaphase Spreads
2.2.2. TC+M-FISH
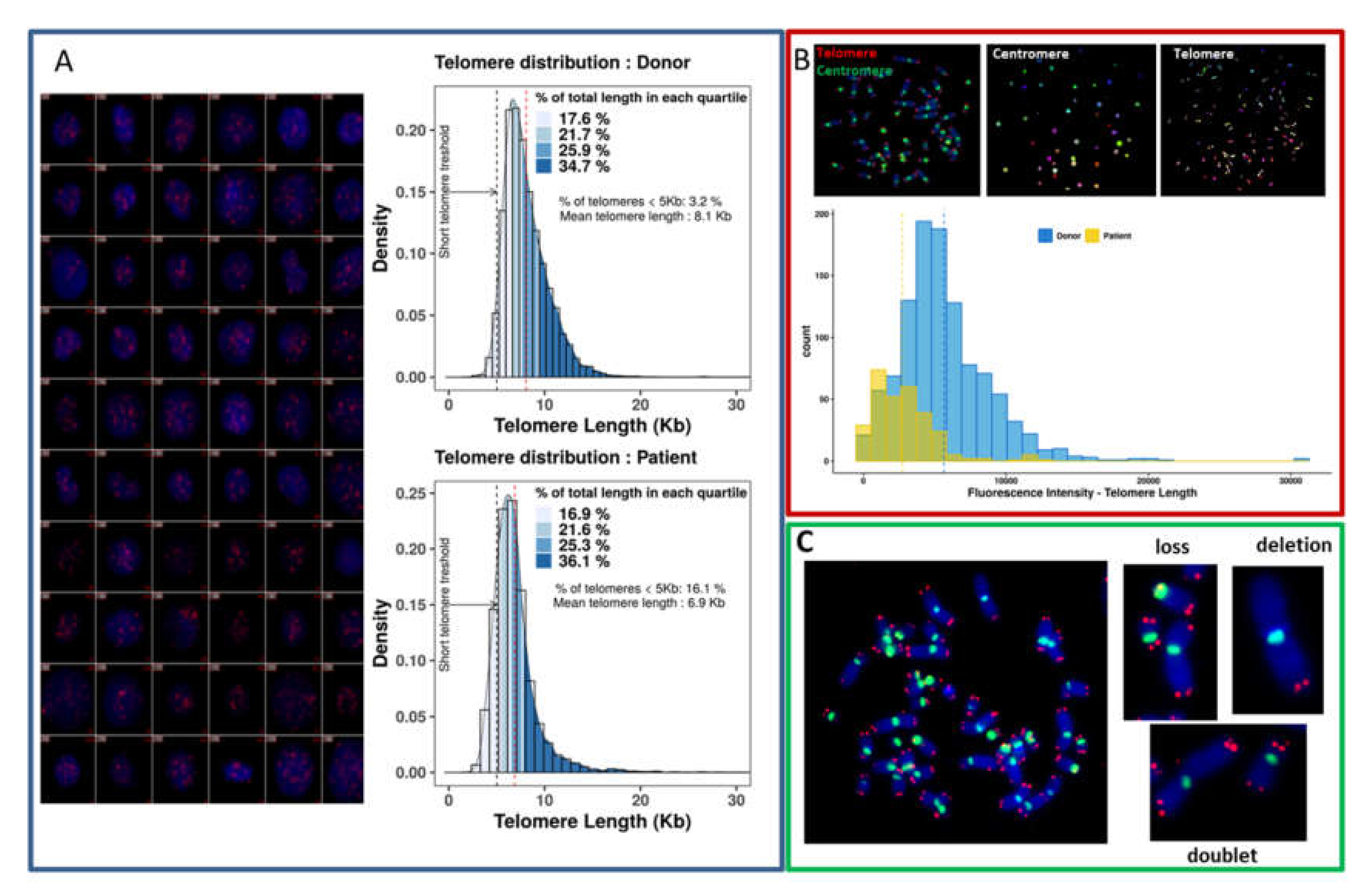
2.2.3. Telomere Quantification
2.2.4. Scoring of Telomeres and Chromosomal Aberrations
2.2.5. Statistical Analysis
3. Results
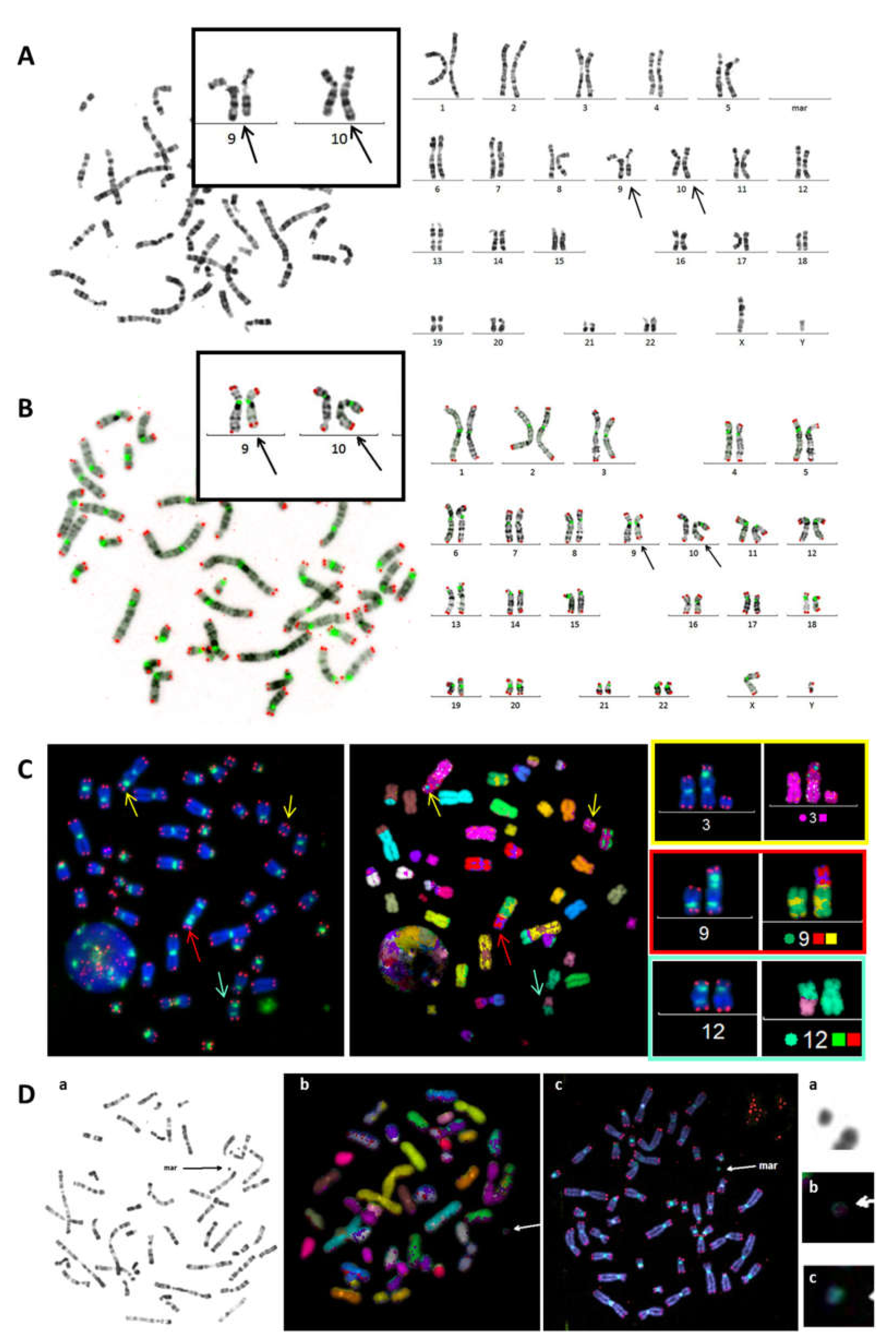
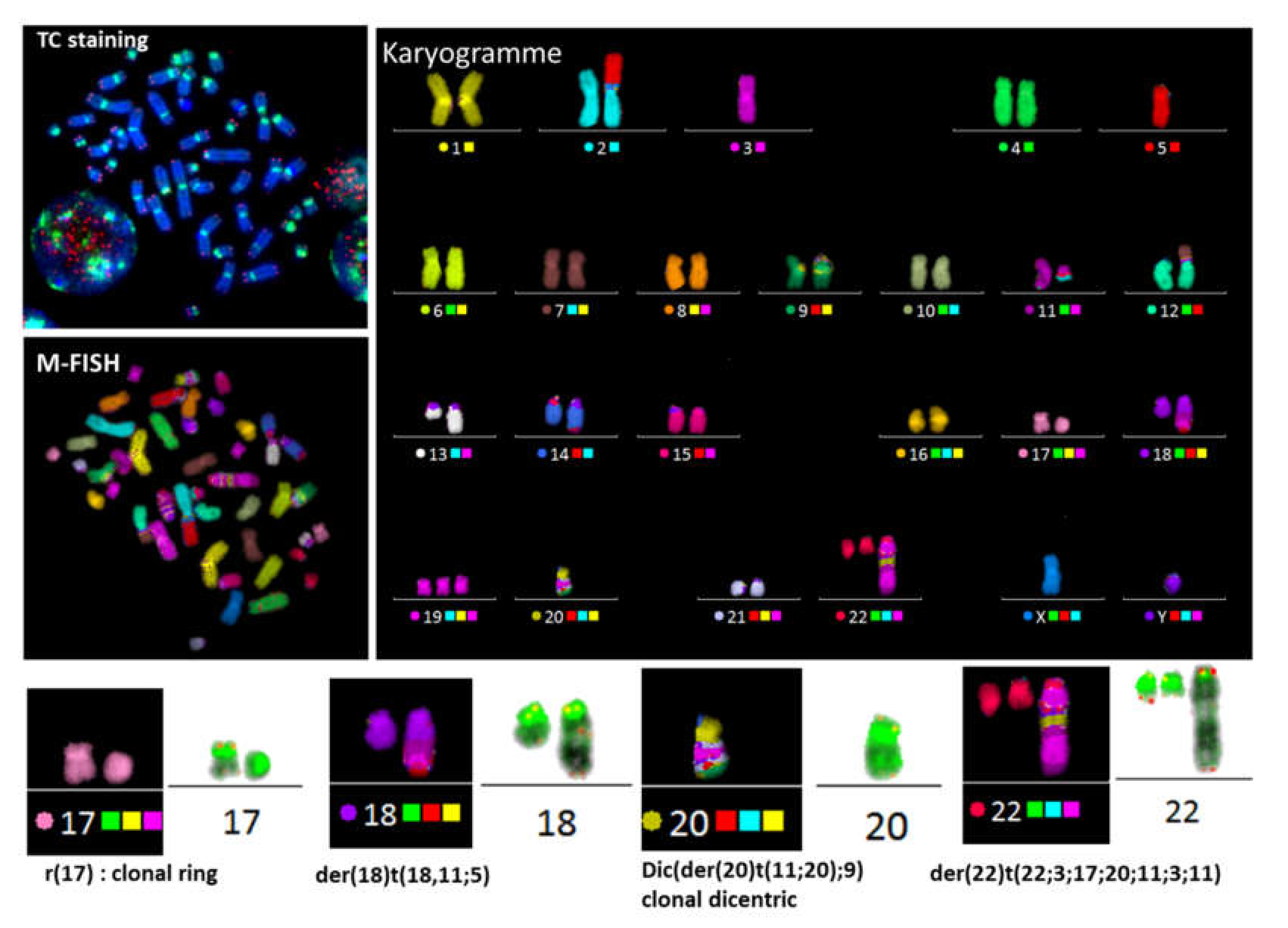
3.1. Improved Identification of Chromosomal Aberrations after TC Staining
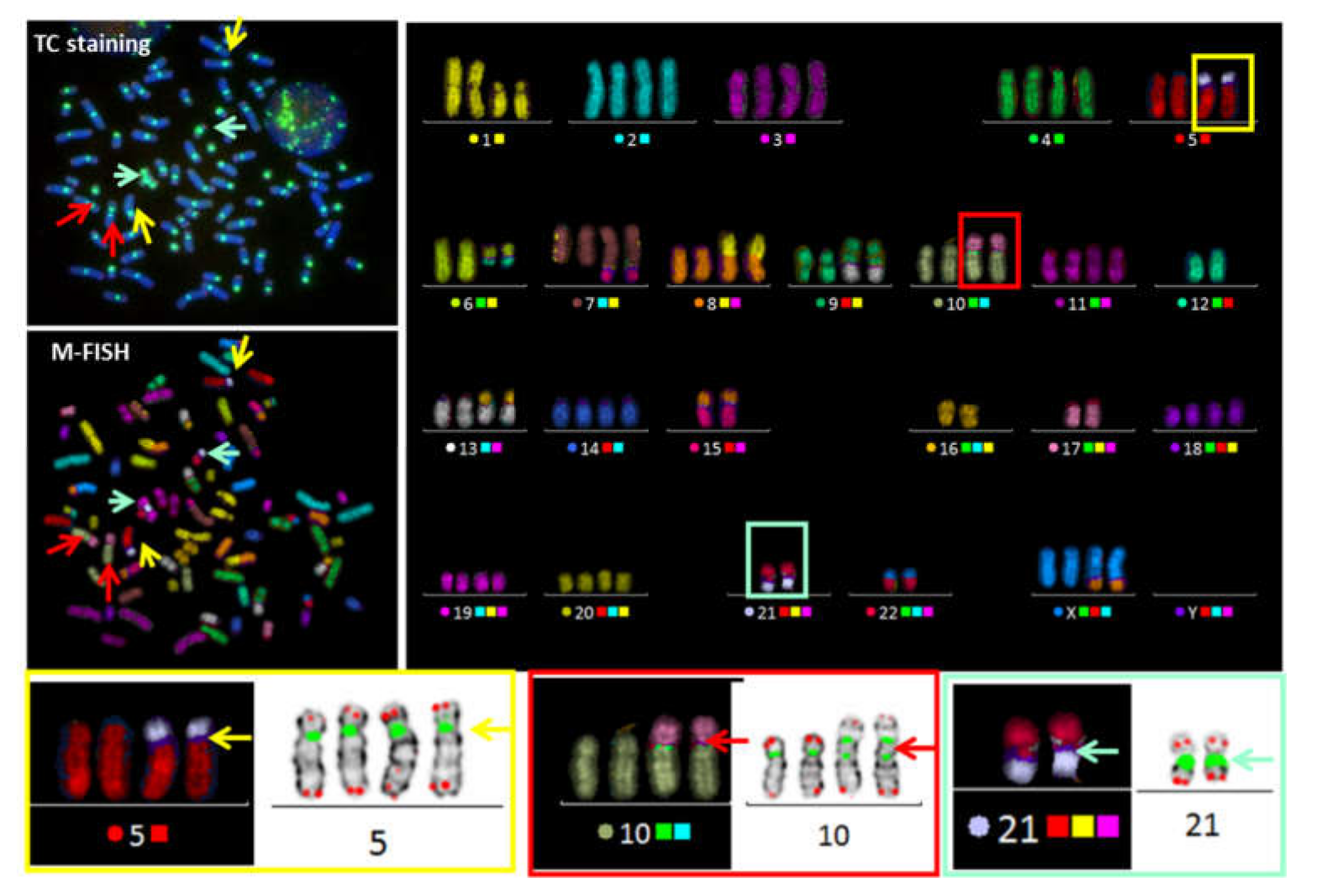
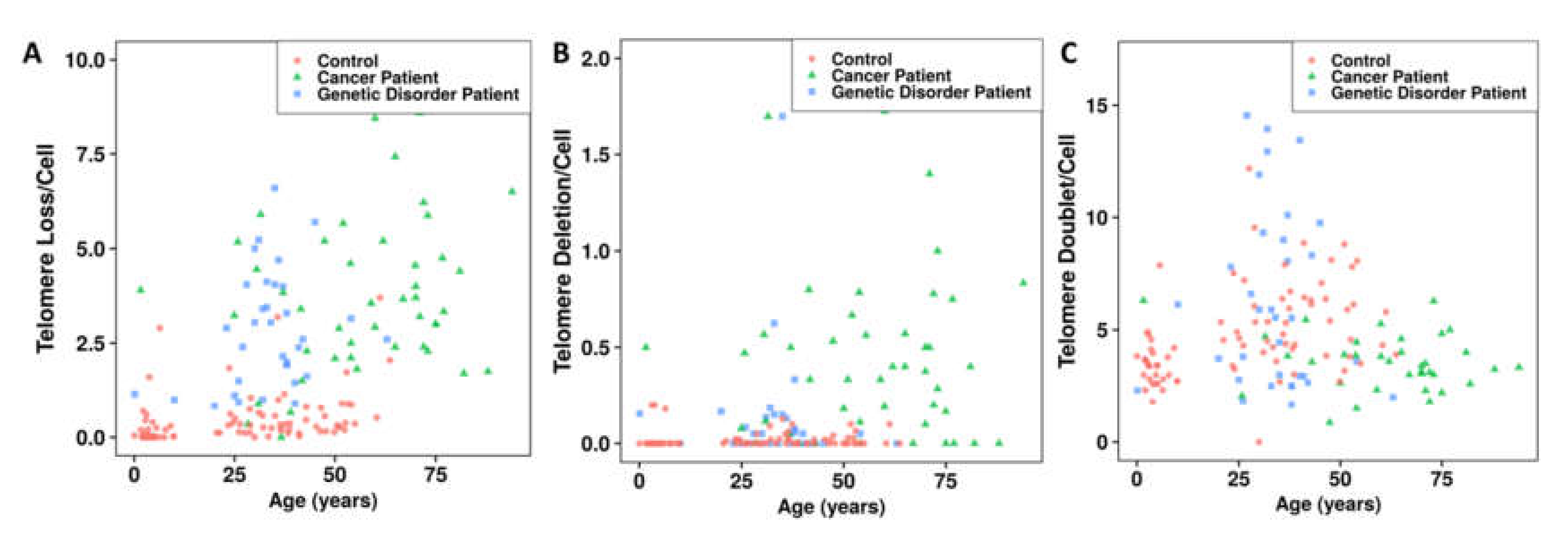
3.2. Telomere Instability Detected by TC+M-FISH Staining
3.3. Validation of the Concept
3.3.1. Telomere Dysfunction
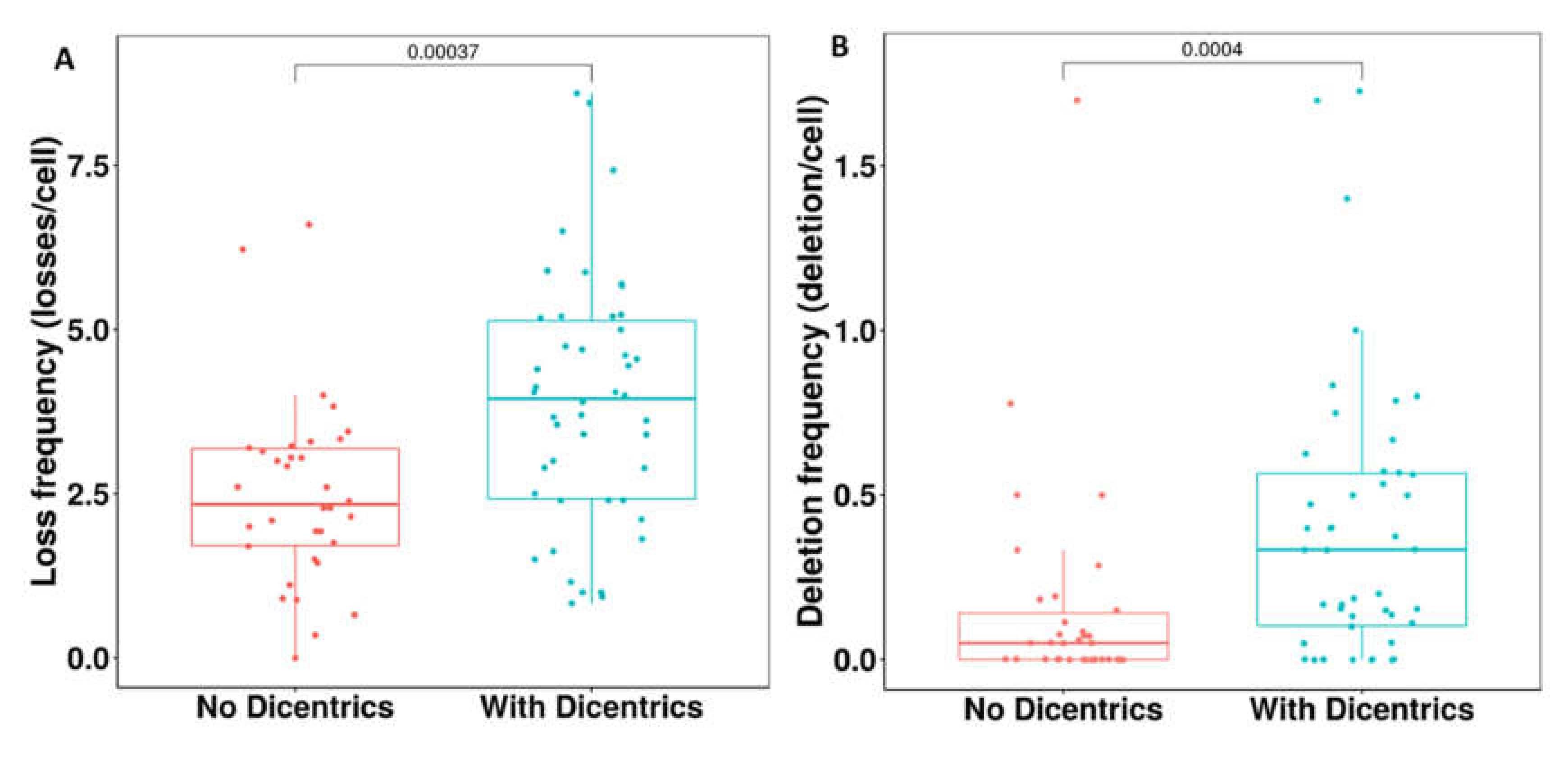
3.3.2. Detection of Dicentric Chromosomes
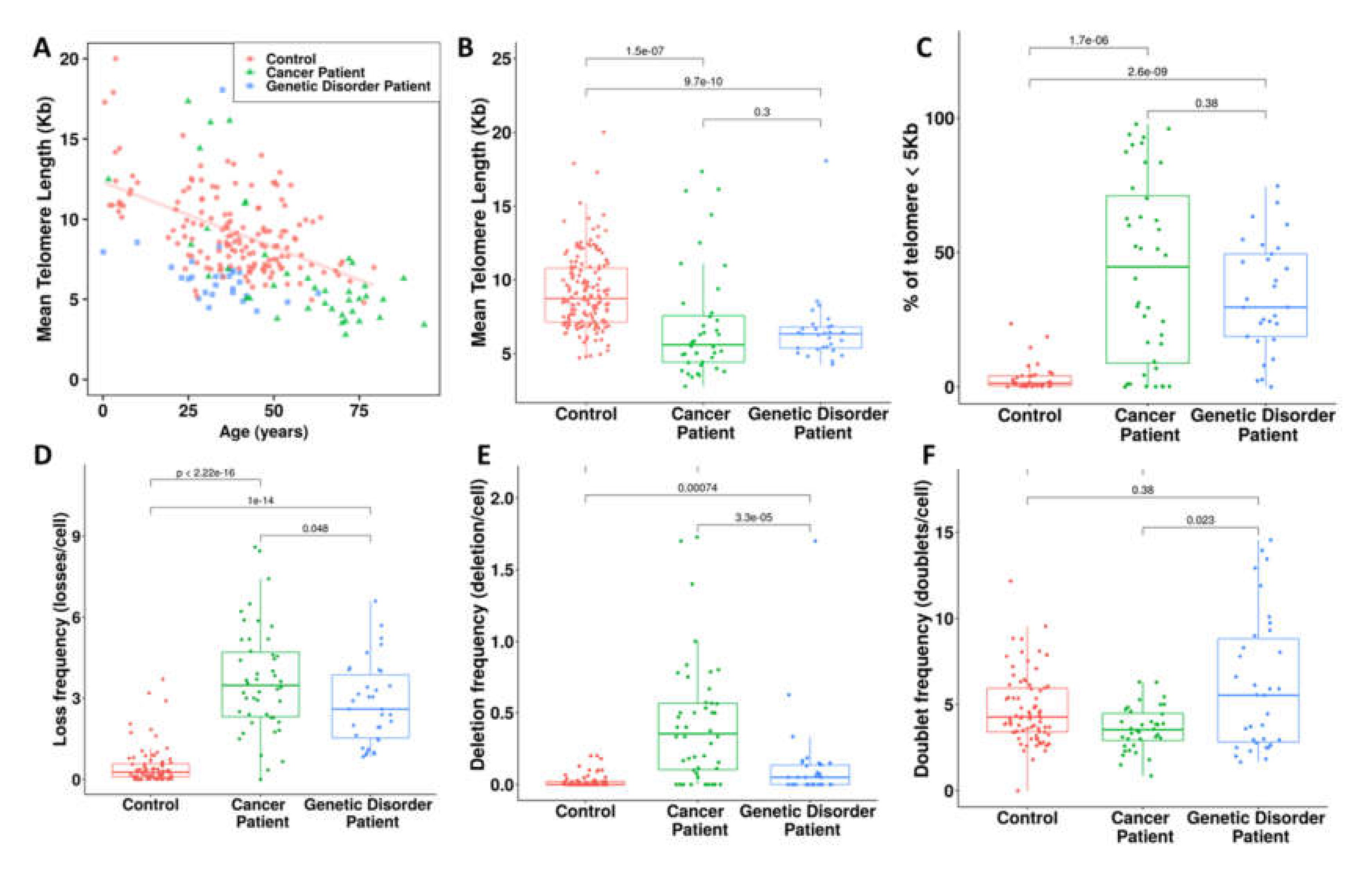
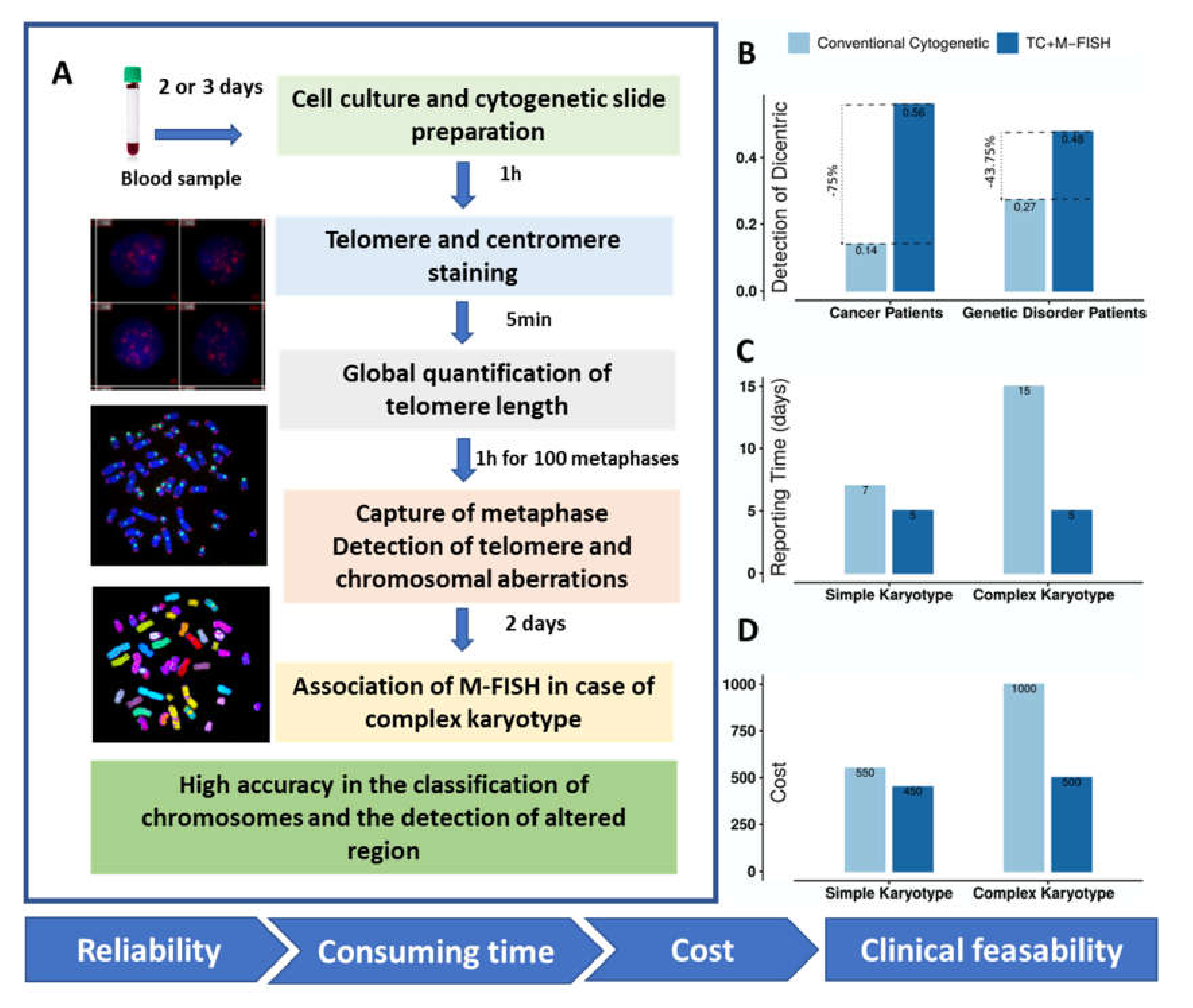
3.4. Proof of Concept in the Clinics
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Burrell, R.A.; McGranahan, N.; Bartek, J.; Swanton, C. The causes and consequences of genetic heterogeneity in cancer evolution. Nature 2013, 501, 338–345. [Google Scholar] [PubMed]
- Bakhoum, S.F.; Landau, D.A. Chromosomal Instability as a Driver of Tumor Heterogeneity and Evolution. Cold Spring Harb. Perspect. Med. 2017, 7, a029611. [Google Scholar] [PubMed]
- Thompson, S.L.; Compton, D.A. Chromosome missegregation in human cells arises through specific types of kinetochore-microtubule attachment errors. Proc. Natl. Acad. Sci. USA 2011, 108, 17974–17978. [Google Scholar] [PubMed] [Green Version]
- Pernot, E.; Hall, J.; Baatout, S.; Benotmane, M.A.; Blanchardon, E.; Bouffler, S.; El Saghire, H.; Gomolka, M.; Guertler, A.; Harms-Ringdahl, M.; et al. Ionizing radiation biomarkers for potential use in epidemiological studies. Mutat. Res. 2012, 751, 258–286. [Google Scholar]
- McGranahan, N.; Burrell, R.A.; Endesfelder, D.; Novelli, M.R.; Swanton, C. Cancer chromosomal instability: Therapeutic and diagnostic challenges. EMBO Rep. 2012, 13, 528–538. [Google Scholar]
- Bashashati, A.; Ha, G.; Tone, A.; Ding, J.; Prentice, L.M.; Roth, A.; Rosner, J.; Shumansky, K.; Kalloger, S.; Senz., J.; et al. Distinct evolutionary trajectories of primary high-grade serous ovarian cancers revealed through spatial mutational profiling. J. Pathol. 2013, 231, 21–34. [Google Scholar]
- Castellarin, M.; Milne, K.; Zeng, T.; Tse, K.; Mayo, M.; Zhao, Y.; Webb, J.R.; Watson, P.H.; Nelson, B.H.; Holt, R.A. Clonal evolution of high-grade serous ovarian carcinoma from primary to recurrent disease. J. Pathol. 2013, 229, 515–524. [Google Scholar]
- Jamal-Hanjani, M.; Wilson, G.A.; McGranahan, N.; Birkbak, N.J.; Watkins, T.B.K.; Veeriah, S.; Shafi, S.; Johnson, D.H.; Mitter, R.; Rosenthal, R.; et al. Tracking the Evolution of Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 376, 2109–2121. [Google Scholar]
- Murugaesu, N.; Wilson, G.A.; Birkbak, N.J.; Watkins, T.; McGranahan, N.; Kumar, S.; Abbassi-Ghadi, N.; Salm, M.; Mitter, R.; Horswell, S.; et al. Tracking the genomic evolution of esophageal adenocarcinoma through neoadjuvant chemotherapy. Cancer Discov. 2015, 5, 821–831. [Google Scholar]
- Benhra, N.; Barrio, L.; Muzzopappa, M.; Milan, M. Chromosomal Instability Induces Cellular Invasion in Epithelial Tissues. Dev. Cell 2018, 47, 161–174. [Google Scholar]
- Bakker, B.; van den Bos, H.; Lansdorp, P.M.; Foijer, F. How to count chromosomes in a cell: An overview of current and novel technologies. BioEssays News Rev. Mol. Cell. Dev. Biol. 2015, 37, 570–577. [Google Scholar]
- M’Kacher, R.; Girinsky, T.; Koscielny, S.; Dossou, J.; Violot, D.; Beron-Gaillard, N.; Ribrag, V.; Bourhis, J.; Bernheim, A.; Parmentier, C.; et al. Baseline and treatment-induced chromosomal abnormalities in peripheral blood lymphocytes of Hodgkin’s lymphoma patients. Int. J. Radiat. Oncol. Biol. Phys. 2003, 57, 321–326. [Google Scholar]
- M’Kacher, R.; Farace, F.; Bennaceur-Griscelli, A.; Violot, D.; Clausse, B.; Dossou, J.; Valent, A.; Parmentier, C.; Ribrag, V.; Bosq, J.; et al. Blastoid mantle cell lymphoma: Evidence for nonrandom cytogenetic abnormalities additional to t(11;14) and generation of a mouse model. Cancer Genet. Cytogenet. 2003, 143, 32–38. [Google Scholar] [PubMed]
- Cuceu, C.; Colicchio, B.; Jeandidier, E.; Junker, S.; Plassa, F.; Shim, G.; Mika, J.; Frenzel, M.; Al Jawhari, M.; Hempel, W.M.; et al. Independent Mechanisms Lead to Genomic Instability in Hodgkin Lymphoma: Microsatellite or Chromosomal Instability. Cancers 2018, 10, 233. [Google Scholar]
- Chin, T.F.; Ibrahim, K.; Thirunavakarasu, T.; Azanan, M.S.; Oh, L.; Lum, S.H.; Yap, T.Y.; Ariffin, H. Nonclonal Chromosomal Aberrations in Childhood Leukemia Survivors. Fetal Pediatric Pathol. 2018, 37, 243–253. [Google Scholar]
- Cantu, E.S.; Dong, H.; Forsyth, D.R.; Espinoza, F.P.; Papenhausen, P.R. Discrepant Cytogenetic and Interphase Fluorescence In Situ Hybridization (I-FISH) Results from Bone Marrow Specimens of Patients with Hematologic Neoplasms. Ann. Clin. Lab. Sci. 2018, 48, 264–272. [Google Scholar]
- Robinson, D.R.; Wu, Y.M.; Lonigro, R.J.; Vats, P.; Cobain, E.; Everett, J.; Cao, X.; Rabban, E.; Kumar-Sinha, C.; Raymond, V.; et al. Integrative clinical genomics of metastatic cancer. Nature 2017, 548, 297–303. [Google Scholar]
- Saha, A.K.; Mourad, M.; Kaplan, M.H.; Chefetz, I.; Malek, S.N.; Buckanovich, R.; Markovitz, D.M.; Contreras-Galindo, R. The Genomic Landscape of Centromeres in Cancers. Sci. Rep. 2019, 9, 11259. [Google Scholar]
- Martinez, A.C.; van Wely, K.H. Centromere fission, not telomere erosion, triggers chromosomal instability in human carcinomas. Carcinogenesis 2011, 32, 796–803. [Google Scholar]
- M’Kacher, R.; Bennaceur-Griscelli, A.; Girinsky, T.; Koscielny, S.; Delhommeau, F.; Dossou, J.; Violot, D.; Leclercq, E.; Courtier, M.H.; Beron-Gaillard, N.; et al. Telomere shortening and associated chromosomal instability in peripheral blood lymphocytes of patients with Hodgkin’s lymphoma prior to any treatment are predictive of second cancers. Int. J. Radiat. Oncol. Biol. Phys. 2007, 68, 465–471. [Google Scholar]
- Aldrup-Macdonald, M.E.; Sullivan, B.A. The past, present, and future of human centromere genomics. Genes 2014, 5, 33–50. [Google Scholar]
- Barra, V.; Fachinetti, D. The dark side of centromeres: Types, causes and consequences of structural abnormalities implicating centromeric DNA. Nat. Commun. 2018, 9, 4340. [Google Scholar] [PubMed] [Green Version]
- Gagliardi, M.; Strazzullo, M.; Matarazzo, M.R. DNMT3B Functions: Novel Insights From Human Disease. Front. Cell Dev. Biol. 2018, 6, 140. [Google Scholar] [PubMed]
- Brown, D.C.; Grace, E.; Sumner, A.T.; Edmunds, A.T.; Ellis, P.M. ICF syndrome (immunodeficiency, centromeric instability and facial anomalies): Investigation of heterochromatin abnormalities and review of clinical outcome. Hum. Genet. 1995, 96, 411–416. [Google Scholar]
- M’Kacher, R.; Maalouf, E.E.; Ricoul, M.; Heidingsfelder, L.; Laplagne, E.; Cuceu, C.; Hempel, W.M.; Colicchio, B.; Dieterlen, A.; Sabatier, L. New tool for biological dosimetry: Reevaluation and automation of the gold standard method following telomere and centromere staining. Mutat. Res. 2014, 770, 45–53. [Google Scholar]
- Kaddour, A.; Colicchio, B.; Buron, D.; El Maalouf, E.; Laplagne, E.; Borie, C.; Ricoul, M.; Lenain, A.; Hempel, W.M.; Morat, L.; et al. Transmission of Induced Chromosomal Aberrations through Successive Mitotic Divisions in Human Lymphocytes after In Vitro and In Vivo Radiation. Sci. Rep. 2017, 7, 3291. [Google Scholar]
- M’Kacher, R.; El Maalouf, E.; Terzoudi, G.; Ricoul, M.; Heidingsfelder, L.; Karachristou, I.; Laplagne, E.; Hempel, W.M.; Colicchio, B.; Dieterlen, A.; et al. Detection and automated scoring of dicentric chromosomes in nonstimulated lymphocyte prematurely condensed chromosomes after telomere and centromere staining. Int. J. Radiat. Oncol. Biol. Phys. 2015, 91, 640–649. [Google Scholar]
- Murnane, J.P. Telomere dysfunction and chromosome instability. Mutat. Res. 2012, 730, 28–36. [Google Scholar]
- Sfeir, A.; Kosiyatrakul, S.T.; Hockemeyer, D.; MacRae, S.L.; Karlseder, J.; Schildkraut, C.L.; de Lange, T. Mammalian telomeres resemble fragile sites and require TRF1 for efficient replication. Cell 2009, 138, 90–103. [Google Scholar]
- Mitchell, T.R.; Glenfield, K.; Jeyanthan, K.; Zhu, X.D. Arginine methylation regulates telomere length and stability. Mol. Cell. Biol. 2009, 29, 4918–4934. [Google Scholar]
- Gascoigne, K.E.; Cheeseman, I.M. Induced dicentric chromosome formation promotes genomic rearrangements and tumorigenesis. Chromosome Res. 2013, 21, 407–418. [Google Scholar] [PubMed] [Green Version]
- Schuler, N.; Palm, J.; Schmitz, S.; Lorat, Y.; Rube, C.E. Increasing genomic instability during cancer therapy in a patient with Li-Fraumeni syndrome. Clin. Transl. Radiat. Oncol. 2017, 7, 71–78. [Google Scholar] [PubMed] [Green Version]
- Sansregret, L.; Vanhaesebroeck, B.; Swanton, C. Determinants and clinical implications of chromosomal instability in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 139–150. [Google Scholar] [PubMed] [Green Version]
- Blackburn, E.H. Switching and signaling at the telomere. Cell 2001, 106, 661–673. [Google Scholar]
- Girinsky, T.; M’Kacher, R.; Lessard, N.; Koscielny, S.; Elfassy, E.; Raoux, F.; Carde, P.; Santos, M.D.; Margainaud, J.P.; Sabatier, L.; et al. Prospective coronary heart disease screening in asymptomatic Hodgkin lymphoma patients using coronary computed tomography angiography: Results and risk factor analysis. Int. J. Radiat. Oncol. Biol. Phys. 2014, 89, 59–66. [Google Scholar]
- Claude, E.; Decottignies, A. Telomere maintenance mechanisms in cancer: Telomerase, ALT or lack thereof. Curr. Opin. Genet. Dev. 2020, 60, 1–8. [Google Scholar]
- Blasco, M.A. The epigenetic regulation of mammalian telomeres. Nat. Rev. Genet. 2007, 8, 299–309. [Google Scholar]
- Armando, R.G.; Mengual Gomez, D.L. Telomeropathies: Etiology, diagnosis, treatment and follow-up. Ethical Leg. Consid. 2019, 96, 3–16. [Google Scholar]
- Savage, S.A. Beginning at the ends: Telomeres and human disease. F1000Research 2018, 7, 524. [Google Scholar]
- Reddel, R.R. Telomere maintenance mechanisms in cancer: Clinical implications. Curr. Pharm. Des. 2014, 20, 6361–6374. [Google Scholar]
- Cawthon, R.M. Telomere length measurement by a novel monochrome multiplex quantitative PCR method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar] [PubMed] [Green Version]
- Ferreira, M.S.V.; Kirschner, M.; Halfmeyer, I.; Estrada, N.; Xicoy, B.; Isfort, S.; Vieri, M.; Zamora, L.; Abels, A.; Bouillon, A.S.; et al. Comparison of flow-FISH and MM-qPCR telomere length assessment techniques for the screening of telomeropathies. Ann. N. Y. Acad. Sci. 2019. [Google Scholar] [CrossRef] [Green Version]
- Canela, A.; Vera, E.; Klatt, P.; Blasco, M.A. High-throughput telomere length quantification by FISH and its application to human population studies. Proc. Natl. Acad. Sci. USA 2007, 104, 5300–5305. [Google Scholar] [PubMed] [Green Version]
- Callet-Bauchu, E.; Salles, G.; Gazzo, S.; Poncet, C.; Morel, D.; Pages, J.; Coiffier, B.; Coeur, P.; Felman, P. Translocations involving the short arm of chromosome 17 in chronic B-lymphoid disorders: Frequent occurrence of dicentric rearrangements and possible association with adverse outcome. Leukemia 1999, 13, 460–468. [Google Scholar]
- Streubel, B.; Sauerland, C.; Heil, G.; Freund, M.; Bartels, H.; Lengfelder, E.; Wandt, H.; Ludwig, W.D.; Nowotny, H.; Baldus, M.; et al. Correlation of cytogenetic, molecular cytogenetic, and clinical findings in 59 patients with ANLL or MDS and abnormalities of the short arm of chromosome 12. Br. J. Haematol. 1998, 100, 521–533. [Google Scholar]
- Ittel, A.; Gervais, C.; Galoisy, A.C.; M’Kacher, R.; Mauvieux, L.; Fornecker, L.M. Triradial and quadriradial chromosomes detected in a case of B-cell prolymphocytic leukaemia. Br. J. Haematol. 2017, 179, 704. [Google Scholar]
- Berger, R.; Busson-Le Coniat, M. Centric and pericentric chromosome rearrangements in hematopoietic malignancies. Leukemia 1999, 13, 671–678. [Google Scholar]
- Mackinnon, R.N.; Campbell, L.J. The role of dicentric chromosome formation and secondary centromere deletion in the evolution of myeloid malignancy. Genet. Res. Int. 2011, 2011, 643628. [Google Scholar]
- MacKinnon, R.N.; Duivenvoorden, H.M.; Campbell, L.J.; Wall, M. The Dicentric Chromosome dic(20;22) Is a Recurrent Abnormality in Myelodysplastic Syndromes and Is a Product of Telomere Fusion. Cytogenet. Genome Res. 2016, 150, 262–272. [Google Scholar]
- Sarova, I.; Brezinova, J.; Zemanova, Z.; Ransdorfova, S.; Izakova, S.; Svobodova, K.; Pavlistova, L.; Berkova, A.; Cermak, J.; Jonasova, A.; et al. Molecular cytogenetic analysis of dicentric chromosomes in acute myeloid leukemia. Leuk. Res. 2016, 43, 51–57. [Google Scholar]
- Sarova, I.; Brezinova, J.; Zemanova, Z.; Ransdorfova, S.; Svobodova, K.; Izakova, S.; Pavlistova, L.; Lizcova, L.; Berkova, A.; Skipalova, K.; et al. High frequency of dicentric chromosomes detected by multi-centromeric FISH in patients with acute myeloid leukemia and complex karyotype. Leuk. Res. 2018, 68, 85–89. [Google Scholar] [PubMed]
- Churpek, J.E.; Bresnick, E.H. Transcription factor mutations as a cause of familial myeloid neoplasms. J. Clin. Investig. 2019, 129, 476–488. [Google Scholar] [PubMed] [Green Version]
- Ganmore, I.; Smooha, G.; Izraeli, S. Constitutional aneuploidy and cancer predisposition. Hum. Mol. Genet. 2009, 18, R84–R93. [Google Scholar]
- Hertzberg, L.; Vendramini, E.; Ganmore, I.; Cazzaniga, G.; Schmitz, M.; Chalker, J.; Shiloh, R.; Iacobucci, I.; Shochat, C.; Zeligson, S.; et al. Down syndrome acute lymphoblastic leukemia, a highly heterogeneous disease in which aberrant expression of CRLF2 is associated with mutated JAK2, a report from the International BFM Study Group. Blood 2010, 115, 1006–1017. [Google Scholar]
- Deakin, J.E.; Potter, S.; O’Neill, R.; Ruiz-Herrera, A.; Cioffi, M.B.; Eldridge, M.D.B.; Fukui, K.; Marshall Graves, J.A.; Griffin, D.; Grutzner, F.; et al. Chromosomics: Bridging the Gap between Genomes and Chromosomes. Genes 2019, 10, 627. [Google Scholar]
- Guinobert, I.; Blondeau, C.; Colicchio, B.; Oudrhiri, N.; Dieterlen, A.; Jeandidier, E.; Deschenes, G.; Bardot, V.; Cotte, C.; Ripoche, I.; et al. The Use of Natural Agents to Counteract Telomere Shortening: Effects of a Multi-Component Extract of Astragalus mongholicus Bunge and Danazol. Biomedicines 2020, 8, 22. [Google Scholar]
- M’Kacher, R.; Breton, L.; Colicchio, B.; Puget, H.; Hempel, W.M.; Al Jawhari, M.; Jeandidier, E.; Frey, M. Benefit of an association of an antioxidative substrate and a traditional chinese medicine on telomere elongation. Cell. Mol. Biol. 2019, 65, 54–58. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | No. of Patients |
|---|---|
| Cancer patients | 50 |
| Male | 32 |
| Age (years) | 56 |
| Type | |
| Hodgkin lymphoma | 18 |
| Non Hodgkin lymphoma | 15 |
| Mantel Cell lymphoma | 8 |
| B-cell prolymphocytic leukemia | 2 |
| Myelodysplasia syndrome | 4 |
| Other | 5 |
| Genetic syndrome | 44 |
| Male | 27 |
| Age (years) | 32 |
| Type | |
| Turner syndrome | 9 |
| Down syndrome | 4 |
| Klinfelter syndrome | 2 |
| Li-Fraumeni | 2 |
| Telomerepathies | 2 |
| Other * | 15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
M’kacher, R.; Colicchio, B.; Borie, C.; Junker, S.; Marquet, V.; Heidingsfelder, L.; Soehnlen, K.; Najar, W.; Hempel, W.M.; Oudrhiri, N.; et al. Telomere and Centromere Staining Followed by M-FISH Improves Diagnosis of Chromosomal Instability and Its Clinical Utility. Genes 2020, 11, 475. https://doi.org/10.3390/genes11050475
M’kacher R, Colicchio B, Borie C, Junker S, Marquet V, Heidingsfelder L, Soehnlen K, Najar W, Hempel WM, Oudrhiri N, et al. Telomere and Centromere Staining Followed by M-FISH Improves Diagnosis of Chromosomal Instability and Its Clinical Utility. Genes. 2020; 11(5):475. https://doi.org/10.3390/genes11050475
Chicago/Turabian StyleM’kacher, Radhia, Bruno Colicchio, Claire Borie, Steffen Junker, Valentine Marquet, Leonhard Heidingsfelder, Kevin Soehnlen, Wala Najar, William M. Hempel, Noufissa Oudrhiri, and et al. 2020. "Telomere and Centromere Staining Followed by M-FISH Improves Diagnosis of Chromosomal Instability and Its Clinical Utility" Genes 11, no. 5: 475. https://doi.org/10.3390/genes11050475