Virucidal Effects of Dielectric Barrier Discharge Plasma on Human Norovirus Infectivity in Fresh Oysters (Crassostrea gigas)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Stock Preparation
2.2. Oyster Sample Preparation and HuNoV Inoculation
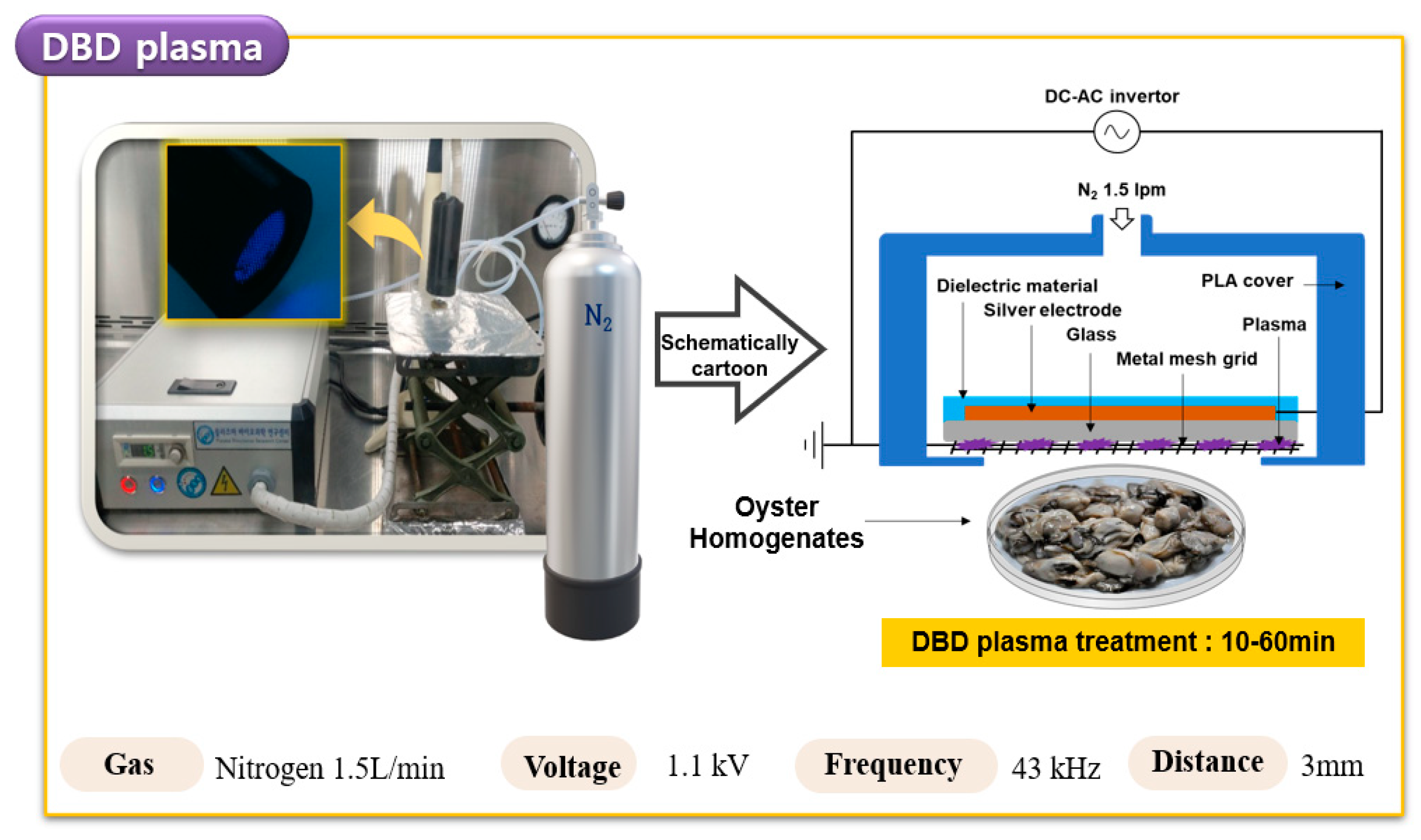
2.3. DBD Plasma Treatment of HuNoV in Oyster
2.4. Propidium Monoazide Treatment on HuNoV in Oyster
2.5. HuNoV Isolation and RNA Extraction
2.6. Quantitative Analysis of HuNoV Using RT-qPCR
2.7. DBD Plasma Reduction Kinetics
2.8. Measurement of pH and Hunter Color
2.9. Statistical Analysis
3. Results
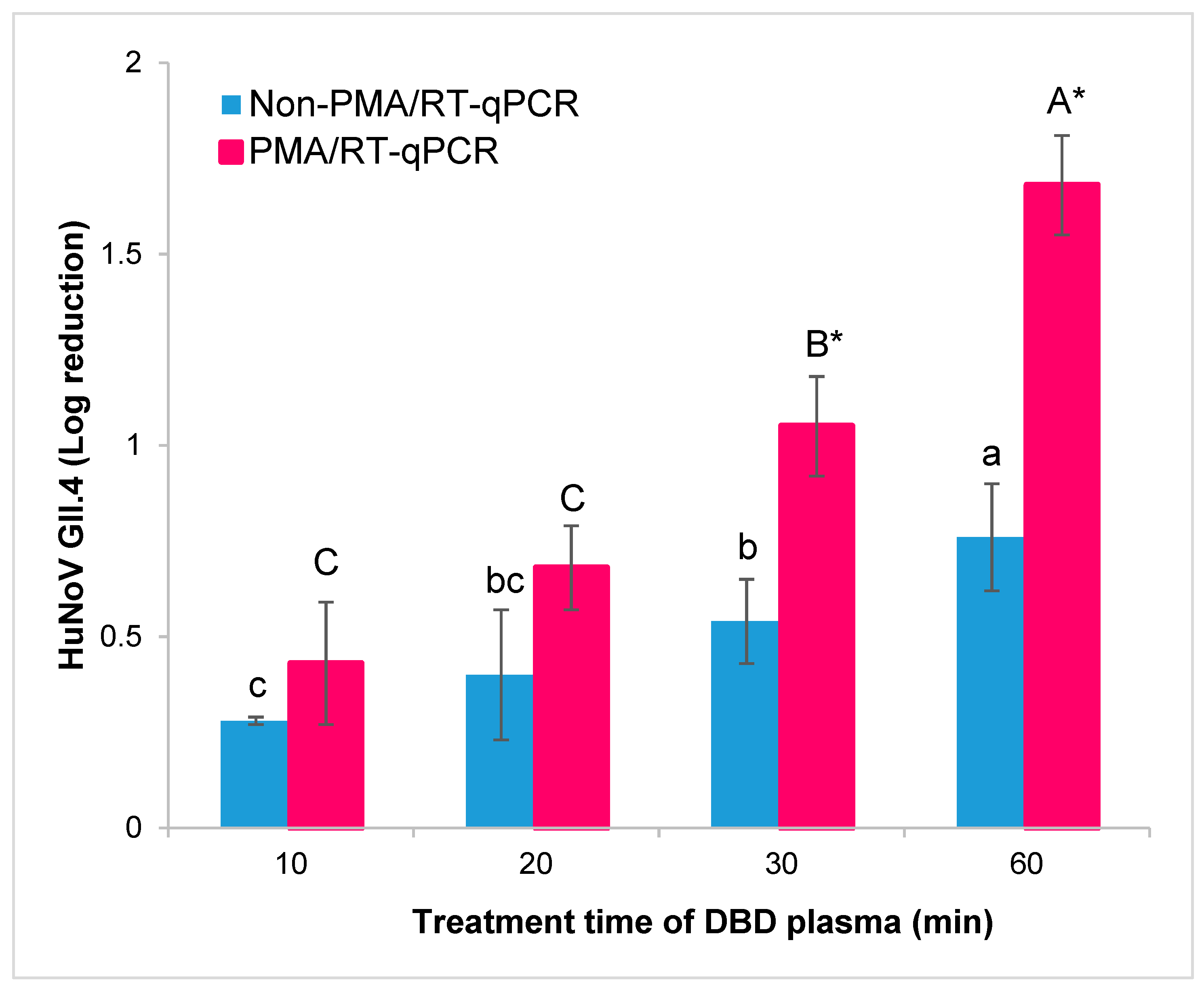
3.1. Influence of DBD Plasma Treatment on HuNoV GII.4 in Oysters by PMA/RT-qPCR
3.2. Influence of DBD Plasma Treatment on HuNoV DR-Values in Oyster
3.3. Changes in pH and Hunter Color When Using DBD Plasma Treatment in Oyster
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ryu, W.-S. Other Positive-Strand RNA Viruses. In Molecular Virology of Human Pathogenic Viruses; Elsevier BV: Amsterdam, The Netherlands, 2017; pp. 177–184. [Google Scholar]
- Tran, T.H.; Trainor, E.; Nakagomi, T.; Cunliffe, N.A.; Nakagomi, O. Molecular epidemiology of noroviruses associated with acute sporadic gastroenteritis in children: Global distribution of genogroups, genotypes and GII.4 variants. J. Clin. Virol. 2013, 56, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Graaf, M.; Van Beek, J.; Koopmans, M.P.G. Human norovirus transmission and evolution in a changing world. Nat. Rev. Genet. 2016, 14, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Cai, H.; Hu, L.; Lei, R.; Pan, Y.; Yan, S.; Wang, Y. Molecular Epidemiology of Oyster-Related Human Noroviruses and Their Global Genetic Diversity and Temporal-Geographical Distribution from 1983 to 2014. Appl. Environ. Microbiol. 2015, 81, 7615–7624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maalouf, H.; Schaeffer, J.; Parnaudeau, S.; Le Pendu, J.; Atmar, R.L.; Crawford, S.E.; Le Guyader, F.S. Strain-Dependent Norovirus Bioaccumulation in Oysters. Appl. Environ. Microbiol. 2011, 77, 3189–3196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, B.S.; Tam, T.-Y. Natural depuration of shellfish for human consumption: A note of caution. Water Res. 2000, 34, 1401–1406. [Google Scholar] [CrossRef]
- Corrêa, A.D.A.; Albarnaz, J.D.; Moresco, V.; Poli, C.R.; Teixeira, A.L.; Simões, C.M.O.; Barardi, C.R.M. Depuration dynamics of oysters (Crassostrea gigas) artificially contaminated by Salmonella enterica serovar Typhimurium. Mar. Environ. Res. 2007, 63, 479–489. [Google Scholar] [CrossRef]
- Huppatz, C.; Munnoch, S.A.; Worgan, T.; Merritt, T.; Dalton, C.B.; Kelly, P.M.; Durrheim, D.N. A norovirus outbreak associated with consumption of NSW oysters: Implications for quality assurance systems. Commun. Dis. Intell. Q. Rep. 2008, 32, 88. [Google Scholar]
- McLeod, C.; Polo, D.; Le Saux, J.-C.; Le Guyader, F.S. Depuration and Relaying: A Review on Potential Removal of Norovirus from Oysters. Compr. Rev. Food Sci. Food Saf. 2017, 16, 692–706. [Google Scholar] [CrossRef] [Green Version]
- Campos, C.J.A.; Lees, D.N. Environmental Transmission of Human Noroviruses in Shellfish Waters. Appl. Environ. Microbiol. 2014, 80, 3552–3561. [Google Scholar] [CrossRef] [Green Version]
- Parshionikar, S.; Laseke, I.; Fout, G.S. Use of Propidium Monoazide in Reverse Transcriptase PCR To Distinguish between Infectious and Noninfectious Enteric Viruses in Water Samples. Appl. Environ. Microbiol. 2010, 76, 4318–4326. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Ko, G. Using propidium monoazide to distinguish between viable and nonviable bacteria, MS2 and murine norovirus. Lett. Appl. Microbiol. 2012, 55, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Coudray-Meunier, C.; Fraisse, A.; Martin-Latil, S.; Guillier, L.; Perelle, S. Discrimination of infectious hepatitis A virus and rotavirus by combining dyes and surfactants with RT-qPCR. BMC Microbiol. 2013, 13, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, E.B.; Choi, M.-S.; Kim, J.Y.; Ha, K.S.; Kwon, J.Y.; Jeong, S.H.; Lee, H.J.; Jung, Y.J.; Ha, J.-H.; Park, S.Y. Characterizing the effects of thermal treatment on human norovirus GII.4 viability using propidium monoazide combined with RT-qPCR and quality assessments in mussels. Food Control 2020, 109, 106954. [Google Scholar] [CrossRef]
- Pankaj, S.; Wan, Z.; Keener, K.M. Effects of Cold Plasma on Food Quality: A Review. Foods 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Patil, S.; Boehm, D.; Milosavljević, V.; Cullen, P.J.; Bourke, P. Mechanisms of Inactivation by High-Voltage Atmospheric Cold Plasma Differ for Escherichia coli and Staphylococcus aureus. Appl. Environ. Microbiol. 2015, 82, 450–458. [Google Scholar] [CrossRef] [Green Version]
- López, M.; Calvo, T.; Prieto, M.; Múgica-Vidal, R.; Muro-Fraguas, I.; Alba-Elías, F.; Alvarez-Ordóñez, A. A Review on Non-thermal Atmospheric Plasma for Food Preservation: Mode of Action, Determinants of Effectiveness, and Applications. Front. Microbiol. 2019, 10, 622. [Google Scholar] [CrossRef]
- Choi, M.-S.; Kim, J.Y.; Jeon, E.B.; Park, S.Y. Antibacterial activity of dielectric barrier discharge plasma against main food-borne bacteria in suspensions. Korean J. Fish Aquat. Sci. 2019, 52, 617–624. [Google Scholar]
- Aboubakr, H.A.; Mor, S.K.; Higgins, L.; Armien, A.; Youssef, M.M.; Bruggeman, P.J.; Goyal, S.M. Cold argon-oxygen plasma species oxidize and disintegrate capsid protein of feline calicivirus. PLoS ONE 2018, 13, e0194618. [Google Scholar] [CrossRef]
- Ahlfeld, B.; Li, Y.; Boulaaba, A.; Binder, A.; Schotte, U.; Zimmermann, J.L.; Morfill, G.; Klein, G. Inactivation of a Foodborne Norovirus Outbreak Strain with Nonthermal Atmospheric Pressure Plasma. mBio 2015, 6, e02300-14. [Google Scholar] [CrossRef] [Green Version]
- Bunz, O.; Mese, K.; Zhang, W.; Piwowarczyk, A.; Ehrhardt, A. Effect of cold atmospheric plasma (CAP) on human adenoviruses is adenovirus type-dependent. PLoS ONE 2018, 13, e0202352. [Google Scholar] [CrossRef] [Green Version]
- Le Guyader, F.S.; Atmar, R.L.; Le Pendu, J. Transmission of viruses through shellfish: When specific ligands come into play. Curr. Opin. Virol. 2012, 2, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, Y.-J.; Park, J.-W.; Han, S.-H.; Cho, H.-G.; Kang, L.-H.; Lee, S.-G.; Ryu, S.-R.; Paik, S.-Y. Full-Genomic Analysis of a Human Norovirus Recombinant GII.12/13 Novel Strain Isolated from South Korea. PLoS ONE 2013, 8, e85063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-S.; Lee, S.-G.; Jin, J.-Y.; Cho, H.-G.; Jheong, W.-H.; Paik, S.-Y. Complete Nucleotide Sequence Analysis of the Norovirus GII.4 Sydney Variant in South Korea. BioMed Res. Int. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Microbiology of the Food Chain Horizontal Method for Determination of Hepatitis A Virus and Norovirus Using Real-Time RT-PCR Part 1: Method for Quantification; ISO 15216-1:2017; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- Choi, M.-S.; Jeon, E.B.; Kim, J.Y.; Choi, E.H.; Lim, J.S.; Choi, J.; Park, S.Y. Impact of non-thermal dielectric barrier discharge plasma on Staphylococcus aureus and Bacillus cereus and quality of dried blackmouth angler (Lophiomus setigerus). J. Food Eng. 2020, 278, 109952. [Google Scholar] [CrossRef]
- Jeong, M.-I.; Park, S.Y.; Ha, S.-D. Thermal inactivation of human norovirus on spinach using propidium or ethidium monoazide combined with real-time quantitative reverse transcription-polymerase chain reaction. Food Control 2017, 78, 79–84. [Google Scholar] [CrossRef]
- Kageyama, T.; Kojima, S.; Shinohara, M.; Uchida, K.; Fukushi, S.; Hoshino, F.B.; Takeda, N.; Katayama, K. Broadly Reactive and Highly Sensitive Assay for Norwalk-Like Viruses Based on Real-Time Quantitative Reverse Transcription-PCR. J. Clin. Microbiol. 2003, 41, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Albertos, I.; Martín-Diana, A.B.; Cullen, P.; Tiwari, B.; Ojha, S.; Bourke, P.; Álvarez, C.; Rico, D. Effects of dielectric barrier discharge (DBD) generated plasma on microbial reduction and quality parameters of fresh mackerel (Scomber scombrus) fillets. Innov. Food Sci. Emerg. Technol. 2017, 44, 117–122. [Google Scholar] [CrossRef]
- Yong, H.I.; Kim, H.-J.; Park, S.; Alahakoon, A.U.; Kim, K.; Choe, W.; Jo, C. Evaluation of pathogen inactivation on sliced cheese induced by encapsulated atmospheric pressure dielectric barrier discharge plasma. Food Microbiol. 2015, 46, 46–50. [Google Scholar] [CrossRef]
- Weltmann, K.D.; Kindel, E.; Von Woedtke, T.; Hähnel, M.; Stieber, M.; Brandenburg, R. Atmospheric-pressure plasma sources: Prospective tools for plasma medicine. Pure Appl. Chem. 2010, 82, 1223–1237. [Google Scholar] [CrossRef]
- D’Souza, D.H. Methods for Estimating Virus Infectivity. In Viruses in Foods; D’Souza, D.H., Ed.; Springer: Cham, Switzerland, 2016; pp. 335–365. [Google Scholar]
- Bae, S.-C.; Park, S.Y.; Choe, W.; Ha, S.-D. Inactivation of murine norovirus-1 and hepatitis A virus on fresh meats by atmospheric pressure plasma jets. Food Res. Int. 2015, 76, 342–347. [Google Scholar] [CrossRef]
- Aboubakr, H.A.; Parra, F.S.; Collins, J.; Bruggeman, P.; Goyal, S.M. Ìn situ inactivation of human norovirus GII.4 by cold plasma: Ethidium monoazide (EMA)-coupled RT-qPCR underestimates virus reduction and fecal material suppresses inactivation. Food Microbiol. 2020, 85, 103307. [Google Scholar] [CrossRef] [PubMed]
- Nayak, G.; Aboubakr, H.A.; Goyal, S.M.; Bruggeman, P.J. Reactive species responsible for the inactivation of feline calicivirus by a two-dimensional array of integrated coaxial microhollow dielectric barrier discharges in air. Plasma Process. Polym. 2018, 15, 1700119. [Google Scholar] [CrossRef]
- Moreno, L.; Aznar, R.; Sánchez, G. Application of viability PCR to discriminate the infectivity of hepatitis A virus in food samples. Int. J. Food Microbiol. 2015, 201, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.-L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef]
- Van Dycke, J.; Ny, A.; Conceição-Neto, N.; Maes, J.; Hosmillo, M.; Cuvry, A.; Goodfellow, I.; Nogueira, T.C.; Verbeken, E.; Matthijnssens, J.; et al. A robust human norovirus replication model in zebrafish larvae. PLoS Pathog. 2019, 15, e1008009. [Google Scholar] [CrossRef] [Green Version]
- Dancho, B.A.; Chen, H.; Kingsley, D.H. Discrimination between infectious and non-infectious human norovirus using porcine gastric mucin. Int. J. Food Microbiol. 2012, 155, 222–226. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on Norovirus (NoV) in oysters: Methods, limits and control options. EFSA J. 2012, 10, 2500. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Ha, S.-D. Assessment of cold oxygen plasma technology for the inactivation of major foodborne viruses on stainless steel. J. Food Eng. 2018, 223, 42–45. [Google Scholar] [CrossRef]
- Son, K.T.; Shim, K.B.; Lim, C.W.; Yoon, N.Y.; Seo, J.H.; Jeong, S.G.; Jeong, W.Y.; Cho, Y.J. Relationship of pH, Glycogen, Soluble Protein, and Turbidity between Freshness of Raw Oyster Crassostrea gigas. Korean J. Fish. Aquat. Sci. 2014, 47, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Cheng, J.; Sunabcd, D.-W. Cold Plasma-Mediated Treatments for Shelf Life Extension of Fresh Produce: A Review of Recent Research Developments. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1312–1326. [Google Scholar] [CrossRef]
- Cao, R.; Xue, C.-H.; Liu, Q.; Xue, Y. Microbiological, chemical, and sensory assessment of Pacific oysters (Crassostrea gigas) stored at different temperatures. Czech J. Food Sci. 2009, 27, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Misra, N. Quality of Cold Plasma Treated Plant Foods. In Cold Plasma in Food and Agriculture; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 253–271. [Google Scholar]
- Oehmigen, K.; Hähnel, M.; Brandenburg, R.; Wilke, C.; Weltmann, K.-D.; Von Woedtke, T. The Role of Acidification for Antimicrobial Activity of Atmospheric Pressure Plasma in Liquids. Plasma Process. Polym. 2010, 7, 250–257. [Google Scholar] [CrossRef]
- Kim, J.-W.; Puligundla, P.; Mok, C. Dielectric barrier discharge plasma for microbial decontamination of dried laver: Effects on physicochemical characteristics. Int. J. Food Sci. Technol. 2015, 50, 2630–2638. [Google Scholar] [CrossRef]
- Leon, J.S.; Kingsley, D.H.; Montes, J.S.; Richards, G.P.; Lyon, G.M.; Abdulhafid, G.M.; Seitz, S.R.; Fernandez, M.L.; Teunis, P.F.; Flick, G.J.; et al. Randomized, Double-Blinded Clinical Trial for Human Norovirus Inactivation in Oysters by High Hydrostatic Pressure Processing. Appl. Environ. Microbiol. 2011, 77, 5476–5482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Chang, S.; Nannapaneni, R.; Zhang, Y.; Coker, R.; Mahmoud, B.S. The effects of X-ray treatments on bioaccumulated murine norovirus-1 (MNV-1) and survivability, inherent microbiota, color, and firmness of Atlantic oysters (Crassostrea virginica) during storage at 5 °C for 20 days. Food Control 2017, 73, 1189–1194. [Google Scholar] [CrossRef] [Green Version]
- Bazaka, K.; Jacob, M.V.; Crawford, R.J.; Ivanova, E.P. Plasma-assisted surface modification of organic biopolymers to prevent bacterial attachment. Acta biomater. 2011, 7, 2015–2028. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| DBD Plasma Treatment (min) | Non-PMA/RT-qPCR | PMA/RT-qPCR | Before/After Using PMA to HuNoV Reduction Difference (Log Titer, % Reduction) |
|---|---|---|---|
| Log Copy Number/µL | |||
| Control | 5.51 ± 0.02 a | 5.51 ± 0.04 a | – |
| 10 | 5.23 ± 0.01 b | 5.08 ± 0.16 b | 0.15 (29.21%) |
| 20 | 5.11 ± 0.17 bc | 4.83 ± 0.11 c | 0.28 (47.52%) |
| 30 | 4.97 ± 0.11 c | 4.46 ± 0.13 d | 0.51 (61.10%) |
| 60 | 4.75 ± 0.14 d | 3.83 ± 0.13 e | 0.92 (87.98%) |
| Enteric Microorganisms | Equation of the First-Order Kinetic Model | D1-Values of DBD Plasma (min) | D2-Values of DBD Plasma (min) | R2 |
|---|---|---|---|---|
| HuNoV GII.4 with PMA | Y = −0.03 + 5.40 | 36.5 ± 1.1 | 73.0 ± 2.3 | 0.98 |
| DBD Plasma Treatment (min) | pH | Hunter Colors | ||
|---|---|---|---|---|
| “L” | “a” | “b” | ||
| Control | 5.49 ± 0.03 NS | 41.57 ± 0.38 NS | −0.21 ± 0.06 NS | 12.56 ± 0.99 NS |
| 10 | 5.49 ± 0.09 | 41.80 ± 1.22 | −0.24 ± 0.03 | 12.79 ± 1.52 |
| 20 | 5.48 ± 0.06 | 41.17 ± 1.13 | −0.19 ± 0.03 | 13.02 ± 1.21 |
| 30 | 5.47 ± 0.02 | 40.29 ± 1.05 | −0.15 ± 0.04 | 13.08 ± 1.43 |
| 60 | 5.46 ± 0.01 | 40.09 ± 1.63 | −0.15 ± 0.09 | 13.43 ± 0.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.-S.; Jeon, E.B.; Kim, J.Y.; Choi, E.H.; Lim, J.S.; Choi, J.; Ha, K.S.; Kwon, J.Y.; Jeong, S.H.; Park, S.Y. Virucidal Effects of Dielectric Barrier Discharge Plasma on Human Norovirus Infectivity in Fresh Oysters (Crassostrea gigas). Foods 2020, 9, 1731. https://doi.org/10.3390/foods9121731
Choi M-S, Jeon EB, Kim JY, Choi EH, Lim JS, Choi J, Ha KS, Kwon JY, Jeong SH, Park SY. Virucidal Effects of Dielectric Barrier Discharge Plasma on Human Norovirus Infectivity in Fresh Oysters (Crassostrea gigas). Foods. 2020; 9(12):1731. https://doi.org/10.3390/foods9121731
Chicago/Turabian StyleChoi, Man-Seok, Eun Bi Jeon, Ji Yoon Kim, Eun Ha Choi, Jun Sup Lim, Jinsung Choi, Kwang Soo Ha, Ji Young Kwon, Sang Hyeon Jeong, and Shin Young Park. 2020. "Virucidal Effects of Dielectric Barrier Discharge Plasma on Human Norovirus Infectivity in Fresh Oysters (Crassostrea gigas)" Foods 9, no. 12: 1731. https://doi.org/10.3390/foods9121731