3.1. Composition of Feeding Substrates
Proximate compositions of the feeding substrates are reported in
Table 1. As expected, lipid, protein, mineral, and fiber contents of middlings (S2) were significantly higher than wheat flour (S1), due to the presence of cereal grain parts other than endosperm (germ, aleurone layer). The olive pomace showed the following composition: 4.51% crude protein, 19.78% crude fat, 35.60% fiber, 31.43% NFE, 8.69% ash, on DM basis. Incorporation of increasing percentages of olive pomace (feeds S3–S5) resulted in a significant increase in moisture (from 8.14% to 53.16%), fiber (from 9.54% to 31.45% DM), and fat (from 5.73% to 7.44% DM), while crude protein percentage decreased from 16.84% to 11.76%, on DM basis. Feed S5 showed the highest moisture, fat, fiber, and ash levels, while feed S2 had the highest protein percentage and feed S1 the highest NFE percentage and energy value.
Table 2 reports the FA composition of total lipids extracted from feeding substrates. FA composition of control feeds (wheat flour, S1; middlings, S2) were in the range of previously published data [
23]. Unsaturated fatty acids (UFAs) were the most abundant in all feeds, which accounted for 80%–83% of total FAs, because of oleic acid (91%–94% of total monounsaturated fatty acids, MUFAs) and linoleic acid (87%–93% of total polyunsaturated fatty acids, PUFAs) amounts. The inclusion of olive pomace increased the oleic acid relative percentage, from 19.26%–21.79% (S1, S2) to 59.98% (S5), and decreased the amount of linoleic acid, from 59.58%–60.43% (S1, S2) to 22.32% (S5), while changes of total saturated fatty acid (SFA) amounts were limited. A decrease of linolenic acid was also observed, as inclusion level of olive pomace increased, thus resulting in non-significant changes of the n-6/n-3 ratio of feeding substrates.
Feed proteins had a total amount of essential amino acids (EAA) ranging from 36.79% (S1) to 44.34% (S5) (
Table 3). Middlings proteins had a higher EAA/NEAA (non-essential amino acids) ratio than wheat flour proteins, in agreement with the higher nutritional value of the middlings substrate. EAA/NEAA ratio was not affected by the substitution of the base feed (middlings) with increasing percentages of olive pomace. In all feeds, the most represented EAA were LEU, THR, MET and PHE, while the most abundant non-essential amino acids (NEAA) were GLU and PRO, which accounted for 26%–45% of total amino acids.
3.2. Composition of Tenebrio Molitor Larvae
Proximate composition of
T. molitor larvae reared on five different feeding substrates are listed in
Table 4. Larvae mirrored the proximate data of their substrates. In fact, larvae L5 showed the highest moisture, fiber, and ash levels; larvae L2 showed the highest protein percentage; larvae L1 had the highest NFE percentage and energy value. The moisture percentage of larvae (60.50%–68.48%) was close to values reported by van Broekhoven et al. [
9], for larvae fed with mixture of organic by-products, and higher than water contents reported by Ghaly et al [
24], Siemianowska et al. [
25], and Alves et al. [
14], for mealworms fed whole wheat flour and brewer’s yeast, oat flakes, soybean and bocaiuva flours, respectively. Protein content observed in larvae receiving 100% middlings (L2) was higher than mealworms analyzed by Ravzanaadii et al. [
26] (50.14% vs. 46.44% DM, respectively), which were only fed wheat bran. Larvae reared on 1:1 middlings/olive-pomace mixture (L4) also showed higher protein level (47.58% DM) than mealworms grown on flours mixture (wheat, soybean, bocaiuva pulp) (44.83%) and on mixtures of by-products from food manufacturing (watermelon, eggshells, banana peels), as reported by Alves et al. [
14] and Tan et al. [
15], respectively. Only experimental feeds made up of 95% whole wheat flour and 5% brewer’s yeast [
24] allowed
T. molitor larvae to reach a protein content as high as observed in the present study. Organic wheat flour (feed S1) was the best substrate to achieve the highest (40.10% DM) crude lipid content of mealworm, despite the higher fat content of feeds S2–S5. No significant differences in the fat content among larvae fed 100% middlings (L2) and pomace enriched middlings (L3–L5) were observed. Lipid levels agreed with values reported by Ravzanaadii et al. [
26], Alves et al. [
14], and Tan et al. [
15]. No significant effect of the feeding substrate was detected for the fiber content of the five groups of larvae. Significant differences were registered for total soluble carbohydrates (NFE), ranging from 4.87% DM in mealworm fed S1 feed to 13.39% DM in larvae reared on 75% olive pomace substrate (L5). Tan et al. [
15] found NFE values less than half in mealworms grown on feeds made up of common food wastes. Total mineral levels (ash) observed in the five mealworm groups were similar to those reported by Ghaly et al. [
24], Siemianowska et al. [
25], Alves et al. [
14], and Tan et al. [
15] for larvae grown on different feeding substrates.
The FA compositions of larvae lipids are reported in
Table 5. UFAs (oleic and linoleic acids) were the most abundant in larvae lipids, while palmitic acid was the most representative SFA. FA composition of mealworm samples agreed with previously published data [
5,
9,
10,
11,
12,
14,
22,
25,
26,
27,
28,
29]. Significant differences were observed between the fat composition of larvae fed control substrates: mealworms grown on organic wheat flour (L1) exhibited higher oleic acid percentage and lower levels of PUFA (linoleic and α-linolenic acids) than larvae collected from middlings (L2), while lipids of larvae grown on feed S2 had the lowest percentages of oleic acid and the highest levels of PUFAs (linoleic and α-linolenic acids) and palmitic acid. The inclusion of pomace in the feed composition did not affect the FA composition of body lipids. Besides, no correlations were observed between the crude lipid content of larvae and of their feeding substrates, according to experimental data published by Francardi et al. [
11], as a result of supplementation of feed with linseed. Dreassi et al. [
10] also noticed that, despite the different fat levels of six cereal-based substrates,
T. molitor larvae had the same fat percentage, whereas van Broekhoven et al. [
9] found that dietary fat affected larval fat content.
Some authors highlighted the possibility of modifying the FA composition, and hence the nutritional quality, of the insect lipids during the breeding [
10,
11]. However, the FA composition of yellow mealworm samples collected from experimental substrates S1–S5 did not reflect the FA composition of feeds. van Broekhoven et al. [
9] also observed that FA profile of mealworms grown on substrates composed of organic by-products of different origin (beer brewing, bread/cookie baking, potato processing, bioethanol production) did not follow the same trend of dietary FAs. Alves et al. [
14] reported that the addition of 50% of bocaiuva flour to control feed (50% wheat flour, 50% soybean flour) did not significantly affect the FAs of the larvae lipids, and Oonincx et al. [
8] highlighted that the fat composition of yellow mealworm seems to be fairly constant, in spite of n-6/n-3 PUFA ratio differences among feeds (various by-products derived from food manufacturing). This behavior might be attributed to the ability of yellow mealworm to de novo synthesize both linoleic and α-linolenic acids [
11,
30,
31], which are essential for humans and other mammalians.
The amino acid composition of mealworm proteins is reported in
Table 6. Larvae fed the five substrates showed a similar amino acid composition, with a total amount of EAA ranging from 48.71% to 53.69% and an EAA/NEAA ratio around 1.0. The most abundant EAA were THR (9.47%–11.14%), LEU (7.31%–8.49%), MET (7.62%–8.36%), and TYR (5.97%–8.40%), while the most represented NEAA were GLU (10.69%–11.38%), PRO (6.33%–10.7%7), ALA (7.89%–9.12%), and ASP (7.10%–8.11%). These values confirmed the well documented ability of insect proteins to meet the human requirement of essential amino acids [
32] and indicated that larvae fed on all tested substrates are high-value protein sources, especially samples L3 and L4 which were characterized by the highest EAA/NEAA ratio (1.16).
Protease inhibitors, specifically trypsin and chymotrypsin inhibitors, represent one of the most relevant antinutritional factors in legume seeds, as they are resistant to digestion and inhibit the activity of key pancreatic enzymes in the gut, thereby reducing digestion and absorption of dietary proteins [
33].
Table 7 reports on the presence of some anti-chymotrypsin activity in
T. molitor larvae reared on the different substrates: higher values were observed in L2 and L4 samples, even if no significant differences (Tukey test,
p < 0.05) were detected among groups. Furthermore, no inhibitory activity of trypsin was detected. However, the measured activities were about one order of magnitude lower than described in legume seeds [
34]. Therefore, the presence of this anti-nutritional factor in
T. molitor larvae did not represent a real safety problem. Notably, the inhibitory activity was resistant to heat treatment, as boiled extracts of larvae fully retained such activity (data not shown).
3.3. Insect Growth Performance
The availability of different nutrients affects important life history traits of insects, such as body weight, development time, and survival rate [
35]. In holometabolous insects, all growth occurs before metamorphosis so that the final weight of the last instar larva determines the size and performance of the adult [
36]. Moreover, pupal weight, which is strongly correlated with potential adult fecundity [
37], was regularly incorporated into mass rearing systems as a measure of larval dietary quality. Survival rate and development time were also indicators of dietary quality [
8].
The mean larval development time of experimental insects ranged between 98 (L2) and 133 days (L5), depending on feed (
Table 8) and in accord with previously published data [
8,
9]. The shortest values were measured for larvae reared on control feed (100% wheat middlings, L2) and on feed having 25% of olive pomace (L3). Further increase of olive pomace percentage in larvae feed lengthened their development time. The last larval instar weight ranged from 0.070 to 0.131 g. It was significantly higher in larvae fed on control substrates (L1, L2) and on 25% pomace enriched feed (L3) than in substrates made up of 50%–75% of olive pomace (L4, L5). Final weights of larvae L1–L3 were similar to those of larvae fed on different substrates containing 10.25%–25.25% protein, 4%–14.92% fat, and 19.4%–61.17% carbohydrates [
9,
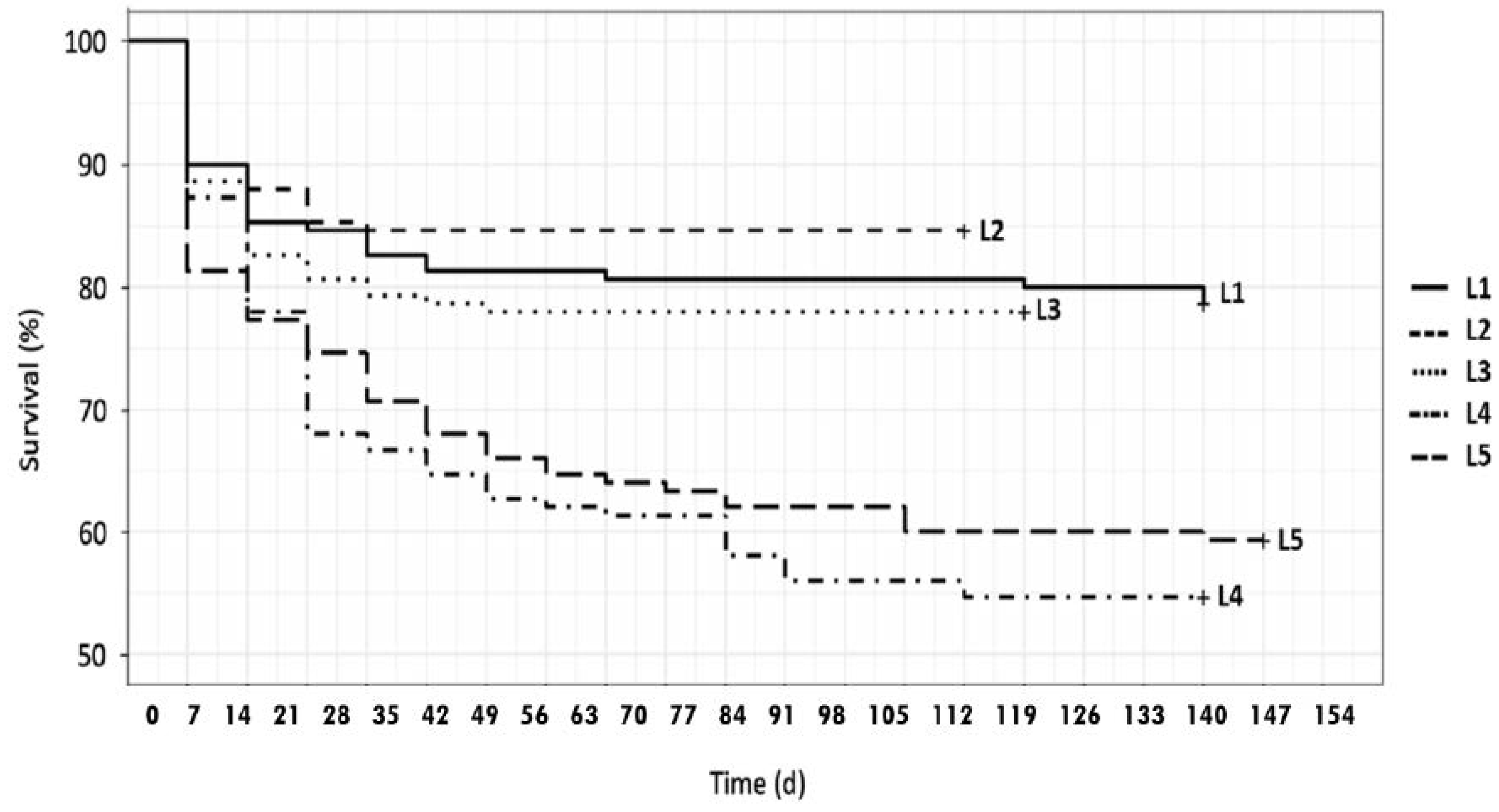
38]. Pupal weight ranged from 0.153 to 0.192 g depending on feed. It was significantly higher in larvae fed on S2 and S3 substrates than on the other rearing substrates. Larval survival rate ranged from 54% (L4) to 85% (L2). In particular, larvae fed on S1, S2 and S3 substrates showed significantly higher survival rates than larvae reared on highly pomace enriched media.
Kaplan–Meier survival curves (
Figure 1) for larvae fed on substrates S1–S3 were significantly different than feeds S4–S5 (S1–S4
p < 0.0001; S1–S5
p = 0.00072; S2–S4
p < 0.0001; S2–S5
p < 0.0001; S3–S4
p = 0.00017; S3–S5
p = 0.00211). Most of the larval mortality occurred within the first four weeks in all experiments, according to the behavior observed by Kim et al [
39].
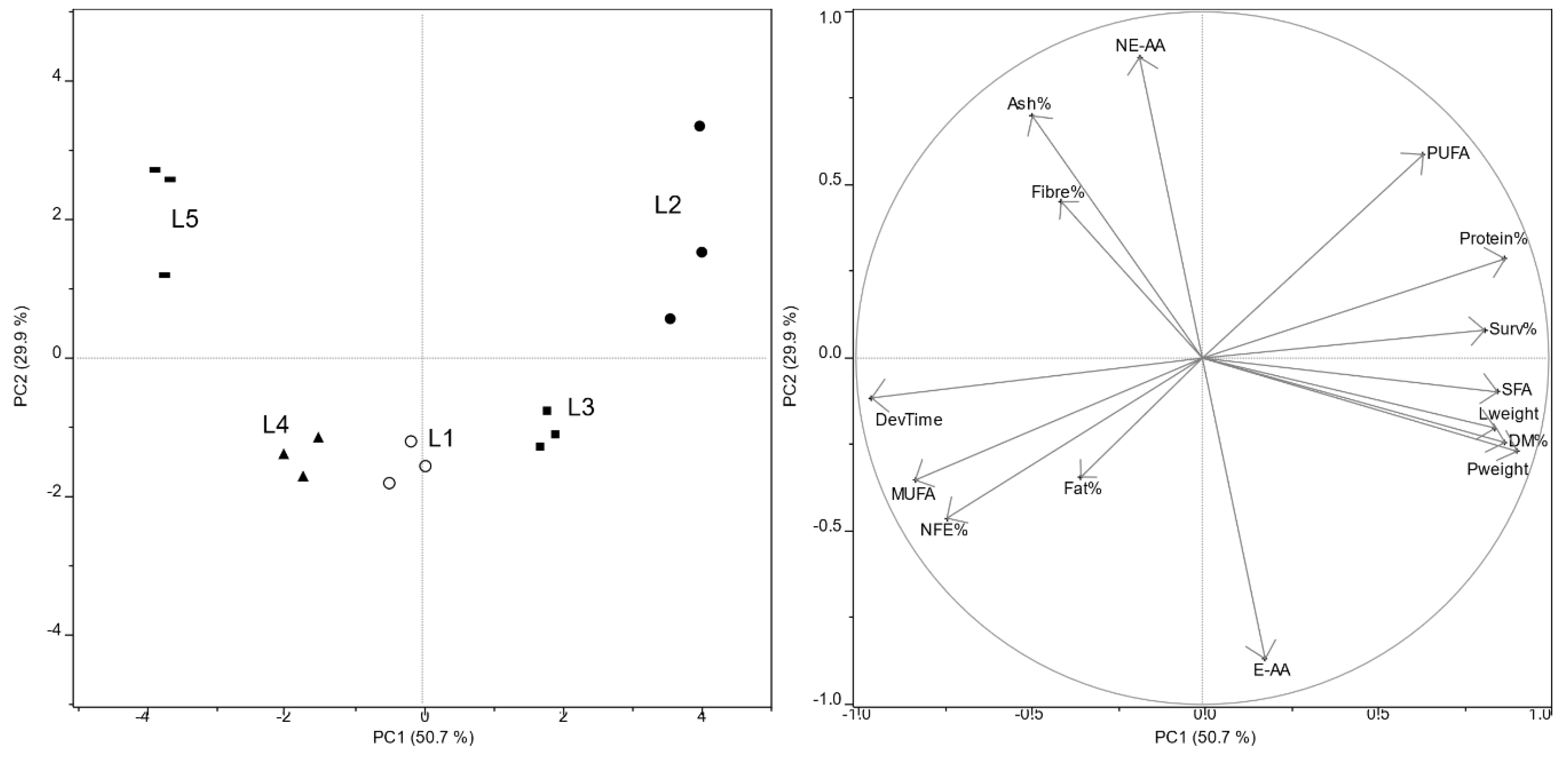
Mealworm larvae are able to self-select optimal nutrients from mixture of heterogeneous substrates in order to reach proper nutritional balance and guarantee best fitness [
40]. Even if some dietary self-selection could not be excluded, mass balance of the
T. molitor larvae reared on substrates S3 and S4 (
Table S2) showed that the total fat intake was greater than the fat contribution of middlings moieties. The FA composition of lipids extracted from the residual products of larvae fed on S3 and S4 (
Table S3) were much more similar to the FA composition of larval feeding substrates (
Table 2) than the FA profile of olive pomace. On the other hand, the residual product of larvae fed on S5 showed a lipid composition very close to olive pomace.
Protein and carbohydrate are the two major nutrients that affect larval growth performance and survival rate in insects [
41]. The carbohydrates/protein ratio affects development time and larval growth of
T. molitor [
40,
42]. Insects can synthetize lipids out of different dietary components such as protein and carbohydrates [
43,
44]. Mealworm larvae have been reared with success on a variety of feeding substrates with a fat range from 4% to 29% and a moisture up to 22% [
9,
10,
14,
45]. In our experiment, mealworm larvae performed better in feeds (S2, S3) having more than 50% carbohydrates, more than 15% proteins and a fat range from 5.73% to 6.18% (
Table 1). Furthermore, the presence of moisture in the diet is very important for the performance of
T. molitor [
46,
47,
48], although it inhabit water-deficient environments too. In our experiments, L4 and L5 fed on diets containing more than 22% of moisture showed the worsts grow performance and survival rate. It could also be hypothesized that the presence of high level of polyphenolic compounds affected the growth performance of mealworm larvae fed on S4 and S5. In facts, phenols are able to form complexes with proteins, thus affecting protein digestibility [
49], amino acid absorption and assimilation and resulting in a decreased larval growth and survival rate [
50,
51].


 ,
, 



{kind=link}
{kind=link}