3.2. Physico-Chemical Characterization
In
Table 2 the chemical–physical parameters of the fermented sausages at the end of ripening are shown. Wide differences in the final pH were observed, with values ranging from a minimum of 4.52 in the ECO samples to a maximum of 6.42 in IM1.
In general, Italian products have the highest pH values (between 5.88 and 6.42), together with Croatian ones (between 5.72 and 6.05). On the other hand, Slovenian sausages showed lower pH values (5.20 and 5.39). These values agree with those reported by Lešić et al. [
20] for Croatian and Slovenian sausages and with the data reported by Cardinali et al. [
21] for Italian Fabriano sausages.
Slovenian products are probably more subjected to northern European production influences. In fact, northern products are generally dried sausages with a pH around or even below 5 and often undergo a smoking phase (that inhibits molds), while Mediterranean sausages are usually long-ripened and with pH values up to 6.2–6.4, given the possible growth of desirable molds [
22,
23].
The Spanish fermented products were widely different in terms of pH, due to the heterogeneous traditional manufacturing process developed in the whole country. In fact, chorizo samples showed a very low pH (between 4.52 and 5.04), while the salchichón had a pH ranging from 5.13 to 5.83. There are different set of data reported in the literature for chorizo characteristics, and among them the pH can vary: results of Galician chorizo showed a pH value of about 5.6 [
24], while Asturian products had pH between 5.0 and 5.1 [
25].
Great variability was also observed in the final aw of the products. In some case, this parameter reached extremely low values, such as in IM2 (0.760), ESB (0.811), IM1 (0.824) and SN (0.823). On the other hand, HNS showed the highest value (0.928). In any case, the aw values reflected the water content of the final products. This parameter ranged from 20.42% in IM2 to 39.04% in ESO. The salt content in the final products ranged from 2.94 (HNS) to 4.48 (IM2).
3.3. Microbiological Analysis
Microbial counts were performed to enumerate in the final product LAB, CNC, enterococci, enterobacteria (including
E. coli) and yeasts (
Table 3).
The largest microbial population was generally represented by LAB, whose counts ranged from 4.4 (ECB) to 8.7 (HNS) log cfu/g. The highest LAB counts were found in samples collected from Croatia and Italy. The lowest numbers were associated with sausages from Spain, particularly ECB and ECE. LAB have been described as the main bacterial population in the dominant microbiota of Croatian traditionally fermented sausages and in Italian spontaneously fermented salami, with loads of 7–8 log cfu/g in the final products [
26,
27,
28].
The counts of CNC were higher in Italian Fabriano salami (7.1 log cfu/g) and in HZK (7.2 log cfu/g). This microbial group was below the detection limit in SN, ECB, ECE and ECO, while in SWO, its concentration was very low (1.4 log cfu/g). The presence of CNC was strongly related to pH, and in particular, they seem to be inhibited by lower pH.
Enterococci were not detected or were detected in sporadic quantities (<1 log cfu/g) in Italian and Slovenian samples and in three Spanish samples (ESA, ECB, ECO). The highest counts of this microbial group were found in Croatian samples (3.2–4.8 log cfu/g).
Yeasts ranged from 2.3 to 5.4 log cfu/g, except for SN sausage. This microbial population can have an important role in sausage-ripening, contributing to the formation of the aroma and to the evolution of product sensory features [
29,
30].
Enterobacteriaceae were present in detectable amounts only in Croatian products, with values ranging between 2.7 (HS) and 5.1 (HZK). In all other samples, this population was below the limit of determination. High levels of
Enterobacteriaceae can indicate the low microbiological quality of the product, and high concentrations (>4 log cfu/g) of this population have been enumerated in several ripened traditional products [
31,
32,
33].
The counts of E. coli were always below the detection limit. In addition, a search for pathogenic microorganisms such as Listeria monocytogenes and Salmonella was carried out, and no positive samples were found.
3.4. Metagenomic Analysis
A more detailed picture of the bacterial composition of spontaneously fermented sausage was obtained through amplicon sequencing and metagenomic analysis.
Only species and genera that reached a concentration higher than 0.5% of amplicon sequence variants (ASVs) in at least one of the samples were considered. A total of more than 500 ASVs were detected, indicating very high biodiversity in the composition of the microbiota of the European salamis taken into consideration. The relative abundance of the ASVs attributed to genera or species is given in
Table 4.
Beyond a large number of microbial species, their relative composition was also very variable among the products, even for those collected in the same geographical area.
While in some samples most of the ASVs were attributed to a single group/species, some sausages were characterized by higher biodiversity, with an important diversification in the microbiota composition (e.g., HZK and ESO).
Among LAB, several genus and species were present. Members of the genus
Latilactobacillus were found in all the sausages.
Latilactobacillus sakei was the dominant species (>50% of ASVs in IM2, IAL, and SN). The
Latilactobacillus sakei group (which included
Lat. curvatus,
Lat. graminis and
Lat. fuchuensis) was found in all the sausages, even if in lower percentage, except for IM1 and ESA.
Lat. sakei, and to a lesser extent
Lat. curvatus, have been reported as the prevailing LAB species in fermented meat products originating from France, Italy and Spain [
6,
34,
35]. In fact, LAB species diversity of fermented sausages is known to be limited, being
Lat. sakei-predominant during the ripening process, due to the species’ excellent adaptation, competitiveness and assertiveness in the meat matrix [
36,
37,
38,
39,
40,
41]. This predominance over other LAB can be attributed to its salt-tolerant and psychrotrophic nature and its specialization in metabolic pathways in the meat environment, including the arginine deiminase pathway and the utilization of nucleosides [
36,
42,
43,
44,
45,
46].
Other lactobacilli were sporadically detected in low amounts, such as
Lacticaseibacillus casei group in ESB,
Lactiplantibacillus plantarum group in ESO,
Ligilactobacillus sp. and
Loigolactobacillus rennini in HS and
Dellaglioa algida in SWO. High levels of the member of the genus
Companilactobacillus (
Com. alimentarius,
Com. heilongjiangensis and
Com. versmoldensis) were present in many Spanish sausages, in particular ESE and ECB, in which they represented 55.3 and 45.0% of the total ASVs. The abundance of
Companilactobacillus found only in Spanish sausages, is a regional peculiarity already reported in the literature [
47,
48]. Moreover, these species have been isolated as spoilage organisms in ready-to-eat meats and from other dry fermented sausages originating from Belgium and Italy [
6,
49,
50] and have been proposed as autochthonous probiotic starter cultures for meat products [
51].
Heterofermentative lactobacilli were present only in ESO and ECE (Levilactobacillus yonginensis group and Limosilactobacillus mucosae, respectively). In addition, ASVs belonging to some dairy LAB (Lactobacillus helveticus, Streptococcus thermophilus and Lactococcus sp.) were found in some Spanish products. These species can be related to the use of powdered milk or other dairy derivatives during manufacturing. The heterofermentative cocci were represented by Leuconostoc carnosum, present in relevant percentages in IM1 (15,8%), SN (19.1%), SWO (23.3%) and HZK (14.8%); Leuconostoc sp. (17.6% in IAL and 15.6% in SWO); and Weissella sp., found only in ESO.
Five species of staphylococci were detected (Staph. epidermidis, Staph. equorum, Staph. saprophyticus, Staph. succinus and Staph. xylosus). They were found only in some sausages, and they were dominant in IM1 (67.7% of ASVs belonging to Staph. equorum and 1.5% to Staph. succinus) and ESA (98.1 of ASVs belonging to Staph. xylosus).
Among CNS,
Staphylococcus xylosus is the prevalent species associated with salamis, even if greater species diversity can be found,
Staphylococcus epidermidis,
Staphylococcus equorum and
Staphylococcus saprophyticus also being reported in these products [
52,
53,
54].
Staphylococci exert several important technological roles in sausage production, such as contribution to flavor and color [
54,
55], and since they are able to use alternative energy sources, such as arginine and nucleoside, their ecological persistence is assured [
37]. These bacteria were detected as subdominant fractions in several fermented meat products, and their presence was associated with mammal skin, being competitive in the acidic and anaerobic conditions prevailing during meat fermentation [
56,
57]. It is noteworthy that many samples showed a very low pH (i.e., Spanish products) that could have hindered their growth. The results therefore underline the sensitivity of this genus to low pH, even in a potentially favorable environment, thanks to the low aw values. These populations probably suffered the strong initial acidification in these products, which was not followed by a significant increase in pH (scarcity of molds, etc.).
Carnobacterium sp. was highlighted only in some Croatian samples, i.e., HS and HNS, albeit with a relevant percentage (19.11% and 9%, respectively). This genus has been found in Fabriano-like products [
21], and it has been associated with meat spoilage phenomena [
58].
Among Actinobacteria, Kocuria sp. and Corynebacterium sp. were present only in HZK, while Brevibacterium casei and Rothia sp. were present in ECE.
The meat spoiler
Brochothrix thermosphacta was detected in several sausages, and its concentration was particularly relevant in HZK (19.58%), IM2 (14.91%) and ESE (14.23%). This species has been reported to be part of the Fabriano-like sausage microbiota by Cardinali et al. [
21], and it is commonly associated with spoiled meat-based products [
59,
60].
Among Gram-negative bacteria, Pseudomonas sp. appeared at a high concentration in the Slovenian samples, in the Spanish ESE (16.7% of ASV) and especially in HNS, in which this genus represented 26.8% of ASVs. Enterobacteria (Klebsiella sp. and Escherichia/Shigella group) were found in ESB, ESO and ECE.
To better evidence the differences in the ASV composition, a cluster analysis was carried out. The results are reported in
Figure 1.
Four different clusters were obtained. The first contained only the Italian sausage IM1. In the second cluster were grouped three Spanish sausages (ESE, ECE and ESO) and the Croatian sample HZK. The third cluster was represented only by ESA. The fourth cluster included nine samples, and in particular the Spanish sausages ECB, ECO and ESB; the Croatian HNS and HS; the Slovenian sausages; and the Italian samples IM2 and IAL.
To highlight the microorganisms responsible for the clustering of sausages, a PCA was applied whose first two components explained 59.76% of the variability (
Figure 2).
The grouping of the sausages reflected the results of the tree diagram. In
Figure 2, factor coordinates of the metagenomic variables are shown. Only the metagenomic variables with coordinates characterized by an absolute value higher than 1 are reported. The
Lat. sakei and the
Lat. sakei group were the main ones responsible for the description of the largest group located in the first quadrant, while
Staph. xylosus determined the collocation of ESA in the left part of the second quadrant. The position of IM1 depends on the variable
Staph. equorum, while the remaining group collocation is mainly influenced by the presence of
Companilactobacillus (especially for the Spanish samples),
Corynebacterium for the Croatian sausage and
Brochothrix thermosphacta for all the samples grouped in this cluster.
3.5. Biogenic Amine Concentrations
The biogenic amine concentrations in the sausages are reported in
Table 5.
Tyramine was present in all the samples, with concentrations ranging from 47.7 mg/kg (IM2) to 366.8 mg/kg (HNS). The mean content of this biogenic amine was 165.5 mg/kg, with a standard deviation of 88.8 mg/kg, indicating a fair variability among the fifteen dry fermented sausages. In general, the Italian products showed lower tyramine concentrations, while the highest amounts were found in the Croatian salamis HNS and HS. These amounts are similar to those reported by EFSA [
61], which indicated tyramine mean concentration of 136 mg/kg, with the 95th percentile of 397 mg/kg, in 400 European salami samples.
Histamine was found only in two Spanish samples (ESB and ECE) at concentrations of 195.8 and 174.7 mg/kg, respectively. This amine is considered the most dangerous for human health as it can cause various adverse effects known as “histamine poisoning” [
62]. The quantities found in the two Spanish samples, although not very high, are nevertheless significant, especially when compared with the maximum amounts allowed in some fish products. These latter are the only ones regulated for histamine presence and generally admit a maximum quantity of 100 mg/kg in fresh fish and of 200 mg/kg for processed products [
63].
The presence of putrescine was more variable: in four samples, this amine was not detected, while two Croatian samples (HNS and HS) showed the highest amounts (about 300 mg/kg). In the same samples, higher quantities of cadaverine were also observed.
Both Gram-negative and Gram-positive bacteria have been described as biogenic amine producers, with wide variability in aminobiogenetic potential between different strains of the same species. Spoilage microorganisms such as enterobacteria and pseudomonads can be strong histamine, cadaverine and putrescine accumulators, and biogenic amines produced by these microbial populations can also be found in fermented sausages [
64,
65]. On the other hand, decarboxylase activity has been found in Gram-positive strains, also belonging to the genus
Staphylococcus and LAB. While
Lat. sakei is usually known for its inability to produce BAs, in many other LAB species, strains with decarboxylase activity are present.
Lat. curvatus, for example, can accumulate both tyramine and histamine as well as
Com. alimentarius [
66,
67,
68]. The ability to produce BAs has been found in other genera found in this investigation, such as
Leuconostoc sp.,
Weissella sp. and
Carnobacterium [
68].
3.6. Sausage Aroma Profile
The aroma profile of the fifteen ripened sausages has been studied. The unidentified compounds accounted for less than 1% of the total peak area in each sample.
The analysis of the volatile organic compounds (VOCs) allowed the clear differentiation of the samples, reflecting the different formulations and production and ripening conditions traditionally adopted in the Mediterranean geographical areas. Indeed, it has been reported that these differences can influence sausage aroma, being dry fermented sausage flavor affected by many processing factors, i.e., raw materials, spices, microbiota composition, smoking, etc. [
55].
Within this wide variability, some common characteristics can be found by grouping the identified molecules in homogeneous chemical groups: ketones, aldehydes, alcohols, acids, and esters (
Table 6), as well as molecules deriving from spices and smoking, reported as
Supplementary Materials (
Tables S1 and S2, respectively).
Concerning the molecules associated with the spices included in the meat batter formulation (
Table S1), they belong to terpenes and terpenoids, phenylpropenes and compounds deriving from garlic (dimethyl disulfide, diallyl sulfide, etc.) [
69]. These latter compounds are particularly present in Slovenian salami, Spanish chorizo and Croatian products, except for the HNS sample. Among terpenes, D-limonene certainly is the prevalent molecule in products in which pepper has been used. Many other terpenes deriving from this spice, such as myrcene, linalool, copaene, carene, o-cymene, etc. [
70], have been detected in these products, while they are absent in chorizo sausages, where oregano has been used. On the other hand, eugenol, safrole and methyl-eugenol are associated with the use of spices such as nutmeg, cinnamon and cloves [
71,
72].
The VOCs derived from smoke (
Table S2) include furans and phenols, already reported for smoked products [
73], and their presence is higher in the Slovenian sausages (SN and SWO) and HS, characterized by a smoking phase during their production. Furthermore, some of these VOCs were detected also in the Spanish chorizo samples, due to the traditional use of smoked paprika [
74].
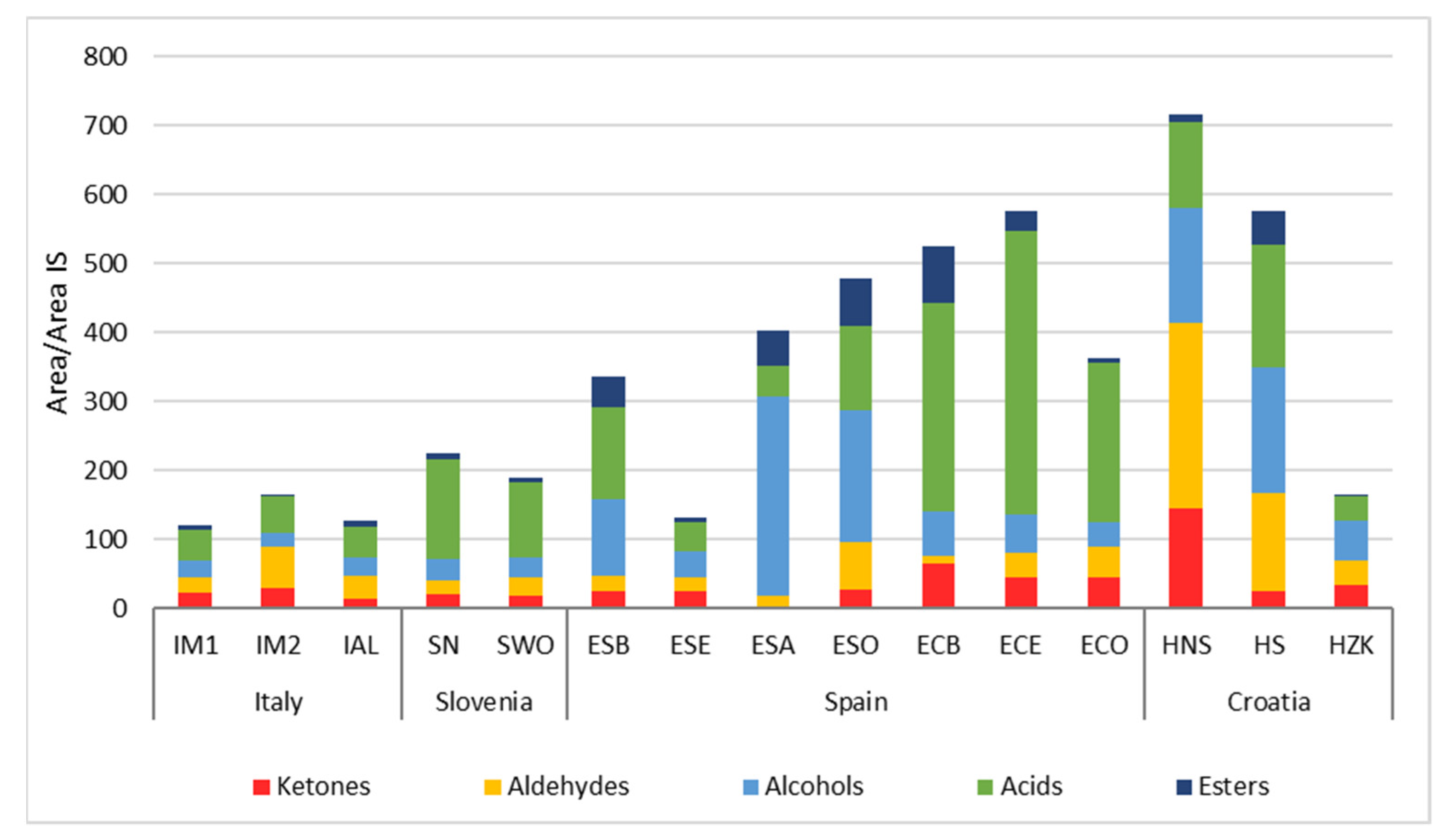
Table 6 reports VOCs derived from the microbial biochemical activities that occurred during sausage fermentation and ripening. The compounds are grouped into chemical families, whose total amounts are shown in
Figure 3. Higher quantities of VOCs were found in some Spanish products and two Croatian samples.
Ketones were evenly distributed in the analysed samples, except for chorizo and HNS samples where their total amount was higher. On the other hand, lower values were found in salchichón ESA. These molecules mostly derive from fatty acid oxidation and, in particular, from β-oxidation. Some microbial groups such as staphylococci and fungi can have a role in these phenomena. It is interesting to observe that products characterized by high concentrations of fat (for example HNS) showed the higher presence of ketones. HS, having the same formulation of HNS but subjected to smoking, presented lower ketone amounts: this can be attributed to the antioxidant role of some of the compounds produced by smoking. It is also interesting to note that chorizo samples are characterized by higher ketone amounts among Spanish products. These fermented sausages did not contain nitrates or nitrites, which exert a well-known antioxidant activity, while these preservatives were present in salchichón formulations.
Among ketones, 2-butanone prevailed in some samples, i.e., ECB and HNS (
Table 6). The presence of this molecule in fermented meat products is common but its contribution to aroma profile can be negative depending on its amounts and the balance with other VOCs [
75]. Diacetyl (2,3-butanedione) and acetoin (3-hydroxy-2-butanone) are particularly present in some Spanish products (ECB and ECO), in Slovenian samples and in two Croatian sausages (HNS and HZK). Diacetyl and acetoin are mainly produced through the catabolism of pyruvic acid by LAB [
10,
76].
Aldehydes are particularly relevant in the Italian sausage IM2 and a Spanish salchichón (ESO), but the maximum amounts were found in Croatian HNS and HS. Most of the detected aldehydes can derive from fatty acids oxidation. Within certain limits, these compounds can contribute to the typical sausage aroma profile. Nevertheless, given their strong aroma perception characterized by herbaceous notes, excessive quantities can lead to organoleptic defects (i.e., rancidity) [
10]. Hexanal, together with nonanal, is certainly the most characteristic molecule of this VOC group. On the other hand, the methyl-branched aldehydes, such as isovaleric aldehyde (3-methyl-butanal), have often been associated with fermented sausage aroma and can derive from the bacterial metabolism of branched amino acids, in particular leucine [
44,
77]. Benzaldehyde and benzeneacetaldehyde are present in most of the samples. In particular, HS contained higher amounts of benzeneacetaldehyde, while benzaldehyde is predominant in HNS and ECO. These VOCs are the result of aromatic amino acid (phenylalanine and tyrosine) metabolism and can contribute to the sausage flavor imparting floral and almond notes [
78].
Alcohols are present in all the samples, but their amounts were higher in HNS and HS and some Spanish samples, particularly in ESA and ESO samples (
Table 6). Among alcohols, ethanol was the most abundant in the samples, with high amounts in some Spanish salchichón (ESB, ESA, ESO) and in Croatian sausages, while a lower presence was highlighted in Italian and Slovenian salamis. The presence of this compound can be influenced by the wine addition in meat batter formulation or it can be the result of several microbial pathways, i.e., pyruvate or amino acid metabolisms [
77,
79], being therefore strongly influenced by the natural microbiota composition of each product. Phenethyl alcohol, which is the results of benzenacetaldehyde reduction and that can give a rose odor, was present in significant amounts in HS and ESA, samples characterized by high amounts of its precursor.
Acids prevailed in chorizo samples but they were the most represented molecular class in Slovenian sausages (
Figure 3). Acetic acid was found in low amounts in Italian salamis and in ESA, ESE and HZK, which showed high pH values. In contrast, significantly higher amounts of acetic acid were detected in the chorizo samples, particularly in ECE. These samples had the lowest pH, ranging from 4.52 to 5.04. Acetic acid can be produced, similarly to ethanol, with many bacterial metabolic pathways and different microbial groups able to be responsible for its accumulation, including LAB, staphylococci and fungi.
It is interesting to underline the presence of isovaleric acid (3-methyl, butanoic acid) (
Table 6), whose occurrence in fermented sausages is well-documented and which can exert a very strong organoleptic impact even in low quantities [
80,
81]. Higher amounts of this VOC have been detected in IM1, while it was not present in very small quantities in HS and HNS.
Esters were present in lower amounts in comparison to other VOC classes and were mainly represented by ethyl acetate, ethyl hexanoate and ethyl octanoate (
Table 6). Their levels showed dependence on sausage type. They were more abundant in some Spanish samples and in HS (
Figure 3), being related to the presence of their precursors and to the esterase activities typical of the microbial communities forming the microbiome.


 ,
, 




{kind=link}
{kind=link}
{kind=link}














