m6A mRNA Destiny: Chained to the rhYTHm by the YTH-Containing Proteins

Laboratoire de Biochimie, Ecole polytechnique, CNRS, Université Paris-Saclay, 91128 Palaiseau CEDEX, France
*
Author to whom correspondence should be addressed.
Genes 2019, 10(1), 49; https://doi.org/10.3390/genes10010049
Submission received: 10 December 2018
/
Revised: 8 January 2019
/
Accepted: 10 January 2019
/
Published: 15 January 2019
(This article belongs to the Special Issue RNA Modifications)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The control of gene expression is a multi-layered process occurring at the level of DNA, RNA, and proteins. With the emergence of highly sensitive techniques, new aspects of RNA regulation have been uncovered leading to the emerging field of epitranscriptomics dealing with RNA modifications. Among those post-transcriptional modifications, N6-methyladenosine (m6A) is the most prevalent in messenger RNAs (mRNAs). This mark can either prevent or stimulate the formation of RNA-protein complexes, thereby influencing mRNA-related mechanisms and cellular processes. This review focuses on proteins containing a YTH domain (for YT521-B Homology), a small building block, that selectively detects the m6A nucleotide embedded within a consensus motif. Thereby, it contributes to the recruitment of various effectors involved in the control of mRNA fates through adjacent regions present in the different YTH-containing proteins.
1. Introduction
The regulation of gene expression plays a central role during development and upon cell response to stress exposure. Hence, living organisms have developed highly complex mechanisms at different steps of gene expression to tune various cellular pathways. Many of those events occur at the post-transcriptional level through the formation of protein-RNA complexes that will influence various aspects of messenger RNA (mRNA) maturation such as alternative splicing, editing, export, and polyadenylation. Therefore, RNA binding domains (RBDs) are key actors in these regulatory mechanisms through their recognition of specific RNA sequences or structures. The most common RBDs are the RNA recognition motif (RRM) (corresponding to about 2% of all RNA binding domains), the hnRNP K Homology domains (KH), Piwi Argonaute and Zwille domains (PAZ), and double stranded RNA-binding domains (dsRBD) [1,2,3,4]. With recent technological developments combining UV cross-linking together with oligo(dT) purification of mRNAs, we now have comprehensive lists of RNA-binding proteins in various eukaryotic organisms [5,6,7,8], indicating that many other RBDs are yet to be uncovered.
The recent identification of several internal post-transcriptional modifications such as m6A (N6-methyladenosine), m1A (N1-methyladenosine), pseudouridine (Ψ), m5C (5-methylcytosine), and ac4C (N4-acetylcytosine) within mRNAs has shed light onto an additional layer of regulation now known as epitranscriptomics [9,10,11,12,13,14,15,16,17,18]. Indeed, similar to the dynamic modifications known to occur in DNA and proteins, epitranscriptomics modifications widely contribute to the regulation of biological pathways [19,20]. At present, the most studied mRNA modification is m6A. This modification was initially identified in mRNAs four decades ago [9]. However, because m6A does not alter base pairing [21], and hence does not result in the introduction of a stall or a mutation during reverse-transcription, the development of advanced techniques to precisely map m6A sites has been a main obstacle for studying its biological significance. This field was reignited in 2011, following the discovery of the m6A demethylase FTO (fat mass and obesity-associated protein), a protein involved in human obesity, as a so-called ’eraser‘ enzyme that removes m6As present on mRNAs [22]. More recently, several laboratories have finally been successful in mapping m6A at individual-nucleotide resolution using cross-linking and immuno-precipitation with m6A-specific antibodies. High-throughput sequencing of the immuno-precipitated RNA fragments revealed the presence of more than 10,000 m6A sites in human cells, affecting more than 25% of the transcriptome. Their detailed mapping showed an enrichment of m6A near the stop codon and in the 3′ untranslated region (3′ UTR) of the target mRNAs. The main ‘writer’ methyltransferase is a multi-protein complex composed of at least METTL3, METTL14, WTAP, and KIAA1429, which is responsible for m6A deposition on the consensus motif DRA*CH (where D is A, G or U; R is A, or G; A* is the methylated A and H is A, C ,or U; [10,13,23,24]). The m6A marks can be deleted by ‘erasers’ such as FTO and ALKBH5 [22,25]. Furthermore, this modification can attract m6A-binding proteins known as ‘readers’, as well as repel various proteins regulating mRNA functions [10,26,27,28]. This differential recruitment of regulatory proteins on m6A marks subsequently determines the fate of m6A-containing mRNAs, such as splicing, translation, degradation, or cellular localization. The most studied RNA binding module known to directly recognize m6A marks is the YTH domain. Indeed, many studies are scrutinizing eukaryotic YTH-containing proteins in order to clarify their roles in the regulation of mRNA fates. The present review aims at summarizing our current knowledge on this class of m6A readers.
2. The YTH Domain, an m6A RNA Grip
The foremost member of this m6A reader protein family is human YT521-B (hereafter termed YTHDC1), which was initially identified as a factor interacting with Tra2β, SC35, SF2, hnRNP G, and SAM68 splicing factors [29,30]. Despite this clear interaction of YTHDC1 with splicing machinery, no known RNA binding domain was detected and the only common feature with splicing factors was the presence of repeats of charged amino acids [30]. Further bioinformatics analyses led to the identification of an additional conserved region, named YTH (for YT521-B Homology) domain. This domain was found exclusively in eukaryotic proteins from fungi (one member in Saccharomyces cerevisiae and Schizosaccharomyces pombe yeasts) through plants (13 members in Arabidopsis thaliana) to higher eukaryotes (five members in human, namely YTHDC 1–2 and YTHDF 1–3) [31,32]. Although this domain was predicted to be a putative RNA binding domain since its identification, its RNA binding property was only demonstrated in 2010 when Zhang et al. showed that YTHDC1 can bind degenerate RNA sequences [33]. YTH-containing proteins became a topic of strong interest when YTHDF1, 2, and 3, three paralogous members of the YTH-containing protein family, were found to be the most enriched human proteins specifically retained by an RNA fragment containing an m6A modification at the heart of the DRA*CH consensus sequence [10,26].
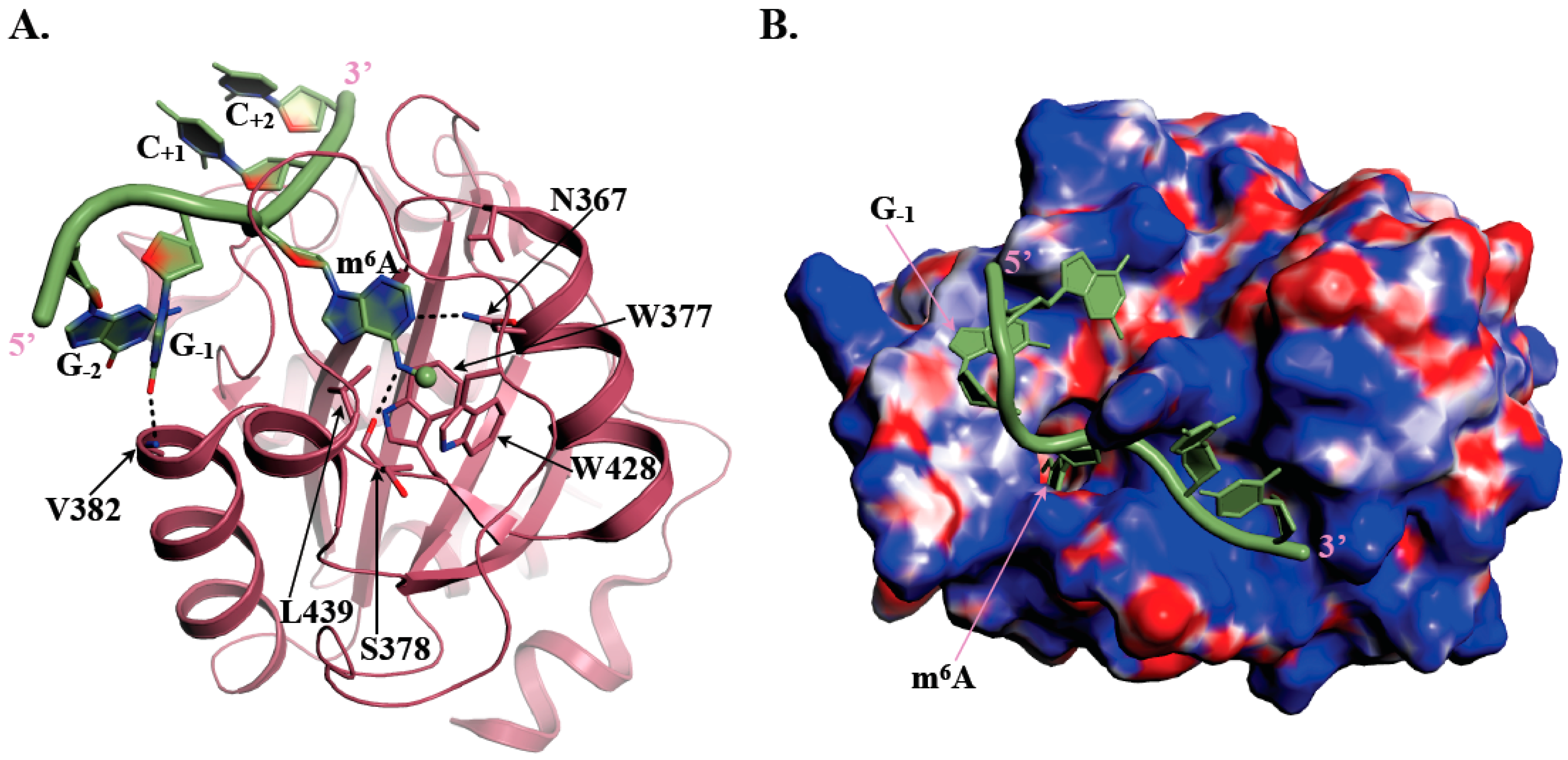
The YTH domain is made of 150 to 200 residues and adopts an α/β fold, with four to five α-helices surrounding a curved six stranded β-sheet as revealed by the NMR structure of YTHDC1 YTH domain determined by the RIKEN Structural Genomics and Proteomics Initiative in 2007 (PDB code: 2YUD). At the center of the β-sheet lies a cavity delineated by conserved hydrophobic residues including three tryptophan amino acid side chains (in some YTH domains, one Trp residue is substituted by either Leu or Tyr) forming a so-called aromatic cage (Figure 1A). Several 3D-structures of different YTH-m6A containing RNA complexes have revealed that this aromatic cage is responsible for the specific recognition of the m6A mark by YTH domains [34,35,36,37,38]. Indeed, in the case of human YTHDC1 bound to an m6A containing RNA, the aromatic cage specifically accommodates m6A via the side chains from W377, W428, and L439 amino acids [37]. The m6A methyl group forms methyl-π interaction with W428 side chain while the purine base is sandwiched between the W377 and L439 side chains (Figure 1A). This interaction of the m6A methyl group with W428 most probably explains the higher affinity (20 to 50-fold difference) of YTH domains for an m6A containing RNA oligonucleotide compared to the same unmodified oligonucleotide [36]. Sequence alignment revealed that the W377 and W428 residues are strictly conserved among the YTH-containing proteins while the position corresponding to L439 can be occupied by either Leu, Tyr, or Trp. The integrity of this aromatic cage is essential for the recognition of m6A since mutation of any of these three residues to alanine disrupts binding [34,37,38]. Polar amino acids (S378, N363, and N367 in human YTHDC1) also contribute to m6A recognition by forming specific hydrogen bonds with nitrogen atoms from the adenine base and then participating together with the aromatic cage to the selective recognition of m6A (Figure 1A). Residues surrounding this aromatic cage are also important for RNA binding as they form a large positively-charged surface interacting with RNA phosphate groups from nucleotides surrounding m6A and also provide some specific interactions with bases. A large majority of m6A marks generated by the main m6A methyltransferase ‘writer’ is found within Gm6AC (70%) or Am6AC (30%) motifs [37]. Interestingly, structural analyses coupled to isothermal titration calorimetry (ITC) measurements using various RNA sequences have shown that the affinity of YTH domain from YTHDC1 for RNA fragments is higher (five- to six-fold difference) when the nucleotide immediately upstream of m6A is a G compared to an A [37,38], suggesting the co-evolution of both the active site of METTL3-METTL14 methyltransferase holoenzyme and of the RNA binding site of human YTHDC1. Indeed, in the structure of YTHDC1 bound to a GGm6ACU RNA fragment [37], the carbonyl group at position six of the G base preceding m6A forms a hydrogen bond with the main chain nitrogen group from V382 (or equivalent position in other YTH domains; Figure 1A). The amine group, present at the same position in A, is less prone to form such hydrogen bond, which could rationalize the differences in measured affinities. However, this seems to be specific of YTHDC1 as all other tested YTH-containing proteins (YTHDF1/2, YTHDC2, and S. cerevisiae Pho92) exhibit similar affinities for RNAs containing any of the four nucleotides at the position immediately upstream m6A [38]. This probably results from the fact that this nucleotide binds into different pockets with no obvious base specificity on YTHDF1 or the Zygosaccharomyces rouxii Mrb1/Pho92 fungal orthologue compared to YTHDC1 [35,38]. So far, structural studies have been performed only with 5- or 7-mers RNAs, which are known to bind more weakly (Kd values in the 2 to 30 µM range) than longer RNAs (9 or 16-mers for instance; Kd values ranging from 0.2 to 0.3 µM for YTHDC1 up to 7.5 µM for YTHDC2 [38]). Therefore, additional structural studies with longer RNA fragments might be of interest to bring more information on RNA recognition by these YTH motifs.
In summary, YTH-domain containing proteins interact with single-stranded RNAs and selectively identify the presence of a modified m6A nucleotide at the center of a consensus signature motif matching that of the major m6A methyltransferase machinery identified to date.
3. YTH, a Building Block Governing the Fates of m6A Containing mRNAs
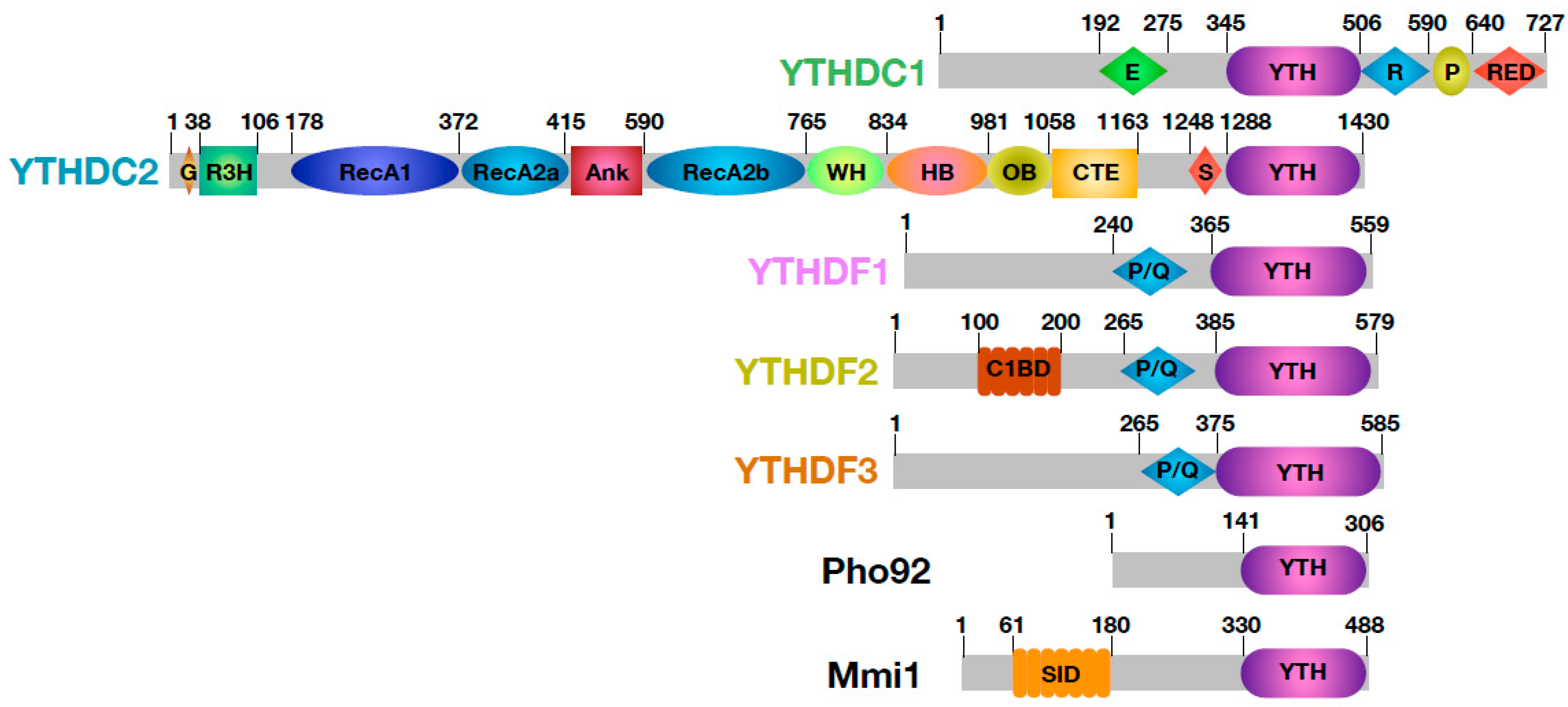
Most RNA binding modules are embedded within larger proteins and are surrounded by either structured domains or low complexity regions, all with various functions [8,40,41]. The YTH domain is no exception to this rule and clearly serves as a building block mostly flanked by regions predicted to be disordered (Figure 2; [42,43,44]). By influencing both the sub-cellular localization of these YTH proteins and their partners, these flanking regions are thereby important for the function of YTH proteins in the regulation of m6A-containing RNA fates, i.e., splicing, mRNA nucleocytoplasmic export, translation and mRNA decay [26,35,45,46,47].
As mentioned above, bioinformatics analyses have identified 13 YTH-containing proteins in A. thaliana, which roles are being clarified. As this topic has been nicely discussed by Bhat et al. in this special issue [32], these plant YTH proteins will not be discussed in this section. On the basis of amino acid sequences, YTH domain proteins from different vertebrates can be divided into three families: YTHDC1, YTHDC2, and YTHDF1–3. There is no significant sequence similarity between members from those different families outside of the YTH domain, hence YTHDC1, YTHDC2, and YTHDF1–3 members cannot be considered as paralogues.
3.1. YTHDC1
YTHDC1 (previously known as YT521B) is the founding member of YTH family of proteins. In this protein, the YTH domain is the only region predicted to be folded and is surrounded by regions rich in charged residues (Glu-rich, Arg-rich and Arg-Asp-Glu-rich, or RED segments; Figure 2) or in proline (P-rich). The human protein contains nuclear localization elements and is found to localize in distinct subnuclear bodies, so-called YT bodies, adjacent to nuclear splicing speckles [48]. This nuclear localization is in agreement with its initially described interactions with various splicing factors (i.e., Tra2β, SC35, SF2, hnRNPG, and SAM68 [29,30]). Recent studies have clearly established a role of YTHDC1 in splicing both in human cells and in Drosophila melanogaster [45,49]. Indeed, this protein contributes to alternative splicing by binding to the pre-mRNAs and by influencing the splice site selection [33]. Mechanistically, YTHDC1 directly interacts with SRSF3 and SRSF10, two serine/arginine-rich splicing factors, in a competitive manner (Figure 3). In doing so, it enhances the binding of SRSF3 to targeted pre-mRNAs resulting in exon inclusion while precluding the binding of SRSF10, which is involved in exon skipping. This YTHDC1-mediated recruitment of SRSF3 is clearly dependent on m6A as either METTL3 silencing or a YTHDC1 double mutant (two Trp residues from the aromatic cage of the YTH domain are substituted by Ala) strongly reduce binding of SRSF3 to RNAs in cellulo [45]. As YTHDC1 has a 30-fold higher affinity for SRSF3 over SRSF10, it interacts predominantly with SRSF3 and hence may favor alternative splicing of m6A-containing pre-mRNAs. This interaction between YTHDC1 and SRSF3 is not only involved in alternative splicing but also in the polyadenylation process of the pre-mRNAs through their association with the pre-mRNA 3′ end processing factors CPSF6 [50]. Moreover, YTHDF1 and SRSF3 collaborate with NXF1 to drive efficient export of transcripts subject to m6A control from the nucleus to the cytoplasm (Figure 3; [47]).
YTHDC1 has also been shown to participate in the regulation of the abundance of MAT2A mRNA, which encodes a subunit of one of the methionine adenosyltransferases responsible for the synthesis of S-adenosyl-L-methionine (SAM) cofactor from methionine and ATP [51]. The MAT2A mRNA is one of the well-characterized targets of the recently identified METTL16 m6A RNA methyltransferase, which introduces m6A marks in the 3′ UTR of this mRNA [52], a modification crucial for mouse embryonic development [53]. Depletion of SAM was shown to enhance the removal of a retained intron in MAT2A pre-mRNA leading to induced expression of this mRNA [52] and to reduce m6A levels in MAT2A 3′ UTR [51]. Whether the role of YTHDC1 in this pathway is to favor splicing, nucleocytoplasmic transfer, or recruitment of mRNA decay enzymes remains to be clarified.
In fruit-fly, the YTHDC1 orthologue is also involved together with m6A methylation in the alternative splicing of Sex lethal (Sxl), which encodes a master regulator of sex determination and dosage compensation [49]. Although Sxl is expressed in males and females, the presence of an additional internal exon in males introduces a premature stop codon that results in both the production of a truncated and non-functional Sxl protein and the rapid elimination of the transcript most probably by the nonsense-mediated mRNA decay pathway [54,55]. Inactivation of either the METTL3 subunit of the m6A mRNA methyltransferase holoenzyme or the YTHDC1 protein in the female but not the male fly results in retention of the male-specific exon concomitant with the decrease of the female-specific isoform, clearly indicating a female-specific splicing defect linked to altered m6A deposition and recognition.
Beyond its role in splicing, human YTHDC1 also helps in transcriptional repression of X chromosome genes by X-inactive specific transcript (XIST), a long non-coding RNA that plays a critical role in inactivation of one X chromosome in female cells [56]. XIST is a heavily methylated RNA with at least 78 m6A sites and the preferential binding of YTHDC1 to m6A marks is necessary for XIST-mediated transcriptional silencing. The depletion of m6A ‘writer’ causes inhibition of XIST function and this defect can be restored by artificially tethering YTHDC1 to XIST in cells lacking m6A methylation machinery. A comprehensive understanding of the mode of action of YTHDC1 on XIST will necessitate further studies.
3.2. YTHDC2
Compared to the other YTH domain-containing proteins, where the YTH domain is embedded within low complexity regions, members of the YTHDC2 family are multi-domain proteins (Figure 2). Apart from the C-terminal YTH domain, there is a N-terminal R3H (arginine and histidine-rich) domain with RNA-binding property [57] preceded by a Gly-rich patch, a central DEAH-box helicase domain (where an ankyrin repeat domain is inserted in the middle of the second RecA domain), an helicase associated 2 domain (HA2), an OB-fold (oligonucleotide / oligosaccharide-binding fold) and a C-terminal extension (CTE) also found in human DHX36, a DNA/RNA DEAH-box helicase involved in G-quadruplex unwinding (Figure 2; [58]). In agreement with its domain composition, human YTHDC2 has RNA dependent ATPase and 3′→5′ RNA helicase activities [59,60,61]. This protein is mainly a diffuse cytoplasmic protein, but it is also enriched in peri-nuclear regions [57]. As expected from the presence of several RNA binding domains, YTHDC2 interacts with mRNAs and in particular with m6A-rich mRNAs through its YTH domain [60,62].
Initially, YTHDC2 was shown to associate with hepatitis C virus protein NS5B to facilitate viral DNA replication [59] and to play an important role in the proliferation of cancer cells by enhancing the translation of metastasis-related genes [63,64]. More recent studies have converged towards an important role of YTHDC2 in the progression of meiotic prophase I, which is a critical and long meiosis stage characterized by many chromosomal events that will ultimately lead to severing of the genome into two halves [60,65]. Consequently, the inactivation of YTHDC2 gene in mice results in gametogenesis defects and infertility [60,61,62,66].
In human cells, YTHDC2 interacts in an RNA-independent manner with the meiosis-specific MEIOC protein as well as with the 5′→3′ exonuclease XRN1 [57,60,62,65,67]. Whether YTHDC2 can interact simultaneously with both MEIOC and XRN1 is unclear but it is tempting to speculate that the MEIOC–YTHDC2 complex interacts with m6A-enriched mRNAs to address them to degradation by the XRN1 exonuclease. However, this model could be restricted to a subclass of mRNAs as various studies have observed different effects on mRNA translation and stability upon inactivation of YTHDC2 gene. Indeed, YTHDC2–MEIOC complex could stabilize meiosis-specific transcripts [67] while destabilizing mitotic mRNAs [60,62,65].
Conversely, YTHDC2 has also been shown to enhance translation efficiency of mRNAs concomitantly to decrease their stability (Figure 3; [62]). This could result from the association of YTHDC2 with the head of the 40S ribosomal subunit both at the level of the 40S but also of the 80S [57]. More precisely, YTHDC2 binding site on the 40S subunit maps in the vicinity of the mRNA entry and exit sites, which could rationalize the dual role of YTHDC2 in enhancing translation efficiency by recruiting m6A-containing mRNAs to the ribosome but also decreasing mRNA stability by recruiting XRN1 to those mRNAs (Figure 3).
3.3. YTHDF Family
Human YTHDF1–3 are cytoplasmic proteins made of a single C-terminal YTH-domain that binds to m6A marks separated from a N-terminal low-complexity domain by a segment rich in Pro, Gln, and Asn amino acids (Figure 2; [26,34,38]). Those three proteins are highly homologous with 65 to 68% and about 85% of sequence identity and similarity, respectively.
YTHDF2 was the first protein to be functionally characterized, especially regarding the repertoire of mRNAs that it binds to, as well as its mode of action. Photo-activable ribonucleoside crosslinking and immunoprecipitation (PAR-CLIP) combined with RNA-immunoprecipitation coupled to sequencing (RIP-Seq) experiments showed that human YTHDF2 selectively targets more than 3000 different transcripts and binds predominantly to their 3′ UTR and around the stop codon [26]. Furthermore, YTHDF2 knock-down results in the accumulation of its targets in translatable or actively translating polysome pools, pointing to a crucial role of YTHDF2 in the translation repression of its targets. This defect in translation is also accompanied by an increase in the global abundance of m6A-modified mRNAs, confirming the intimate link existing between the number of m6A sites and the instability of the targeted mRNA [26]. This activity of YTHDF2 in mRNA destabilization requires both its N- and C-terminal regions as over-expression of full-length YTHDF2 leads to decay of m6A containing mRNAs, while expression of only the N-terminal or the C-terminal region does not have the same effect [26]. This role of YTHDF2 in the degradation of m6A-containing mRNAs is further supported by its localization in Processing bodies (P-bodies) in which YTHDF2 co-localizes with DCP1a and DDX6 proteins known to be involved in mRNA decapping [26]. YTHDF2 also directly interacts with CNOT1, the scaffolding subunit of the CCR4-NOT mRNA deadenylase [43]. This interaction relies on the SH domain from CNOT1 and the YTHDF2 N-terminal domain, which is also responsible for the localization of at least YTHDF2 to P-bodies (Figure 3; [43]). Interestingly, Pho92, the only YTH-containing protein from S. cerevisiae, also interacts with Pop2, another component of the CCR4-NOT complex [68]. The similarities between human YTHDF2 and S. cerevisiae Pho92 are emphasized by (1) the ability of human YTHDF2 gene (but not YTHDC1) to complement for the deletion of PHO92 gene in S. cerevisiae and (2) the role of Pho92 as an enhancer of mRNA decay [68]. Altogether, this suggests that the main role of YTHDF2 is in the regulation of m6A-containing mRNA decay and that this role has been conserved throughout evolution. Interestingly, upon heat shock, YTHDF2 relocalizes to the nucleus and this is accompanied by a specific increase of m6A in the 5′ UTR of stress-inducible mRNAs and an increased ribosome occupancy in their coding region [69]. This could then contribute to the stimulation of translation by the direct recruitment of the translation initiation factor 3 complex (eIF3) to m6A sites located within mRNA 5′ UTRs [70].
Despite the strong sequence similarity with YTHDF2, YTHDF1 knock-down does not affect the m6A/A ratio, indicating that this protein is unlikely to be involved in m6A-containing mRNA decay [42]. On the contrary, YTHDF1 seems to enhance the translation efficiency of a population of transcripts encoded by around 1200 genes, to which it associates in an m6A-dependent manner. This mechanism is likely to occur through the recognition of m6A sites by YTHDF1 within the 3′ UTR of mRNAs on the one hand, as well as with the 40S subunit and components of the eIF3 complex bound in the vicinity of the start codon on the other (Figure 3; [42]). This mechanism differs from the one described above for 5′ UTR m6As that directly recruit eIF3 under stress conditions and that is independent of YTHDF1 [70]. Hence, the effect of YTHDF1 on translation of m6A-mRNAs may be limited to a small subset of RNAs depending on various physiological situations. In the nervous system for example, transcriptome-wide mapping of YTHDF1-binding sites, combined with nascent protein labelling, revealed that YTHDF1 enhances translation of key hippocampal m6A-methylated mRNAs in response to neuronal stimulation, thus contributing to learning and memory [71].
Several observations indicate that YTHDF3 interacts with both YTHDF1 and YTHDF2 proteins and thereby works together with those factors to up-regulate translation (YTHDF1) or enhance degradation of mRNAs (YTHDF2), respectively [46,72]. Indeed, PAR-CLIP coupled to RIP-seq showed that YTHDF3 shares more than half of its targets with YTHDF1 but also YTHDF2, which is not surprising considering the high degree of sequence identity (86% to 89%) between the YTH domains from these three proteins [46,72]. However, human YTHDF3 might be also mobilized independently of YTHDF1 and YTHDF2 under specific conditions. Notably YTHDF3 co-localizes exclusively with the stress granules under oxidative stress, whereas YTHDF1 and YTHDF2 retain their cytoplasmic localization with only marginal presence in stress granules. In this specific context, YTHDF3 selectively recognizes a pool of oxidative stress-induced methylated mRNAs in order to mediate triaging of mRNAs from the translatable pool to stress granules (Figure 3; [73]). Overall, this indicates that all three YTHDF family proteins may participate in a complex regulatory mechanism that results first in a higher translational efficiency of m6A-mRNAs followed by their rapid degradation. This complex interplay between these three YTHDF proteins and interacting proteins involved in these different cellular processes will need to be clarified in the future.
Interestingly, m6A marks are not restricted to cellular RNAs but are also found in viral mRNAs, where they are recognized by the YTHDF1–3 proteins [74,75,76,77,78]. Several recent studies have focused on the roles of m6A and YTHDFs on the regulation of viral infection leading to the description of various mechanisms. For instance, the recognition of m6A marks on viral mRNAs by YTHDFs has been shown to block reverse transcription of Zika virus genome [77]. In the case of Hepatitis C virus, YTHDFs inhibit HCV infection without affecting RNA replication by a mechanism that could be common to most Flaviviridae (Figure 3; [75]). Finally, the role of YTHDFs on HIV-1 infection is not clear, as Tirumuru et al. have shown that YTHDFs inhibit infection by decreasing reverse transcription of the viral genome [78] while Kennedy et al. presented data supporting a role of YTHDFs as enhancers of the expression of both viral RNA and proteins and of viral replication [76]. Further studies are then clearly needed to clarify the role of m6A marks on the infection of human cells by various viruses.
4. Schizosaccharomyces pombe Mmi1, a Fission within YTH Family Proteins
Comparative genomics led to the conclusion that the m6A ‘writers’ homologous to human METTL3 and METTL14 proteins are absent in S. pombe fission yeast [79,80]. However, S. pombe Mmi1 (for Meiotic mRNA interception 1), a YTH-domain containing protein, is present in this organism. Mmi1 has the same organization as YTHDFs with a low-complexity N-terminal region rich in Prolines and charged residues followed by a C-terminal YTH (Figure 2).
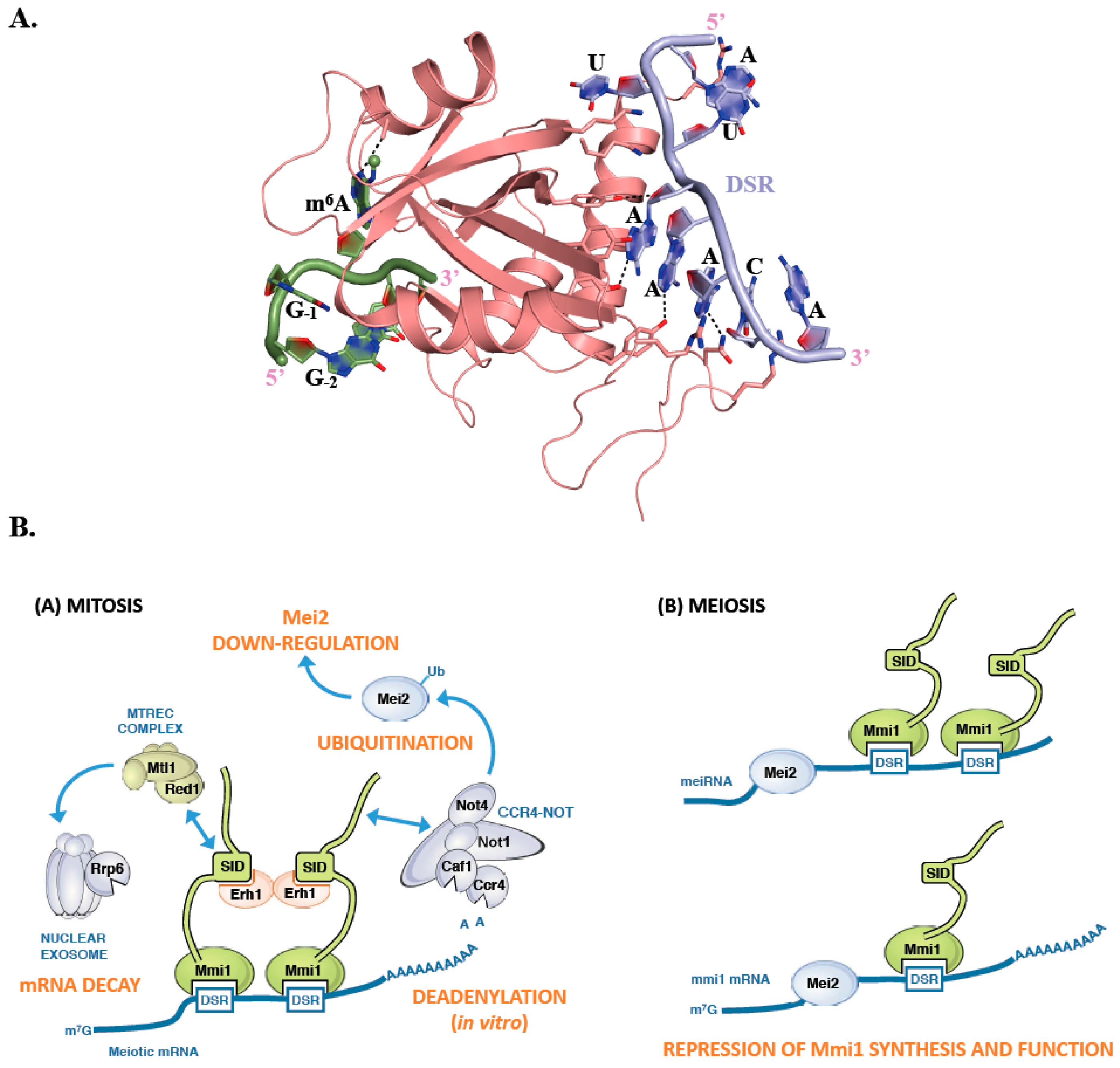
Mmi1 is an RNA binding protein that specifically recognizes a consensus U(U/C/G)AAAC motif, present in multiple copies within larger regions called Determinant of Selective Removal (DSR). This motif radically differs from the m6A consensus binding site recognized by canonical YTH domains [81]. Mmi1 YTH domain is responsible for the binding to the DSR and is assisted in this function by a low complexity region located upstream this YTH domain [82]. Although this domain is structurally similar to YTH domains from other proteins such as YTHDC1, YTHDF2, and Pho92, it is using a different surface to interact with its RNA consensus motif (Figure 4A). This binding surface is located on the opposite face of the Mmi1 YTH domain compared to the region involved in m6A binding by classical YTH domains (Figure 4A; [82,83,84]). This region is conserved in fission yeasts (S. pombe, Schizosaccharomyces japonicus, Schizosaccharomyces octosporus, and Schizosaccharomyces cryophilus) but not in other YTH containing proteins. Mmi1 YTH domain is incapable of binding a GGm6AC containing RNA, and N6-methylation at any position of the DSR motif weakens binding to the RNA [83]. Similarly, the YTH domains from YTHDC1, YTHDF2 and Pho92 do not bind DSR consensus motif. Detailed comparison of the Mmi1 pocket corresponding to m6A binding site in other YTH domain proteins shows that two Trp residues are conserved while the position corresponding to Trp, Leu, or Tyr in canonical YTH domains is occupied by a His in Mmi1. However, two main differences that could explain the Mmi1 YTH domain inability to bind m6A containing RNAs have been observed. First, the amino acid corresponding to the Asn, Asp, or His residues forming a hydrogen bond with N1 atom of the adenosine ring is substituted by Ala in Mmi1. Second, while the surface surrounding the m6A binding pocket is positively charged in canonical YTH domains, the corresponding region in Mmi1 is negatively charged, which is not favorable for RNA binding [83]. Hence, Mmi1 is an exception among YTH domains as it cannot recognize m6A modification and interacts with RNA in a completely different manner.
Mmi1 is a key cell fate regulator that promotes degradation of meiosis-specific transcripts in mitotic cells and hence seems to fulfill a similar function as human YTHDF2 and S. cerevisiae Pho92, namely the regulation of the stability of its RNA targets [26,68,81,85]. During mitosis, Mmi1 localizes in the nucleus and stably interacts with Erh1, a small protein with still unclear function, to form the Erh1-Mmi1 Complex (EMC) (Figure 4B; [86]). The EMC associates with MTREC (Mtl1-Red1 core), a multi-subunit complex containing, among others, the zinc-finger protein Red1, the Mtr4-like RNA helicase Mtl1, and the serine and proline-rich protein Pir1/Iss10 [86,87,88,89,90]. EMC and MTREC associate and cooperate with the Rrp6 subunit of the nuclear exosome for the selective elimination of meiotic DSR-containing transcripts (Figure 4B; [81,91,92]). In addition, EMC sequesters meiotic mRNAs in nuclear foci, preventing their export to the cytoplasm and their translation [90]. Therefore, Mmi1 silences the expression of meiotic genes in at least two different ways, through RNA degradation and nuclear retention.
Recent works have also reported a tight association between Mmi1 and the CCR4-NOT complex, resulting in its recruitment to meiosis specific transcripts in vivo as well as in the stimulation of deadenylation activity in vitro [44,86,93,94]. However, this function of the CCR4-NOT complex is not mandatory for the degradation of meiotic mRNAs. Rather, Mmi1 recruits CCR4-NOT complex to promote ubiquitination and down-regulation of its own inhibitor, the meiosis inducer Mei2, via the Not4/Mot2 E3 ubiquitin ligase subunit [89]. This regulatory circuit preserves the activity of Mmi1, ensuring efficient meiotic mRNA degradation in mitotic cells. Upon entry into meiosis, instead, Mmi1 is sequestered in an RNP (ribonucleoprotein) complex formed by the Mei2 protein and the long noncoding meiRNA, thereby allowing expression of the meiotic program (Figure 4B; [81]). Mei2 may also inactivate Mmi1 at the mRNA level as it binds to its transcripts during early meiosis [95]. Further studies are needed to clarify the relationships between these two main effectors of the mitosis-meiosis switch in S. pombe.
5. Conclusions
The technological progresses made during the last decade have allowed the identification of RNA modifications present at low abundance and on RNA species with limited stability such as mRNAs and long noncoding RNAs in eukaryotic cells. This research has created the basis for the characterization of proteins and multi-protein complexes involved in the deposition, elimination, and detection of these modifications. So far, much of the studies have focused on the most abundant modification, namely m6A. As a consequence, the best characterized m6A ‘readers’ are YTH-containing proteins and studies in human cells have shown that the five human YTH proteins are affecting mRNA fates such as splicing, export, translation, and decay by recruiting various protein partners. However, there is increasing evidence that proteins lacking YTH domains can also selectively recognize m6A marks. This is indeed the case for instance of the eIF3 initiation factor [70], the FMR1 RNA binding protein, which loss results in fragile X-linked mental retardation syndrome [28,96] or the insulin-like growth factor 3 mRNA-binding proteins IGF2BPs [97]. Undoubtedly, epitranscriptomics is a highly dynamic field and future work will identify new ‘readers’ of epitranscriptomics marks as has recently been the case for ALYREF, an m5C ‘reader’ [98], paving the way for the description of highly sophisticated mechanisms to control gene expression at the mRNA level.
Author Contributions
D.H., C.C. and M.G. drafted the article and approved the version to be published. We apologize for the many studies that could not be cited due to space constraints. We are indebted to Mathieu Rougemaille for critical reading of the manuscript.
Funding
This work was supported by Ecole Polytechnique, the Centre National pour la Recherche Scientifique, the Agence Nationale pour la Recherche [grant ANR-16-CE11-0003]. D.H. was supported by a PhD fellowship from the French Ministère de l’Enseignement Supérieur et de la Recherche (MESR).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lingel, A.; Sattler, M. Novel modes of protein-RNA recognition in the RNAi pathway. Curr. Opin. Struct. Biol. 2005, 15, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Daubner, G.M.; Clery, A.; Allain, F.H. RRM-RNA recognition: NMR or crystallography...and new findings. Curr. Opin. Struct. Biol. 2013, 23, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Masliah, G.; Barraud, P.; Allain, F.H. RNA recognition by double-stranded RNA binding domains: A matter of shape and sequence. Cell. Mol. Life Sci. 2013, 70, 1875–1895. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, G.; Taylor, I.A.; Ramos, A. KH-RNA interactions: Back in the groove. Curr. Opin. Struct. Biol. 2015, 30, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Baltz, A.G.; Munschauer, M.; Schwanhausser, B.; Vasile, A.; Murakawa, Y.; Schueler, M.; Youngs, N.; Penfold-Brown, D.; Drew, K.; Milek, M.; et al. The mRNA-bound proteome and its global occupancy profile on protein-coding transcripts. Mol. Cell 2012, 46, 674–690. [Google Scholar] [CrossRef] [PubMed]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA biology from an atlas of mammalian mRNA-binding proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.F.; Jain, S.; She, M.; Parker, R. Global analysis of yeast mRNPs. Nat. Struct. Mol. Biol. 2013, 20, 127–133. [Google Scholar] [CrossRef]
- Castello, A.; Fischer, B.; Frese, C.K.; Horos, R.; Alleaume, A.M.; Foehr, S.; Curk, T.; Krijgsveld, J.; Hentze, M.W. Comprehensive identification of RNA-binding domains in human cells. Mol. Cell 2016, 63, 696–710. [Google Scholar] [CrossRef]
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Squires, J.E.; Patel, H.R.; Nousch, M.; Sibbritt, T.; Humphreys, D.T.; Parker, B.J.; Suter, C.M.; Preiss, T. Widespread occurrence of 5-methylcytosine in human coding and non-coding RNA. Nucleic Acids Res. 2012, 40, 5023–5033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.; Agarwala, S.D.; Mumbach, M.R.; Jovanovic, M.; Mertins, P.; Shishkin, A.; Tabach, Y.; Mikkelsen, T.S.; Satija, R.; Ruvkun, G.; et al. High-resolution mapping reveals a conserved, widespread, dynamic mRNA methylation program in yeast meiosis. Cell 2013, 155, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Carlile, T.M.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.; Bernstein, D.A.; Mumbach, M.R.; Jovanovic, M.; Herbst, R.H.; Leon-Ricardo, B.X.; Engreitz, J.M.; Guttman, M.; Satija, R.; Lander, E.S.; et al. Transcriptome-wide mapping reveals widespread dynamic-regulated pseudouridylation of ncRNA and mRNA. Cell 2014, 159, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Nachtergaele, S.; Moshitch-Moshkovitz, S.; Peer, E.; Kol, N.; Ben-Haim, M.S.; Dai, Q.; Di Segni, A.; Salmon-Divon, M.; Clark, W.C.; et al. The dynamic N(1)-methyladenosine methylome in eukaryotic messenger RNA. Nature 2016, 530, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Safra, M.; Sas-Chen, A.; Nir, R.; Winkler, R.; Nachshon, A.; Bar-Yaacov, D.; Erlacher, M.; Rossmanith, W.; Stern-Ginossar, N.; Schwartz, S. The m1A landscape on cytosolic and mitochondrial mRNA at single-base resolution. Nature 2017, 551, 251–255. [Google Scholar] [CrossRef]
- Arango, D.; Sturgill, D.; Alhusaini, N.; Dillman, A.A.; Sweet, T.J.; Hanson, G.; Hosogane, M.; Sinclair, W.R.; Nanan, K.K.; Mandler, M.D.; et al. Acetylation of cytidine in mRNA promotes translation efficiency. Cell 2018. [Google Scholar] [CrossRef]
- Gilbert, W.V.; Bell, T.A.; Schaening, C. Messenger RNA modifications: Form, distribution, and function. Science 2016, 352, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Peer, E.; Rechavi, G.; Dominissini, D. Epitranscriptomics: Regulation of mRNA metabolism through modifications. Curr. Opin. Chem. Biol. 2017, 41, 93–98. [Google Scholar] [CrossRef]
- Roost, C.; Lynch, S.R.; Batista, P.J.; Qu, K.; Chang, H.Y.; Kool, E.T. Structure and thermodynamics of N6-methyladenosine in RNA: A spring-loaded base modification. J. Am. Chem. Soc. 2015, 137, 2107–2115. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vagbo, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef]
- Edupuganti, R.R.; Geiger, S.; Lindeboom, R.G.H.; Shi, H.; Hsu, P.J.; Lu, Z.; Wang, S.Y.; Baltissen, M.P.A.; Jansen, P.; Rossa, M.; et al. N(6)-methyladenosine (m(6)A) recruits and repels proteins to regulate mRNA homeostasis. Nat. Struct. Mol. Biol. 2017, 24, 870–878. [Google Scholar] [CrossRef]
- Imai, Y.; Matsuo, N.; Ogawa, S.; Tohyama, M.; Takagi, T. Cloning of a gene, YT521, for a novel RNA splicing-related protein induced by hypoxia/reoxygenation. Brain research. Mol. Brain Res. 1998, 53, 33–40. [Google Scholar] [CrossRef]
- Hartmann, A.M.; Nayler, O.; Schwaiger, F.W.; Obermeier, A.; Stamm, S. The interaction and colocalization of Sam68 with the splicing-associated factor YT521-B in nuclear dots is regulated by the Src family kinase p59(fyn). Mol. Biol. Cell 1999, 10, 3909–3926. [Google Scholar] [CrossRef]
- Stoilov, P.; Rafalska, I.; Stamm, S. YTH: A new domain in nuclear proteins. Trends Biochem. Sci. 2002, 27, 495–497. [Google Scholar] [CrossRef]
- Bhat, S.S.; Bielewicz, D.; Jarmolowski, A.; Szweykowska-Kulinska, Z. N(6)-methyladenosine (m(6)A): Revisiting the old with focus on new, an Arabidopsis thaliana centered review. Genes 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Theler, D.; Kaminska, K.H.; Hiller, M.; de la Grange, P.; Pudimat, R.; Rafalska, I.; Heinrich, B.; Bujnicki, J.M.; Allain, F.H.; et al. The YTH domain is a novel RNA binding domain. J. Biol. Chem. 2010, 285, 14701–14710. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhao, D.; Wu, J.; Shi, Y. Structure of the YTH domain of human YTHDF2 in complex with an m(6)A mononucleotide reveals an aromatic cage for m(6)A recognition. Cell Res. 2014, 24, 1490–1492. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Tong, L. Molecular basis for the recognition of methylated adenines in RNA by the eukaryotic YTH domain. Proc. Natl. Acad. Sci. USA 2014, 111, 13834–13839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theler, D.; Dominguez, C.; Blatter, M.; Boudet, J.; Allain, F.H. Solution structure of the YTH domain in complex with N6-methyladenosine RNA: A reader of methylated RNA. Nucleic Acids Res. 2014, 42, 13911–13919. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, X.; Liu, K.; Roundtree, I.A.; Tempel, W.; Li, Y.; Lu, Z.; He, C.; Min, J. Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat. Chem. Biol. 2014, 10, 927–929. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, K.; Ahmed, H.; Loppnau, P.; Schapira, M.; Min, J. Structural basis for the discriminative recognition of N6-Methyladenosine RNA by the human YT521-B homology domain family of proteins. J. Biol. Chem. 2015, 290, 24902–24913. [Google Scholar] [CrossRef]
- Brooks, B.R.; Brooks, C.L., 3rd; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [Green Version]
- Lunde, B.M.; Moore, C.; Varani, G. RNA-binding proteins: Modular design for efficient function. Nature reviews. Mol. Cell Biol. 2007, 8, 479–490. [Google Scholar]
- Achsel, T.; Bagni, C. Cooperativity in RNA-protein interactions: The complex is more than the sum of its partners. Curr. Opin. Neurobiol. 2016, 39, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N(6)-methyladenosine modulates messenger RNA translation efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m(6)A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef] [PubMed]
- Stowell, J.A.W.; Webster, M.W.; Kogel, A.; Wolf, J.; Shelley, K.L.; Passmore, L.A. Reconstitution of targeted deadenylation by the CCR4-NOT complex and the YTH domain protein Mmi1. Cell Rep. 2016, 17, 1978–1989. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.S.; Hao, Y.J.; Sun, B.F.; Sun, H.Y.; Li, A.; Ping, X.L.; Lai, W.Y.; et al. Nuclear m(6)A reader YTHDC1 regulates mRNA splicing. Mol. Cell 2016, 61, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Chen, Y.S.; Ping, X.L.; Yang, X.; Xiao, W.; Yang, Y.; Sun, H.Y.; Zhu, Q.; Baidya, P.; Wang, X.; et al. Cytoplasmic m(6)A reader YTHDF3 promotes mRNA translation. Cell Res. 2017, 27, 444–447. [Google Scholar] [CrossRef]
- Roundtree, I.A.; Luo, G.Z.; Zhang, Z.; Wang, X.; Zhou, T.; Cui, Y.; Sha, J.; Huang, X.; Guerrero, L.; Xie, P.; et al. YTHDC1 mediates nuclear export of N(6)-methyladenosine methylated mRNAs. eLife 2017, 6, e31311. [Google Scholar] [CrossRef]
- Nayler, O.; Hartmann, A.M.; Stamm, S. The ER repeat protein YT521-B localizes to a novel subnuclear compartment. J. Cell Biol. 2000, 150, 949–962. [Google Scholar] [CrossRef]
- Lence, T.; Akhtar, J.; Bayer, M.; Schmid, K.; Spindler, L.; Ho, C.H.; Kreim, N.; Andrade-Navarro, M.A.; Poeck, B.; Helm, M.; et al. m(6)A modulates neuronal functions and sex determination in Drosophila. Nature 2016, 540, 242–247. [Google Scholar] [CrossRef]
- Kasowitz, S.D.; Ma, J.; Anderson, S.J.; Leu, N.A.; Xu, Y.; Gregory, B.D.; Schultz, R.M.; Wang, P.J. Nuclear m6A reader YTHDC1 regulates alternative polyadenylation and splicing during mouse oocyte development. PLoS Genet. 2018, 14, e1007412. [Google Scholar] [CrossRef]
- Shima, H.; Matsumoto, M.; Ishigami, Y.; Ebina, M.; Muto, A.; Sato, Y.; Kumagai, S.; Ochiai, K.; Suzuki, T.; Igarashi, K. S-adenosylmethionine synthesis is regulated by selective N(6)-adenosine methylation and mRNA degradation involving METTL16 and YTHDC1. Cell Rep. 2017, 21, 3354–3363. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Conrad, N.K. The U6 snRNA m(6)A methyltransferase METTL16 regulates SAM synthetase intron retention. Cell 2017, 169, 824–835. [Google Scholar] [CrossRef]
- Mendel, M.; Chen, K.M.; Homolka, D.; Gos, P.; Pandey, R.R.; McCarthy, A.A.; Pillai, R.S. Methylation of structured RNA by the m(6)A writer METTL16 is essential for mouse embryonic development. Mol. Cell 2018, 71, 986–1000. [Google Scholar] [CrossRef]
- Salz, H.K.; Maine, E.M.; Keyes, L.N.; Samuels, M.E.; Cline, T.W.; Schedl, P. The Drosophila female-specific sex-determination gene, Sex-lethal, has stage-, tissue-, and sex-specific RNAs suggesting multiple modes of regulation. Genes Dev. 1989, 3, 708–719. [Google Scholar] [CrossRef]
- Moschall, R.; Gaik, M.; Medenbach, J. Promiscuity in post-transcriptional control of gene expression: Drosophila sex-lethal and its regulatory partnerships. FEBS Lett. 2017, 591, 1471–1488. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.P.; Chen, C.K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, J.; Rao, H.; Hackert, P.; Sloan, K.E.; Hobartner, C.; Bohnsack, M.T. The m(6)A reader protein YTHDC2 interacts with the small ribosomal subunit and the 5′-3′ exoribonuclease XRN1. RNA 2018, 24, 1339–1350. [Google Scholar] [CrossRef]
- Chen, M.C.; Tippana, R.; Demeshkina, N.A.; Murat, P.; Balasubramanian, S.; Myong, S.; Ferre-D’Amare, A.R. Structural basis of G-quadruplex unfolding by the DEAH/RHA helicase DHX36. Nature 2018, 558, 465–469. [Google Scholar] [CrossRef]
- Morohashi, K.; Sahara, H.; Watashi, K.; Iwabata, K.; Sunoki, T.; Kuramochi, K.; Takakusagi, K.; Miyashita, H.; Sato, N.; Tanabe, A.; et al. Cyclosporin A associated helicase-like protein facilitates the association of hepatitis C virus RNA polymerase with its cellular cyclophilin B. PLoS ONE 2011, 6, e18285. [Google Scholar] [CrossRef]
- Wojtas, M.N.; Pandey, R.R.; Mendel, M.; Homolka, D.; Sachidanandam, R.; Pillai, R.S. Regulation of m(6)A transcripts by the 3′->5′ RNA helicase YTHDC2 is essential for a successful meiotic program in the mammalian germline. Mol. Cell 2017, 68, 374–387. [Google Scholar] [CrossRef]
- Jain, D.; Puno, M.R.; Meydan, C.; Lailler, N.; Mason, C.E.; Lima, C.D.; Anderson, K.V.; Keeney, S. ketu mutant mice uncover an essential meiotic function for the ancient RNA helicase YTHDC2. eLife 2018, 7, e30919. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.J.; Zhu, Y.; Ma, H.; Guo, Y.; Shi, X.; Liu, Y.; Qi, M.; Lu, Z.; Shi, H.; Wang, J.; et al. YTHDC2 is an N(6)-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017, 27, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, A.; Konno, J.; Tanikawa, K.; Sahara, H. Transcriptional machinery of TNF-α-inducible YTH domain containing 2 (YTHDC2) gene. Gene 2014, 535, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, A.; Tanikawa, K.; Tsunetomi, M.; Takai, K.; Ikeda, H.; Konno, J.; Torigoe, T.; Maeda, H.; Kutomi, G.; Okita, K.; et al. RNA helicase YTHDC2 promotes cancer metastasis via the enhancement of the efficiency by which HIF-1α mRNA is translated. Cancer Lett. 2016, 376, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Soh, Y.Q.S.; Mikedis, M.M.; Kojima, M.; Godfrey, A.K.; de Rooij, D.G.; Page, D.C. Meioc maintains an extended meiotic prophase I in mice. PLoS Genet. 2017, 13, e1006704. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.S.; Batista, P.J.; Gold, R.S.; Chen, Y.G.; de Rooij, D.G.; Chang, H.Y.; Fuller, M.T. The conserved RNA helicase YTHDC2 regulates the transition from proliferation to differentiation in the germline. eLife 2017, 6, e26116. [Google Scholar] [CrossRef] [PubMed]
- Abby, E.; Tourpin, S.; Ribeiro, J.; Daniel, K.; Messiaen, S.; Moison, D.; Guerquin, J.; Gaillard, J.C.; Armengaud, J.; Langa, F.; et al. Implementation of meiosis prophase I programme requires a conserved retinoid-independent stabilizer of meiotic transcripts. Nat. Commun. 2016, 7, 10324. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.J.; Jeong, S.J.; Kim, K.N.; Baek, I.J.; Chang, M.; Kang, C.M.; Park, Y.S.; Yun, C.W. A novel protein, Pho92, has a conserved YTH domain and regulates phosphate metabolism by decreasing the mRNA stability of PHO4 in Saccharomyces cerevisiae. Biochem. J. 2014, 457, 391–400. [Google Scholar] [CrossRef]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.B. Dynamic m(6)A mRNA methylation directs translational control of heat shock response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5′ UTR m(6)A promotes Cap-independent translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, X.; Weng, Y.L.; Lu, Z.; Liu, Y.; Lu, Z.; Li, J.; Hao, P.; Zhang, Y.; Zhang, F.; et al. m(6)A facilitates hippocampus-dependent learning and memory through YTHDF1. Nature 2018, 563, 249–253. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 facilitates translation and decay of N(6)-methyladenosine-modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Anders, M.; Chelysheva, I.; Goebel, I.; Trenkner, T.; Zhou, J.; Mao, Y.; Verzini, S.; Qian, S.B.; Ignatova, Z. Dynamic m(6)A methylation facilitates mRNA triaging to stress granules. Life Sci. Alliance 2018, 1, e201800113. [Google Scholar] [CrossRef] [PubMed]
- Krug, R.M.; Morgan, M.A.; Shatkin, A.J. Influenza viral mRNA contains internal N6-methyladenosine and 5′-terminal 7-methylguanosine in cap structures. J. Virol. 1976, 20, 45–53. [Google Scholar]
- Gokhale, N.S.; McIntyre, A.B.R.; McFadden, M.J.; Roder, A.E.; Kennedy, E.M.; Gandara, J.A.; Hopcraft, S.E.; Quicke, K.M.; Vazquez, C.; Willer, J.; et al. N6-methyladenosine in Flaviviridae viral RNA genomes regulates infection. Cell Host Microbe 2016, 20, 654–665. [Google Scholar] [CrossRef]
- Kennedy, E.M.; Bogerd, H.P.; Kornepati, A.V.; Kang, D.; Ghoshal, D.; Marshall, J.B.; Poling, B.C.; Tsai, K.; Gokhale, N.S.; Horner, S.M.; et al. Posttranscriptional m(6)A editing of HIV-1 mRNAs enhances viral gene expression. Cell Host Microbe 2016, 19, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Lichinchi, G.; Zhao, B.S.; Wu, Y.; Lu, Z.; Qin, Y.; He, C.; Rana, T.M. Dynamics of human and viral RNA methylation during Zika virus infection. Cell Host Microbe 2016, 20, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Tirumuru, N.; Zhao, B.S.; Lu, W.; Lu, Z.; He, C.; Wu, L. N(6)-methyladenosine of HIV-1 RNA regulates viral infection and HIV-1 Gag protein expression. eLife 2016, 5, e15528. [Google Scholar] [CrossRef]
- Bujnicki, J.M.; Feder, M.; Radlinska, M.; Blumenthal, R.M. Structure prediction and phylogenetic analysis of a functionally diverse family of proteins homologous to the MT-A70 subunit of the human mRNA:m(6)A methyltransferase. J. Mol. Evol. 2002, 55, 431–444. [Google Scholar] [CrossRef]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene expression regulation mediated through reversible m(6)A RNA methylation. Nature reviews. Genetics 2014, 15, 293–306. [Google Scholar]
- Harigaya, Y.; Tanaka, H.; Yamanaka, S.; Tanaka, K.; Watanabe, Y.; Tsutsumi, C.; Chikashige, Y.; Hiraoka, Y.; Yamashita, A.; Yamamoto, M. Selective elimination of messenger RNA prevents an incidence of untimely meiosis. Nature 2006, 442, 45–50. [Google Scholar] [CrossRef]
- Stowell, J.A.W.; Wagstaff, J.L.; Hill, C.H.; Yu, M.; McLaughlin, S.H.; Freund, S.M.V.; Passmore, L.A. A low-complexity region in the YTH domain protein Mmi1 enhances RNA binding. J. Biol. Chem. 2018, 293, 9210–9222. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhu, Y.; Bao, H.; Jiang, Y.; Xu, C.; Wu, J.; Shi, Y. A novel RNA-binding mode of the YTH domain reveals the mechanism for recognition of determinant of selective removal by Mmi1. Nucleic Acids Res. 2016, 44, 969–982. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Xu, J.; Su, S.; Liu, H.; Gan, J.; Ma, J. Structural insights into the specific recognition of DSR by the YTH domain containing protein Mmi1. Biochem. Biophys. Res. Commun. 2017, 491, 310–316. [Google Scholar] [CrossRef]
- McPheeters, D.S.; Cremona, N.; Sunder, S.; Chen, H.M.; Averbeck, N.; Leatherwood, J.; Wise, J.A. A complex gene regulatory mechanism that operates at the nexus of multiple RNA processing decisions. Nat. Struct. Mol.Biol. 2009, 16, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, T.; Thillainadesan, G.; Chalamcharla, V.R.; Meng, Z.; Balachandran, V.; Dhakshnamoorthy, J.; Zhou, M.; Grewal, S.I.S. Enhancer of rudimentary cooperates with conserved RNA-processing factors to promote meiotic mRNA decay and facultative heterochromatin assembly. Mol. Cell 2016, 61, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Egan, E.D.; Braun, C.R.; Gygi, S.P.; Moazed, D. Post-transcriptional regulation of meiotic genes by a nuclear RNA silencing complex. RNA 2014, 20, 867–881. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhu, J.; Schermann, G.; Ohle, C.; Bendrin, K.; Sugioka-Sugiyama, R.; Sugiyama, T.; Fischer, T. The fission yeast MTREC complex targets CUTs and unspliced pre-mRNAs to the nuclear exosome. Nat. Commun. 2015, 6, 7050. [Google Scholar] [CrossRef] [Green Version]
- Simonetti, F.; Candelli, T.; Leon, S.; Libri, D.; Rougemaille, M. Ubiquitination-dependent control of sexual differentiation in fission yeast. eLife 2017, 6, e28046. [Google Scholar] [CrossRef]
- Shichino, Y.; Otsubo, Y.; Kimori, Y.; Yamamoto, M.; Yamashita, A. YTH-RNA-binding protein prevents deleterious expression of meiotic proteins by tethering their mRNAs to nuclear foci. eLife 2018, 7, e32155. [Google Scholar] [CrossRef]
- Houseley, J.; LaCava, J.; Tollervey, D. RNA-quality control by the exosome. Nature reviews. Mol. Cell Biol. 2006, 7, 529–539. [Google Scholar]
- Yamanaka, S.; Yamashita, A.; Harigaya, Y.; Iwata, R.; Yamamoto, M. Importance of polyadenylation in the selective elimination of meiotic mRNAs in growing S. pombe cells. EMBO J. 2010, 29, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Cotobal, C.; Rodriguez-Lopez, M.; Duncan, C.; Hasan, A.; Yamashita, A.; Yamamoto, M.; Bahler, J.; Mata, J. Role of CCR4-NOT complex in heterochromatin formation at meiotic genes and subtelomeres in fission yeast. Epigenetics Chromatin 2015, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Ukleja, M.; Cuellar, J.; Siwaszek, A.; Kasprzak, J.M.; Czarnocki-Cieciura, M.; Bujnicki, J.M.; Dziembowski, A.; Valpuesta, J.M. The architecture of the Schizosaccharomyces pombe CCR4-NOT complex. Nat. Commun. 2016, 7, 10433. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Futcher, B.; Leatherwood, J. Mmi1 and rep2 mRNAs are novel RNA targets of the Mei2 RNA-binding protein during early meiosis in Schizosaccharomyces pombe. Open Biol. 2018, 8, 180110. [Google Scholar] [CrossRef] [PubMed]
- Penagarikano, O.; Mulle, J.G.; Warren, S.T. The pathophysiology of fragile X syndrome. Annu. Rev. Genom. Hum. Genet. 2007, 8, 109–129. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Sun, B.F.; Chen, Y.S.; Xu, J.W.; Lai, W.Y.; Li, A.; Wang, X.; Bhattarai, D.P.; Xiao, W.; et al. 5-methylcytosine promotes mRNA export—NSUN2 as the methyltransferase and ALYREF as an m(5)C reader. Cell Res. 2017, 27, 606–625. [Google Scholar] [CrossRef]
Figure 1.
Recognition of m6A (N6-methyladenosine) containing RNAs by human YTHDC1 YTH domain. (A) Ribbon representation of a GGm6ACC RNA oligonucleotide (green) bound to human YTHDC1 (PDB code: 4R3I; [37]). The methyl group grafted on N6-adenosine is shown as a sphere. The side chains from residues involved in the formation of the m6A aromatic cage and the hydrogen bonds responsible for specificity of m6A as well as for increased affinity of YTHDC1 for RNAs harboring a G just upstream of the m6A mark are shown as sticks. Hydrogen bonds are depicted by black dashed lines. (B) Mapping of the electrostatic surface at the surface of human YTHDC1 YTH domain with the GGm6ACC RNA oligonucleotide shown in green. Positively (8 kBT/e) and negatively (−8 kBT/e) charged regions are colored blue and red, respectively. The potential was calculated using the CHARMM-GUI server [39].
Figure 1.
Recognition of m6A (N6-methyladenosine) containing RNAs by human YTHDC1 YTH domain. (A) Ribbon representation of a GGm6ACC RNA oligonucleotide (green) bound to human YTHDC1 (PDB code: 4R3I; [37]). The methyl group grafted on N6-adenosine is shown as a sphere. The side chains from residues involved in the formation of the m6A aromatic cage and the hydrogen bonds responsible for specificity of m6A as well as for increased affinity of YTHDC1 for RNAs harboring a G just upstream of the m6A mark are shown as sticks. Hydrogen bonds are depicted by black dashed lines. (B) Mapping of the electrostatic surface at the surface of human YTHDC1 YTH domain with the GGm6ACC RNA oligonucleotide shown in green. Positively (8 kBT/e) and negatively (−8 kBT/e) charged regions are colored blue and red, respectively. The potential was calculated using the CHARMM-GUI server [39].

Figure 2.
Schematic representation and domain composition of human, Saccharomyces cerevisiae (Pho92) and Schizosaccharomyces pombe (Mmi1) YTH-domain containing proteins. The predicted or experimentally determined limits of domains are indicated. E: Glu-rich domain. R: Arg-rich domain. P: Pro-rich domain. RED: Arg/Glu/Asp-rich domain. G: Gly-rich domain. R3H: small domain containing an invariant Arg sported from a highly conserved His by three residues. RecA1 and RecA2: RecA domains found in helicases. Ank: Ankyrin repeats. WH: Winged-helix domain. HB: Helical bundle. OB: OB-fold. CTE: C-terminal extension. S: Ser-rich domain. P/Q: Pro and Gln-rich domain. C1BD: CNOT1 binding domain. SID: Self-Interacting domain.
Figure 2.
Schematic representation and domain composition of human, Saccharomyces cerevisiae (Pho92) and Schizosaccharomyces pombe (Mmi1) YTH-domain containing proteins. The predicted or experimentally determined limits of domains are indicated. E: Glu-rich domain. R: Arg-rich domain. P: Pro-rich domain. RED: Arg/Glu/Asp-rich domain. G: Gly-rich domain. R3H: small domain containing an invariant Arg sported from a highly conserved His by three residues. RecA1 and RecA2: RecA domains found in helicases. Ank: Ankyrin repeats. WH: Winged-helix domain. HB: Helical bundle. OB: OB-fold. CTE: C-terminal extension. S: Ser-rich domain. P/Q: Pro and Gln-rich domain. C1BD: CNOT1 binding domain. SID: Self-Interacting domain.

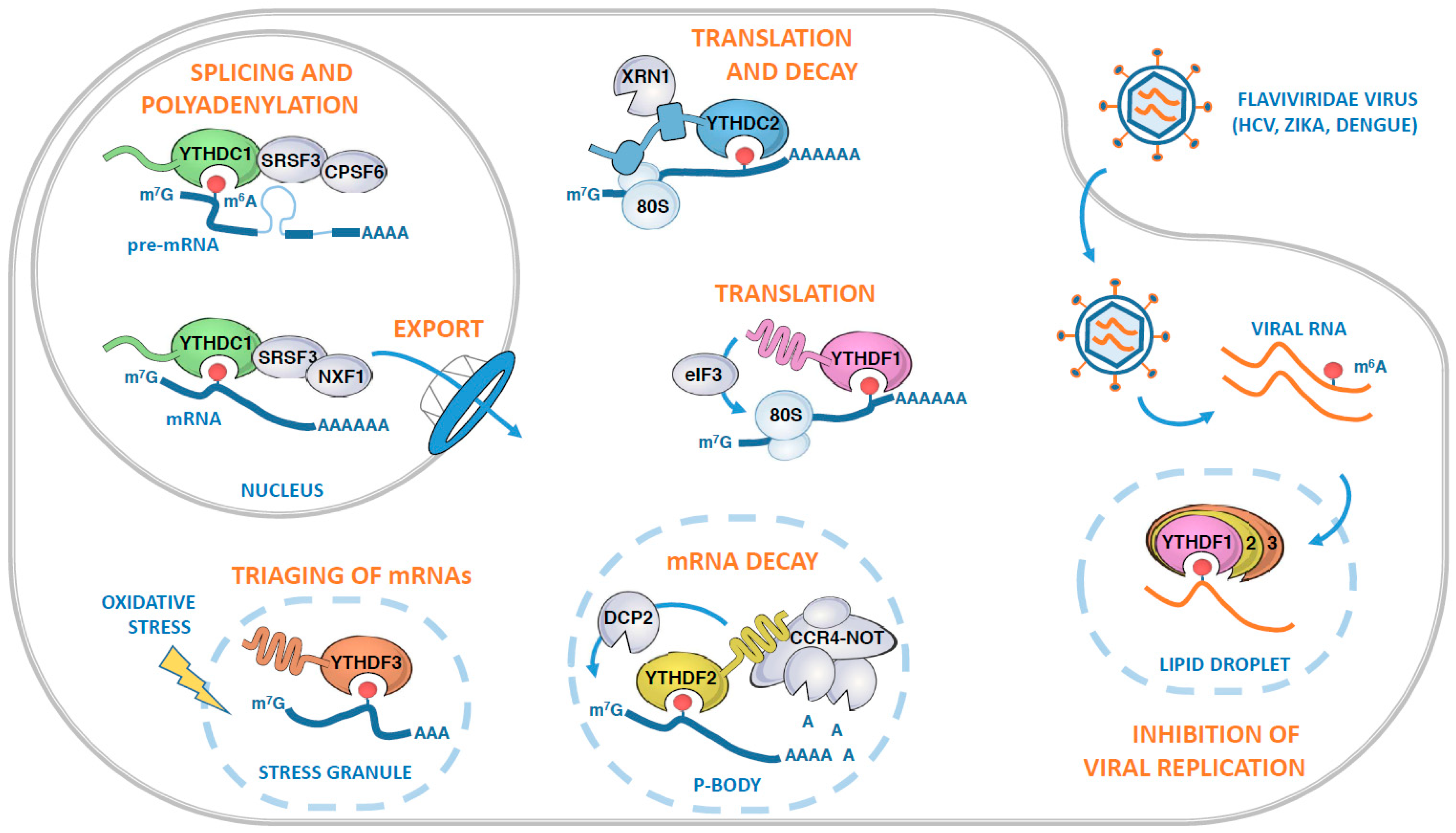
Figure 3.
Roles of human YTH-containing proteins in various aspects of messenger RNA (mRNA) fates. In the nucleus, YTHDC1 recognizes m6A-modified pre-mRNAs and orchestrates their splicing, polyadenylation, and nuclear export through its association with SRSF3, CPSF6, and NXF1. Once in the cytosol, the modified mRNAs can be bound by YTHDC2, which in turn recruits both the ribosome and the XRN1 exoribonuclease. Alternatively, the mRNA can be targeted by the YTHDF proteins, either to be actively translated in an YTHDF1-dependent manner, or subjected to mRNA decay through YTHDF2 and its ability to recruit the CCR4-NOT deadenylase complex. YTHDF2-mediated mRNA decay is likely to occur in Processing bodies (P-bodies) where it co-localizes with the decapping enzyme DCP2. Following oxidative stress, m6A-modified mRNAs can also be recognized by YTHDF3, which facilitates the triaging of mRNAs into the stress granules. The YTHDF proteins can also target viral m6A-modified RNAs during infection, as illustrated by the case of hepatitis C virus (HCV) infection. During infection, the YTHDF proteins relocalize to lipid droplets, sites of viral assembly, and sequester m6A-modified HCV RNAs, preventing their interaction with HCV core protein and subsequent virus particle production.
Figure 3.
Roles of human YTH-containing proteins in various aspects of messenger RNA (mRNA) fates. In the nucleus, YTHDC1 recognizes m6A-modified pre-mRNAs and orchestrates their splicing, polyadenylation, and nuclear export through its association with SRSF3, CPSF6, and NXF1. Once in the cytosol, the modified mRNAs can be bound by YTHDC2, which in turn recruits both the ribosome and the XRN1 exoribonuclease. Alternatively, the mRNA can be targeted by the YTHDF proteins, either to be actively translated in an YTHDF1-dependent manner, or subjected to mRNA decay through YTHDF2 and its ability to recruit the CCR4-NOT deadenylase complex. YTHDF2-mediated mRNA decay is likely to occur in Processing bodies (P-bodies) where it co-localizes with the decapping enzyme DCP2. Following oxidative stress, m6A-modified mRNAs can also be recognized by YTHDF3, which facilitates the triaging of mRNAs into the stress granules. The YTHDF proteins can also target viral m6A-modified RNAs during infection, as illustrated by the case of hepatitis C virus (HCV) infection. During infection, the YTHDF proteins relocalize to lipid droplets, sites of viral assembly, and sequester m6A-modified HCV RNAs, preventing their interaction with HCV core protein and subsequent virus particle production.

Figure 4.
S. pombe Mmi1 contains an atypical YTH domain. (A) Ribbon representation of the complex between S. pombe Mmi1 YTH domain (pink) and a Determinant of Selective Removal (DSR) RNA sequence (light blue; [84]). An m6A containing RNA fragment (green) has been modeled by superimposing the crystal structure of RNA-bound YTH domain from YTHDC1 onto the structure of S. pombe Mmi1. Some residues and hydrogen bonds important for the interaction between S. pombe Mmi1 and the RNA DSR sequence are shown as sticks or black dashed lines, respectively. (B) Cartoons summarizing how Mmi1 controls the decay of meiotic mRNAs during mitosis or is inactivated during meiosis.
Figure 4.
S. pombe Mmi1 contains an atypical YTH domain. (A) Ribbon representation of the complex between S. pombe Mmi1 YTH domain (pink) and a Determinant of Selective Removal (DSR) RNA sequence (light blue; [84]). An m6A containing RNA fragment (green) has been modeled by superimposing the crystal structure of RNA-bound YTH domain from YTHDC1 onto the structure of S. pombe Mmi1. Some residues and hydrogen bonds important for the interaction between S. pombe Mmi1 and the RNA DSR sequence are shown as sticks or black dashed lines, respectively. (B) Cartoons summarizing how Mmi1 controls the decay of meiotic mRNAs during mitosis or is inactivated during meiosis.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hazra, D.; Chapat, C.; Graille, M. m6A mRNA Destiny: Chained to the rhYTHm by the YTH-Containing Proteins. Genes 2019, 10, 49. https://doi.org/10.3390/genes10010049
AMA Style
Hazra D, Chapat C, Graille M. m6A mRNA Destiny: Chained to the rhYTHm by the YTH-Containing Proteins. Genes. 2019; 10(1):49. https://doi.org/10.3390/genes10010049
Chicago/Turabian StyleHazra, Ditipriya, Clément Chapat, and Marc Graille. 2019. "m6A mRNA Destiny: Chained to the rhYTHm by the YTH-Containing Proteins" Genes 10, no. 1: 49. https://doi.org/10.3390/genes10010049
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.