
Asparagus Fructans as Emerging Prebiotics
by
 , , , , and
, , , , and
Amel Hamdi
1,2,
Isabel Viera-Alcaide
1,
Rafael Guillén-Bejarano
1,
Rocío Rodríguez-Arcos
1,
Manuel Jesús Muñoz
2,
Jose Manuel Monje Moreno
2 and
Ana Jiménez-Araujo
1,* 1
Instituto de la Grasa, Consejo Superior de Investigaciones Científicas (CSIC), Pablo de Olavide Universitary Campus, Building 46, Carretera de Utrera Km 1, 41013 Seville, Spain
2
Molecular Biology and Biochemical Engineering Department, Centro Andaluz de Biología del Desarrollo (CABD), University Pablo de Olavide (UPO), CSIC/UPO/JA, Carretera de Utrera Km 1, 41013 Seville, Spain
*
Author to whom correspondence should be addressed.
Foods 2023, 12(1), 81; https://doi.org/10.3390/foods12010081
Submission received: 18 November 2022
/
Revised: 16 December 2022
/
Accepted: 20 December 2022
/
Published: 23 December 2022
(This article belongs to the Special Issue Extraction, Chemistry and Utilization of Active Ingredients from Agri-Food Wastes and Byproducts)
Abstract
:Commercial fructans (inulin and oligofructose) are generally obtained from crops such as chicory, Jerusalem artichoke or agave. However, there are agricultural by-products, namely asparagus roots, which could be considered potential sources of fructans. In this work, the fructans extracted from asparagus roots and three commercial ones from chicory and agave were studied in order to compare their composition, physicochemical characteristics, and potential health effects. Asparagus fructans had similar chemical composition to the others, especially in moisture, simple sugars and total fructan contents. However, its contents of ash, protein and phenolic compounds were higher. FTIR analysis confirmed these differences in composition. Orafti®GR showed the highest degree of polymerization (DP) of up to 40, with asparagus fructans (up to 25) falling between Orafti®GR and the others (DP 10–11). Although asparagus fructan powder had a lower fructan content and lower DP than Orafti®GR, its viscosity was higher, probably due to the presence of proteins. The existence of phenolic compounds lent antioxidant activity to asparagus fructans. The prebiotic activity in vitro of the four samples was similar and, in preliminary assays, asparagus fructan extract presented health effects related to infertility and diabetes diseases. All these characteristics confer a great potential for asparagus fructans to be included in the prebiotics market.
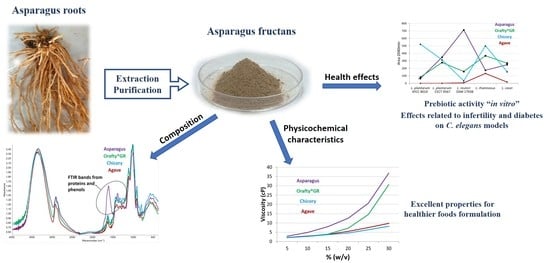
1. Introduction
Fructans, present in nature as storage polysaccharides in plants, are widely used in the food, nutraceutical and pharmaceutical industries. They are basically made up of fructose units linked to the glucose moiety of a sucrose molecule. Due to structural differences, these polymers can be classified into five groups: (i) inulin-type fructans, based on a 1-kestose structure; (ii) levan-type fructans, from a 6-kestose molecule; (iii) graminans, with branched inulin or levan structure; (iv) inulin neoseries, synthesized from a neo-kestose molecule, and (v) mixed-type levans, from a neo-levan structure. Fructans have applications as functional food and as prebiotic nutrients [1,2]. They are resistant to digestion in the upper portion of the human intestinal tract due to the β configuration of anomeric C-2 but they can be fermented by some beneficial colonic bacteria, popularly known as probiotics [3]. This fermentation process brings beneficial health effects, such as enhancement in the intestinal absorption of calcium, magnesium and iron [4], improvement in obesity prevention or control, blood glucose level, and lipid metabolism by the production of short-chain fatty acids [5], stimulation of the immune system [6] and reduction in the chances of cancer and allergic reactions [7].
In addition to their interesting nutritional and health benefits, fructans are also used in food formulations for their techno-functional properties such as a replacement for sugar in low-calorie food [8]. Due to their gelling capacity, they can easily be used as a fat substitute in food preparations. The product formulated with fructans instead of fat has a similar mouthfeel, with an appropriate creamy and consistent texture. Considered dietary fiber, they also have a low-calorie content, about 1–1.5 kcal per gram [9]. Fructans have synergistic effects with most gelling agents, e.g., gelatin, alginate, k- and i-carrageenans, gellan gum and maltodextrins. In addition, they also work as foam and emulsion stabilizers. Therefore, they can be included in the formulation of aerated desserts, ice creams, table spreads and sauces, replacing synthetic additives. Related to their gelling characteristics, fructans have humectant properties, affecting boiling and freezing points and reducing water activity. In this way, they can also act as an antimicrobial agent [10].
Due to their wide distribution in the plant kingdom and their varied applications in food technology, the extraction, isolation and characterization of fructans are gaining interest. The chemical characteristics of fructans depend on the species and also vary with the environmental conditions and developmental stage of plants [11]. Currently, chicory root (Cichorium intybus L.) is the main feedstock for the industrial production of inulin in Europe, with 15–20% of inulin on a fresh weight basis. Jerusalem artichoke (Helianthus tuberosus L.) and agave (Agave tequilana Weber) are also considered natural sources of inulin in other world zones, especially in North America. Jerusalem artichoke and its by-products contain a large amount of fructans (11–14% fresh weight basis) [12] and the remaining feedstock is utilized for the production of ethanol [13]. Fructans are also extracted from agave (higher than 37% on a fresh weight basis), which is often utilized as a bulking agent and can serve as a sugar replacement [14].
Recent studies conducted by Viera-Alcaide et al. [15], Weerasingha et al. [16], Sun et al. [17] and Mudannayake et al. [8] found that the roots and the rhizome of some Asparagus species such as, A. officinalis, A. falcatus, A. racemosus and A. cochinchinensis have the potential to be used as alternative inulin sources. The global cultivation area of different Asparagus species is higher than that of chicory and Jerusalem artichoke [8,16,18]. Compared to the commercial fructan sources, A. falcatus, A. racemosus and A. officinalis have shown 17.74, 11.83 and 15.3% of inulin, respectively, in the roots on a fresh weight basis [8,16,18].
The addition of fructans to different food systems is a worldwide practice. However, asparagus fructans and their prebiotic properties remain almost unknown. Weerasingha et al. [16] added A. officinalis fructan as an ingredient in the production of yogurt and concluded that its addition did not change the nutritional quality of this probiotic food. After a sensory analysis, asparagus fructan-added yogurts scored higher than plain yogurts in all the evaluated attributes and also improved their physicochemical characteristics. Moreover, the prebiotic properties of asparagus fructans were stated in this same study, since the viability of yogurt probiotic bacteria (Bifidobacterium bifidum, Lactobacillus bulgaricus ssp. bulgaricus and Streptococcus thermophilus) increased proportionally to the fructan concentration during the storage period. In another study [17], a purified fructan from the roots of A. cochinchinensis was subjected to in vitro fermentation by human fecal microbiota. As the main physicochemical effects, a decrease in the pH of the culture medium was detected, together with an increase in the short-chain fatty acid concentration, especially that of acetic, propionic, i-valeric and n-valeric acids. Some changes were also observed in the microbiota composition after 24 h fermentation: the beneficial genus Prevotella, Megamonas, and Bifidobacterium increased proportionally, while the genus Haemophilus, related to respiratory and nervous diseases, decreased. These results pointed to a health-promoting effect linked to asparagus fructan consumption.
As recent research has demonstrated good perspectives for the use of asparagus fructans in food and nutraceuticals applications, and considering that an agro-food by-product such as roots and rhizomes from asparagus cultivation are the feedstock for their isolation, there is increasing interest in developing this alternative fructan source. In our previous work, we established the concept of roots and rhizomes of asparagus as a valuable source of fructans [15]. As a continuation, this paper aimed to study the chemical composition, physicochemical characteristics, and in vitro prebiotic activity of the asparagus root fructans and three commercial ones in order to verify the capability of asparagus fructans in contrast to others currently on the market, and to determine their potential health effects using Caenorhabditis elegans as a model organism.
2. Materials and Methods
2.1. Plant Material
Underground asparagus organs were harvested during autumn, 2019, at the end of the harvest season, at different asparagus exploitation fields in Huétor-Tájar, Granada (Spain). The rhizomes and roots were sent to the Instituto de la Grasa labs, washed to remove soil and other foreign material, and left to dry at room temperature. The samples were chopped with an IKRA Mogatec shredder model EG 2500 in order to obtain a homogeneous raw material for fructan extraction and frozen at −20 °C.
2.2. Fructan Extraction from Asparagus Roots
Fructans were extracted as previously described [15]: 500 g of frozen sample were homogenized with 4 L of boiling water and placed in a shaking water bath set at 80 °C and 60 rpm for 30 min. Afterward, the sample was filtered through filter paper and the slurry was extracted again under the same conditions with 2 L of hot water. Both filtrates were mixed and stored at −20 °C.
2.3. Purification of Asparagus Fructans by Adsorption Chromatography
A glass column was filled with 100 mL (3 × 15 cm) of Diaion® HP20, a synthetic adsorbent resin of the highly porous type obtained from Vivaqua International S.L. (Barberà del Vallès, Spain). The resin was activated with 96% ethanol and then equilibrated with distilled water. In total, 1 L of extract was loaded into the column at a flow rate of 2-bed volumes/h and the column was washed with 200 mL water, 400 mL of 20% ethanol, and 400 mL of 80% ethanol. The non-retained fraction, water and 20% ethanol washes were put together, concentrated under vacuum and freeze-dried. The resultant pale-brown solid was considered to be purified asparagus fructans (PAF).
2.4. Commercial Fructan Sources
Orafti®GR from Beneo GmbH (Mannheim, Germany) was kindly supplied by Ferrer Alimentación S.A. (Barcelona, Spain). Chicory inulin from SaludViva (Elche, Spain) and agave inulin (OligofructineTM) from Tentorium Energy S.L. (Ulldecona, Spain) were purchased from local health food stores.
2.5. Determination of Chemical Composition
2.5.1. Moisture
Aliquots of PAF and commercial inulins (1 g × 3) were dried in an infrared moisture analyzer (Ohaus, MB45) until constant weight. The result is expressed as a g/100 g fresh weight.
2.5.2. Simple Sugar Composition
Solutions of 1 mg/mL of the different samples were analyzed in a Dionex (Sunnyvale, CA, USA) high-performance anion-exchange chromatograph (HPAEC) using a Carbopac PA-10 column (4 × 250 mm, 10 μm) in combination with a Carbopac PA guard column (4 × 50 mm, 10 μm) as described by Jaramillo-Carmona et al. [19]. The mobile phase consisted of 100 mM NaOH (eluent A) and 100 mM NaOH and 700 mM sodium acetate (eluent B). The elution conditions were as follows: 0–15 min, 100% A (re-equilibration); injection at 15 min and start acquisition at 16 min; linear gradient over 45 min to 25% A, 75% B. The flow rate was maintained at 0.9 mL/min. A Dionex pulsed electrochemical detector in the pulsed amperometric detection (PAD) mode was used. Calibration curves of standard glucose, fructose and sucrose were used for the quantification of mono- and disaccharides.
2.5.3. Ash
Ashes were determined by incinerating samples in a muffle furnace Nabertherm model B180 (Bremen, Germany) at 550 °C until white ash was obtained.
2.5.4. Proteins
The concentration of protein was determined as % nitrogen content × 6.25 by elemental microanalysis using a Leco CHNS932 analyzer (St. Joseph, MI, USA).
2.5.5. Fructans
The fructan content in each product was analyzed by the K-FRUC assay Kit from Megazyme (Bray, Ireland). The complete method is described in detail on the Megazyme website [20]. The results were expressed as g/100 g fresh weight product.
2.5.6. Phenols
Solutions of 10 mg/mL of each sample were analyzed by HPLC [21]. Briefly, the analyses were carried out using a Jasco-LC-Net II ADC liquid chromatograph system equipped with a diode array detector (DAD). Phenols were separated using a Mediterranea Sea C18 reverse-phase analytical column (25 cm length × 4.6 mm i.d., 5 μm particle size; Teknokroma, Barcelona, Spain). The gradient profile was formed using solvent A (water with 1% formic acid) and solvent B (acetonitrile with 1% formic acid): from 0% B to 20% B for the first 20 min, to 21% B over the next 8 min, maintained at 21% B for 2 min, and then to 30% B over the next 10 min, and to 100% over the next 5 min, and finally maintained at 100% B for 5 min. The flow rate was 1 mL/min and the column temperature was 30 °C. Spectra from all peaks were recorded in the 200–600 nm range and the chromatograms were acquired at 360 nm. The quantification of individual phenols was performed using an eight-point regression curve in the range of 0–250 μg on the basis of standards.
2.6. pH
Solutions of 100 mg/mL were made from PAF and other commercial samples to determine pH values. A pHmeter Mettler-Toledo (Barcelona, Spain) model FiveEasy was used for the measurements.
2.7. Antioxidant Activity
The antioxidant activity was measured against DPPH· free radical [22]. The efficient concentration (EC50), which represents the amount of antioxidants necessary to decrease the DPPH· initial absorbance by 50%, was calculated from a calibration curve by linear regression for each sample. The activity was expressed as millimoles of Trolox equivalent (TE) per kilogram of sample by means of a dose–response curve for Trolox.
2.8. Degree of Polymerization of Fructans
The samples were analyzed in a Dionex (Sunnyvale, CA, USA) high-performance anion-exchange chromatograph (HPAEC) under similar conditions to those described for simple sugar determination. The elution conditions were as described in Viera-Alcaide et al. [15]: 0–15 min, 100% A (re-equilibration); injection at 15 min and start acquisition at 16 min; linear gradient over 55 min to 20% A, 80% B. The flow rate was maintained at 0.9 mL/min. The identification of the different DP was made using Orafti®GR.
2.9. Viscosity
Viscosity measurements were taken with a Cannon-Fenske viscometer, series 150. A set of solutions of the four inulin samples (5–30% w/v) were prepared at 80 °C with continuous stirring. The viscosity assays were developed at three temperatures 80, 20 and 4 °C in a water bath with a thermostat. The time the sample took to pass between the two marks in the viscometer (×3), in seconds, is used to calculate the dynamic viscosity using the following formula:
where η is the dynamic viscosity, expressed in cP; C is the viscometer constant at the assay temperature; t is the time to pass in seconds; and ρ is the density of the sample at the assay temperature in g/mL.
η = C × t × ρ
2.10. Oil Holding Capacity (OHC)
OHC was measured as previously described [23], with some modifications: 500 mg of each sample, in triplicate, were weighed into 15 mL centrifuge tubes and 5 g of sunflower oil was added to each tube. The tubes were stirred and left to settle for 60 min. Afterward, the samples were centrifuged in a Digtor 22-R centrifuge from Orto-Alresa (Daganzo de Arriba, Spain) set at 3122× g, 20 °C, for 20 min. After centrifugation, the tubes were placed upside down for 30 min to rinse the non-retained oil. The tubes were then weighed and the retained oil was calculated by weight difference. OHC was expressed as g oil/100 g inulin.
2.11. Color
The color determinations of the four inulin samples were measured using a color measurement spectrophotometer BYK-Gardner, Color-view model (Columbia, MD, USA) set for Hunter L* (lightness), a* (redness), b* (yellowness).
2.12. Fourier-Transform Infrared (FTIR) Spectroscopy
Around 2 mg of each sample was milled carefully with 200 mg of KBr using a clean and dry mortar and pestle. The dry and milled powders were pressed into discs in a Graseby hydraulic press (Specac, Orpington, UK) fitted with a 13 mm evacuable pellet die [24]. The clear KBr-fructan discs, once loaded into the holder, were exposed horizontally to the IR beam in a Bruker vertex 70 FT-IR spectrometer (Bruker Optics, Ettlingen, Germany) equipped with a deuterated triglycine sulfate (DTGS) detector. The experiments were run in duplicate to ensure consistency of the results. All the spectra were collected in the range of 5000–400 cm−1 by co-addition of 50 scans and a resolution of 4 cm−1 using weak Norton–Beer apodization [25]. The spectra were collected and manipulated with OPUS version 7.2 (Bruker Optics, Ettlingen, Germany). The differences between the FTIR spectra obtained were studied with Omnic 7.3 (Thermo Electon Inc., Madison, WI, USA).
2.13. Prebiotic Effect In Vitro
The prebiotic activity of the different fructan sample was evaluated against five probiotic Lactobacillus strains: L. plantarum ATCC 8014 (American Type culture collection, Manassas, VA, USA), L. plantarum CECT 9567 (Colección Española de Cultivos Tipo, Valencia University, Valencia, Spain), L. reuteri DSM 17,938 (Casen Recordati S.L., Madrid, Spain), L. rhamnosus, and L. casei. The culture media were MRS, which replaced the carbohydrate source for the different fructans at 2% (MRS+Asparagus, MRS+Orafti®GR, MRS+Chicory, MRS+Agave). As a control for each sample, the different strains were also incubated in MRS but only with the amount of simple sugar that comes with each fructan sample. The presented results are the difference between both assays, to discuss only the growth due to fructan fermentation. Two more control were included, MRS + 2% glucose and MRS without any carbon source [26]. In each microplate well, 10 μL of inoculum and 340 μL of medium were dosified to reach a final concentration of 106 cfu/mL. The optical density (OD) at 600 nm was measured every half an hour in a SPECTROstar spectrophotometer (BMG Labtech, Ortenberg, Germany). The software includes the Reader Control and MARS data analysis interfaces, which served to measure the area under the growth curve for 36 h. All the assays were performed in triplicate.
2.14. C. elegans Assays
2.14.1. C. elegans Strains and Maintenance Conditions
The C. elegans strains used in the study were obtained from CGC (Caenorhabditis Genetics Center, University of Minnesota, Minneapolis, MN, USA): CB1370: daf-2(e1370ts)III and GMC101: dvIs100[Punc-54::amyloid-ß1-42:3′UTR unc-54 + Pmtl-2::GFP]II. Nematode growth medium (NGM) agar plates containing Agar N◦1 (2.5% w/v, Oxoid Limited, Basingstoke, UK), Pepton N-Z-Soy(R) BL4 (0.25% w/v, Sigma Aldrich, St.-Louis, MO, USA), NaCl (0.3% w/v), cholesterol (0.0005% v/v), CaCl2 (1 mM), MgSO4 (1 mM), K2HPO4/KH2PO4 (25 mM, pH 6.0) were seeded with OP50 as a food source for all the strains, and the living temperature was set at 16 °C. The asparagus fructans extract stock solutions (6, 7 mg/mL in ultrapure water) were prepared, filtered with 0.22 µm (Syringe-driven Filters membrane CA) and stored at −20 °C. The utilization of water as a substitute for asparagus fructans extract was the control group.
2.14.2. Paralysis Assay in GMC101 Strain
Transgenic C. elegans strain GMC101 was synchronized by treating gravid hermaphrodites with alkaline hypochlorite solution [27]. The eggs obtained were shaken (120 rpm) at 16 °C overnight in M9 buffer (22 mM KH2PO4 42 mM Na2HPO4 86 mM NaCl 1 mM MgSO4) to allow hatching. The next day, L1 animals (200 L1/plate) were transferred to fresh NGM agar plates (60 mm diameter) seeded with OP50 and treated with 100 µL of fructans extract (168 µg/mL) or with water (control) and allowed to reach L4 stage at 16 °C. The synchronized L4 was transferred to fresh NGM plates spotted with OP50 containing either fructans extracts or water (100 µL) (n = 60 worms/group, 30 worms/plate). The worms were incubated at 25 °C to induce amyloid-β aggregation for 16 h. The amyloid-β peptides are aggregated in the muscle cells of GMC101 strain, which causes paralysis in the mutants. The number of paralyzed worms was scored under the microscope. To identify the paralysis, each worm was gently touched with a platinum wire. Worms were considered to be paralyzed if they moved neither spontaneously nor in response to three prods on the head, at least one time their full body length.
2.14.3. Determination of Progeny Production in daf-2(e1370)
Synchronized L1 worms (200 L1/plate) were transferred to fresh NGM agar plates (60 mm diameter) spotted with OP50 containing either 100 µL fructans extracts (168 µg/mL) or water (control) and were allowed to develop to the late L4 stage (around 2.5 days) at 16 °C. This low temperature was chosen to avoid dauer formation in the temperature-sensitive daf-2(e1370) mutant. At L4 stage (around 2.5 days later), thirty worms were transferred to small fresh NGM agar plates (35 mm diameter) spotted with OP50, also with or without 50 µL of fructans extracts (168 µg/mL) (n = 30 worms/group, 1 worm/plate), the worms were incubated for 72 h at 25 °C. The progeny production was determined by quantifying the number of progenies hatched out of eggs. This assay was performed at least three times independently.
2.15. Statistical Analysis
All samples were analyzed at least in triplicate. To assess the differences among samples, a multiple-sample comparison was performed using the Statgraphics® Plus program Version 2.1. The level of significance was p < 0.05.
GraphPad Prism 9 (Version 9.0a) was used to analyze the data from C. elegans. Experiments yielding quantitative data for statistical analysis were performed independently at least three times, all with similar results. Micrograph images shown in the figures are representative of three independent experiments, all with similar results for progeny production assay and the average of three independent experiments for paralysis assay.
3. Results and Discussion
3.1. Asparagus Inulin Yield and Chemical Composition
As described in the Methods section, 500 g of roots and rhizomes from asparagus plants were treated with 4 L of water and then a second extraction was carried out with 2 L of water. Both filtrates were pooled (4.8 L) and subjected to a purification step by adsorption chromatography. This step was necessary because asparagus roots are also a potential source of steroidal saponins [21,28,29,30], which must be removed from root extracts. This saponin extract also has its own interest for its technological and functional characteristics [28,30,31]. After purification, fructan-containing fractions were concentrated under a vacuum and freeze-dried. The complete process was repeated eight times at least and an average yield of 17.51 ± 1.95 g/100 g fresh roots was determined (48.57% dry weight). The obtained residue (PAF) was a dry light-brown powder that was compared with other commercial inulins in order to know its possible technological and dietetic applications.
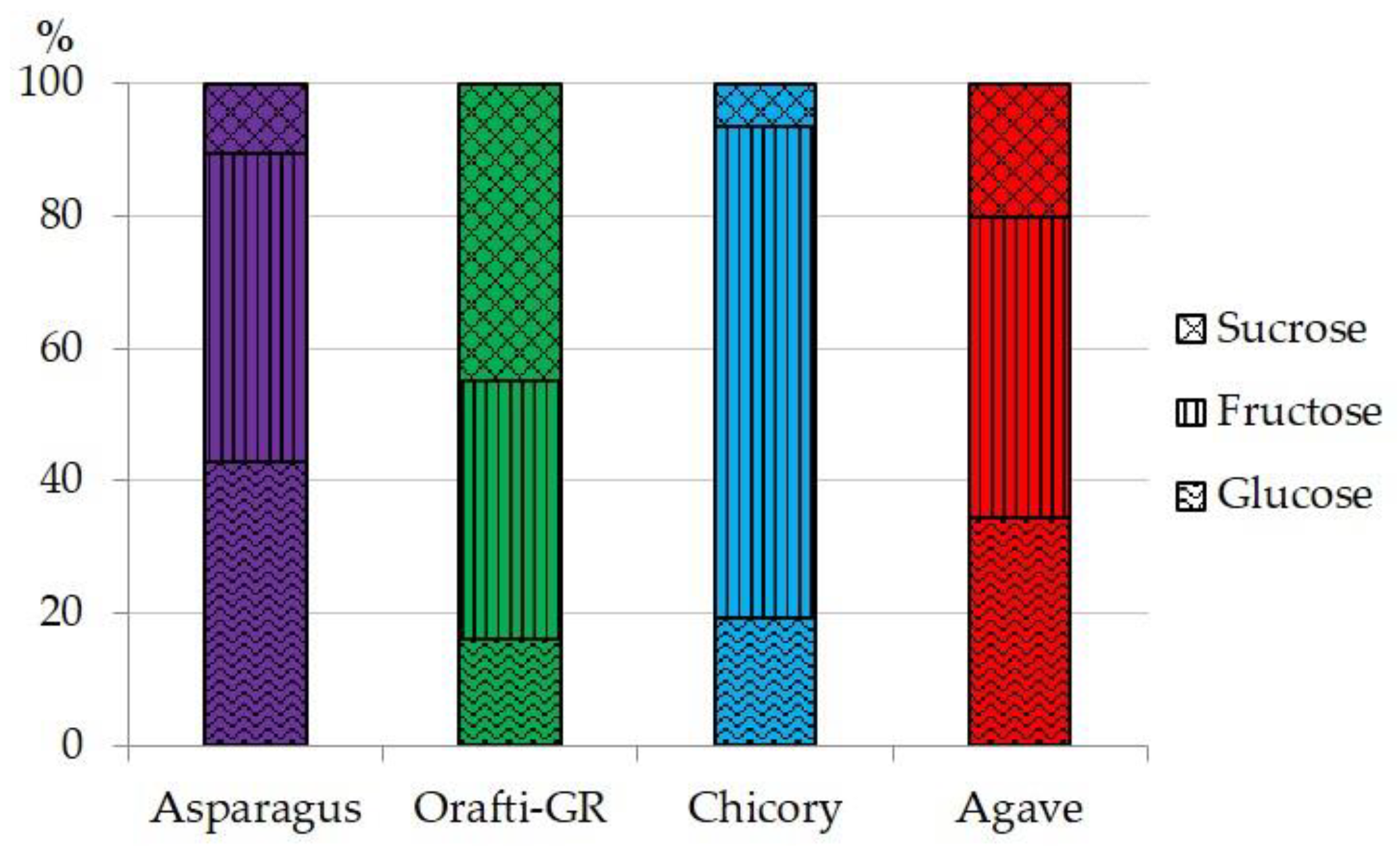
PAF and the commercial samples were analyzed for their content in moisture, simple sugars, ash, protein and fructan and the results are presented in Table 1. Although the last step for PAF obtaining was freeze-drying, 9% of its moisture remained. This content was the highest of the four samples analyzed, with the inulin from chicory at 6%, and that from agave and Orafti®GR at 4.6%. The high content of moisture could be related to the protein content in PAF, as the good water-holding capacity of proteins is widely known [8]. The amounts of simple sugars (glucose, fructose and sucrose) are very important, especially from a dietetic point of view. Orafti®GR had the lowest contents and the inulin from chicory showed the highest. PAF had twice the sugar content of Orafti®GR, around 4%, but half the amount of sugar was quantified in the other commercial samples. The composition of simple sugars is presented in Figure 1. The inulin from asparagus and agave were in similar proportions with similar amounts of glucose and fructose and only small amounts of sucrose. Sucrose was the major component in Orafti®GR, and fructose was the most abundant in the sample from chicory. Considering fructans as healthy food ingredients, it is important to maintain the level of simple sugar as low as possible, as fructans are often used as sugar replacements in diabetic and low-calorie foods [32]. Even more important than sugar content is the fructan content in the PAF and other commercial samples. Two groups could be clearly established from the four samples analyzed: Orafti®GR and inulin from agave, which have nearly 80% fructan content, and PAF and inulin from chicory, which were around 60%. From a dietetic point of view, the ratio of fructan/simple sugar should be as high as possible. This value was the highest for Orafti®GR (39.25), followed by PAF and agave (13.83 and 11.44, respectively), and finally chicory (6.95) inulins.
Therefore, we have a 48.57% yield on asparagus root dry weight basis and 58% fructans in PAF. With these percentages, we can assume that asparagus roots contain around 28% fructans on a dry weight basis. Chicory roots, Jerusalem artichoke tubers and agave stems, the usual industrial sources of fructans, have a much higher yield of fructans as expressed in dry weight: 63–80% for chicory [33,34]; 65–84% for Jerusalem artichoke [35,36], and 38–73% for agave [37,38]. However, other published works on agricultural by-products such as fructan sources reported lower amounts for this component, e.g., 16–28% in artichoke waste [39], around 5% in onion by-product [40] and 1.5% in cashew apple [41]. The extract obtained from artichoke waste contained 60–80% fructans, and that of onion, around 10%. If additional purification steps were applied to PAF, its fructan percentage would increase. Ion exchange chromatography can be implemented for ash and protein removal [8], and also treatment with calcium hydroxide and phosphoric acid for deproteinization [42]. Considering these facts, asparagus roots and rhizomes are similar to artichoke waste, the best agricultural by-product described as a fructan source in the bibliography, and its industrial exploitation could increase the income of asparagus spear producers.
Phenolics are also present in PAF (Table 2) but not in the other samples and, as a consequence, PAF also showed antioxidant activity. Chelidonic acid was the major component, followed by caffeic acid and caffeoyl-glycoside. Plants containing chelidonic acid are widely used in folk medicine as anti-inflammatory agents [43,44,45], with therapeutic potential for allergic disorders [43], mild analgesic, antimicrobial, oncostatic and sedative [44], and even in ethnoveterinary uses [46]. Caffeic acid is commonly known as a natural healing agent with medicinal properties because it possesses antioxidant, anti-inflammatory, anticancer, and neuroprotective properties [47]. Therefore, the phenolic compounds present in PAF cannot be considered contaminants to be removed but as bioactive constituents with valuable activities.
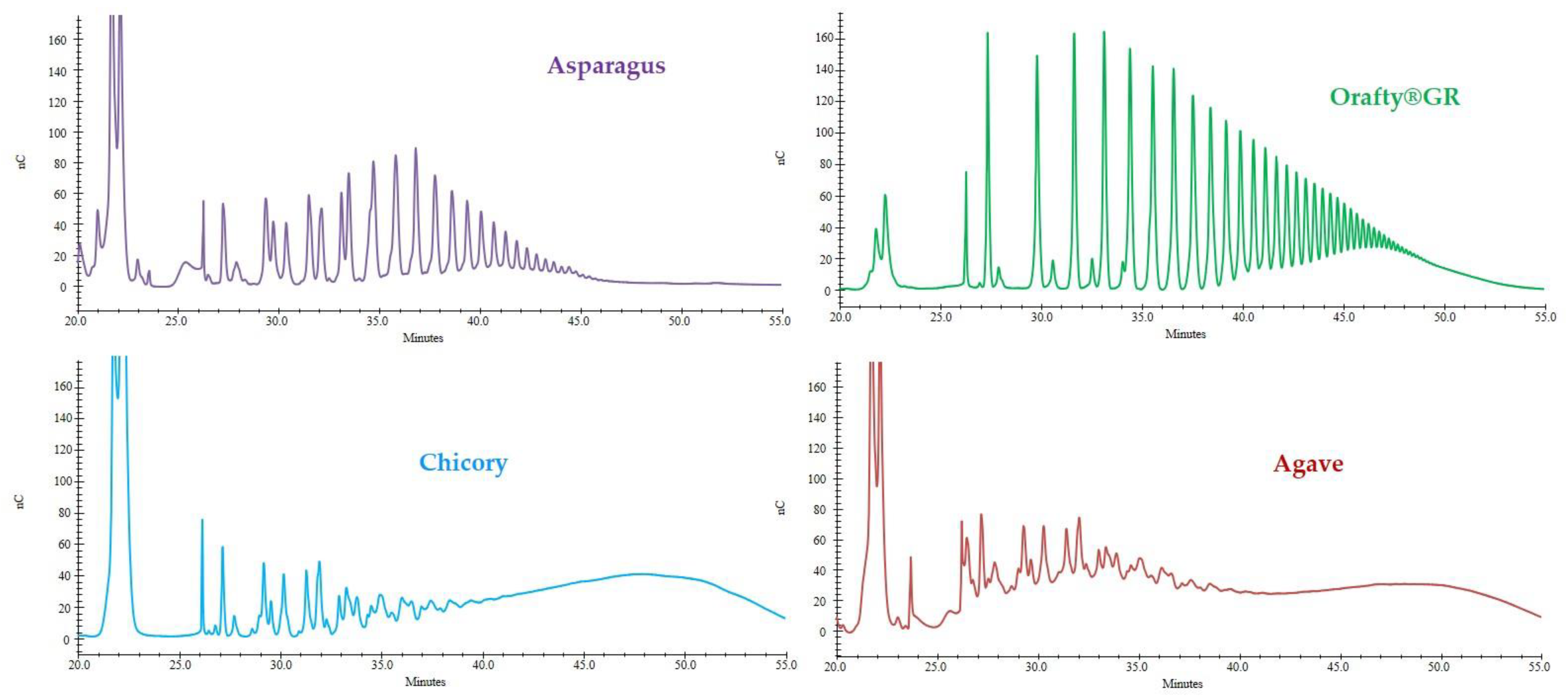
3.2. Degree of Polymerization of PAF and Commercial Inulins
The elution profiles of the different fructans analyzed by HPAEC are presented in Figure 2. PAF showed similar characteristics to Orafti®GR, and inulins from chicory and agave showed almost identical profiles. Orafti®GR, which is obtained from chicory roots, is widely used in food technology as a fat replacer in the formulation of baked goods, breakfast cereals, candy and chocolates, soups and sauces, dairy and meat products, etc. [48] without losing their creamy mouthfeel and providing low caloric content. Chicory fructans are of the inulin-series, linear polymers of fructose linked to the fructose unit of a sucrose molecule by β(2→1) linkage [49]. In Asparagales (asparagus and agave), apart from the inulin series, there is also a variable percentage of inulin-neo series (fructose is bonded β(2→6) to glucose in sucrose and β(2→1) on the fructose residue). In these plants, a great variety of isomers were described [15,50,51,52]. The case of agave is even more complex because this crop survives in a severe drought environment, which boosts the synthesis of all kinds of fructans [49].
In the present study (Figure 2), fructans from asparagus roots contained polymers of up to 25 sugar units but also a variety of iso-isomers were identified, as previously reported [15]. In Orafti®GR, polymers of higher molecular weight were found (up to 40 DP). In this case, the presence of iso-isomers was reduced to a minimum. The commercial inulins from chicory and agave looked very similar and were identified as oligomers of up to 10–11 DP. In fact, they should be labeled as “oligofructose” or “fructooligosaccharides” (FOS) rather than “inulin” because the term “inulin” refers to polymers with DP > 10 [53].
DP is a very important fructan characteristic because it marks the possible applications of the product. Oligofructose with DP ≤ 10 have some degree of sweetness (30–65% compared to sucrose), they are very soluble in water, not texturizing [54], but are easily and quickly fermented by colonic microbiota, with high prebiotic activity. However, the sweetness of inulins with DP > 10 is very low (0–30%), they produce viscous solutions in water [48], and are fermented more slowly in the gut. When ingested, oligofructose is fermented mainly in the proximal zone of the colon. On the contrary, inulin arrives at the distal portion where most chronic diseases occur and its beneficial prebiotic activity can be performed [55]. It is clear that fructan DP influences not only its technological applications but also its prebiotic characteristics.
3.3. Physicochemical Characteristics of PAF and Other Commercial Inulins
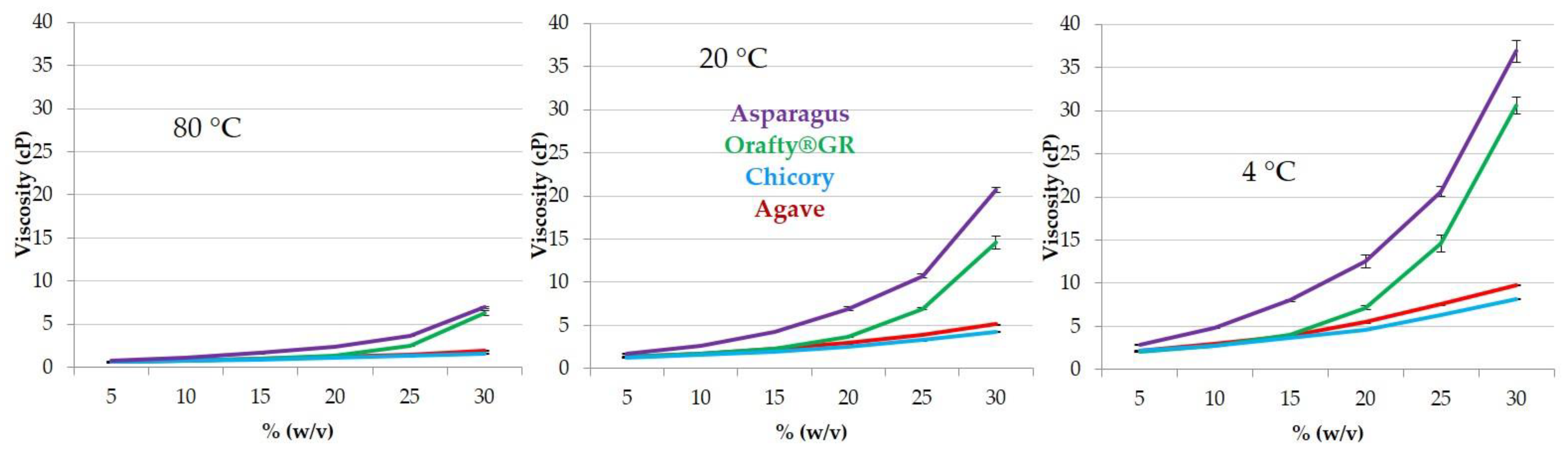
As mentioned above, viscosity is a key property for the use of inulins as fat replacers and it is directly linked to DP. In Figure 3, the viscosity of inulins from different origins at several concentrations and temperatures is presented. At the three tested temperatures, the behavior of samples was similar: in all cases, PAF and Orafti®GR showed close values, although PAF solutions were always the most viscous ones. Inulin from agave and chicory presented significant differences only at 4 °C. It is very clear that the correlation between DP and viscosity is that the lower the DP, the lower the viscosity.
Both factors, temperature and concentration, had a great influence on sample viscosity. For all samples, the decrease in temperature from 80 to 4 °C increased viscosity between 300 and 500 times (depending on concentration, lower concentrations corresponded to lower increases). However, the increase in concentration from 5 to 30% at the three temperatures resulted in differences among the samples. In chicory and agave, the viscosity increased 250–280 times at 80 °C, 340–380 times at 20 °C, and 380–440 times at 4 °C. When PAF and Orafti®GR were studied, greater increases were detected: 900–1000, 1000–1200 and around 1400 times, respectively, at 80, 20 and 4 °C. The effect of concentration was higher in samples with high DP than in those with low DP and was obviously also higher at lower temperatures.
Although the PAF extract had lower fructan content with lower DP than Orafti®GR, its viscosity results were always higher. This fact could be due to the presence of proteins in PAF (17.5%). Interactions such as charge–dipole, charge-induced dipole, and hydrogen bonds are mainly responsible for the viscosity of protein solutions [56]. Furthermore, it is widely known that the addition of sugars or polyols acts as a protein stabilizer by increasing the viscosity of the solution [57,58]. The presence of fructans and proteins in PAF resulted in more viscous solutions when compared to Orafti®GR. The creamy texture of both products at medium–high concentrations (>20%) and medium–low temperatures (<40 °C) allows for their application as fat replacers in multiple food formulations (ice creams, dressings, dairy products and juices) [59,60,61].
The results obtained for pH, OHC and color are presented in Table 3. There are significant differences in pH among the samples, between 4.6 and 6.4 units. These values are in the same range as another published previously [8,38]. OHC bundled the samples into groups: Orafti®GR and agave (around 79 g oil/100 g sample), and PAF and chicory with a significantly higher OCH (about 94 g/100 g). This fact could be related to the purity of the samples because those with lower purity (near 58% in fructans, Table 1) showed higher OHC results. The presence of other compounds such as proteins, simple sugars or phenols was likely to improve this property. Higher values were found for inulin isolated from Jerusalem artichoke tubers [62], up to 145 g oil/100 g product.
The color of PAF was clearly different from that of the other fructan sources (Table 3) because the asparagus extract was pale brown. The L* (lightness) value decreased and the b* value (yellowness) went up. The same happened for other unpurified fructans from natural sources such as Jerusalem artichoke tubers [62], Asparagus falcatus and Taraxacum javanicum plants [8].
3.4. FT-IR Spectrum for Different Inulins
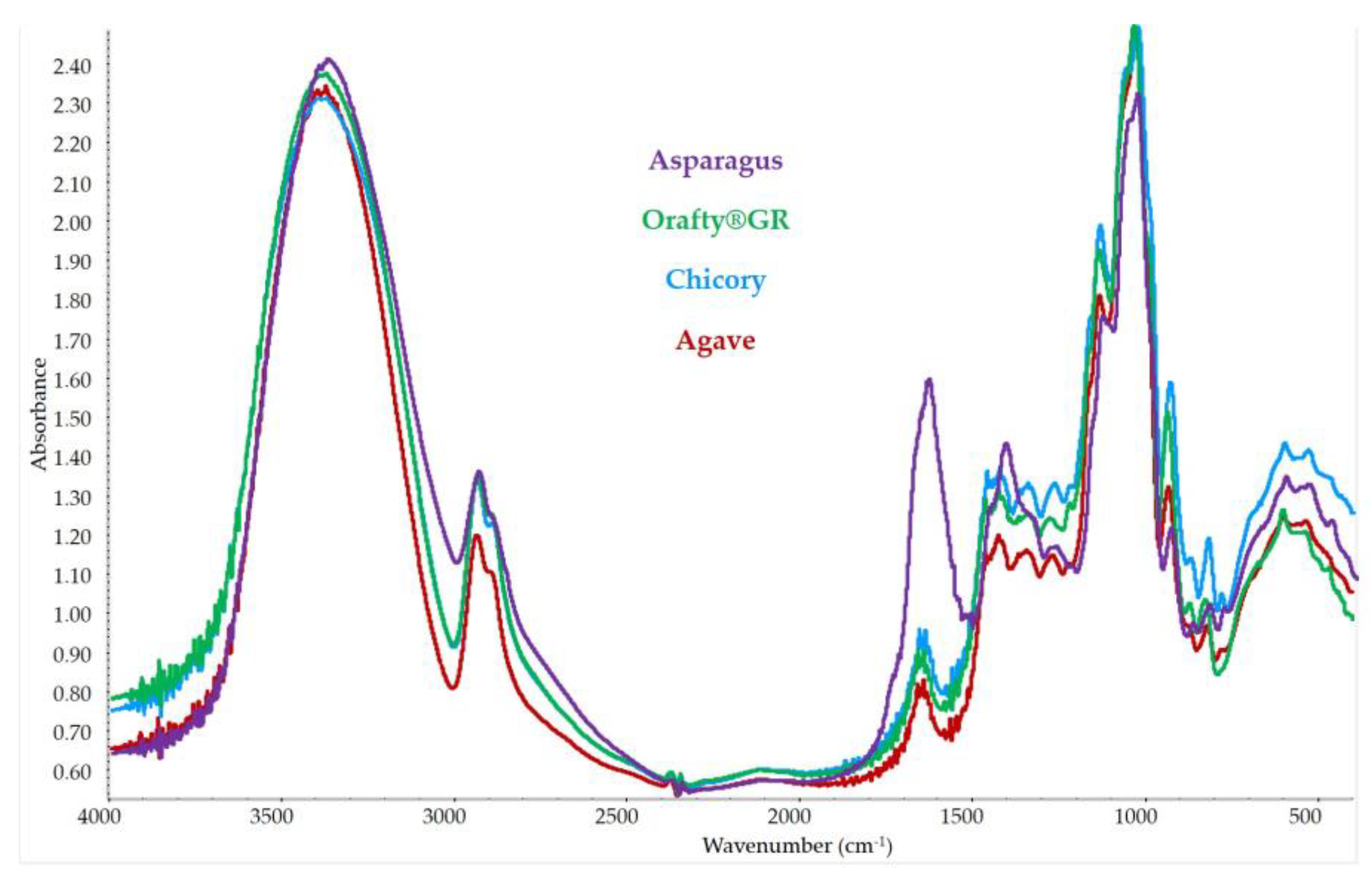
In Figure 4, the spectra of the different samples are presented. They presented the typical bands described previously for inulin [63]. The broad band between 3600 and 3200 cm−1 was due to the stretching of OH- groups from both carbohydrate and phenolic groups. The lower band at 3000–2800 cm−1 is attributed to C-H vibrations. In the spectral region of 1500–800 cm−1, there is another group of bands related to carbohydrates: 1500–1250 cm−1 for C-C-C stretching, and 1250–800 cm−1 for C-O stretching, C-O-H and C-O-C bending [38]. PAF showed a more intense band in this zone that could be due to bending and stretching vibrations in aromatic rings [64].
There was another band in the spectral region of 1700–1500 cm−1, where PAF looked different. A band was present in all samples in this zone but that of PAF was much higher than that of the other ones. In this zone, there was a band of C=O stretch, which is characteristic of inulin [8]. However, in this zone, there were bands assigned to proteins [38] (amide I and amide II vibrations) and to aromatic compounds (C-H and C=C-C aromatic bond stretching) [65]. The presence of both groups of compounds in PAF could explain this high-intensity band at 1700–1500 cm−1. These results supported the presence of purified fructans in the root extract of asparagus plants. The comparison of the PAF spectrum with those of other commercial fructans confirmed that all the samples had very similar compositions but with higher amounts of proteins and phenolics in asparagus roots.
3.5. Prebiotic Activity of Inulins In Vitro
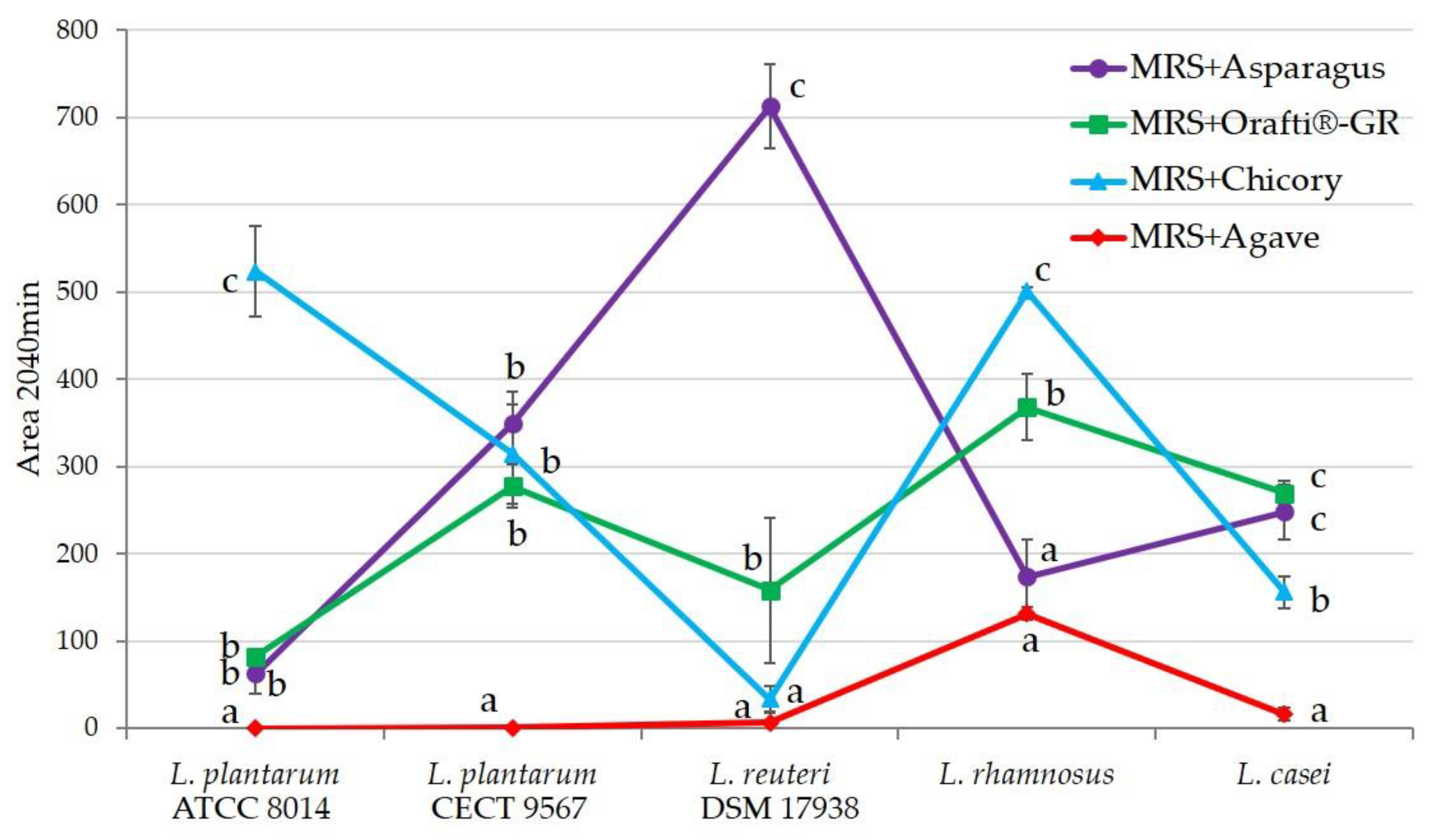
The obtained results are presented in Figure 5. Each Lactobacillus strain showed different behavior against the different fructans assayed. In general, fructans from agave were hardly fermented, only L. rhamnosus showed some capacity. The other three samples demonstrated prebiotic activity depending on the strains. L. reuteri, which was the strain with the lowest fermentation capacity, also showed the highest one on PAF. Orafti®GR was mildly fermented by almost all the strains, except for L. plantarum ATCC 8014. Fructans from chicory presented a very good fermentability with L. plantarum ATCC 8014 and L. rhamnosus. PAF showed excellent results with L. reuteri, low fermentability against L. plantarum ATCC 8014, and moderate against the other three strains.
This variability in the activity of the different prebiotic/probiotic pairings was previously described. Working with chicory inulin and its hydrolysis products (oligofructose), Roberfroid et al. [55] discussed that Bifidobacterium infantis optimally fermented both substrates, Bifidobacterium animalis only oligofructose, and Bifidobacterium bifidum any of them. Similar results were found when fructans from Agave angustifolia [38] and artichoke waste [39] were assayed against several Lactobacillus and Bifidobacterium species.
3.6. Effect of Fructans Extracts against Aβ-Induced Paralysis
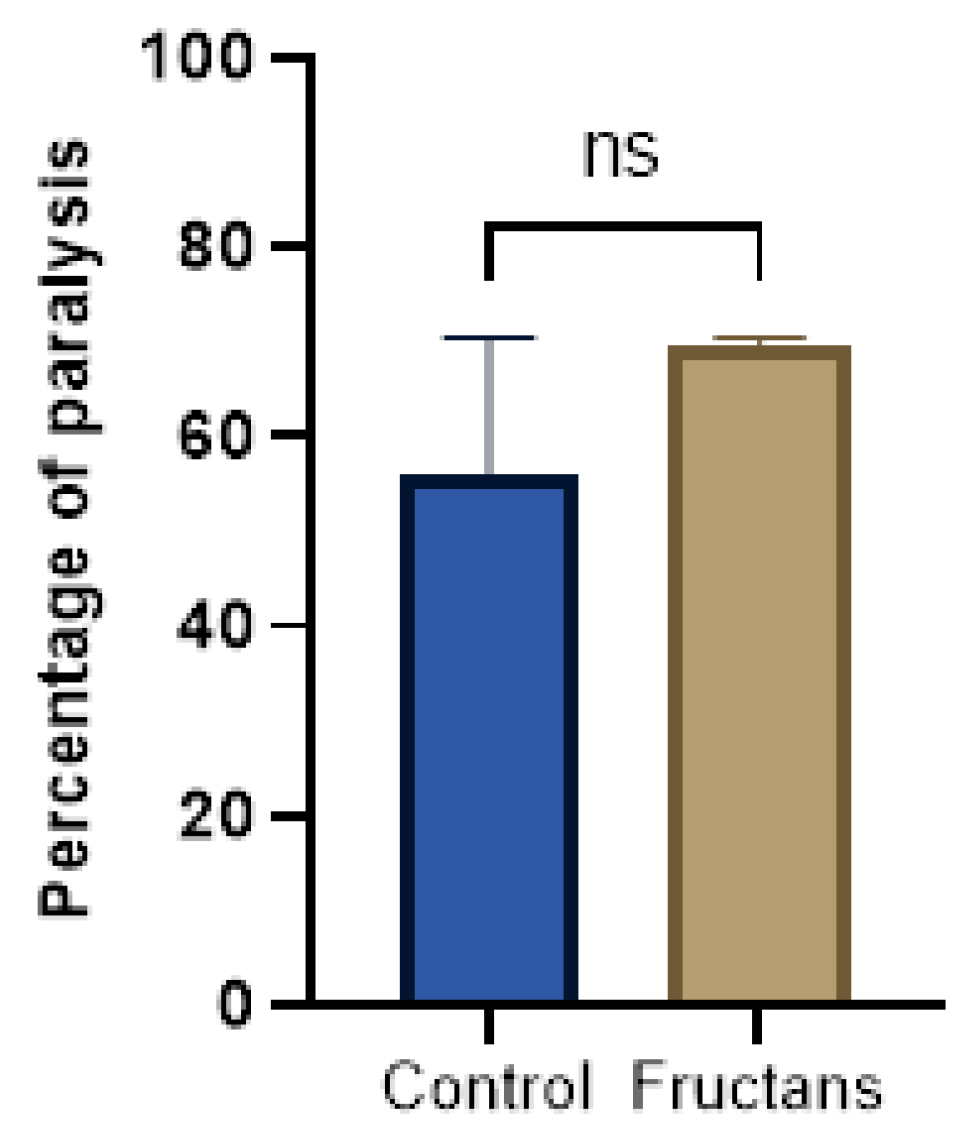
A paralysis test was performed to analyze any potential effect of the fructan extracts on the transgenic C. elegans model of Alzheimer’s disease. For this purpose, the GMC101 strain was used. This strain expresses the human amyloid-β peptide in muscle cells in a temperature-inducible way, which leads to a paralyzed phenotype. L1 GMC101 was grown on nematode growth medium with or without fructan extracts at 16 °C until they reach the L4 stage and then shifted to 25 °C to induce Aβ aggregation. Figure 6 illustrates the percentage of paralyzed worms at a non-permissive temperature (25 °C). The result showed that asparagus fructans at a concentration of 168 µg/mL do not ameliorate the paralysis phenotype.
The broad variety of chemical compounds found in plant natural products, which can be employed as sources for new Alzheimer’s disease therapies, may perhaps play a crucial role in drug discovery. Numerous natural compounds’ abilities to reduce amyloid-β toxicity were investigated in several C. elegans strains. Using the same strain of our study, a large variety of plant products was evaluated in these models which showed potential in reducing Aβ-induced paralysis such as triterpenoids and flavonoid glycosides from Dillenia suffruticosa leaves [66], flavonoids from sour Jujube seed [67], Danshen (Salvia miltiorrhiza) water extract [68], flavonoid glucosides from Ziziphus jujuba seeds [69] and so on. Nevertheless, few studies have explored the effect of soluble polysaccharides on toxicity and aggregation of amyloid-β in the transgenic C. elegans model of Alzheimer’s disease [70,71].
3.7. Effect of Fructan Extracts on the Fertility of the Mutant daf-2(e-1370)
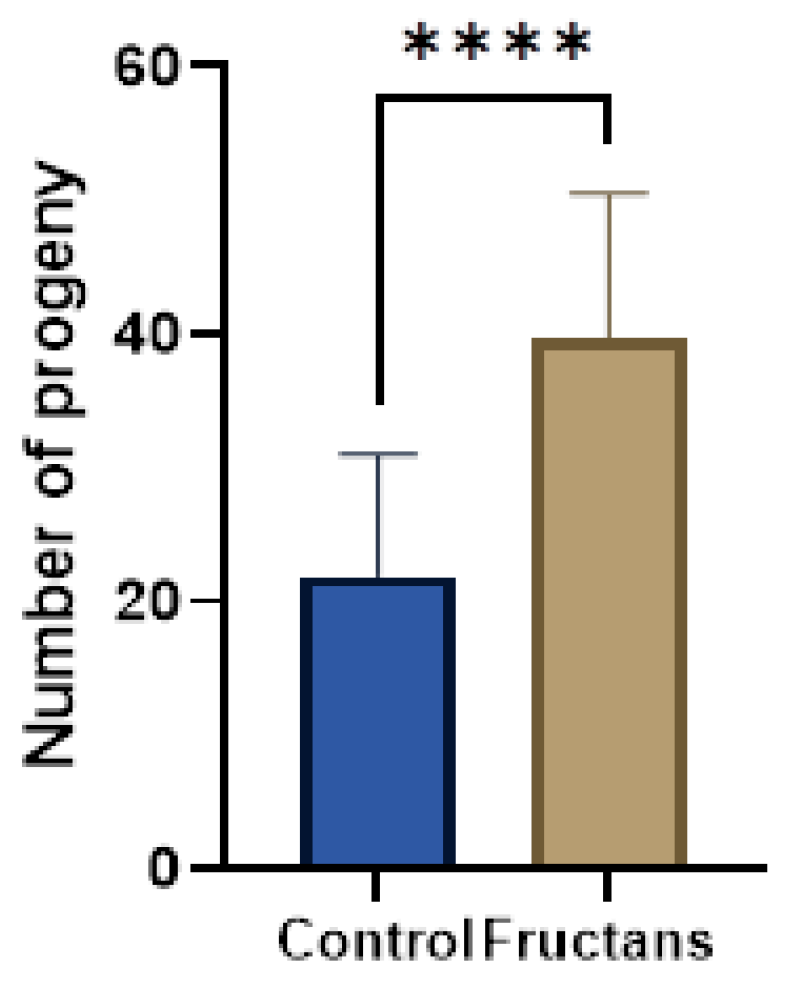
A crucial regulator of growth, development, and longevity is the insulin/IGF-1 signaling (IIS) pathway, which has been conserved throughout evolution [72]. In humans, high levels of insulin signaling are linked to tumor growth and cancer, whereas low levels of insulin signaling cause insulin resistance and diabetes [73]. Given its consequences, diabetes mellitus poses a serious risk to human health on a global scale [74]. All over the world, it is one of the top five most common causes of death [75]. Hyperglycemia and glucose intolerance lead to the development of a group of metabolic diseases known as diabetes mellitus. There are two types of diabetes mellitus that are frequently recognized. Insufficient insulin release from the pancreatic cells characterizes type-1 diabetes, whereas the emergence of insulin resistance in the body characterizes type-2 diabetes [76]. An excellent animal model for studying insulin-like signaling is the nematode C. elegans. Reduced or partial loss of function on the insulin pathway in C. elegans can alter stress responses or affect life span, among other pleiotropic effects [77]. Low or non-existent fertility in C. elegans were linked to mutations in the insulin signaling pathway [78]. This pathway is evolutionarily conserved in all metazoans and similarly to C. elegans, other organisms, including humans, also showed a correlation between reduced fertility and deficiencies in the activity of the insulin signaling pathway [79]. In the present work, we selected a C. elegans mutant strain named daf-2(e1370) affected in the human homolog of the insulin and insulin-like growth factor-I (IGF-I) receptors, which exhibits a severe reduction in fertility and altered embryonic development [80]. To prove the effects of asparagus fructans extracts on the fertility of daf-2(e1370), progeny numbers were quantified compared to control and represented in Figure 7 and Figure S1.
The results showed that asparagus fructan extracts double the progeny in this mutant, presenting a significant increase in the average descendent per worm (40 vs. 20). To the author’s knowledge, no research has been carried out on the improvement of daf-2(e1370) fertility by plants source fructans. In a previous work [81], the wild-type C. elegans treated with 100 µg/mL fructans from an exopolysaccharide secreted by Weissella cibaria lived 64% longer than the control under juglone oxidative stress. The forkhead transcription factor DAF-16, which is crucial for the insulin pathway, mediates this rise in oxidative stress resistance. Under normal conditions, the insulin pathway keeps DAF-16 inactive and outside of the nucleus. When there is an impairment of the insulin pathway, DAF-16 goes to the nucleus to control most of the insulin pathway-related phenotypes. Consequently, a mutation in daf-16 can suppress most of the known insulin pathway mutants’ phenotypes, including daf-2(e1370) infertility [82]. Paradoxically, the effect of fructans from Weissella cibaria and those from asparagus seems to be opposite in their relation with daf-16, but as far as we are looking at different phenotypes and different types of fructans, this point needs further investigation.
Insulin affects blood glucose levels in humans but it also plays a significant role in the reproductive system by interacting with insulin and insulin-like growth factor receptors in the brain and testis. Studies have shown that insulin plays crucial roles in spermatogenesis, prostatic growth, Sertoli cell proliferation and differentiation, testicular descent, and sexual behavior [83,84,85,86,87]. Infertility and impaired testis growth and maturation may also result from the absence of insulin receptors in GnRH neurons or Sertoli cells [88,89]. In addition, decreased levels of gonadotropins and testosterone, altered androgen-to-estrogen ratios, impaired semen parameters, and erectile dysfunction occurs in obese men with insulin resistance [90,91]. Interestingly, recent studies showed that inulin-type fructans improve glycemic control and moderate insulin resistance and lipid metabolism in prediabetic and type 2 diabetic patients [92,93].
4. Conclusions
Fructans are commercially obtained from chicory, agave and Jerusalem artichoke, depending on the production zone (Europe, South America and North America, respectively). However, asparagus underground organs are agricultural by-products that could be considered sources of fructans. As discussed in this work, a purified extract of asparagus roots could be regarded as a fructan extract with technological potential. Its composition and physicochemical characteristics are similar to other commercial fructans and show proven prebiotic activity. Therefore, asparagus fructans could be used as a fat replacement for medium–low-temperature foods (salad dressings, ice cream, pastry fillers, etc.) with its low sugar and calorie content. Due to its molecular weight and prebiotic activity, its use could provide in addition beneficial effects on colon proximal and distal portions. The preliminary results on its therapeutic possibilities opened a broad field for possible health applications. Changing the idea of asparagus roots as a troublesome agricultural by-product for that of a valuable feedstock could be a great goal for asparagus cultivation sustainability and a successful endeavor from the point of view of the circular economy.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/foods12010081/s1, Figure S1: Asparagus fructans extracts enhances fertility of the mutant daf-2(e-1370) (168µg/mL). The total number of progeny produced increased significantly following asparagus fructans extracts treatment. A: the second independent experiment and B: the third independent experiment. Graphs represent mean ± SD n = 30. ** p ≤ 0.01, *** p ≤ 0.001 using Two-tailed t-test.
Author Contributions
Conceptualization: A.H., I.V.-A., R.G.-B., R.R.-A., M.J.M., J.M.M.M. and A.J.-A.; methodology: A.H., R.G.-B., R.R.-A., M.J.M., J.M.M.M. and A.J.-A.; data analysis: R.G.-B., M.J.M., J.M.M.M. and A.J.-A.; data curation: R.G.-B., R.R.-A. and A.J.-A.; writing—original draft preparation: A.H., I.V.-A., J.M.M.M. and A.J.-A.; writing—review and editing: R.G.-B., R.R.-A., M.J.M., J.M.M.M. and A.J.-A.; project coordinator: R.R.-A. and R.G.-B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grant AGL2017-82428-R and PID2020-120463RB-I00 funded by MCIN/AEI/10.13039/501100011033 and by “ERDF A way of making Europe”; by grant PDC2021-121674-I00 funded by MCIN/AEI/10.13039/501100011033 by “European Union NextGenerationEU/PRTR”, and by European Union “NextGenerationEU” by the grant “Margarita Salas” for the Requalification of the Spanish university system 2021-2023 convened by the University Pablo de Olavide (Sevilla, Spain).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article or the Supplementary Material.
Acknowledgments
The authors gratefully thank Dámaso Hornero-Méndez and Diego L. García-González for their assistance in FT-IR analysis, and Belén Caballero-Guerrero for prebiotic activity assays.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Hughes, R.L.; Alvarado, D.A.; Swanson, K.S.; Holscher, H.D. The Prebiotic Potential of Inulin-Type Fructans: A Systematic Review. Adv. Nutr. 2022, 13, 492–529. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Shehzad, A.; Omar, M.; Rakha, A.; Raza, H.; Sharif, H.R.; Shakeel, A.; Ansari, A.; Niazi, S. Inulin: Properties, Health Benefits and Food Applications. Carbohydr. Polym. 2016, 147, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Apolinário, A.C.; de Lima Damasceno, B.P.G.; de Macêdo Beltrão, N.E.; Pessoa, A.; Converti, A.; da Silva, J.A. Inulin-Type Fructans: A Review on Different Aspects of Biochemical and Pharmaceutical Technology. Carbohydr. Polym. 2014, 101, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Lepczyński, A.; Herosimczyk, A.; Barszcz, M.; Ożgo, M.; Michałek, K.; Grabowska, M.; Tuśnio, A.; Szczerbińska, D.; Skomiał, J. Diet Supplemented Either with Dried Chicory Root or Chicory Inulin Significantly Influence Kidney and Liver Mineral Content and Antioxidative Capacity in Growing Pigs. Animal 2021, 15, 100129. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Nakanishi, N.; Miyoshi, T.; Okamura, T.; Hashimoto, Y.; Senmaru, T.; Majima, S.; Ushigome, E.; Asano, M.; Yamaguchi, M.; et al. Inulin Reduces Visceral Adipose Tissue Mass and Improves Glucose Tolerance through Altering Gut Metabolites. Nutr. Metab. 2022, 19, 50. [Google Scholar] [CrossRef]
- De Giani, A.; Sandionigi, A.; Zampolli, J.; Michelotti, A.; Tursi, F.; Labra, M.; Di Gennaro, P. Effects of Inulin-Based Prebiotics Alone or in Combination with Probiotics on Human Gut Microbiota and Markers of Immune System: A Randomized, Double-Blind, Placebo-Controlled Study in Healthy Subjects. Microorganisms 2022, 10, 1256. [Google Scholar] [CrossRef]
- Mauro, N.; Giammona, G.; Scialabba, C. Inulin for Cancer Therapy: Present and Perspectives. Int. J. Pharma Res. Rev. 2016, 5, 63–69. [Google Scholar]
- Mudannayake, D.C.; Wimalasiri, K.M.S.; Silva, K.F.S.T.; Ajlouni, S. Comparison of Properties of New Sources of Partially Purified Inulin to Those of Commercially Pure Chicory Inylin. J. Food Sci. 2015, 80, C950–C960. [Google Scholar] [CrossRef]
- Alaei, F.; Hojjatoleslamy, M.; Hashemi Dehkordi, S.M. The Effect of Inulin as a Fat Substitute on the Physicochemical and Sensory Properties of Chicken Sausages. Food Sci. Nutr. 2018, 6, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Gunenc, A.; Hosseinian, F. Developing Emulsion Gels by Incorporating Jerusalem Artichoke Inulin and Investigating Their Lipid Oxidative Stability. Food Prod. Process Nutr. 2020, 2, 2. [Google Scholar] [CrossRef]
- Benkeblia, N. Fructooligosaccharides and Fructans Analysis in Plants and Food Crops. J. Chrom. A 2013, 1313, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Cuenca, A.; Herrera-Vázquez, S.E.; Condezo-Hoyos, L.; Gómez-Ordóñez, E.; Rupérez, P. Inulin Extraction from Common Inulin-Containing Plant Sources. Ind. Crops Prod. 2021, 170, 113726. [Google Scholar] [CrossRef]
- Bhagia, S.; Akinosho, H.; Ferreira, J.F.S.; Ragauskas, A.J. Biofuel Production from Jerusalem Artichoke Tuber Inulins: A Review. Biofuel Res. J. 2017, 4, 587–599. [Google Scholar] [CrossRef]
- Bouaziz, M.A.; Rassaoui, R.; Besbes, S. Chemical Composition, Functional Properties, and Effect of Inulin from Tunisian Agave Americana L. Leaves on Textural Qualities of Pectin Gel. J. Chem. 2014, 2014, 758697. [Google Scholar] [CrossRef] [Green Version]
- Viera-Alcaide, I.; Hamdi, A.; Guillén-Bejarano, R.; Rodríguez-Arcos, R.; Espejo-Calvo, J.A.; Jiménez-Araujo, A. Asparagus Roots: From an Agricultural By-Product to a Valuable Source of Fructans. Foods 2022, 11, 652. [Google Scholar] [CrossRef]
- Weerasingha, V.; Wimalasiri, K.M.S.; Jayasumana, L. Enhanced Probiotic Activity, Physicochemical and Sensory Properties of Set-Yoghurt Incorporated with Asparagus Officinalis Inulin. J. Diet. Food Technol. 2020, 1, 1–6. [Google Scholar] [CrossRef]
- Sun, Q.; Zhu, L.; Li, Y.; Cui, Y.; Jiang, S.; Tao, N.; Chen, H.; Zhao, Z.; Xu, J.; Dong, C. A Novel Inulin-Type Fructan from Asparagus Cochinchinensis and Its Beneficial Impact on Human Intestinal Microbiota. Carbohydr. Polym. 2020, 247, 116761. [Google Scholar] [CrossRef]
- Singh, R.S.; Dhaliwal, R.; Puri, M. Production of High Fructose Syrup from Asparagus Inulin Using Immobilized Exoinulinase from Kluyveromyces Marxianus YS-1. J. Ind. Microbiol. Biotechnol. 2007, 34, 649–655. [Google Scholar] [CrossRef]
- Jaramillo-Carmona, S.M.; Javier Tejero-Maján, F.; Jiménez-Araujo, A.; Guillén-Bejarano, R.; Rodríguez-Arcos, R. Comparative Analysis of Chemical Compounds Related to Quality of Canned Asparagus. JFNR 2019, 7, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Megazyme Fructan Assay Kit-Measurement of Fructan in Plants Food|Megazyme. Available online: https://www.megazyme.com/fructan-assay-kit (accessed on 27 October 2021).
- Hamdi, A.; Jaramillo-Carmona, S.; Beji, R.; Tej, R.; Zaoui, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Kasri, M.; Lachaal, M.; Bouraoui, N.K.; et al. The Phytochemical and Bioactivity Profiles of Wild Asparagus Albus L. Plant. Food Res. Int. 2017, 99, 720–729. [Google Scholar] [CrossRef]
- Rodríguez, R.; Jaramillo, S.; Rodríguez, G.; Espejo, J.A.; Guillén, R.; Fernández-Bolaños, J.; Heredia, A.; Jiménez, A. Antioxidant Activity of Ethanolic Extracts from Several Asparagus Cultivars. J. Agric. Food Chem. 2005, 53, 5212–5217. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Alventosa, J.M.; Rodríguez-Gutiérrez, G.; Jaramillo-Carmona, S.; Espejo-Calvo, J.A.; Rodríguez-Arcos, R.; Fernandez-Bolaños, J.; Guillén-Bejarano, R.; Jiménez-Araujo, A. Effect of Extraction Method on Chemical Composition and Functional Characteristics of High Dietary Fibre Powders Obtained from Asparagus By-Products. Food Chem. 2009, 113, 665–671. [Google Scholar] [CrossRef]
- Hornero Méndez, D. Cambios en la Composición de Pigmentos en Frutos de Capsicum annuum Durante la Maduración y Procesado Para Pimentón: Variedades Bola y Agridulce. Doctoral Dissertation, Universidad de Sevilla, Sevilla, Spain, 1994. [Google Scholar]
- García-González, D.L.; Van De Voort, F.R. A Novel Wire Mesh “Cell” for Studying Lipid Oxidative Processes by Fourier Transform Infrared Spectroscopy. Appl. Spectrosc. 2009, 63, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.; Reiner, J.; Fleischhacker, L.; Viernstein, H.; Loeppert, R.; Praznik, W. Growth of Selected Probiotic Strains with Fructans from Different Sources Relating to Degree of Polymerization and Structure. J. Funct. Foods 2016, 24, 264–275. [Google Scholar] [CrossRef]
- Sulston, J.; Hodgkin, J. The Nematode Caenorhabditis Elegans; Wood, W., Ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1988. [Google Scholar]
- Viera-Alcaide, I.; Hamdi, A.; Rodríguez-Arcos, R.; Guillén-Bejarano, R.; Jiménez-Araujo, A. Asparagus Cultivation Co-Products: From Waste to Chance. FSN 2020, 6, 57. [Google Scholar] [CrossRef]
- Hamdi, A.; Jaramillo-Carmona, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Lachaal, M.; Karray-Bouraoui, N.; Guillén-Bejarano, R. Phytochemical Characterization and Bioactivity of Asparagus Acutifolius: A Focus on Antioxidant, Cytotoxic, Lipase Inhibitory and Antimicrobial Activities. Molecules 2021, 26, 3328. [Google Scholar] [CrossRef]
- Hamdi, A.; Jiménez-Araujo, A.; Rodríguez-Arcos, R.; Jaramillo-Carmona, S.; Lachaal, M.; Bouraoui, N.K.; Guillén-Bejarano, R. Asparagus Saponins; Chemical Characterization, Bioavailability and Intervention in Human Health. Nutr. Food Sci. Int. J. 2018, 7, 555704. [Google Scholar] [CrossRef]
- Jaramillo, S.; Muriana, F.J.G.; Guillén, R.; Jiménez-Araujo, A.; Rodríguez-Arcos, R.; López, S. Saponins from Edible Spears of Wild Asparagus Inhibit AKT, P70S6K, and ERK Signalling, and Induce Apoptosis through G0/G1 Cell Cycle Arrest in Human Colon Cancer HCT-116 Cells. J. Funct. Foods 2016, 26, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Verma, D.K.; Patel, A.R.; Thakur, M.; Singh, S.; Tripathy, S.; Srivastav, P.P.; Chávez-González, M.L.; Gupta, A.K.; Aguilar, C.N. A Review of the Composition and Toxicology of Fructans, and Their Applications in Foods and Health. J. Food Compos. Anal. 2021, 99, 103884. [Google Scholar] [CrossRef]
- Tewari, S.; Ramalakshmi, K.; Methre, L.; Mohan Rao, L.J. Microwave-Assisted Extraction of Inulin from Chicory Roots Using Response Surface Methodology. J. Nutr. Food Sci. 2015, 5, 342. [Google Scholar] [CrossRef] [Green Version]
- Monti, A.; Amaducci, M.T.; Pritoni, G.; Venturi, G. Growth, Fructan Yield, and Quality of Chicory (Cichorium Intybus L.) as Related to Photosynthetic Capacity, Harvest Time, and Water Regime. J. Exp. Bot. 2005, 56, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Lingyun, W.; Jianhua, W.; Xiaodong, Z.; Da, T.; Yalin, Y.; Chenggang, C.; Tianhua, F.; Fan, Z. Studies on the Extracting Technical Conditions of Inulin from Jerusalem Artichoke Tubers. J. Food Eng. 2007, 79, 1087–1093. [Google Scholar] [CrossRef]
- Puangbut, D.; Jogloy, S.; Vorasoot, N.; Srijaranai, S.; Holbrook, C.C.; Patanothai, A. Variation of Inulin Content, Inulin Yield and Water Use Efficiency for Inulin Yield in Jerusalem Artichoke Genotypes under Different Water Regimes. Agric. Water Manag. 2015, 152, 142–150. [Google Scholar] [CrossRef]
- Flores-Girón, E.; Salazar-Montoya, J.A.; Ramos-Ramírez, E.G. Application of a Box-Behnken Design for Optimizing the Extraction Process of Agave Fructans ( Agave Tequilana Weber Var. Azul): Box-Behnken Design for Optimizing the Extraction Process of Agave Fructans. J. Sci. Food Agric. 2016, 96, 3860–3866. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Martínez, J.; González-Cervantes, R.; Hernández-Gallegos, M.; Mendiola, R.; Aparicio, A.; Ocampo, M. Prebiotic Potential of Agave Angustifolia Haw Fructans with Different Degrees of Polymerization. Molecules 2014, 19, 12660–12675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeaiter, Z.; Regonesi, M.E.; Cavini, S.; Labra, M.; Sello, G.; Di Gennaro, P. Extraction and Characterization of Inulin-Type Fructans from Artichoke Wastes and Their Effect on the Growth of Intestinal Bacteria Associated with Health. BioMed Res. Int. 2019, 2019, 1083952. [Google Scholar] [CrossRef] [Green Version]
- Roldán-Marín, E.; Jensen, R.I.; Krath, B.N.; Kristensen, M.; Poulsen, M.; Cano, M.P.; Sánchez-Moreno, C.; Dragsted, L.O. An Onion Byproduct Affects Plasma Lipids in Healthy Rats. J. Agric. Food Chem. 2010, 58, 5308–5314. [Google Scholar] [CrossRef]
- Duarte, F.N.D.; Rodrigues, J.B.; da Costa Lima, M.; dos S. Lima, M.; Pacheco, M.T.B.; Pintado, M.M.E.; de Souza Aquino, J.; de Souza, E.L. Potential Prebiotic Properties of Cashew Apple (Anacardium Occidentale L.) Agro-Industrial Byproduct on Lactobacillus Species: Prebiotic Effects of Cashew Byproduct on Lactobacilli. J. Sci. Food Agric. 2017, 97, 3712–3719. [Google Scholar] [CrossRef]
- Li, W.; Zhang, J.; Yu, C.; Li, Q.; Dong, F.; Wang, G.; Gu, G.; Guo, Z. Extraction, Degree of Polymerization Determination and Prebiotic Effect Evaluation of Inulin from Jerusalem Artichoke. Carbohyd. Polym. 2015, 121, 315–319. [Google Scholar] [CrossRef]
- Singh, D.K.; Gulati, K.; Ray, A. Effects of Chelidonic Acid, a Secondary Plant Metabolite, on Mast Cell Degranulation and Adaptive Immunity in Rats. Int. Immunopharmacol. 2016, 40, 229–234. [Google Scholar] [CrossRef]
- Kim, D.-S.; Kim, S.-J.; Kim, M.-C.; Jeon, Y.-D.; Um, J.; Hong, S.-H. The Therapeutic Effect of Chelidonic Acid on Ulcerative Colitis. Biol. Pharm. Bull. 2012, 35, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zheng, Y.; Li, C.; Zheng, Y.; Wang, D.; Wu, Z.; Huang, L.; Wang, Y.; Li, P.; Peng, W.; et al. Discovery of Anti-Inflammatory Ingredients in Chinese Herbal Formula Kouyanqing Granule Based on Relevance Analysis between Chemical Characters and Biological Effects. Sci. Rep. 2015, 5, 18080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGaw, L.J.; Eloff, J.N. Ethnoveterinary Use of Southern African Plants and Scientific Evaluation of Their Medicinal Properties. J. Ethnopharmacol. 2008, 119, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Beneo Inulin from Orafti®|BENEO Prebiotic, Dietary Chicory Root Fibre. Available online: https://www.beneo.com/ingredients/human-nutrition/functional-fibres/inulin (accessed on 27 October 2021).
- Yoshida, M. Fructan Structure and Metabolism in Overwintering Plants. Plants 2021, 10, 933. [Google Scholar] [CrossRef]
- Shiomi, N. Structure of Fructopolysaccharide (Asparagosin) from Roots of Asparagus (Asparagus Officinalis L.). New Phytol. 1993, 123, 263–270. [Google Scholar] [CrossRef]
- Shiomi, N.; Yamada, J.; Izawa, M. Isolation and Identification of Fructo-Oligosaccharides in Roots of Asparagus (Asparagus Officinalis L.). Agric. Biol. Chem. 1976, 40, 567–575. [Google Scholar] [CrossRef]
- Witzel, K.; Matros, A. Fructans Are Differentially Distributed in Root Tissues of Asparagus. Cells 2020, 9, 1943. [Google Scholar] [CrossRef]
- Nobre, C.; Teixeira, J.A.; Rodrigues, L.R. New Trends and Technological Challenges in the Industrial Production and Purification of Fructo-Oligosaccharides. Crit. Rev. Food Sci. 2015, 55, 1444–1455. [Google Scholar] [CrossRef]
- Oligofructose the 100% Natural Sugar Replacer|BENEO. Available online: https://www.beneo.com/ingredients/human-nutrition/functional-fibres/oligofructose (accessed on 25 May 2022).
- Roberfroid, M.B.; Van Loo, J.A.E.; Gibson, G.R. The Bifidogenic Nature of Chicory Inulin and Its Hydrolysis Products. J. Nutr. 1998, 128, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Hong, T.; Iwashita, K.; Shiraki, K. Viscosity Control of Protein Solution by Small Solutes: A Review. CPPS 2018, 19, 746–758. [Google Scholar] [CrossRef] [PubMed]
- Minton, A.P. Influence of Macromolecular Crowding upon the Stability and State of Association of Proteins: Predictions and Observations. J. Pharm. Sci. 2005, 94, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Woods, C.E.; Litowski, J.R.; Roschen, L.A.; Gadgil, H.S.; Razinkov, V.I.; Kerwin, B.A. Effect of Sugar Molecules on the Viscosity of High Concentration Monoclonal Antibody Solutions. Pharm. Res. 2011, 28, 1552–1560. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Bayarri, S.; Tárrega, A.; Costell, E. Inulin as Texture Modifier in Dairy Products. Food Hydrocoll. 2011, 25, 1881–1890. [Google Scholar] [CrossRef]
- Yousaf, M.S.; Yusof, S.; Manap, M.Y.B.A.; Abd-Aziz, S. Storage Stabiliy of Clarified Banana Juice Fortified with Inulin and Oligofructose. J. Food Process. Pres. 2010, 34, 599–610. [Google Scholar] [CrossRef]
- Cardarelli, H.R.; Buriti, F.C.A.; Castro, I.A.; Saad, S.M.I. Inulin and Oligofructose Improve Sensory Quality and Increase the Probiotic Viable Count in Potentially Synbiotic Petit-Suisse Chesse. LWT-Food Sci. Technol. 2008, 41, 1037–1046. [Google Scholar] [CrossRef]
- Abou-Arab, A.A.; Talaat, H.A.; Abu-Salem, F.M. Physico-Chemical Properties of Inulin Produced from Jerusalem Artichoke Tubers on Bench and Pilot Plant Scale. Aust. J. Basic Appl. Sci. 2011, 5, 1297–1309. [Google Scholar]
- Barhatova, T.-; Nazarenko, M.-; Koguhova, M.-; Hripko, I. Obtaining and identification of inulin from jerusalem artichoke (Helianthus tuberosus) tubers. Foods Raw Mater. 2015, 3, 13–22. [Google Scholar] [CrossRef]
- Oliveira, R.N.; Mancini, M.C.; de Oliveira, F.C.S.; Passos, T.M.; Quilty, B.; Thiré, R.M.D.S.M.; McGuinness, G.B. FTIR Analysis and Quantification of Phenols and Flavonoids of Five Commercially Available Plants Extracts Used in Wound Healing. Matéria 2016, 21, 767–779. [Google Scholar] [CrossRef] [Green Version]
- Wongsa, P.; Phatikulrungsun, P.; Prathumthong, S. FT-IR Characteristics, Phenolic Profiles and Inhibitory Potential against Digestive Enzymes of 25 Herbal Infusions. Sci. Rep. 2022, 12, 6631. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, S.; Khor, B.-K.; Khaw, K.-Y.; Murugaiyah, V.; Chan, K.-L. Cholinesterase Inhibitory Potential of Dillenia Suffruticosa Chemical Constituents and Protective Effect against Aβ−induced Toxicity in Transgenic Caenorhabditis Elegans Model. Phytomed. Plus 2021, 1, 100022. [Google Scholar] [CrossRef]
- Yang, T.; Fang, L.; Lin, T.; Li, J.; Zhang, Y.; Zhou, A.; Xie, J. Ultrasonicated Sour Jujube Seed Flavonoids Extract Exerts Ameliorative Antioxidant Capacity and Reduces Aβ-Induced Toxicity in Caenorhabditis Elegans. J. Ethnopharmacol. 2019, 239, 111886. [Google Scholar] [CrossRef] [PubMed]
- Yuen, C.W.; Murugaiyah, V.; Najimudin, N.; Azzam, G. Danshen (Salvia Miltiorrhiza) Water Extract Shows Potential Neuroprotective Effects in Caenorhabditis Elegans. J. Ethnopharmacol. 2021, 266, 113418. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Li, Q.; Yang, T.; Qi, W.; Zhang, Y.; Wang, L.; Sun, X.; Xie, J. Flavonoid Glucosides from Ziziphus Jujuba Seeds Improve Learning and Memory in Mice. Rev. Bras. Farmacogn. 2022, 32, 99–110. [Google Scholar] [CrossRef]
- Kang, N.; Luan, Y.; Jiang, Y.; Cheng, W.; Liu, Y.; Su, Z.; Liu, Y.; Tan, P. Neuroprotective Effects of Oligosaccharides in Rehmanniae Radix on Transgenic Caenorhabditis Elegans Models for Alzheimer’s Disease. Front. Pharmacol. 2022, 13, 878631. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, R.; Li, H.; Xiang, Y.; Xiao, L.; Hu, M.; Ma, F.; Ma, C.W.; Huang, Z. Antioxidant and Neuroprotective Effects of Dictyophora Indusiata Polysaccharide in Caenorhabditis Elegans. J. Ethnopharmacol. 2016, 192, 413–422. [Google Scholar] [CrossRef]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical Nodes in Signalling Pathways: Insights into Insulin Action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef]
- Lindhurst, M.J.; Parker, V.E.R.; Payne, F.; Sapp, J.C.; Rudge, S.; Harris, J.; Witkowski, A.M.; Zhang, Q.; Groeneveld, M.P.; Scott, C.E.; et al. Mosaic Overgrowth with Fibroadipose Hyperplasia Is Caused by Somatic Activating Mutations in PIK3CA. Nat. Genet. 2012, 44, 928–933. [Google Scholar] [CrossRef]
- Iravani, S. Green Synthesis of Metal Nanoparticles Using Plants. Green Chem. 2011, 13, 2638. [Google Scholar] [CrossRef]
- Mohammadi Arvanag, F.; Bayrami, A.; Habibi-Yangjeh, A.; Rahim Pouran, S. A Comprehensive Study on Antidiabetic and Antibacterial Activities of ZnO Nanoparticles Biosynthesized Using Silybum Marianum L Seed Extract. Mater. Sci. Eng. C 2019, 97, 397–405. [Google Scholar] [CrossRef]
- Hussein, J.; Attia, M.F.; El Bana, M.; El-Daly, S.M.; Mohamed, N.; El-Khayat, Z.; El-Naggar, M.E. Solid State Synthesis of Docosahexaenoic Acid-Loaded Zinc Oxide Nanoparticles as a Potential Antidiabetic Agent in Rats. Int. J. Biol. Macromol. 2019, 140, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, S.; Ruvkun, G. Daf-2, Daf-16 and Daf-23: Genetically Interacting Genes Controlling Dauer Formation in Caenorhabditis Elegans. Genetics 1994, 137, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Gems, D.; Sutton, A.J.; Sundermeyer, M.L.; Albert, P.S.; King, K.V.; Edgley, M.L.; Larsen, P.L.; Riddle, D.L. Two Pleiotropic Classes of Daf-2 Mutation Affect Larval Arrest, Adult Behavior, Reproduction and Longevity in Caenorhabditis Elegans. Genetics 1998, 150, 129–155. [Google Scholar] [CrossRef] [PubMed]
- Oghbaei, H.; Rastgar Rezaei, Y.; Nikanfar, S.; Zarezadeh, R.; Sadegi, M.; Latifi, Z.; Nouri, M.; Fattahi, A.; Ahmadi, Y.; Bleisinger, N. Effects of Bacteria on Male Fertility: Spermatogenesis and Sperm Function. Life Sci. 2020, 256, 117891. [Google Scholar] [CrossRef] [PubMed]
- Tissenbaum, H.A.; Ruvkun, G. An Insulin-like Signaling Pathway Affects Both Longevity and Reproduction in Caenorhabditis Elegans. Genetics 1998, 148, 703–717. [Google Scholar] [CrossRef]
- Lakra, A.K.; Ramatchandirane, M.; Kumar, S.; Suchiang, K.; Arul, V. Physico-Chemical Characterization and Aging Effects of Fructan Exopolysaccharide Produced by Weissella Cibaria MD2 on Caenorhabditis Elegans. LWT 2021, 143, 111100. [Google Scholar] [CrossRef]
- Rasulova, M.; Zečić, A.; Monje Moreno, J.M.; Vandemeulebroucke, L.; Dhondt, I.; Braeckman, B.P. Elevated Trehalose Levels in C. elegans Daf-2 Mutants Increase Stress Resistance, Not Lifespan. Metabolites 2021, 11, 105. [Google Scholar] [CrossRef]
- Chandrashekar, V.; Bartke, A. The Impact of Altered Insulin-like Growth Factor-I Secretion on the Neuroendocrine and Testicular Functions. Minerva. Ginecol. 2005, 57, 87–97. [Google Scholar]
- Hamidi, M.; Ziaee, M.; Delashoub, M.; Marjani, M.; Karimitabar, F.; Khorami, A.; Ahmadi, N.A. The Effects of Essential Oil of Lavandula Angustifolia on Sperm Parameters Quality and Reproductive Hormones in Rats Exposed to Cadmium. J. Rep. Pharma. Sci. 2015, 4, 135–142. [Google Scholar]
- Keyhanmanesh, R.; Hamidian, G.; Alipour, M.R.; Ranjbar, M.; Oghbaei, H. Protective Effects of Sodium Nitrate against Testicular Apoptosis and Spermatogenesis Impairments in Streptozotocin-Induced Diabetic Male Rats. Life Sci. 2018, 211, 63–73. [Google Scholar] [CrossRef]
- Nef, S.; Verma-Kurvari, S.; Merenmies, J.; Vassalli, J.-D.; Efstratiadis, A.; Accili, D.; Parada, L.F. Testis Determination Requires Insulin Receptor Family Function in Mice. Nature 2003, 426, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Kushwaha, S.; Jena, G.B. Relative Influence of Testosterone and Insulin in the Regulation of Prostatic Cell Proliferation and Growth. Steroids 2011, 76, 416–423. [Google Scholar] [CrossRef]
- Brüning, J.C.; Gautam, D.; Burks, D.J.; Gillette, J.; Schubert, M.; Orban, P.C.; Klein, R.; Krone, W.; Müller-Wieland, D.; Kahn, C.R. Role of Brain Insulin Receptor in Control of Body Weight and Reproduction. Science 2000, 289, 2122–2125. [Google Scholar] [CrossRef] [PubMed]
- Pitetti, J.-L.; Calvel, P.; Zimmermann, C.; Conne, B.; Papaioannou, M.D.; Aubry, F.; Cederroth, C.R.; Urner, F.; Fumel, B.; Crausaz, M.; et al. An Essential Role for Insulin and IGF1 Receptors in Regulating Sertoli Cell Proliferation, Testis Size, and FSH Action in Mice. Mol. Endocrinol. 2013, 27, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Hammoud, A.; Meikle, A.; Reis, L.; Gibson, M.; Peterson, C.; Carrell, D. Obesity and Male Infertility: A Practical Approach. Semin. Reprod. Med. 2012, 30, 486–495. [Google Scholar] [CrossRef]
- Khorrami, A.; Ghanbarzadeh, S.; Ziaee, M.; Arami, S.; Vajdi, R.; Garjani, A. Dietary Cholesterol and Oxidised Cholesterol: Effects on Sperm Characteristics, Antioxidant Status and Hormonal Profile in Rats. Andrologia 2015, 47, 310–317. [Google Scholar] [CrossRef]
- Liu, F.; Prabhakar, M.; Ju, J.; Long, H.; Zhou, H.-W. Effect of Inulin-Type Fructans on Blood Lipid Profile and Glucose Level: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Eur. J. Clin. Nutr. 2017, 71, 9–20. [Google Scholar] [CrossRef]
- Guess, N.D.; Dornhorst, A.; Oliver, N.; Frost, G.S. A Randomised Crossover Trial: The Effect of Inulin on Glucose Homeostasis in Subtypes of Prediabetes. Ann. Nutr. Metab. 2016, 68, 26–34. [Google Scholar] [CrossRef]
Figure 1.
Percentual composition of simple sugars in the different fructan samples.

Figure 2.
HPAEC profiles of the different fructan samples: asparagus fructan, Orafti®GR, chicory inulin from SaludViva, and agave inulin from Tentorium Energy S.L.
Figure 2.
HPAEC profiles of the different fructan samples: asparagus fructan, Orafti®GR, chicory inulin from SaludViva, and agave inulin from Tentorium Energy S.L.

Figure 3.
Viscosity patterns of the different fructan samples at different concentrations (5–30% w/v) and temperatures (80 °C, 20 °C, and 4 °C).
Figure 3.
Viscosity patterns of the different fructan samples at different concentrations (5–30% w/v) and temperatures (80 °C, 20 °C, and 4 °C).

Figure 4.
FT-IR spectrum of the different fructan samples.

Figure 5.
Area under the growth curve for the different inulins studied and several Lactobacillus strains. The bar for each value indicates the standard deviation. All analyses were performed at least in triplicate. Values bearing the same letter are not significantly different at the 5% level, as determined by the Duncan multiple range test.
Figure 5.
Area under the growth curve for the different inulins studied and several Lactobacillus strains. The bar for each value indicates the standard deviation. All analyses were performed at least in triplicate. Values bearing the same letter are not significantly different at the 5% level, as determined by the Duncan multiple range test.

Figure 6.
Percentage of paralyzed worms at a non-permissive temperature (25 °C). Fructans extracts (168 µg/mL) do not ameliorate the paralysis phenotype of GMC101 nematode model of Alzheimer’s disease. The percentage of paralyzed worms at a non-permissive temperature (25 °C) in non-treated (blue bar) and treated (brown bar) GMC101 strain was presented. Results are expressed as mean ± SD. ns represents no significant effect of fructans compared to the control using two-tailed t-test. Final data represented the average of three independent assays.
Figure 6.
Percentage of paralyzed worms at a non-permissive temperature (25 °C). Fructans extracts (168 µg/mL) do not ameliorate the paralysis phenotype of GMC101 nematode model of Alzheimer’s disease. The percentage of paralyzed worms at a non-permissive temperature (25 °C) in non-treated (blue bar) and treated (brown bar) GMC101 strain was presented. Results are expressed as mean ± SD. ns represents no significant effect of fructans compared to the control using two-tailed t-test. Final data represented the average of three independent assays.

Figure 7.
Progeny of daf-2(e1370) strain in presence or absence (control) of asparagus fructans. Asparagus fructan extracts (168µg/mL) enhance fertility of the mutant daf-2(e-1370). In the X-axis, the blue bar corresponding to daf-2(e-1370) without fructan extract treatment and the brown bar corresponding to daf-2(e-1370) treated with 168 µg/mL of fructan extract. The Y-axis indicates the average total number of progenies per worm produced during 72 h. The total number of progeny produced increased significantly following asparagus fructans extracts treatment. Graphs represent mean ± SD n = 30. **** represents p < 0.001 using two-tailed t-test. Two more replicates with similar results were carried out to confirm these effects (Supplementary Material).
Figure 7.
Progeny of daf-2(e1370) strain in presence or absence (control) of asparagus fructans. Asparagus fructan extracts (168µg/mL) enhance fertility of the mutant daf-2(e-1370). In the X-axis, the blue bar corresponding to daf-2(e-1370) without fructan extract treatment and the brown bar corresponding to daf-2(e-1370) treated with 168 µg/mL of fructan extract. The Y-axis indicates the average total number of progenies per worm produced during 72 h. The total number of progeny produced increased significantly following asparagus fructans extracts treatment. Graphs represent mean ± SD n = 30. **** represents p < 0.001 using two-tailed t-test. Two more replicates with similar results were carried out to confirm these effects (Supplementary Material).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Chemical composition of inulin from different sources.
| Moisture | Simple Sugars | Ash | Protein | Fructans | |
|---|---|---|---|---|---|
| Asparagus | 9.00 ± 0.13 c | 4.19 ± 0.43 b | 6.69 ± 0.06 | 17.50 ± 0.83 | 57.94 ± 0.41 a |
| Orafti-GR | 4.61 ± 0.37 a | 1.99 ± 0.01 a | t | n.d. | 78.11 ± 0.51 b |
| Chicory | 6.00 ± 0.57 b | 8.48 ± 0.59 d | t | n.d. | 58.96 ± 1.44 a |
| Agave | 4.60 ± 0.11 a | 7.05 ± 0.39 c | t | n.d. | 80.65 ± 2.47 b |
All analyses were performed at least in triplicate. The results are presented as mean ± standard deviation. Means bearing the same letter are not significantly different at the 5% level, as determined by the Duncan multiple range test. n.d.: not detected. t: traces.
Table 2.
Phenolic composition and antioxidant activity of asparagus fructan extract. TE: Trolox equivalents.
Table 2.
Phenolic composition and antioxidant activity of asparagus fructan extract. TE: Trolox equivalents.
| mg/g Asparagus Fructan | |
| Chelidonic acid | 1.13 ± 0.07 |
| Caffeic acid glycoside | 0.30 ± 0.00 |
| Caffeic acid | 0.64 ± 0.03 |
| p-Coumaric acid | 0.07 ± 0.00 |
| t-Ferulic acid | 0.11 ± 0.01 |
| Total | 2.25 ± 0.03 |
| μmols TE/g asparagus fructan | |
| Antioxidant activity | 43.62 ± 3.16 |
All analyses were performed at least in triplicate. The results are presented as mean ± standard deviation.
Table 3.
pH, oil holding capacity (OHC) and color of the different fructan samples.
| Color | |||||
|---|---|---|---|---|---|
| pH | OHC | L* | a* | b* | |
| Asparagus | 6.45 ± 0.01 d | 93.63 ± 1.41 b | 50.5203 | 3.9032 | 17.3581 |
| Orafti®GR | 6.04 ± 0.02 c | 78.46 ± 3.01 a | 88.8906 | −0.3873 | 2.6834 |
| Chicory | 5.12 ± 0.03 b | 95.15 ± 4.76 b | 86.9714 | −0.1935 | 3.039 |
| Agave | 4.59 ± 0.23 a | 79.93 ± 2.78 a | 88.1635 | −0.6182 | 4.652 |
All analyses were performed at least in triplicate. The results are presented as mean ± standard deviation. Means bearing the same letter are not significantly different at the 5% level, as determined by the Duncan multiple range test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hamdi, A.; Viera-Alcaide, I.; Guillén-Bejarano, R.; Rodríguez-Arcos, R.; Muñoz, M.J.; Monje Moreno, J.M.; Jiménez-Araujo, A. Asparagus Fructans as Emerging Prebiotics. Foods 2023, 12, 81. https://doi.org/10.3390/foods12010081
AMA Style
Hamdi A, Viera-Alcaide I, Guillén-Bejarano R, Rodríguez-Arcos R, Muñoz MJ, Monje Moreno JM, Jiménez-Araujo A. Asparagus Fructans as Emerging Prebiotics. Foods. 2023; 12(1):81. https://doi.org/10.3390/foods12010081
Chicago/Turabian StyleHamdi, Amel, Isabel Viera-Alcaide, Rafael Guillén-Bejarano, Rocío Rodríguez-Arcos, Manuel Jesús Muñoz, Jose Manuel Monje Moreno, and Ana Jiménez-Araujo. 2023. "Asparagus Fructans as Emerging Prebiotics" Foods 12, no. 1: 81. https://doi.org/10.3390/foods12010081
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.