Molecular Cloning and Characterization of SYCP3 and TSEG2 Genes in the Testicles of Sexually Mature and Immature Yak

Abstract
:1. Introduction
2. Materials and Methods
2.1. Total RNA Extraction and cDNA Synthesis
2.2. Molecular Cloning
2.3. Exploration through Semi-Quantitative PCR
2.4. Exploration through Quantitative Real-Time PCR
2.5. Primer Designing and Bioinformatics Analysis
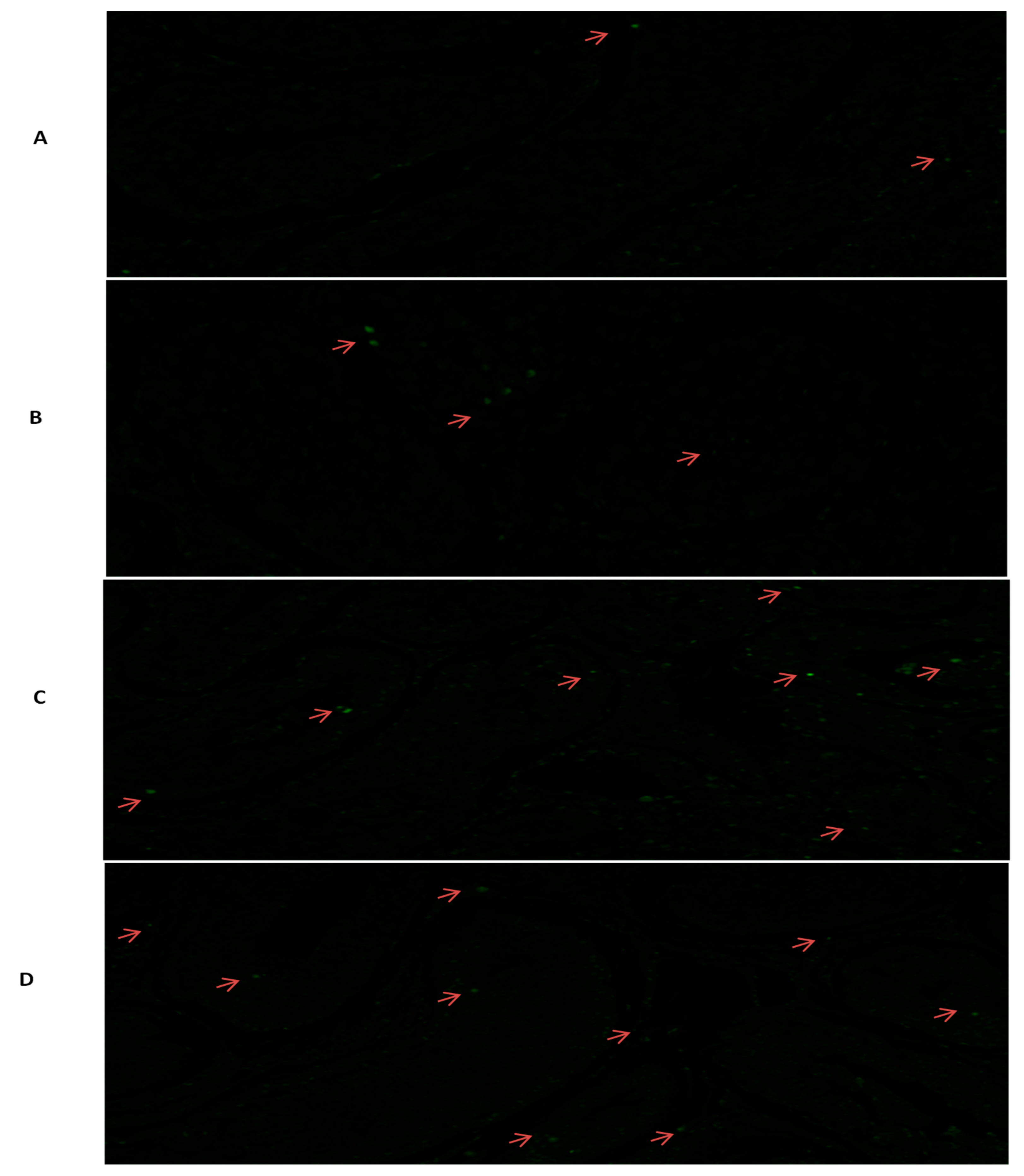
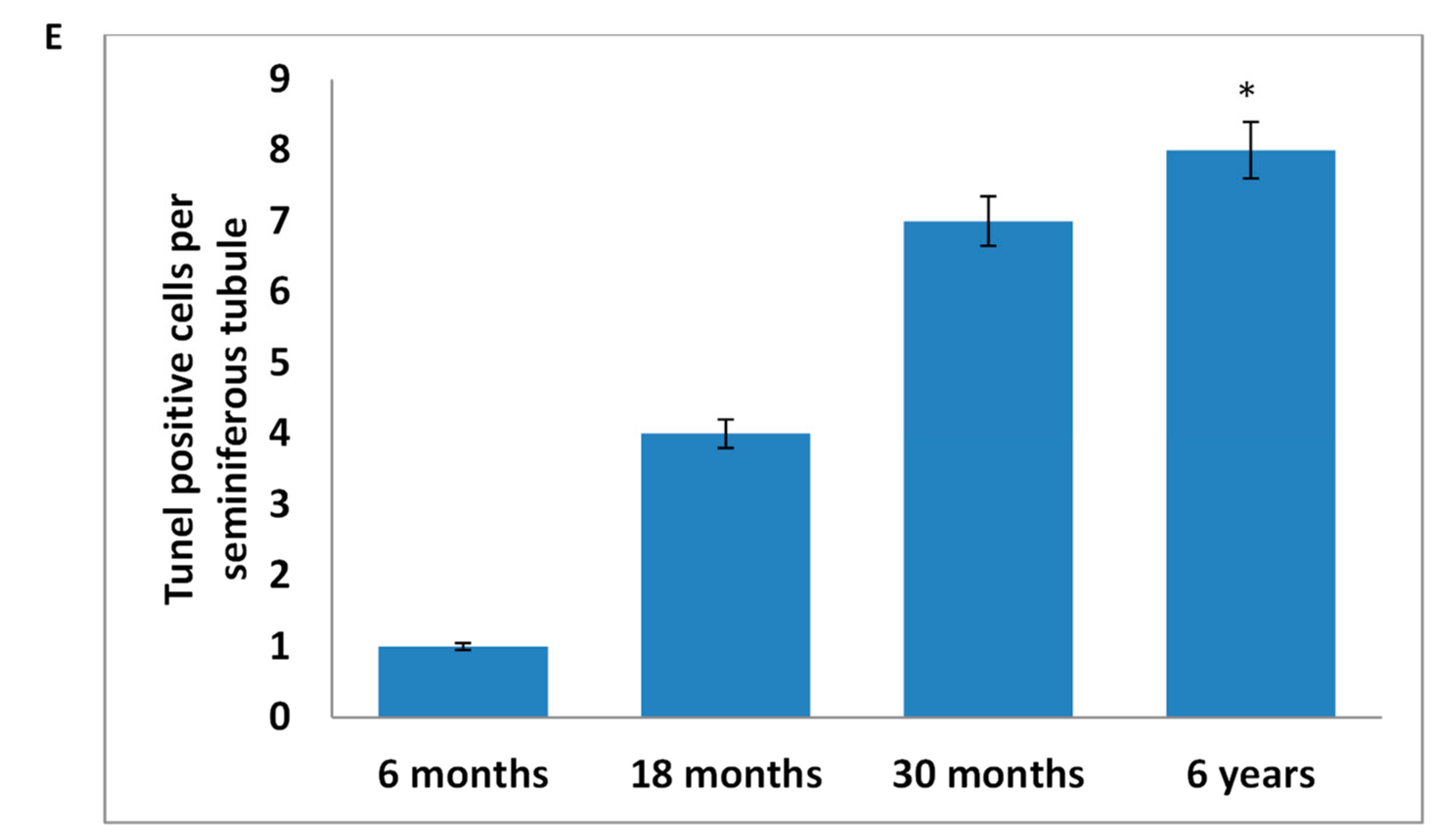
2.6. Exploration through Tunnel Assay
2.7. Tissue Immunostaining
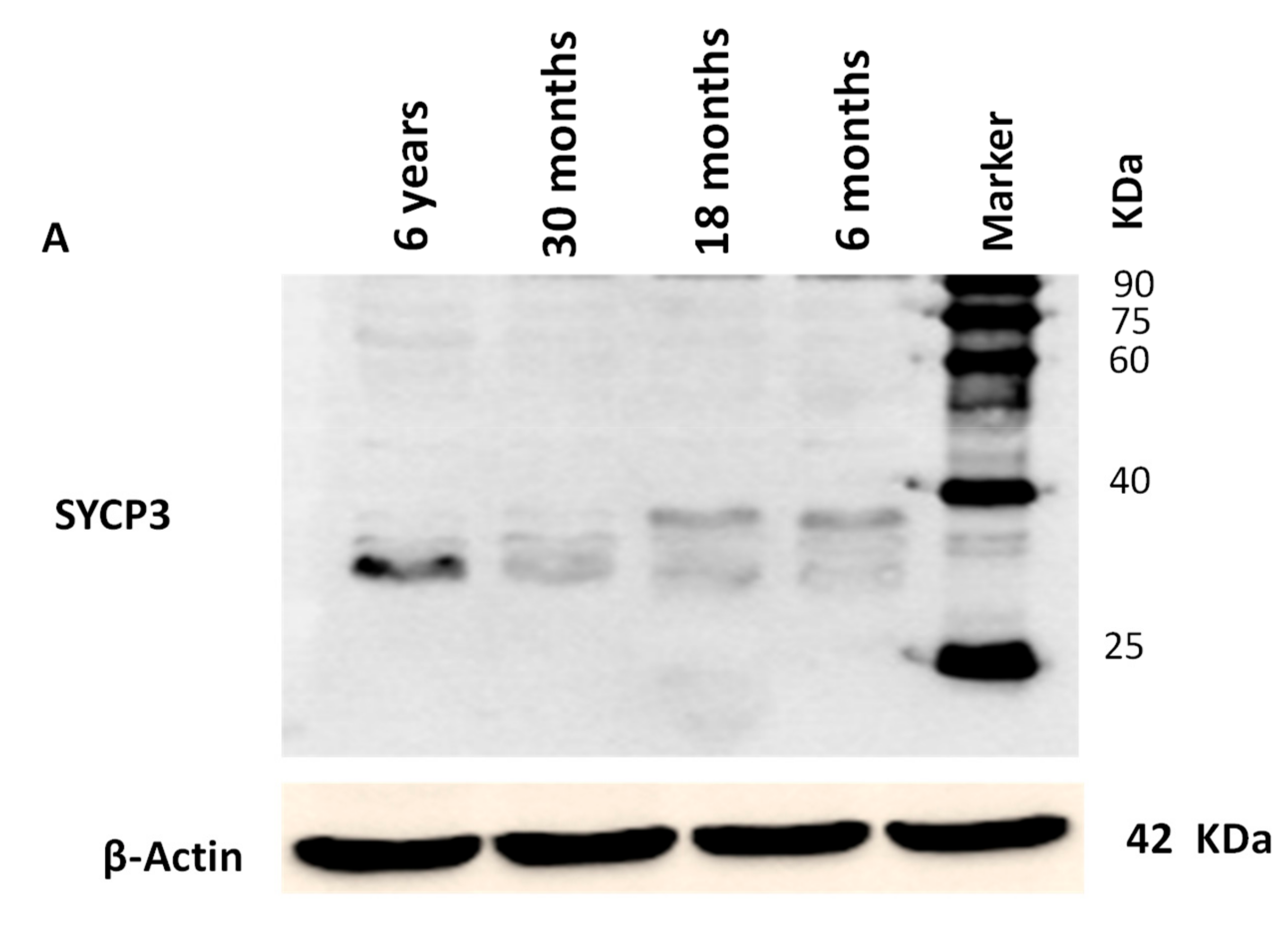
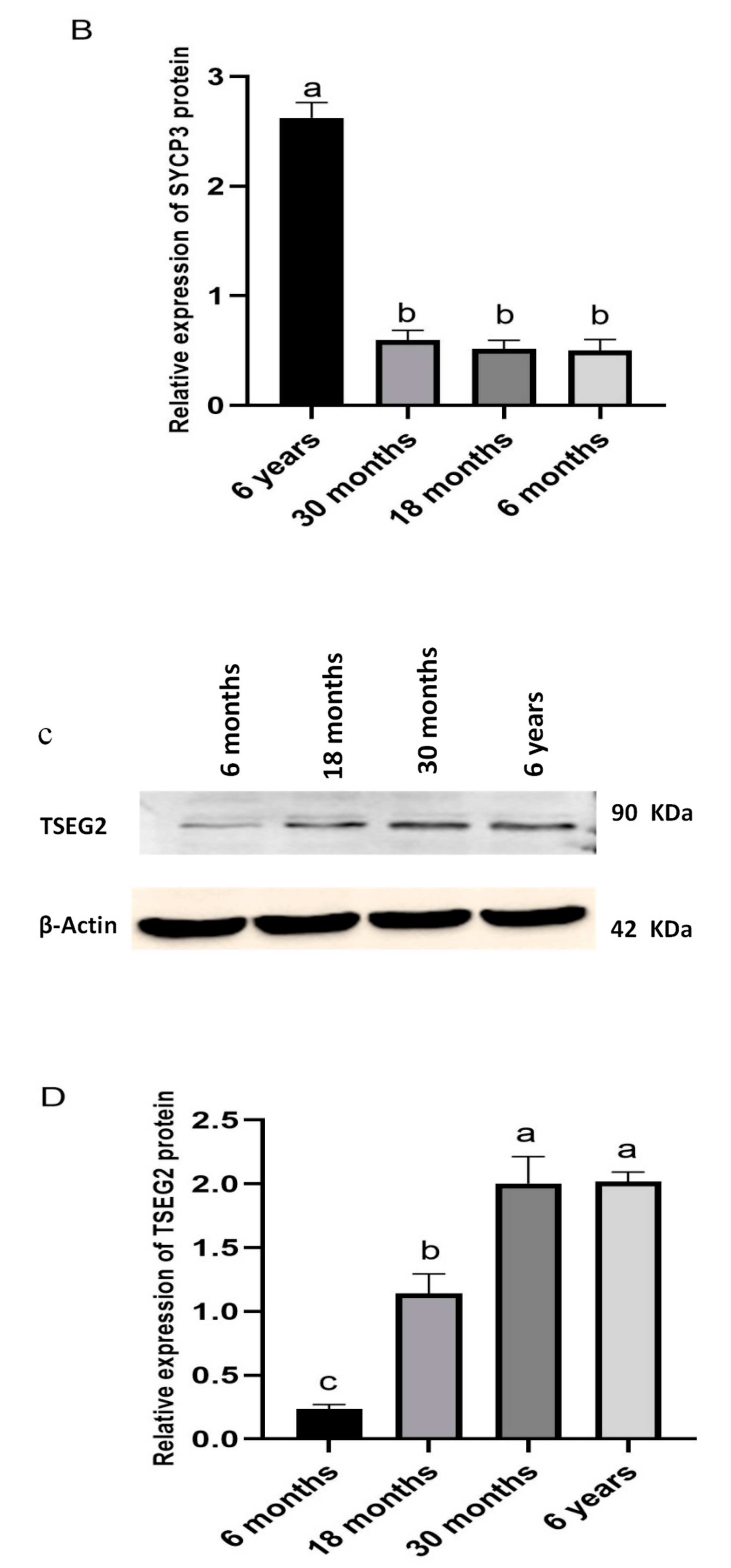
2.8. Western Blotting
2.9. Statistical Analyses
3. Results
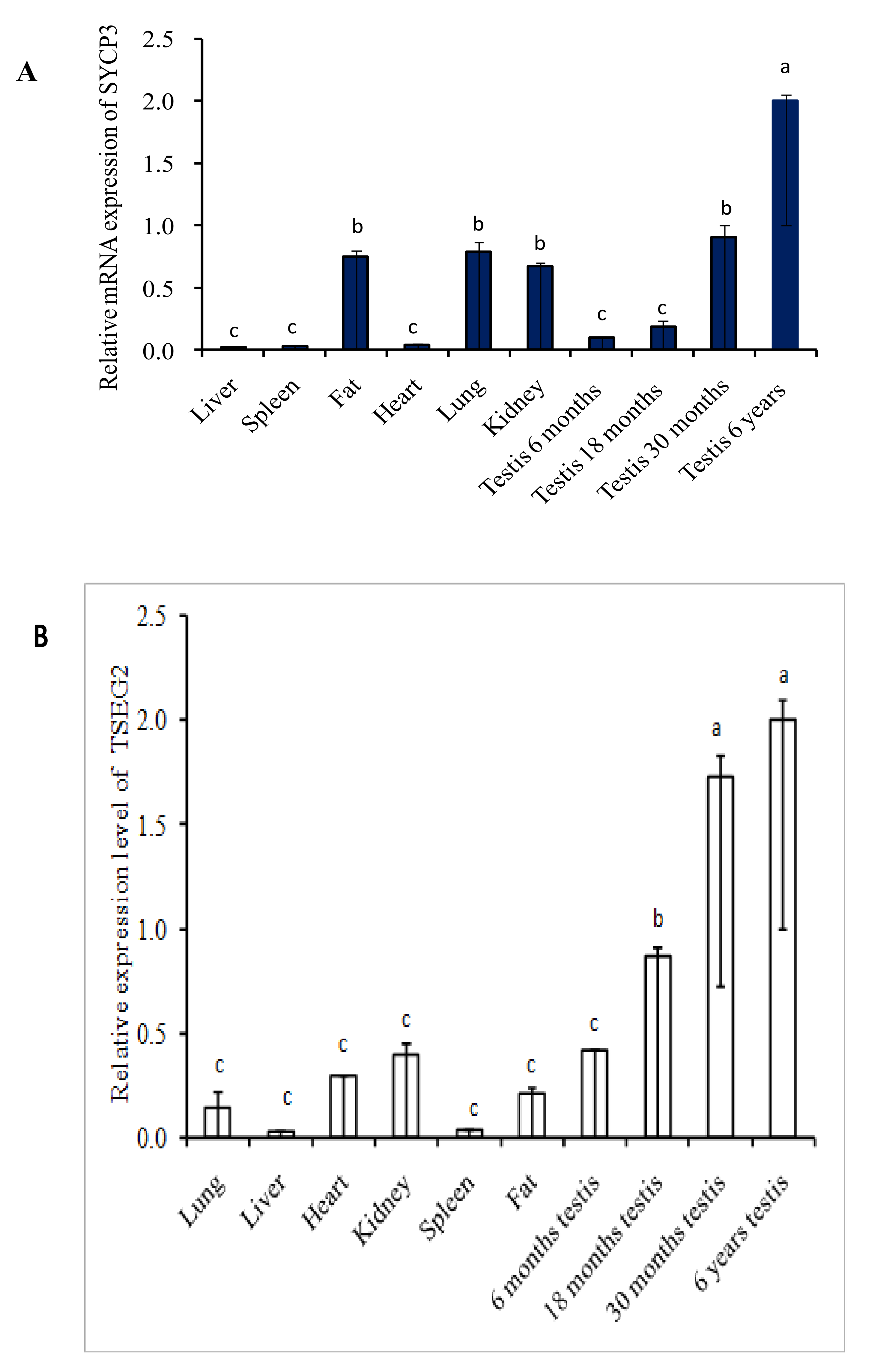
3.1. Expression Profile of SYCP3 and TSEG2 by Semi-Quantitative PCR and Quantitative Real Time PCR
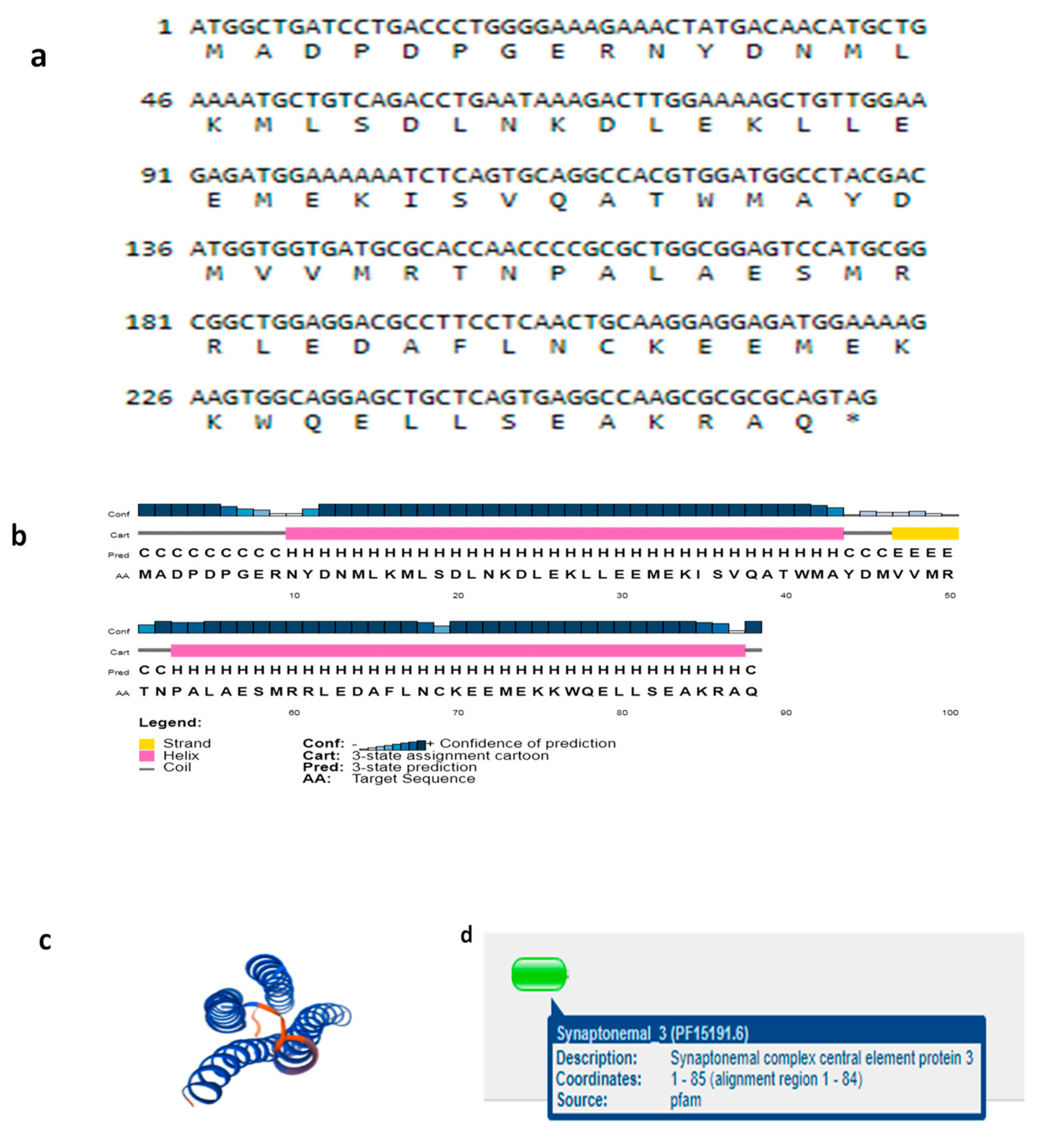
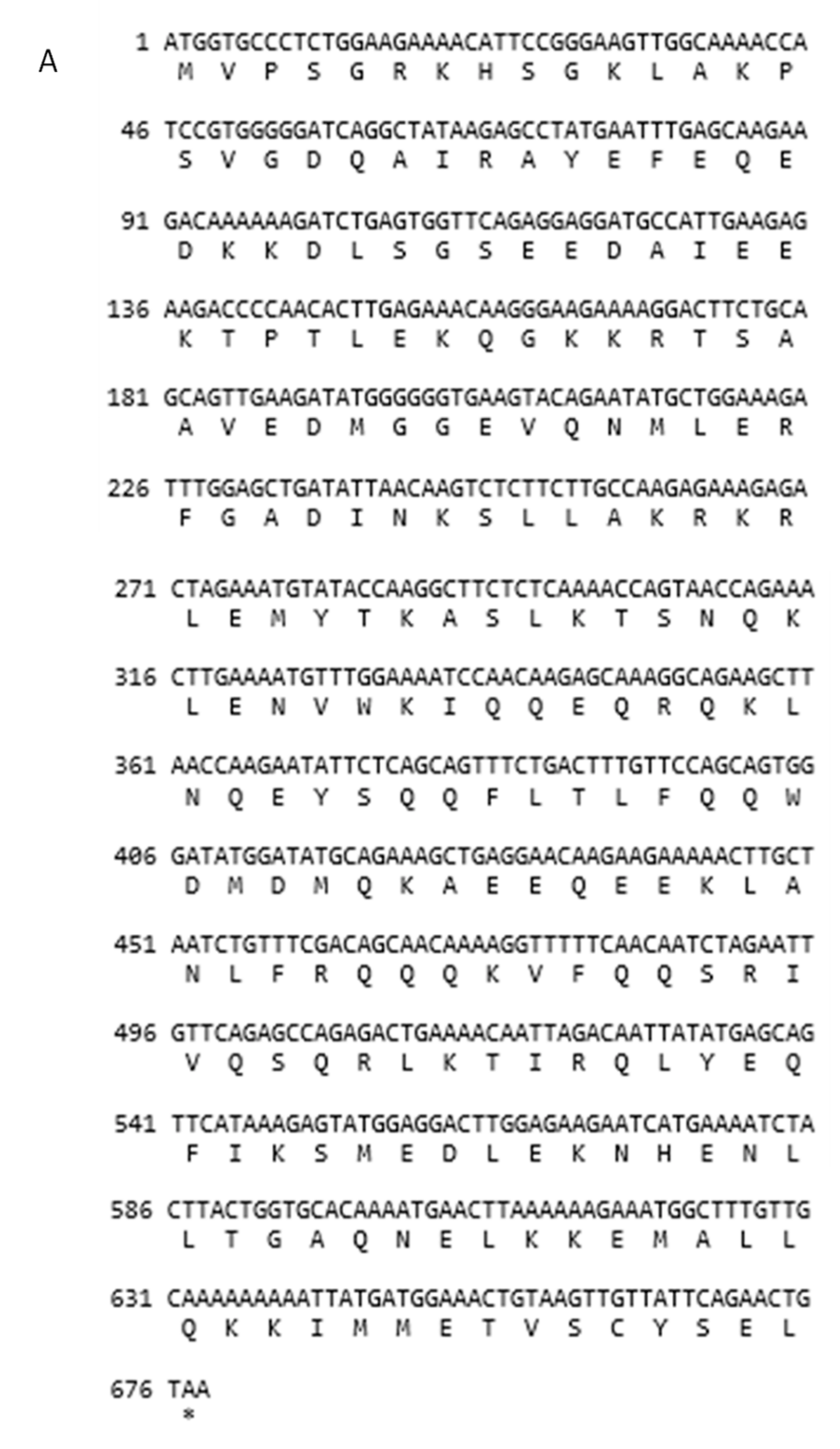
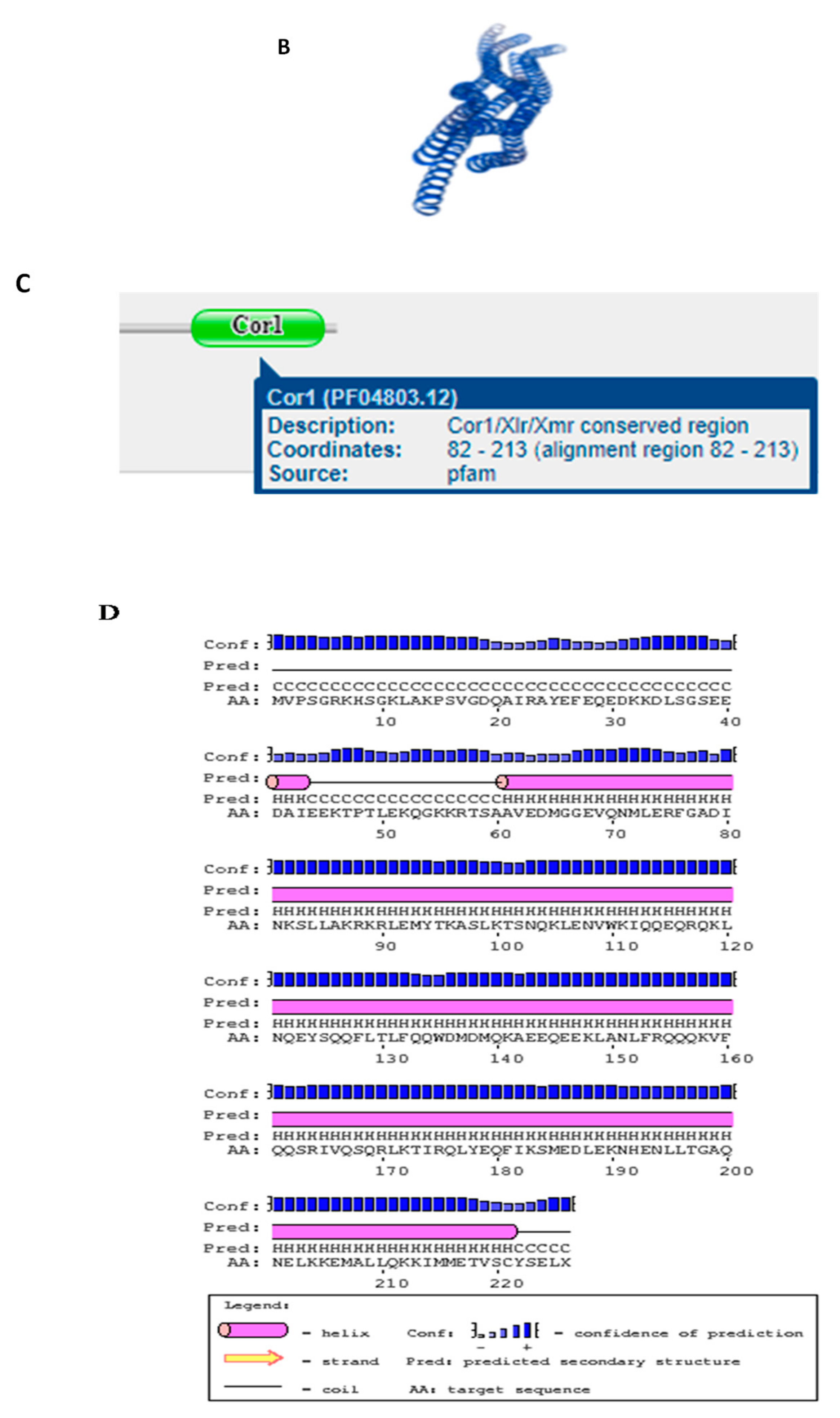
3.2. Structures of SYCP3 and TSEG2 Gene
3.3. Tunel Assay Analysis
3.4. Western Blotting Analysis
3.5. Immunostaining Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Syrjänen, J.L.; Pellegrini, L.; Davies, O.R. A molecular model for the role of SYCP3 in meiotic chromosome organisation. eLife 2014, 3, e02963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahoon, C.K.; Hawley, R.S. Regulating the construction and demolition of the synaptonemal complex. Nat. Struct. Mol. Biol. 2016, 23, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, J.G.; Hoja, M.R.; Wilbertz, J.; Nordqvist, K.; Hoog, C. Female germ cell aneuploidy and embryo death in mice lacking the meiosis-specific protein SCP3. Science 2002, 296, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- De Vries, F.A.; de Boer, E.; van den Bosch, M.; Baarends, W.M.; Ooms, M.; Yuan, L.; Liu, J.G.; van Zeeland, A.A.; Heyting, C.; Pastink, A. Mouse Sycp1 functions in synaptonemal complex assembly, meiotic recombination, and XY body formation. Genes Dev. 2005, 19, 1376–1389. [Google Scholar] [CrossRef] [PubMed]
- Kouznetsova, A.; Benavente, R.; Pastink, A.; Hoog, C. Meiosis in mice without a synaptonemal complex. PLoS ONE 2011, 6, e28255. [Google Scholar] [CrossRef]
- Bolor, H.; Mori, T.; Nishiyama, S.; Ito, Y.; Hosoba, E.; Inagaki, H.; Kogo, H.; Ohye, T.; Tsutsumi, M.; Kato, T.; et al. Mutations of the SYCP3 gene in women with recurrent pregnancy loss. Am. J. Hum. Genet. 2009, 84, 14–20. [Google Scholar] [CrossRef]
- Costa, Y.; Speed, R.; Ollinger, R.; Alsheimer, M.; Semple, C.A.; Gautier, P.; Maratou, K.; Novak, I.; Höög, C.; Benavente, R.; et al. Two novel proteins recruited by synaptonemal complex protein 1 (SYCP1) are at the centre of meiosis. J. Cell Sci. 2005, 118, 2755–2762. [Google Scholar] [CrossRef] [Green Version]
- Hamer, G.; Gell, K.; Kouznetsova, A.; Novak, I.; Benavente, R.; Höög, C. Characterization of a novel meiosis-specific protein within the central element of the synaptonemal complex. J. Cell Sci. 2006, 119, 4025–4032. [Google Scholar] [CrossRef] [Green Version]
- Schramm, S.; Fraune, J.; Naumann, R.; Hernandez-Hernandez, A.; Höög, C.; Cooke, H.J.; Alsheimer, M.; Benavente, R. A novel mouse synaptonemal complex protein is essential for loading of central element proteins, recombination, and fertility. PLoS Genet. 2011, 7, e1002088. [Google Scholar] [CrossRef]
- Garcia-Cruz, R.; Roig, I.; Caldes, M.G. Maternal origin of the human aneuploidies. Are homolog synapsis and recombination to blame? Notes (learned) from the underbelly. Genome Dyn. 2009, 5, 128–136. [Google Scholar]
- Handel, M.A.; Schimenti, J.C. Genetics of mammalian meiosis: Regulation, dynamics and impact on fertility. Nat. Rev. Genet. 2010, 11, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Page, S.L.; Hawley, R.S. The genetics and molecular biology of the synaptonemal complex. Annu. Rev. Cell Dev. Biol. 2004, 20, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Syrjänen, J.L.; Heller, I.; Candelli, A.; Davies, O.R.; Peterman, E.J.G.; Wuite, G.J.L.; Pellegrini, L. Single-molecule observation of DNA compaction by meiotic protein SYCP3. eLife 2017, 6, e22582. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.Y.; Yanling, G.; Feng, J.; Zhou, W.; Xue, Y.; Yuequan, S. Structural Insight into the Central Element Assembly of the Synaptonemal Complex. Sci. Rep. 2014, 4, 7059. [Google Scholar] [CrossRef]
- Hu, T.; Wang, Z.; Zeng, F.; Chen, X.; Gu, Z.; Zheng, L.; Tong, Q. Expression Pattern of Testis-specific Expressed Gene 2 in Cryptorchidism Model and Its Role in Apoptosis of Spermatogenic Cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2010, 30, 193–197. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Tong, Q.S.; Zeng, F.Q.; Liu, Y.; Gu, Z.H.; Zheng, L.D.; Cai, J.B.; Jiang, G.S. Cloning and expression of a novel mouse testis gene TSEG-2. Natl. J. Androl. 2009, 15, 99–105. (In Chinese) [Google Scholar]
- Fraune, J.; Schramm, S.; Alsheimer, M.; Benavente, R. The mammalian synaptonemal complex: Protein components, assembly and role in meiotic recombination. Exp. Cell Res. 2012, 318, 1340–1346. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, J.G.; Zhao, J.; Brundell, E.; Daneholt, B.; Hoog, C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol. Cell 2000, 5, 73–83. [Google Scholar] [CrossRef]
- Miyamoto, T.; Hasuike, S.; Yogev, L.; Maduro, M.R.; Ishikawa, M.; Westphal, H.; Lamb, D.J. Azoospermia in patients heterozygous for a mutation in SYCP3. Lancet 2003, 362, 1714–1719. [Google Scholar] [CrossRef]
- Wang, S.; Pan, Z.; Zhang, Q.; Xie, Z.; Liu, H.; Li, Q. Differential mRNA Expression and Promoter Methylation Status of SYCP3 Gene in Testes of Yaks and Cattle-Yaks. Reprod. Domest. Anim. 2012, 47, 455–462. [Google Scholar] [CrossRef]
- Shi, Y.Q.; Zhuang, X.J.; Xu, B.; Hua, J.; Liao, S.Y.; Shi, Q.; Cooke, H.J.; Han, C. SYCP3-like X-linked 2 is expressed in meiotic germ cells and interacts with synaptonemal complex central element protein 2 and histone acetyl transferase TIP60. Gene 2013, 527, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Xiang, L.; Guo, X.; Bao, P.-J.; Jin, S.; Wu, X.Y. The low expression of Dmrt7 is associated with spermatogenic arrest in cattle-yak. Mol. Biol. Rep. 2014, 41, 7255–7263. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, H.; Yao, B.; Xu, W.; Chen, J.; Zhou, X. Lncrna H19/MIR-675 axis regulates cardiomyocyte apoptosis by targeting vdac1 in diabetic cardiomyopathy. Sci. Rep. 2016, 6, 36340. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1107. [Google Scholar] [CrossRef]
- Coulter, G.H.; Kastelic, J.P. Management programs for developing bulls. In Current Veterinary Therapy-Food Animal Practice, 4th ed.; Howard, J.L., Smith, R., Eds.; W.B. Saunders: Philadelphia, PA, USA, 1999; pp. 127–136. [Google Scholar]
- Hassold, T.; Hunt, P. To err (meiotically) is human: The genesis of human aneuploidy. Nat. Rev. Genet. 2001, 2, 280–291. [Google Scholar] [CrossRef]
- Judis, L.; Chan, E.R.; Schwartz, S.; Seftel, A.; Hassold, T. Meiosis I arrest and azoospermia in an infertile male explained by failure of formation of a component of the synaptonemal complex. Fertil. Steril. 2004, 81, 205–209. [Google Scholar] [CrossRef]
- Aarabi, M.; Modarressi, M.H.; Soltanghoraee, H.; Behjati, R.; Amirjannati, N.; Akhondi, M.M. Testicular expression of synaptonemal complex protein 3 (SYCP3) messenger ribonucleic acid in 110 patients with nonobstructive azoospermia. Fertil. Steril. 2006, 86, 325–331. [Google Scholar] [CrossRef]
- Calenda, A.; Allenet, B.; Escalier, D.; Bach, J.F.; Garchon, H.J. The meiosis-specific Xmr gene product is homologous to the lymphocyte Xlr protein and is a component of the XY body. EMBO J. 1994, 13, 100–109. [Google Scholar] [CrossRef]
- Abrahan, H.; Masich, S.; Fukuda, T.; Kouznetsova, A.; Sandin, S.; Daneholt, B.; Höög, C. The central element of the synaptonemal complex in mice is organized as a bilayer junction structure. J. Cell Sci. 2016, 129, 2239–2249. [Google Scholar]
- Schmekel, K.; Wahrman, J.; Skoglund, U.; Daneholt, B. The central region of the synaptonemal complex in Blaps cribrosa studied by electron microscope tomography. Chromosoma 1993, 102, 669–681. [Google Scholar] [CrossRef]
- Dobson, M.J.; Pearlman, R.E.; Karaiskakis, A.; Spyropoulos, B.; Moens, P.B. Synaptonemal complex proteins: Occurrence, epitope mapping and chromosome disjunction. J. Cell Sci. 1994, 107, 2749–2760. [Google Scholar] [PubMed]
- Izadyar, F.; den Ouden, K.; Creemers, L.B.; Creemers, L.B.; Posthuma, G.; Parvinen, M.; de Rooij, D.G. Proliferation and Differentiation of Bovine Type A Spermatogonia During Long-Term Culture. Biol. Reprod. 2003, 68, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, Y.; Saito, K.; Shinya, M.; Kawasaki, T.; Sakai, N. Evaluation of Sycp3, Plzf and Cyclin B3 expression and suitability as spermatogonia and spermatocyte markers in zebrafish. Gene Expr. Patterns 2011, 11, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Reynard, L.N.; Cocquet, J.; Burgoyne, P.S. The multi-copy mouse gene Sycp3-like Y-linked (Sly) encodes an abundant spermatid protein that interacts with a histone acetyltransferase and an acrosomal protein. Biol. Reprod. 2009, 81, 250–257. [Google Scholar] [CrossRef]
- Sazegari, A.; Kalantar, S.M.; Pashaiefar, H.; Mohtaram, S.; Honarvar, N.; Feizollahi, Z.; Ghasemi, N. The T657C polymorphism on the SYCP3 gene is associated with recurrent pregnancy loss. J. Assist. Reprod. Genet. 2014, 31, 1377–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession no | Gene | Primers Sequence (5’- >3’) | Product Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| NM_001206333.1 | TSEG2 (For cloning) | F: TGGCAGGTAGCTGAACAGGA R: GTTGGGGTCGCTGTGGTTC | 328 | 60.84 61.26 |
| XM_005903404.1 | TSEG2 (For gene expression) | F:CTGACCCTGGGGAAAGAAACT R: ACCATGTCGTAGGCCATCCA | 130 | 59.57 60.98 |
| XM_024992196.1 | SYCP3 (For cloning) | F: TACGCCTGTCCGGAGACATT R: GACTTTCGGACACTTGCCATC | 942 | 60.97 59.54 |
| XM_019961171.1 | SYCP3 (For gene expression) | F:TCCGGGAAGTTGGCAAAACC R: GGTCTTCTCTTCAATGGCATCC | 117 | 61.11 59.05 |
| NM_001034034.2 F: AATGAAAGGGCCATCACCATC | GAPDH | F:AATGAAAGGGCCATCACCATC R:CACCACCCTGTTGCTGTAGCCA | 204 | 55.85 60.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalwar, Q.; Chu, M.; Ahmad, A.A.; Ma, X.; Zhang, R.; Ma, F.; Xie, J.; Ding, X.; Wu, X.; Bao, P.; et al. Molecular Cloning and Characterization of SYCP3 and TSEG2 Genes in the Testicles of Sexually Mature and Immature Yak. Genes 2019, 10, 867. https://doi.org/10.3390/genes10110867
Kalwar Q, Chu M, Ahmad AA, Ma X, Zhang R, Ma F, Xie J, Ding X, Wu X, Bao P, et al. Molecular Cloning and Characterization of SYCP3 and TSEG2 Genes in the Testicles of Sexually Mature and Immature Yak. Genes. 2019; 10(11):867. https://doi.org/10.3390/genes10110867
Chicago/Turabian StyleKalwar, Qudratullah, Min Chu, Anum Ali Ahmad, Xiaoming Ma, Renzheng Zhang, Fulong Ma, Jianpeng Xie, Xuezhi Ding, Xiaoyun Wu, Pengjia Bao, and et al. 2019. "Molecular Cloning and Characterization of SYCP3 and TSEG2 Genes in the Testicles of Sexually Mature and Immature Yak" Genes 10, no. 11: 867. https://doi.org/10.3390/genes10110867