The Soybean Laccase Gene Family: Evolution and Possible Roles in Plant Defense and Stem Strength Selection

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Laccase Gene Family Members in Soybean
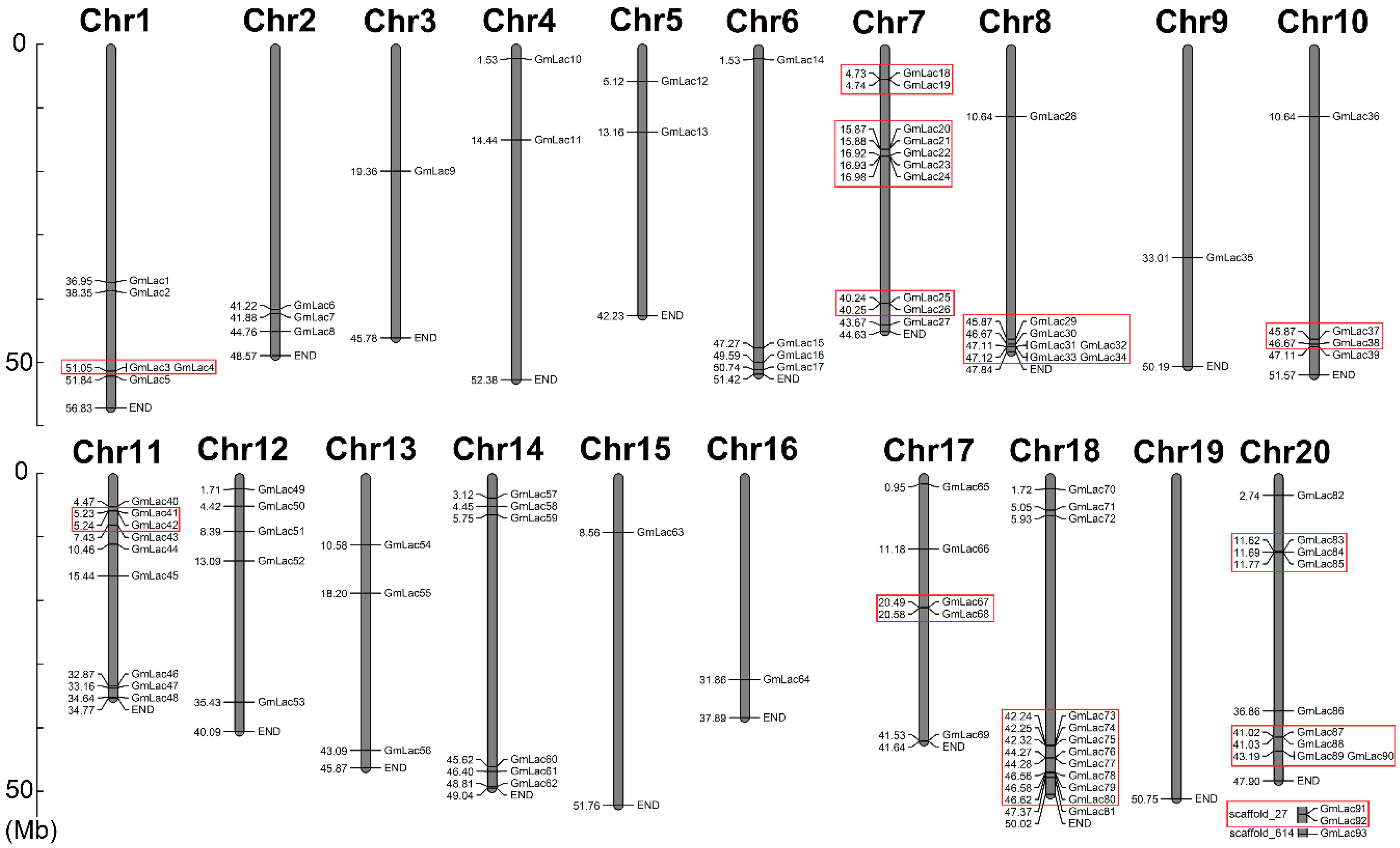
2.2. Physical Map of the Soybean Laccase Genes and Properties of the Laccase Proteins
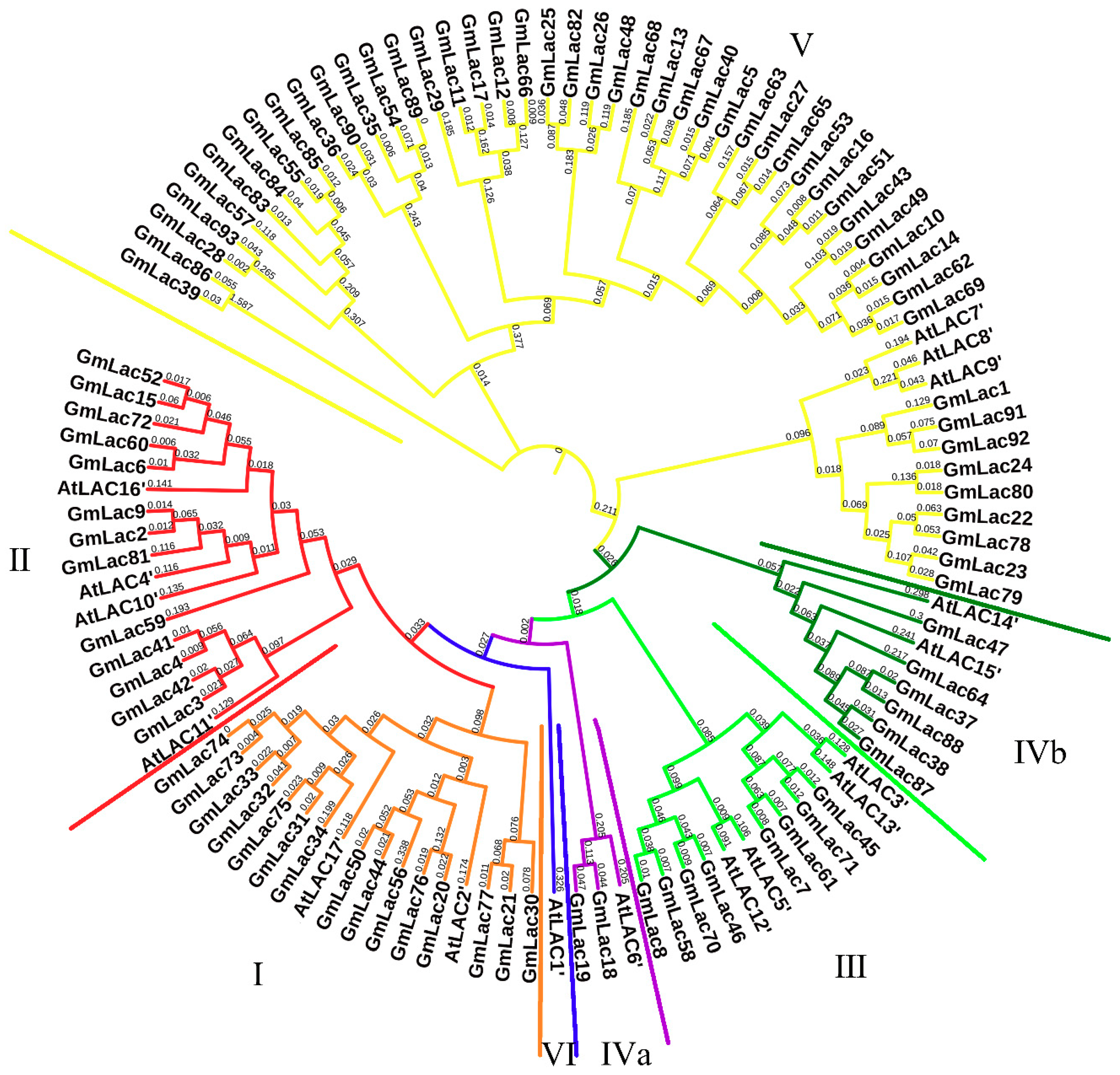
2.3. Sequence Alignment and Phylogenetic Analysis
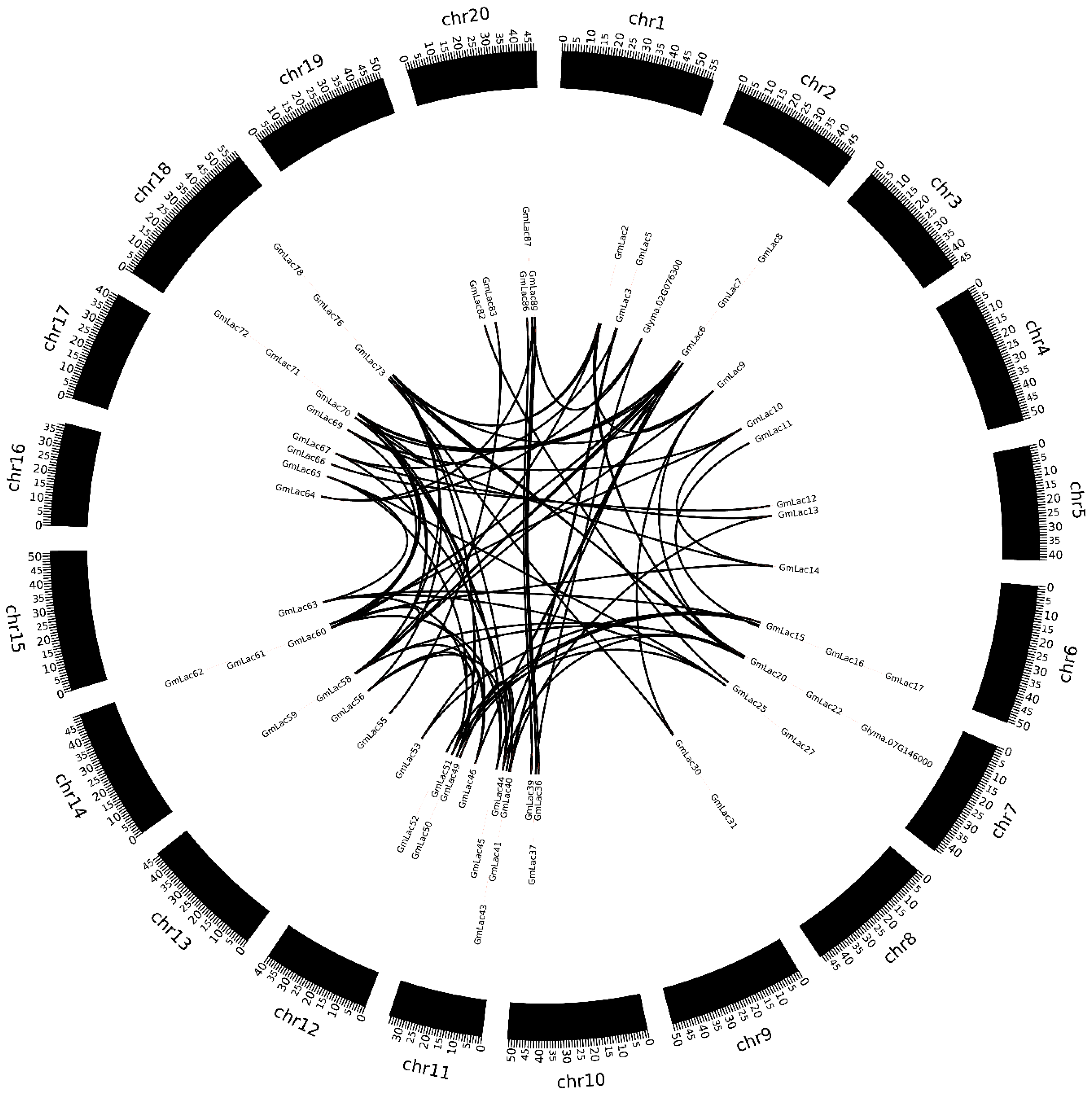
2.4. Gene Duplication and Expansion Analysis
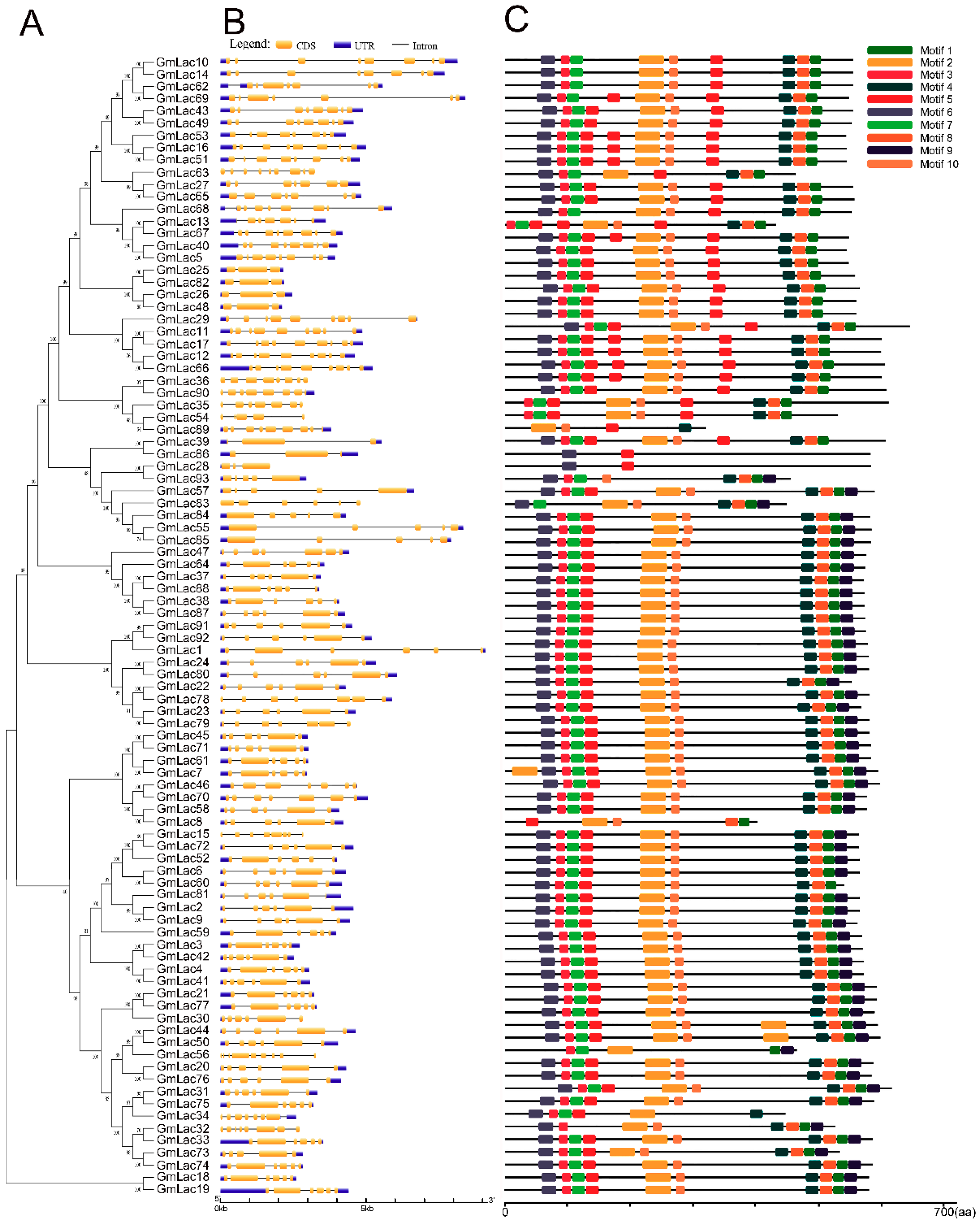
2.5. Gene Structure and Motif Compositions Analysis
2.6. Expression Pattern of Soybean Laccase Family Genes
2.7. Expression Patterns of GmLacs Following Phytophthora sojae Infection
2.8. qRT-PCR Analysis of GmLacs Expression Following Phytophthora sojae Infection
2.9. Selective Pressure Analysis
3. Results
3.1. Identification and Classification of Laccase Genes in Soybean
3.2. Phylogenetic Analysis of the Soybean Laccase Gene Family
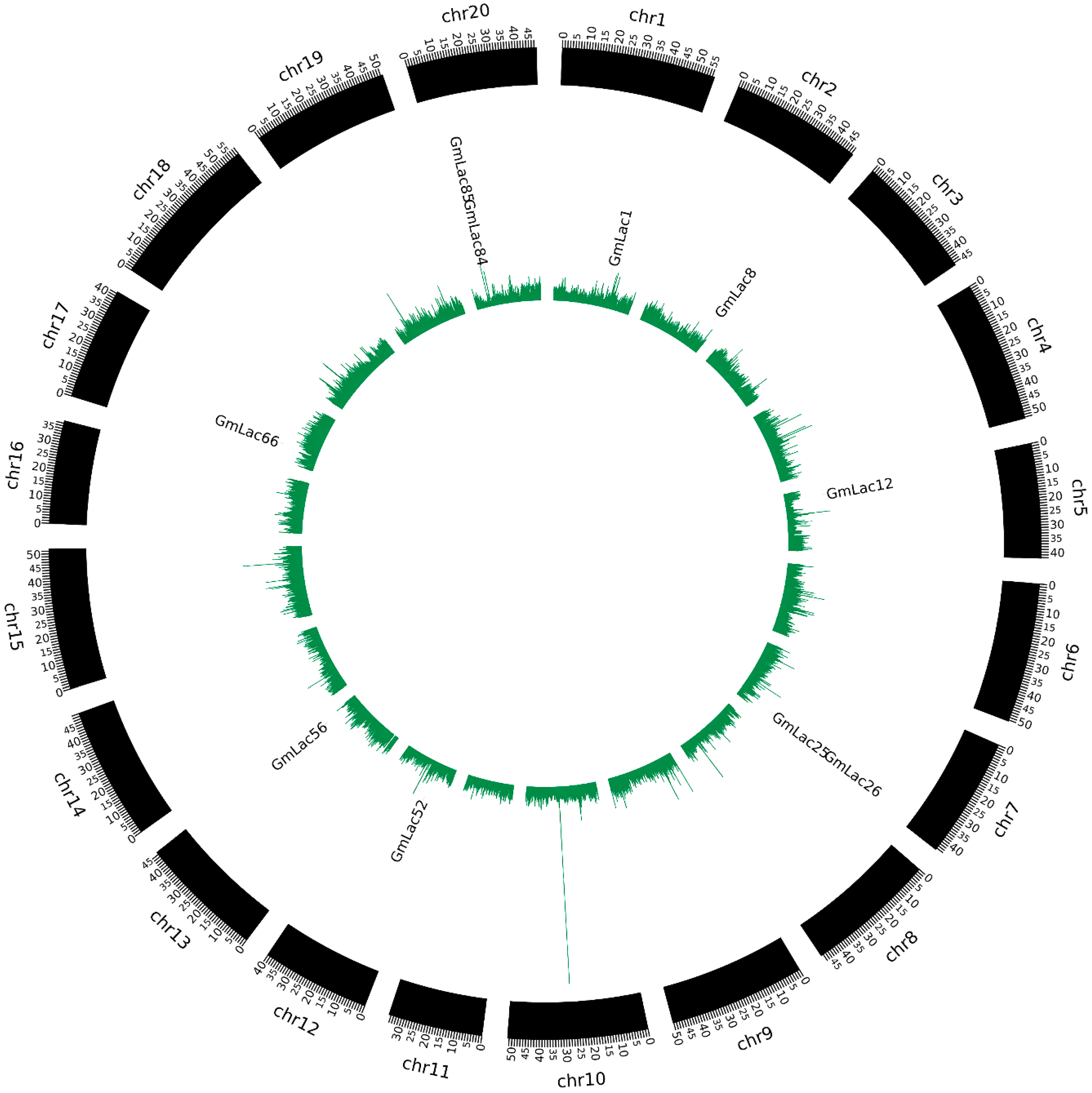
3.3. Duplication and Expansion Events in Soybean Laccase Gene Family
3.4. Gene Structure and Motif Compositions of GmLacs
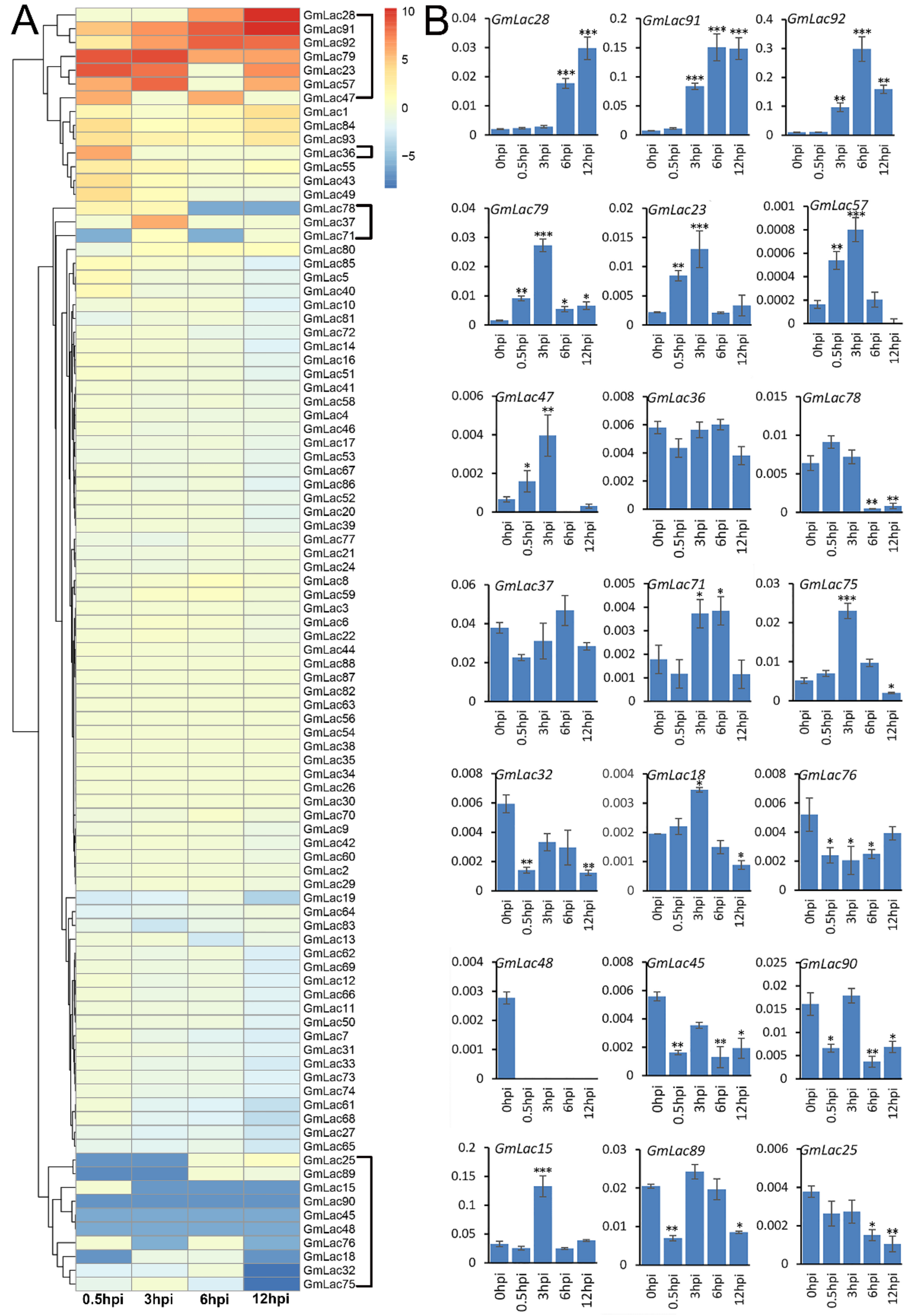
3.5. Expression Patterns of GmLac Genes
3.6. Expression Changes of GmLacs after Inoculation with Phytophthora sojae
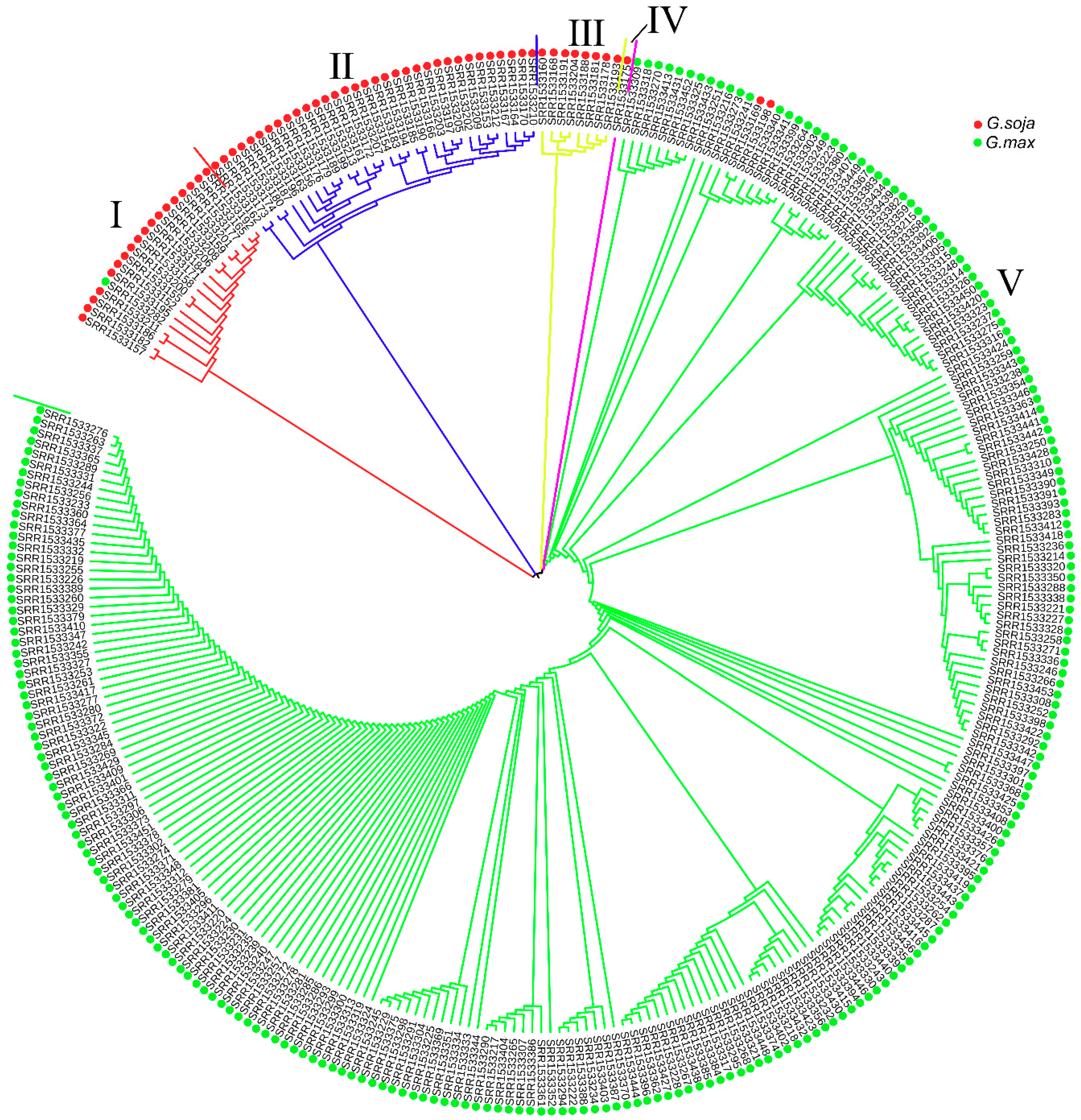
3.7. Selection Pressure Analysis of Laccase Gene Family during Soybean Domestication
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yaropolov, A.I.; Skorobogat, O.V.; Vartanov, S.S.; Varfolomeyev, S.D. Laccase. Appl. Biochem. Biotechnol. 1994, 49, 257–280. [Google Scholar] [CrossRef]
- Alfred, M.M.; Richard, C.S. Laccase: New functions for an old enzyme. Phytochemistry 2002, 60, 551–565. [Google Scholar]
- Cai, X.; Davis, E.J.; Ballif, J.; Liang, M.; Bushman, E.; Haroldsen, V.; Torabinejad, J.; Wu, Y. Mutant identification and characterization of the laccase gene family in Arabidopsis. J. Exp. Bot. 2006, 57, 2563–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandre, G.; Zhulin, I.B. Laccases are widespread in bacteria. Trends Biotechnol. 2000, 18, 41–42. [Google Scholar] [CrossRef]
- Diamantidis, G.; Effosse, A.; Potier, P.; René, B. Purification and characterization of the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biol. Biochem. 2000, 32, 919–927. [Google Scholar] [CrossRef]
- Claus, H. Laccases and their occurrence in prokaryotes. Arch. Microbiol. 2003, 179, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Ausec, L.; Zakrzewski, M.; Goesmann, A.; Schlueter, A.; Mandic-Mulec, I. Bioinformatic analysis reveals high diversity of bacterial genes for laccase-like enzymes. PLoS ONE 2011, 6, e25724. [Google Scholar] [CrossRef]
- Yoshida, H. Chemistry of lacquer (urushi). J. Chem. Soc. 1883, 43, 472–486. [Google Scholar] [CrossRef]
- Bertrand, G. Sur la laccase et sur le pouvoir oxydant de cette diastase. CR Acad. Sci. 1895, 120, 266–269. [Google Scholar]
- Kudanga, T.; Nyanhongo, G.S.; Guebitz, G.; Burton, S. Potential applications of laccase-mediated coupling and grafting reactions: A review. Enzyme Microb. Technol. 2011, 48, 195–208. [Google Scholar] [CrossRef]
- Bento, I.; Carrondo, M.A.; Lindley, P.F. Reduction of dioxygen by enzymes containing copper. J. Biol. Inorg. Chem. 2006, 11, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Shiba, T.; Xiao, L.; Miyakoshi, T.; Chen, C.L. Oxidation of isoeugenol and coniferyl alcohol catalyzed by laccases isolated from rhus vernicifera stokes and pycnoporus coccineus. J. Mol. Catal. B Enzym. 2000, 10, 605–615. [Google Scholar] [CrossRef]
- Rodriguez, C.S.; Toca-Herrera, J.L. Industrial and biotechnological applications of laccases: A review. Biotechnol. Adv. 2006, 24, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Chirivì, C.; Fontana, G.; Monti, D.; Ottolina, G.; Riva, S.; Danieli, B. The quest for new mild and selective modifications of natural structures: Laccase-catalysed oxidation of ergot alkaloids leads to unexpected stereoselective C-4 hydroxylation. Chem. Eur. J. 2012, 18, 10355–10361. [Google Scholar] [CrossRef]
- Claus, H. Laccases: Structure, reaction, distribution. Micron 2004, 35, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.; Shumakovich, G.P.; Gorbacheva, M.; Yaropolov, A.I. “Blue” laccases. Biochemistry 2007, 72, 1136–1150. [Google Scholar] [CrossRef] [PubMed]
- Linares, N.; Fernández, F.; Loske, A.; Gómez-Lim, M.A. Enhanced delignification of lignocellulosic biomass by recombinant fungus phanerochaete chrysosporium overexpressing laccases and peroxidases. J. Mol. Microb. Biotechnol. 2018, 28, 1–13. [Google Scholar]
- Leonowicz, A.; Cho, N.S.; Luterek, J.; Jarosz-Wilkołazka, A.; Wojtas-Wasilewska, M.; Matuszewska, A.; Hofrichter, M.; Dobritzsch, D.; Rogalski, J. Fungal laccase: Properties and activity on lignin. J. Basic Microb. 2001, 41, 185–227. [Google Scholar] [CrossRef]
- Baldrian, P. Fungal laccases—Occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef]
- Hoegger, P.J.; Kilaru, S.; James, T.Y.; Thacker, J.R.; Kües, U. Phylogenetic comparison and classification of laccase and related multicopper oxidase protein sequences. FEBS J. 2006, 273, 2308–2326. [Google Scholar] [CrossRef]
- Wösten, H.A.B.; Wessels, J.G.H. The Emergence of Fruiting Bodies in Basidiomycetes. In Growth, Differentiation and Sexuality; Ursula, K., Reinhard, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 393–414. [Google Scholar]
- Lettera, V.; Piscitelli, A.; Leo, G.; Birolo, L.; Pezzella, C.; Sannia, G. Identification of a new member of pleurotus ostreatus laccase family from mature fruiting body. Fungal Biol. UK 2010, 114, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, H.; Chen, M.; Ren, A.; Huang, J.; Wang, H.; Zhao, M.; Feng, Z. Cloning and functional analysis of a laccase gene during fruiting body formation in Hypsizygus marmoreus. Microbiol. Res. 2015, 179, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Cao, Z.; Cao, K.; Ma, S.; Gong, X.; Jia, H.; Dai, D.; Dong, J. Identification of laccase-like multicopper oxidases from the pathogenic fungus Setosphaeria turcica and their expression pattern during growth and infection. Eur. J. Plant Pathol. 2019, 153, 1149–1163. [Google Scholar] [CrossRef]
- Cloete, M.; Mostert, L.; Fischer, M.; Halleen, F. Pathogenicity of south african hymenochaetales taxa isolated from esca-infected grapevines. Phytopathol. Mediterr. 2015, 54, 368–379. [Google Scholar]
- Temp, U.; Eggert, C. Novel interaction between laccase and cellobiose dehydrogenase during pigment synthesis in the white rot fungus Pycnoporus cinnabarinus. Appl. Environ. Microb. 1999, 65, 389–395. [Google Scholar]
- Díaz, L.; Del Río, J.A.; Pérez-Gilabert, M.; Ortuño, A. Involvement of an extracellular fungus laccase in the flavonoid metabolism in Citrus fruits inoculated with Alternaria alternata. Plant Physiol. Biochem. 2015, 89, 11–17. [Google Scholar] [CrossRef]
- Berthet, S.; Thévenin, J.; Baratiny, D.; Demont-Caulet, N.; Debeaujon, I.; Bidzinski, P.; Leplé, J.-C.; Huis, R.; Hawkins, S.; Gomez, L.-D.; et al. Role of plant laccases in lignin polymerization. In Lignins—Biosynthesis, Biodegradation and Bioengineering; Lise, J., Catherine, L., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2012; Chapter 5; pp. 145–172. [Google Scholar]
- Rodgers, C.J.; Blanford, C.F.; Giddens, S.R.; Skamnioti, P.; Armstrong, F.A.; Gurr, S.J. Designer laccases: A vogue for high-potential fungal enzymes? Trends Biotechnol. 2010, 28, 63–72. [Google Scholar] [CrossRef]
- Meehan, T. The lacquer tree. Proc. Acad. Natl. Sci. Phila. 2014, 29, 279. [Google Scholar]
- Nitta, K.; Kataoka, K.; Sakurai, T. Primary structure of a Japanese lacquer tree laccase as a prototype enzyme of multicopper oxidases. J. Inorg. Biochem. 2002, 91, 125–131. [Google Scholar] [CrossRef]
- Berthet, S.; Demontcaulet, N.; Pollet, B.; Bidzinski, P.; Cézard, L.; Bris, P.L.; Borrega, N.; Hervé, J.; Blondet, E.; Balzergue, S. Disruption of Laccase4 and 17 results in tissue-specific alterations to lignification of arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef]
- Sedbrook, J.C.; Carroll, K.L.; Kai, F.H.; Masson, P.H.; Somerville, C.R. The arabidopsis SKU5 gene encodes an extracellular glycosyl phosphatidylinositol-anchored glycoprotein involved in directional root growth. Plant Cell 2002, 14, 1635–1648. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.R.L.; Ferrar, P.H. Diphenol oxidases, enzyme-catalysed browning and plant disease resistance. Biotechnol. Genet. Eng. 1998, 15, 457–498. [Google Scholar] [CrossRef]
- Iqbal, M.J.; Ahsan, R.; Afzal, A.J.; Jamai, A.; Meksem, K.; Elshemy, H.A.; Lightfoot, D.A.; Elshemy, H.A. Multigeneic QTL: The laccase encoded within the soybean Rfs2/rhg1 locus inferred to underlie part of the dual resistance to cyst nematode and sudden death syndrome. Curr. Issues Mol. Biol. 2009, 11 (Suppl. S1), 11. [Google Scholar]
- Bao, W.; O’Malley, D.M.; Whetten, R.; Sederoff, R.R. A laccase associated with lignification in loblolly pine xylem. Science 1993, 260, 672–674. [Google Scholar] [CrossRef] [PubMed]
- Sterjiades, R.; Dean, J.F.; Eriksson, K.E. Laccase from sycamore maple (acer pseudoplatanus) polymerizes monolignols. Plant Physiol. 1992, 99, 1162–1168. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, C.; Zhu, M.; Yu, Y.; Zhang, Y.; Wei, Z. Generation and characterization of transgenic poplar plant overexpressing a cotton laccase gene. Plant Cell Tiss. Org. 2008, 93, 303–310. [Google Scholar] [CrossRef]
- Sharma, P.; Goel, R.; Capalash, N. Bacterial Laccases. World J. Microb. Biot. 2007, 23, 823–832. [Google Scholar] [CrossRef]
- Hullo, M.F.; Moszer, I.; Danchin, A.; Martin, V.I. CotA of bacillus subtilis Is a copper-dependent laccase. J. Bacteriol. 2001, 183, 5426–5430. [Google Scholar] [CrossRef]
- Fang, Z.; Li, T.; Wang, Q.; Zhang, X.; Peng, H.; Fang, W.; Hong, Y.; Ge, H.; Xiao, Y. A bacterial laccase from marine microbial metagenome exhibiting chloride tolerance and dye decolorization ability. Appl. Microbiol. Biot. 2011, 89, 1103–1110. [Google Scholar] [CrossRef]
- Wang, J.; Feng, J.; Jia, W.; Fan, P.; Bao, H.; Li, S.; Li, Y. Genome-wide identification of sorghum bicolor laccases reveals potential targets for lignin modification. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Wang, X.; Shen, Z.; Zheng, L. Comprehensive analysis of rice laccase gene (OsLAC) family and ectopic expression of OsLAC10 enhances tolerance to copper stress in arabidopsis. Int. J. Mol. Sci. 2017, 18, 209. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.K.; Rai, K.M.; Thu, S.W.; Hii, M.M.; Mendu, V. Genome-wide identification of multifunctional laccase gene family in cotton (Gossypium spp.); Expression and biochemical analysis during fiber development. Sci. Rep. UK 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; Humana Press: Totowa, NJ, USA, 2005. [Google Scholar]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Krogh, A.; Sonnhammer, E.L. Advantages of combined transmembrane topology and signal peptide prediction—The Phobius web server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, msw054. [Google Scholar] [CrossRef] [PubMed]
- Ivica, L.; Peer, B. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar]
- Wang, Y.; Li, J.; Paterson, A.H. MCScanX-transposed: Detecting transposed gene duplications based on multiple colinearity scans. Bioinformatics 2013, 29, 1458–1460. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating γ-series methods and sliding window strategies. Genom. Proteom. Bioinf. 2010, 8, 77–80. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef]
- Guo, A.; Zhu, Q.H.; Chen, X.; Luo, J.C. GSDS: A gene structure display server. Yi chuan = Hereditas/Zhongguo yi chuan xue hui bian ji 2007, 29, 1023–1026. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Farmer, A.; Joshi, T.; Takahashi, K.; Langley, R.J.; Franklin, L.D.; He, J.; Xu, D.; May, G.; Stacey, G. An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants. Plant J. 2010, 63, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Jiang, Y.; Wang, Z.; Gou, Z.; Lyu, J.; Li, W.; Yu, Y.; Shu, L.; Zhao, Y.; Ma, Y.; et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 2015, 33, 408. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Sun, L.; Li, S.; Wang, W.; Ding, Y.; Swarm, S.A.; Li, L.; Wang, X.; Tang, X.; Zhang, Z.; et al. Elevation of soybean seed oil content through selection for seed coat shininess. Nat. Plants 2018, 4, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, W.; Fang, C.; Xu, F.; Liu, Y.; Wang, Z.; Yang, R.; Zhang, M.; Liu, S.; Lu, S.; et al. Parallel selection on a dormancy gene during domestication of crops from multiple families. Nat. Genet. 2018, 50, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- McCaig, B.C.; Meagher, R.B.; Dean, J.F. Gene structure and molecular analysis of the laccase-like multicopper oxidase (LMCO) gene family in Arabidopsis thaliana. Planta 2005, 221, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of gene duplication. Proc. Natl. Acad. Sci. USA 2004, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed]
- Lavin, M.; Herendeen, P.; Wojciechowski, M. Evolutionary rates analysis of leguminosae implicates a rapid diversification of lineages during the tertiary. Syst. Biol. 2005, 54, 575–594. [Google Scholar] [CrossRef] [PubMed]
- Pfeil, B.E.; Schlueter, J.A.; Shoemaker, R.C.; Doyle, J.J. Placing paleopolyploidy in relation to taxon divergence: A phylogenetic analysis in legumes using 39 gene families. Syst. Biol. 2005, 54, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, X.; Hu, R.; Wang, Y.; Xiao, C.; Jiang, Y.; Zhang, X.; Zheng, C.; Fu, Y.F. The pattern of phosphate transporter 1 genes evolutionary divergence in Glycine max L. BMC Plant Biol. 2013, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.A.; Lin, J.Y.; Schlueter, S.D.; Vasylenko-Sanders, I.F.; Deshpande, S.; Yi, J.; O’Bleness, M.; Roe, B.A.; Nelson, R.L.; Scheffler, B.E.; et al. Gene duplication and paleopolyploidy in soybean and the implications for whole genome sequencing. BMC Genom. 2007, 8, 330. [Google Scholar] [CrossRef] [PubMed]
- Dai, A.H.; Yang, S.X.; Zhou, H.K.; Tang, K.Q.; Li, G.; Leng, J.T.; Yu, H.; Zhang, Y.H.; Gao, J.S.; Yang, X.; et al. Evolution and expression divergence of the CYP78A subfamily genes in soybean. Genes 2018, 9, 611. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Landherr, L.; Frohlich, M.; Leebens-Mack, J.; Ma, H.; dePamphilis, C.W. Patterns of gene duplication in the plant SKP1 gene family in angiosperms: Evidence for multiple mechanisms of rapid gene birth. Plant J. 2007, 50, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4. [Google Scholar] [CrossRef] [PubMed]
- Van de Peer, Y.; Maere, S.; Meyer, A. The evolutionary significance of ancient genome duplications. Nat. Rev. Genet. 2009, 10, 725–732. [Google Scholar] [CrossRef] [Green Version]
- Giannuzzi, G.; D’Addabbo, P.; Gasparro, M.; Martinelli, M.; Carelli, F.; Antonacci, D.; Ventura, M. Analysis of high-identity segmental duplications in the grapevine genome. BMC Genom. 2011, 12. [Google Scholar] [CrossRef]
- Lupski, J.R.; Stankiewicz, M.D. Genomic Disorders-The Genomic Basis of Disease; Humana Press: New York, NY, USA, 2006; pp. 73–88. [Google Scholar]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.A.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 465, 120. [Google Scholar] [CrossRef]
- Hu, Q.; Min, L.; Yang, X.; Jin, S.; Zhang, L.; Li, Y.; Ma, Y.; Qi, X.; Li, D.; Liu, H.Y.; et al. Laccase GhLac1 modulates broad-spectrum biotic stress tolerance via manipulating phenylpropanoid pathway and jasmonic acid synthesis. Plant Physiol. 2018, 176, 1808–1823. [Google Scholar] [CrossRef]
- Zhang, S.; Ju, C.L.; Wang, X. Arabidopsis laccase Gene AtLAC4 regulates plant growth and responses to abiotic stress. CBB 2013, 47, 357–365. [Google Scholar]
- Gijzen, M.; Qutob, D. Phytophthora Sojae and Soybean. In Oomycete Genetics and Genomics: Diversity, Interactions, and Research Tools; Kurt, L.S., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 303–329. [Google Scholar]
- Forster, H. Phytophthora sojae Races have arisen by clonal evolution and by rare outcrosses. Mol. Plant Microbe Interact. 1994, 7, 780. [Google Scholar] [CrossRef]
- Dashiell, K. SOYBEANS: Improvement, Production, and Uses, 3rd ed.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2005; pp. 110–111. [Google Scholar]
- Zohary, D.; Hopf, M. Domestication of plants in the old world. J. Appl. Ecol. 1993, 130, 365–372. [Google Scholar]
- Liu, B.; Fujita, T.; Yan, Z.; Sakamoto, S.; Xu, D.J. QTL mapping of domestication-related traits in soybean (Glycine max). Ann. Bot. Lond. 2007, 100, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Li, P.; Cai, D.; Chen, Q.; Qin, P.; Tan, T.; Cao, H. Comparison of performances of corn fiber plastic composites made from different parts of corn stalk. Ind. Crop Prod. 2016, 95, 521–527. [Google Scholar] [CrossRef]
- Dean, J.F.; Eriksson, K.E. Laccase and the deposition of lignin in vascular plants. Holzforschung 1994, 48, 21–33. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. Transparent testa10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [PubMed]
- Ranocha, P.; Chabannes, M.; Chamayou, S.; Danoun, S.; Jauneau, A.; Boudet, A.M.; Goffner, D. Laccase down-regulation causes alterations in phenolic metabolism and cell wall structure in poplar. Plant Physiol. 2002, 129, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Nakashima, J.; Chen, F.; Yin, Y.; Fu, C.; Yun, J.; Shao, H.; Wang, X.; Wang, Z.Y.; Dixon, R. Laccase is necessary and nonredundant with peioxidase for lignin polymerization during vascular development in Arabidopsis. Plant Cell 2013, 25, 3976–3987. [Google Scholar] [CrossRef]
- Wang, C.Y.; Zhang, S.; Yu, Y.; Luo, Y.C.; Liu, Q.; Ju, C.; Zhang, Y.C.; Qu, L.H.; Lucas, W.J.; Wang, X.; et al. MiR397b regulates both lignin content and seed number in Arabidopsis via modulating a laccase involved in lignin biosynthesis. Plant Biotechnol. J. 2014, 12, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bouchabké-Coussa, O.; Lenris, P.; Antelme, S.; Soulhat, C.; Gineau, E.; Dalmais, M.; Bendahmane, A.; Morin, H.; Mouille, G.; et al. Lassase 5 is required for lignification of the brachypodium distachyon culm. Plant Physiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Li, Q.; Wei, H.; Chang, M.J.; Tunlaya-Anukit, S.; Kim, H.; Liu, J.; Song, J.; Sun, Y.H.; Yuan, L.; et al. Ptr-miR397a is a negative regulator of laccase genes affecting lignin content in Populus trichocarpa. Proc. Natl. Acad. Sci. USA 2013, 110, 10848–10853. [Google Scholar] [CrossRef] [PubMed]
- The Arabidopsis Genome, I. Analysis of the genome sequence of the flowering plant arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [PubMed]
- The International Brachypodium Initiative; Vogel, J.P.; Garvin, D.F.; Mockler, T.C.; Schmutz, J.; Rokhsar, D.; Bevan, M.W.; Barry, K.; Lucas, S.; Harmon-Smith, M.; et al. Genome sequencing and analysis of the model grass brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the oryza sativa nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]
- Wang, K.; Wang, Z.; Li, F.; Ye, W.; Wang, J.; Song, G.; Yue, Z.; Cong, L.; Shang, H.; Zhu, S.; et al. The draft genome of a diploid cotton Gossypium raimondii. Nat. Genet. 2012, 44, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fan, G.; Wang, K.; Sun, F.; Yuan, Y.; Song, G.; Li, Q.; Ma, Z.; Lu, C.H.; Zou, C.; et al. Genome sequence of the cultivated cotton Gossypium arboreum. Nat. Genet. 2014, 46, 567–574. [Google Scholar] [CrossRef]
- Ngaki, M.N.; Wang, B.; Sahu, B.; Srivastava, S.; Farooqi, M.; Sekhar, K.; Swaminathan, S.; Bhattacharyya, M.K. Tanscriptomic study of the soybean-fusarium virguliforme interaction revealed a novel ankyrin-repeat containing defense gene, expression of whose during infection led to enhanced resistance to the fungal pathogen in transgenic soybean plants. PLoS ONE 2016, 11, e0163106. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Root | RH | Nodules | Pod | Seed | Stem | SAM | Leaves | Flower | Scale Bar |
|---|---|---|---|---|---|---|---|---|---|---|---|
| GmLac5 | Glyma.01G183100 | 29.87 | 66.04 | 33.99 | 37.23 | 14.55 | 46.41 | 24.14 | 2.00 | 6.78 | 0.00 |
| GmLac10 | Glyma.04G019500 | 11.59 | 44.89 | 11.61 | 73.40 | 111.75 | 53.38 | 23.71 | 2.26 | 11.45 | 5.00 |
| GmLac12 | Glyma.05G056100 | 54.73 | 65.77 | 60.52 | 23.36 | 22.16 | 47.63 | 55.08 | 6.07 | 5.52 | 10.00 |
| GmLac16 | Glyma.06G307100 | 36.50 | 49.47 | 10.29 | 129.80 | 4.81 | 99.05 | 114.40 | 14.28 | 18.43 | 15.00 |
| GmLac40 | Glyma.11G059200 | 33.53 | 59.28 | 40.51 | 33.92 | 10.26 | 87.08 | 43.94 | 6.10 | 9.01 | 20.00 |
| GmLac55 | Glyma.13G076900 | 162.50 | 50.75 | 30.23 | 12.38 | 3.09 | 36.75 | 8.77 | 12.19 | 4.63 | 25.00 |
| GmLac62 | Glyma.14G223000 | 27.32 | 53.26 | 32.22 | 21.12 | 15.68 | 33.04 | 8.42 | 1.00 | 2.10 | 30.00 |
| GmLac65 | Glyma.17G012300 | 69.51 | 127.58 | 54.73 | 26.11 | 33.73 | 26.31 | 62.91 | 1.39 | 4.16 | |
| GmLac66 | Glyma.17G138300 | 66.83 | 56.05 | 48.36 | 56.52 | 35.88 | 84.13 | 43.27 | 8.40 | 9.79 | |
| GmLac69 | Glyma.17G261500 | 47.09 | 65.88 | 29.72 | 2.73 | 0.36 | 28.53 | 16.04 | 3.85 | 4.14 | |
| GmLac85 | Glyma.20G051900 | 93.65 | 67.15 | 30.28 | 32.47 | 44.21 | 48.51 | 13.49 | 12.04 | 2.55 |
| Gene Name | Gene ID | Root | RH | Nodules | Pod | Seed | Stem | SAM | Leaves | Flower | Scale Bar |
|---|---|---|---|---|---|---|---|---|---|---|---|
| GmLac1 | Glyma.01G108200 | 41.64 | 4.94 | 6.82 | 0.06 | 0.00 | 0.04 | 0.01 | 0.03 | 0.14 | 0.00 |
| GmLac2 | Glyma.01G112600 | 0.94 | 6.30 | 12.58 | 10.45 | 0.45 | 70.53 | 3.31 | 5.93 | 1.92 | 5.00 |
| GmLac8 | Glyma.02G261600 | 2.95 | 4.36 | 3.94 | 7.97 | 0.69 | 32.58 | 0.56 | 0.98 | 1.91 | 10.00 |
| GmLac9 | Glyma.03G077900 | 1.00 | 7.28 | 15.93 | 10.78 | 0.61 | 75.27 | 3.25 | 5.62 | 2.18 | 15.00 |
| GmLac24 | Glyma.07G142600 | 38.53 | 11.66 | 21.13 | 0.08 | 0.00 | 0.03 | 0.09 | 2.86 | 0.21 | 20.00 |
| GmLac25 | Glyma.07G225300 | 0.20 | 0.00 | 0.00 | 0.01 | 0.00 | 1.06 | 0.20 | 0.03 | 505.03 | 25.00 |
| GmLac77 | Glyma.18G183700 | 0.16 | 1.65 | 2.13 | 35.66 | 0.05 | 16.90 | 0.30 | 1.16 | 0.32 | 30.00 |
| GmLac87 | Glyma.20G172600 | 13.60 | 0.03 | 0.00 | 0.00 | 0.01 | 2.87 | 117.91 | 0.00 | 2.54 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Li, G.; Zheng, K.; Zhu, X.; Ma, J.; Wang, D.; Tang, K.; Feng, X.; Leng, J.; Yu, H.; et al. The Soybean Laccase Gene Family: Evolution and Possible Roles in Plant Defense and Stem Strength Selection. Genes 2019, 10, 701. https://doi.org/10.3390/genes10090701
Wang Q, Li G, Zheng K, Zhu X, Ma J, Wang D, Tang K, Feng X, Leng J, Yu H, et al. The Soybean Laccase Gene Family: Evolution and Possible Roles in Plant Defense and Stem Strength Selection. Genes. 2019; 10(9):701. https://doi.org/10.3390/genes10090701
Chicago/Turabian StyleWang, Quan, Guang Li, Kaijie Zheng, Xiaobin Zhu, Jingjing Ma, Dongmei Wang, Kuanqiang Tang, Xingxing Feng, Jiantian Leng, Hui Yu, and et al. 2019. "The Soybean Laccase Gene Family: Evolution and Possible Roles in Plant Defense and Stem Strength Selection" Genes 10, no. 9: 701. https://doi.org/10.3390/genes10090701