Lactoferrin Isolation and Hydrolysis from Red Deer (Cervus elaphus) Milk and the Antibacterial Activity of Deer Lactoferrin and Its Hydrolysates



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
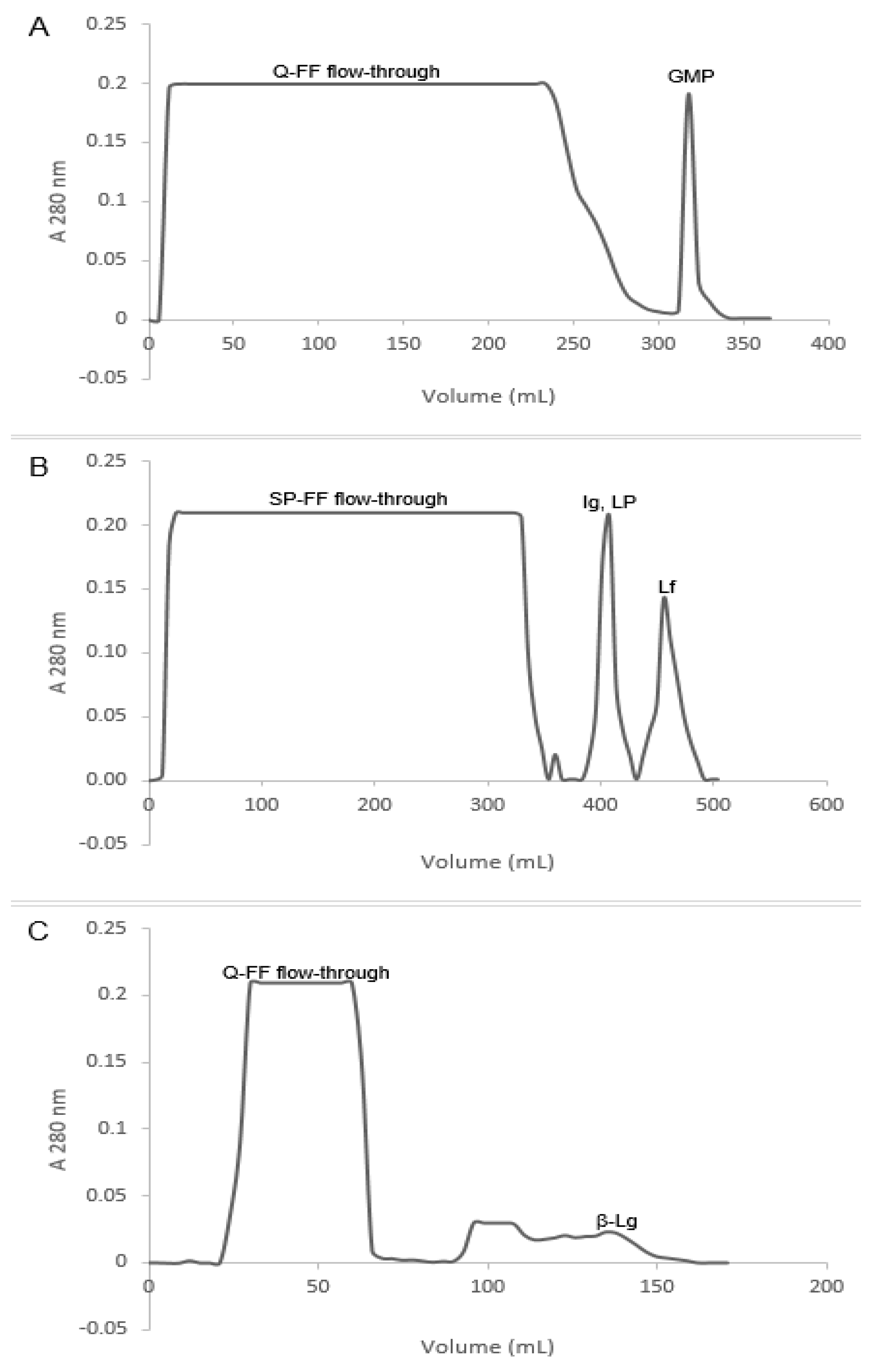
2.2. Fast Protein Liquid Chromatography
2.3. In Vitro Digestion of Lf
2.4. OPA Assay for Lf Peptide Production
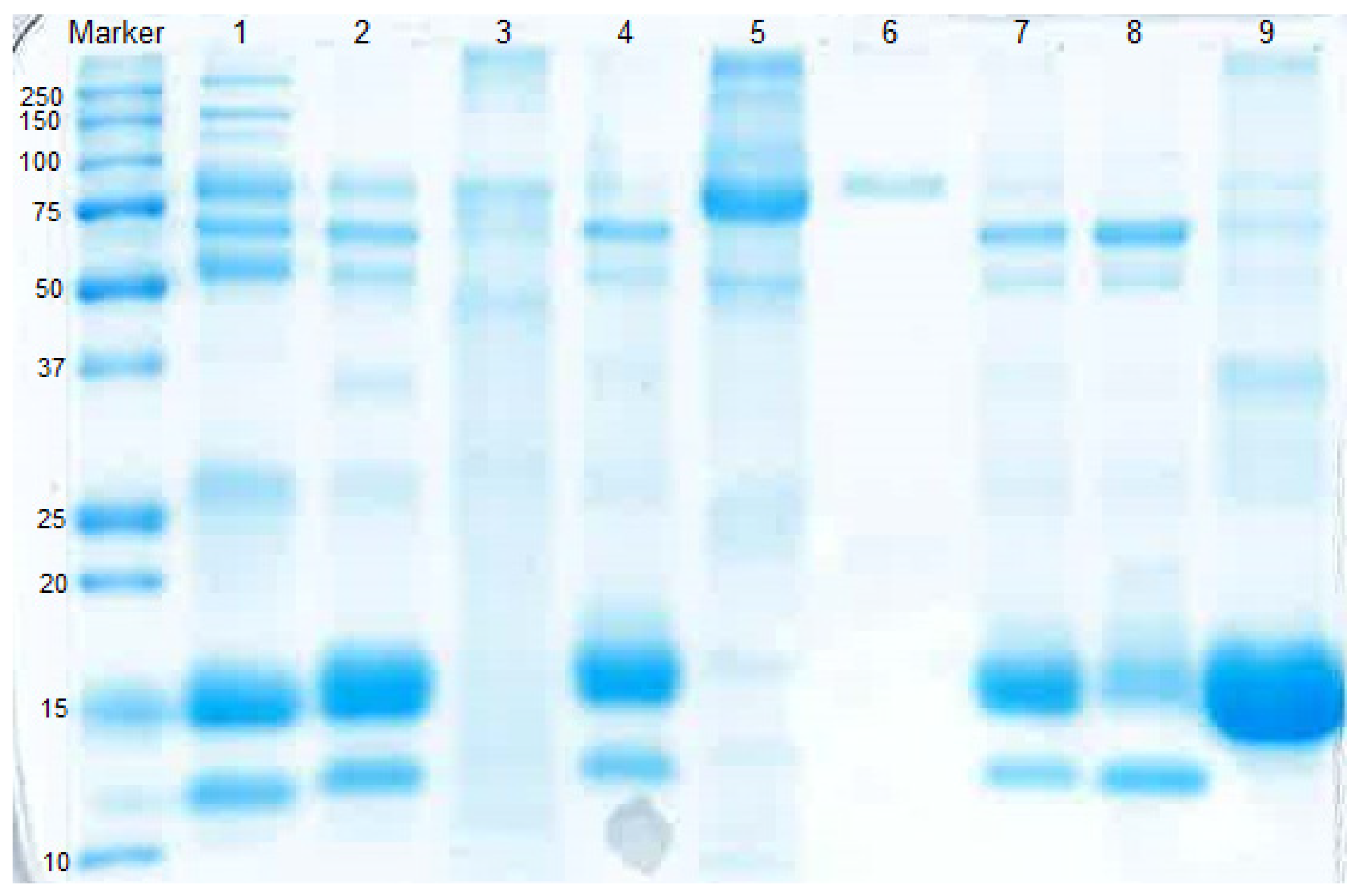
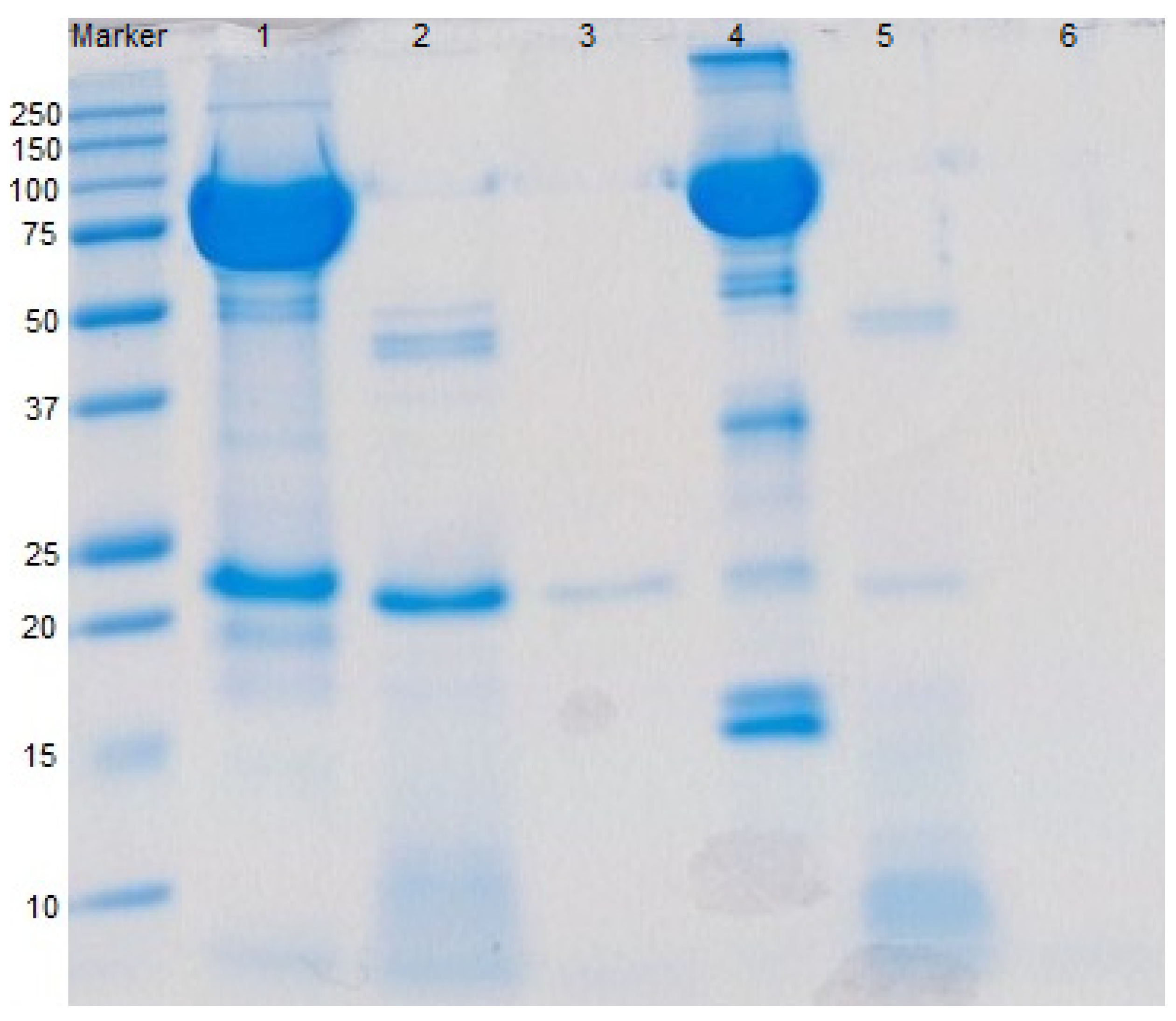
2.5. SDS-PAGE
2.6. Bacterial Strains
2.7. Determination of Antibacterial Activity
2.8. Statistical Analysis
3. Results
3.1. Fractionation of Whey Proteins from Deer Milk
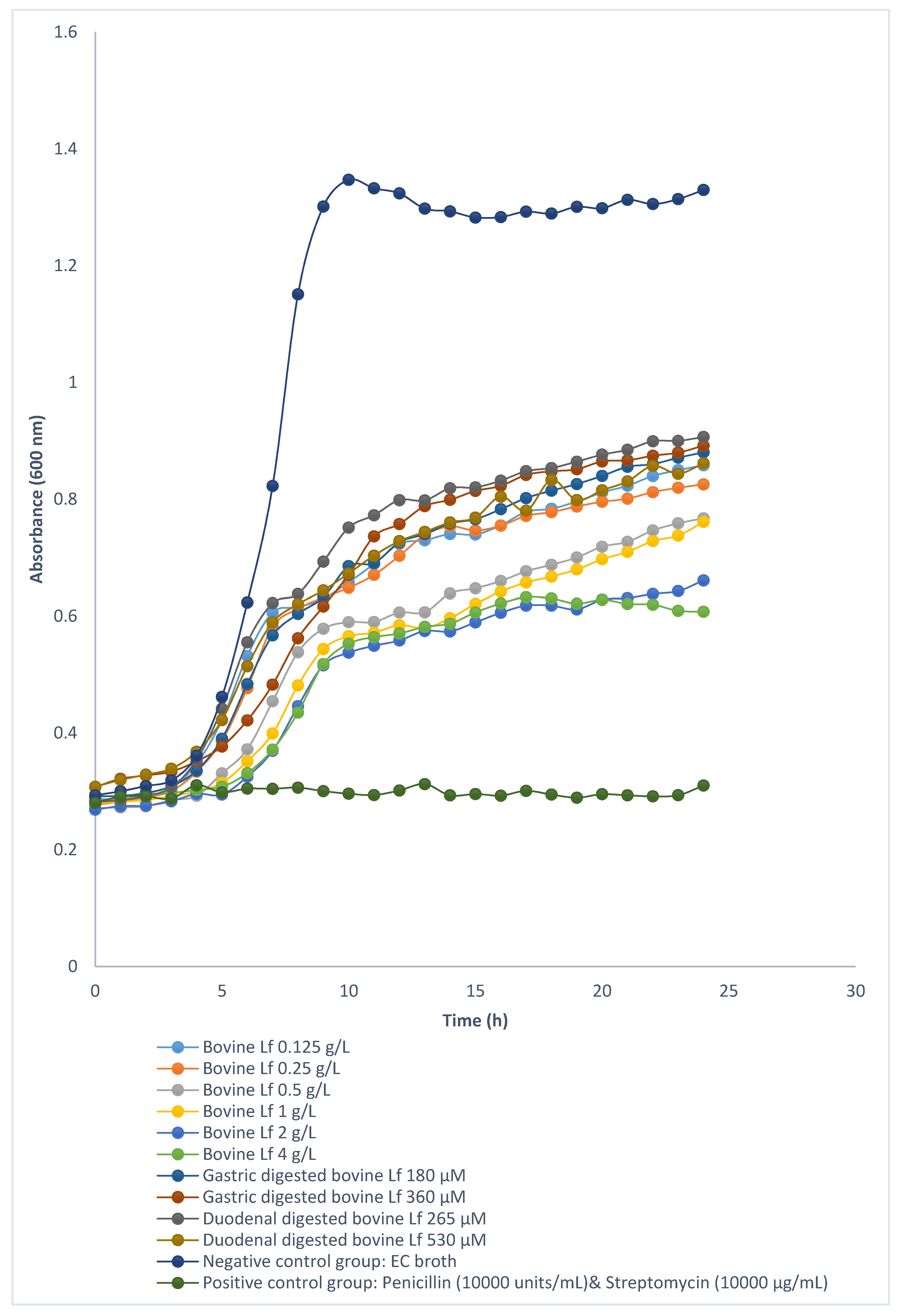
3.2. Digestibility of Deer and Cow Lf
3.3. Antibacterial Activities of Deer and Cow Lf and Their Hydrolysates
3.3.1. Antibacterial Activity of Deer and Cow Lf
3.3.2. Antibacterial Activity of Deer and Cow Lf Hydrolysates
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Lf | Lactoferrin |
| α-La | Alpha-lactalbumin |
| β-Lg | Beta-lactoglobulin |
| BSA | Bovine serum albumin |
| DSA | Deer serum albumin |
| E. coli ATCC 25922 | Escherichia coli ATCC 25922 |
| FPLC | Fast protein liquid chromatography |
| GMP | Glycocaseinomacropeptide |
| Ig | Immunoglobulin |
| L. acidophilus ATCC 4356 | Lactobacillus acidophilus ATCC 4356 |
| Lf | Lactoferrin |
| Lp | Lactoperoxidase |
| MBC | Minimun bactericidal concentration |
| MIC | Minimum inhibitory concentration |
| MRSc | MRS broth plus 0.05% cysteine |
| OPA | o-Phthaldialdehyde |
| S. aureus ATCC 25923 | Staphylococcus aureus ATCC 25923 |
| SDS-PAGE | Sodium dodecyl sulphate-polyacrylamide gel electrophoresis |
References
- Otago Daily Times. Available online: http://www.odt.co.nz/regions/north-otago/253360/potential-cheese-deers-milk (accessed on 21 November 2020).
- Smithers, G.W. Whey and whey proteins—From ‘gutter-to-gold’. Int. Dairy J. 2008, 18, 695–704. [Google Scholar] [CrossRef]
- Holand, O.; Gjostein, H.; Nieminen, M. Handbook of Milk of Non-Bovine Mannals; Wiley-Blackwell: Hoboken, NJ, USA, 2006; pp. 355–370. [Google Scholar]
- Wang, Y.; Bekhit, A.E.-D.A.; Morton, J.D.; Mason, S. Nutritional Value of Deer Milk. In Nutrients in Dairy and their Implications on Health and Disease; Elsevier BV: Amsterdam, The Netherlands, 2017; pp. 363–375. [Google Scholar]
- Goodman, R.E.; Schanbacher, F.L. Bovine lactoferrin mRNA: Sequence, analysis, and expression in the mammary gland. Biochem. Biophys. Res. Commun. 1991, 180, 75–84. [Google Scholar] [CrossRef]
- Lambert, L.A.; Perri, H.; Halbrooks, P.J.; Mason, A.B. Evolution of the transferrin family: Conservation of residues associated with iron and anion binding. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 142, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Bekhit, A.E.-D.A.; McConnell, M.; Mason, S.; Carne, A. Fractionation of whey proteins from red deer (Cervus elaphus) milk and comparison with whey proteins from cow, sheep and goat milks. Small Rumin. Res. 2014, 120, 125–134. [Google Scholar] [CrossRef]
- Opatha Vithana, N.L. A comparative study of immunomodulatory activity of deer and cow milk proteins. Ph.D. Thesis, Lincoln University, Canterbury, UK.
- García-Montoya, I.A.; Cendón, T.S.; Arévalo-Gallegos, S.; Rascón-Cruz, Q. Lactoferrin a multiple bioactive protein: An overview. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 226–236. [Google Scholar] [CrossRef]
- Korhonen, H.J.; Marnila, P. Milk Proteins | Lactoferrin. In Encyclopedia of Dairy Sciences; Elsevier BV: Amsterdam, The Netherlands, 2011; pp. 801–806. [Google Scholar]
- Ochoa, T.J.; Cleary, T.G. Effect of lactoferrin on enteric pathogens. Biochimie 2009, 91, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Gifford, J.L.; Hunter, H.N.; Vogel, H.J. Lactoferricin: A lactoferrin-derived peptide with antimicrobial, antiviral, antitumor and immunological properties. Cell. Mol. Life Sci. 2005, 62, 2588–2598. [Google Scholar] [CrossRef]
- Pilbrow, J.; Bekhit, A.E.-D.A.; Carne, A. Fractionation of sheep cheese whey by a scalable method to sequentially isolate bioactive proteins. Food Chem. 2016, 203, 165–174. [Google Scholar] [CrossRef]
- Mandalari, G.; Adel-Patient, K.; Barkholt, V.; Baro, C.; Bennett, L.; Bublin, M.; Gaier, S.; Graser, G.; Ladics, G.S.; Mierzejewska, D.; et al. In vitro digestibility of β-casein and β-lactoglobulin under simulated human gastric and duodenal conditions: A multi-laboratory evaluation. Regul. Toxicol. Pharmacol. 2009, 55, 372–381. [Google Scholar] [CrossRef]
- Hasselmann, C. Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, ix–xv. [Google Scholar]
- Conesa, C.; Sánchez, L.; Rota, C.; Pérez, M.-D.; Calvo, M.; Farnaud, S.; Evans, R.W. Isolation of lactoferrin from milk of different species: Calorimetric and antimicrobial studies. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Tavares, T.G.; Malcata, F.X. Whey proteins as source of bioactive peptides against hypertension. Bioact. Food Pept. Health Dis. 2013, 75–114. [Google Scholar]
- El-Sayed, M.M.H.; Chase, H.A. Simulation of the breakthrough curves for the adsorption of alpha-lactalbumin and beta-lactoglobulin to SP Sepharose FF cation-exchanger. Biochem. Eng. J. 2010, 49, 221–228. [Google Scholar] [CrossRef]
- MilkAMP Database. Available online: http://milkampdb.org/entrieslist.php?x_Native_Protein=Lactoferrin(LF) (accessed on 11 November 2019).
- Yen, C.-C.; Shen, C.-J.; Hsu, W.-H.; Chang, Y.-H.; Lin, H.-T.; Chen, H.-L.; Chen, C.-M. Lactoferrin: An iron-binding antimicrobial protein against Escherichia coli infection. BioMetals 2011, 24, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- Kieckens, E.; Rybarczyk, J.; Barth, S.; Menge, C.; Cox, E.; Vanrompay, D. Effect of lactoferrin on release and bioactivity of Shiga toxins from different Escherichia coli O157:H7 strains. Veter Microbiol. 2017, 202, 29–37. [Google Scholar] [CrossRef]
- Ochoa, T.J.; Brown, E.L.; Guion, C.E.; Chen, J.Z.; McMahon, R.J.; Cleary, T.G. Effect of lactoferrin on enteroaggregative E-coli (EAEC). Biochem. Cell Biol. Biochim. Biol. Cell. 2006, 84, 369–376. [Google Scholar] [CrossRef]
- Yekta, M.A.; Verdonck, F.; Broeck, W.V.D.; Goddeeris, B.; Cox, E.; Vanroy, D. Lactoferrin inhibits E. coli O157:H7 growth and attachment to intestinal epithelial cells. Vet. Med. 2010, 55, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Murata, M.; Wakabayashi, H.; Yamauchi, K.; Abe, F. Identification of milk proteins enhancing the antimicrobial activity of lactoferrin and lactoferricin. J. Dairy Sci. 2013, 96, 4891–4898. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.-W.; Jheng, T.; Shyu, C.-L.; Mao, F. Antimicrobial potential for the combination of bovine lactoferrin or its hydrolysate with lactoferrin-resistant probiotics against foodborne pathogens. J. Dairy Sci. 2013, 96, 1438–1446. [Google Scholar] [CrossRef]
- Udekwu, K.I.; Parrish, N.; Ankomah, P.; Baquero, F.; Levin, B.R. Functional relationship between bacterial cell density and the efficacy of antibiotics. J. Antimicrob. Chemother. 2009, 63, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Hoek, K.S.; Milne, J.M.; Grieve, P.A.; Dionysius, D.A.; Smith, R. Antibacterial activity in bovine lactoferrin-derived peptides. Antimicrob. Agents Chemother. 1997, 41, 54–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, M.; Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K. Potent Antibacterial Peptides Generated by Pepsin Digestion of Bovine Lactoferrin. J. Dairy Sci. 1991, 74, 4137–4142. [Google Scholar] [CrossRef]
- Bolscher, J.G.; Van Der Kraan, M.I.; Nazmi, K.; Kalay, H.; Grün, C.H.; Hof, W.V.; Veerman, E.C.; Amerongen, A.V.N. A one-enzyme strategy to release an antimicrobial peptide from the LFampin-domain of bovine lactoferrin. Peptides 2006, 27, 1–9. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration | Escherichia coli ATCC 25922 | Staphylococcus aureus ATCC 25923 | Lactobacillus acidophilus ATCC 4356 | |||
|---|---|---|---|---|---|---|
| Lag Time (h) | Generation Time (min) | Lag Time (h) | Generation Time (min) | Generation Time (h) | ||
| Deer Lf (g/L) | 0.125 | 3.00 ± 0.00 | * 34.12 ± 0.01 c | 4.67 ± 0.33 | 37.57 ± 0.09 | NA |
| 0.25 | 3.33 ± 0.33 | * 34.00 ± 0.07 d | 5.00 ± 0.00 | 37.70 ± 0.01 k | 10.43 ± 0.07 | |
| 0.5 | * 4.33 ± 0.33 | * 33.55 ± 0.02 e | 5.00 ± 0.00 h | 37.78 ± 0.01 l | 9.62 ± 0.20 | |
| 1 | * 7.00 ± 0.00 a | * 34.83 ± 0.10 f | 5.00 ± 0.00 i | 37.29 ± 0.02 m | * 13.28 ± 0.87 | |
| 2 | * 10.33 ± 0.33 b | * 34.68 ± 0.08 g | 5.33 ± 0.33 j | 37.07 ± 0.12 n | 12.50 ± 2.04 o | |
| Deer Lf hydrolysates (µM) | 140 gastric | * 5.67 ± 0.33 | * 35.24 ± 0.06 | NA | NA | NA |
| 280 gastric | MIC | MIC | 5.00 ± 0.00 | 37.11 ± 0.09 | NA | |
| 560 gastric | NA | NA | * 8.33 ± 0.33 | 37.99 ± 0.11 | NA | |
| 201 duodenal | * 4.67 ± 0.33 | * 35.47 ± 0.01 | NA | NA | NA | |
| 402 duodenal | MIC | MIC | 5.00 ± 0.00 | 36.99 ± 0.17 | NA | |
| 804 duodenal | NA | NA | * 8.00 ± 0.00 | 37.62 ± 0.05 | NA | |
| Cow Lf (g/L) | 0.125 | 3.00 ± 0.00 | * 35.27 ± 0.04 c | NA | NA | NA |
| 0.25 | 3.00 ± 0.00 | * 35.25 ± 0.01 d | 5.00 ± 0.00 | * 38.40 ± 0.01 k | 12.33 ± 1.00 | |
| 0.5 | 4.00 ± 0.00 | * 35.59 ± 0.02 e | * 6.33 ± 0.33 h | * 38.83 ± 0.09 l | * 12.99 ± 0.29 | |
| 1 | * 4.67 ± 0.33 a | * 35.77 ± 0.01 f | * 6.67 ± 0.33 i | * 38.76 ± 0.06 m | * 17.19 ± 6.60 | |
| 2 | * 5.00 ± 0.00 b | * 35.95 ± 0.01 g | * 7.33 ± 0.33 j | * 38.38 ± 0.10 n | * 17.04 ± 0.91 o | |
| 4 | * 4.67 ± 0.33 | * 35.86 ± 0.01 | * 9.33 ± 0.33 | * 38.17 ± 0.02 | * 15.72 ± 0.35 | |
| Cow Lf hydrolysates (µM) | 180 gastric | 3.33 ± 0.33 | * 35.11 ± 0.09 | NA | NA | NA |
| 360 gastric | 3.33 ± 0.33 | * 35.23 ± 0.16 | * 11.00 ± 0.00 | * 38.93 ± 0.28 | NA | |
| 720 gastric | NA | NA | 5.33 ± 2.33 | 38.00 ± 0.22 | NA | |
| 265 duodenal | 3.00 ± 0.00 | * 34.80 ± 0.05 | NA | NA | NA | |
| 530 duodenal | 3.00 ± 0.00 | * 35.16 ± 0.12 | * 7.00 ± 0.00 | * 38.77 ± 0.03 | NA | |
| 1060 duodenal | NA | NA | * 8.33 ± 0.33 | * 38.29 ± 0.02 | NA | |
| Broth control group | 3.00 ± 0.00 | 32.01 ± 0.01 | 4.00 ± 0.00 | 37.46 ± 0.01 | 9.38 ± 0.17 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Bekhit, A.E.-D.A.; Mason, S.L.; Morton, J.D. Lactoferrin Isolation and Hydrolysis from Red Deer (Cervus elaphus) Milk and the Antibacterial Activity of Deer Lactoferrin and Its Hydrolysates. Foods 2020, 9, 1711. https://doi.org/10.3390/foods9111711
Wang Y, Bekhit AE-DA, Mason SL, Morton JD. Lactoferrin Isolation and Hydrolysis from Red Deer (Cervus elaphus) Milk and the Antibacterial Activity of Deer Lactoferrin and Its Hydrolysates. Foods. 2020; 9(11):1711. https://doi.org/10.3390/foods9111711
Chicago/Turabian StyleWang, Ye, Alaa El-Din A. Bekhit, Susan L. Mason, and James D. Morton. 2020. "Lactoferrin Isolation and Hydrolysis from Red Deer (Cervus elaphus) Milk and the Antibacterial Activity of Deer Lactoferrin and Its Hydrolysates" Foods 9, no. 11: 1711. https://doi.org/10.3390/foods9111711