
Functional Characterization of Saccharomyces Yeasts from Cider Produced in Hardanger
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Media, and Culture Conditions
2.2. Screening for Sulfite Reductase Activity Using BiGGY Agar
2.3. Micro-Fermentations
2.4. Determination of Sugars, Acids, and Ethanol
Reagents, Materials, and Standards for HPLC–UV/RI Analyses
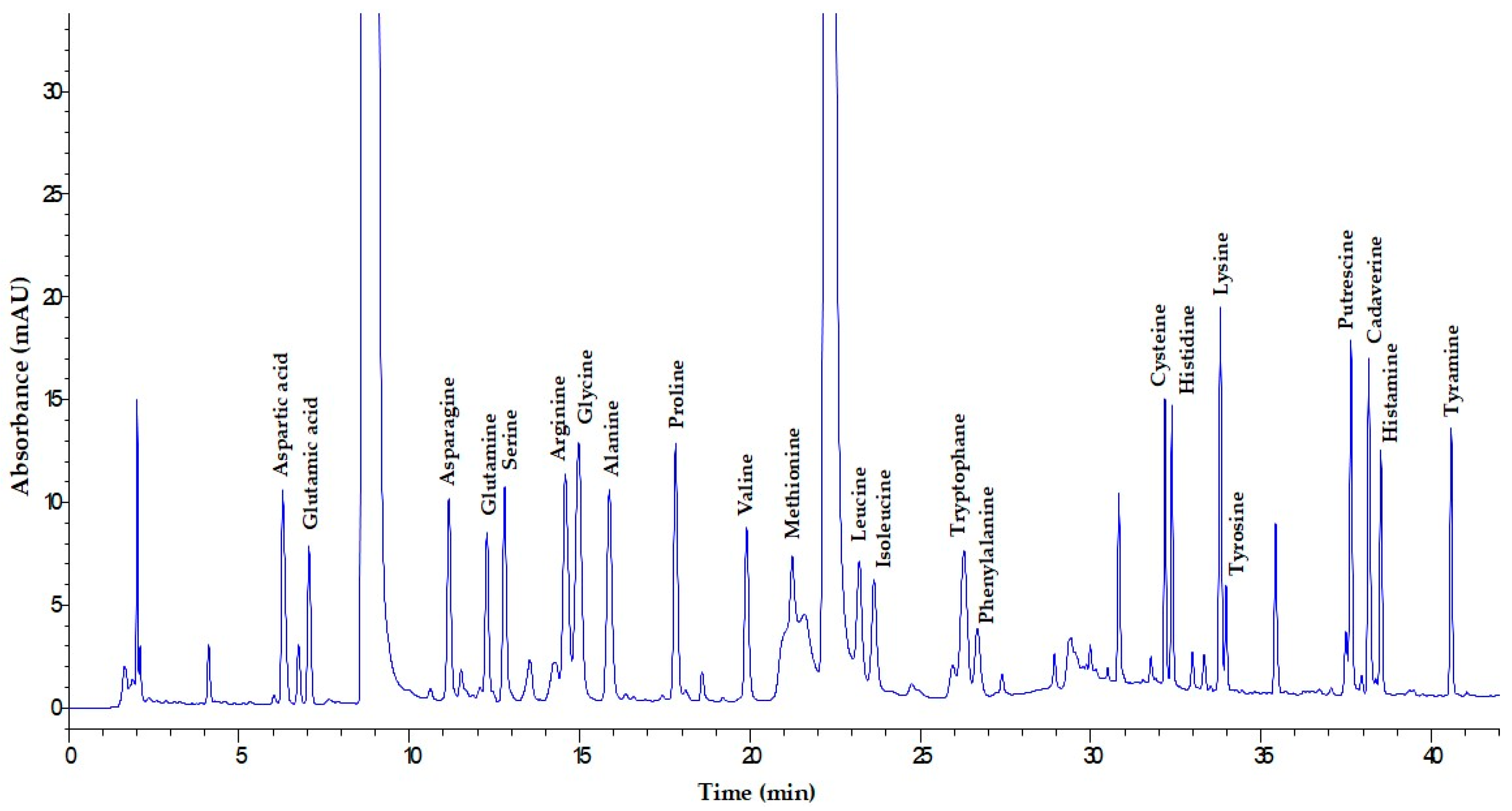
2.5. HPLC–UV Determination of Amino Acids and Biogenic Amines
2.5.1. Reagents, Materials, and Standards for HPLC–UV Analyses
2.5.2. Derivatization Procedure for Determining Amino Acids and Biogenic Amines
2.5.3. HPLC–UV Analysis of AAs and BAs
2.5.4. Method Validation
2.6. Determination of Volatile Compounds Using HS-SPME-GC-MS
2.7. Multivariate Data and Statistical Analysis
3. Results and Discussion
3.1. H2S Production
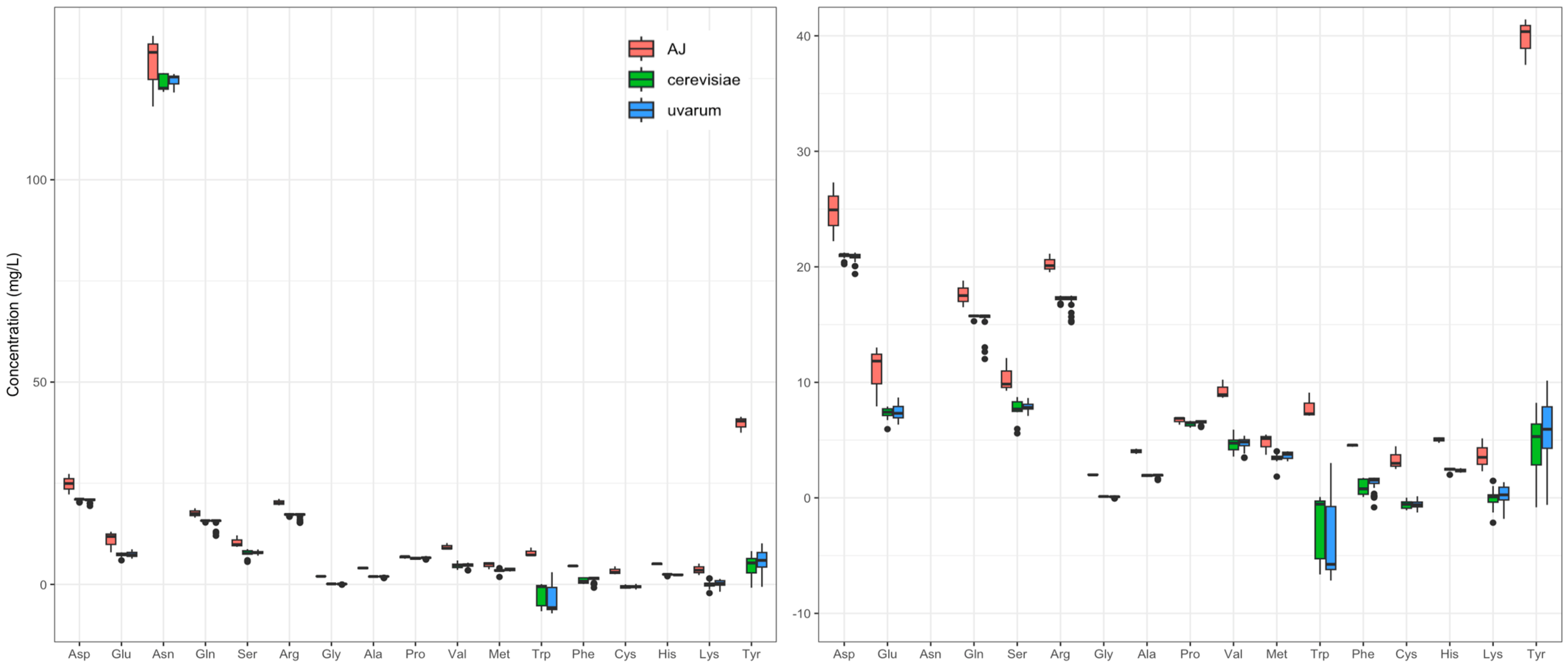
3.2. HPLC–UV Determination of Amino Acids and Biogenic Amines
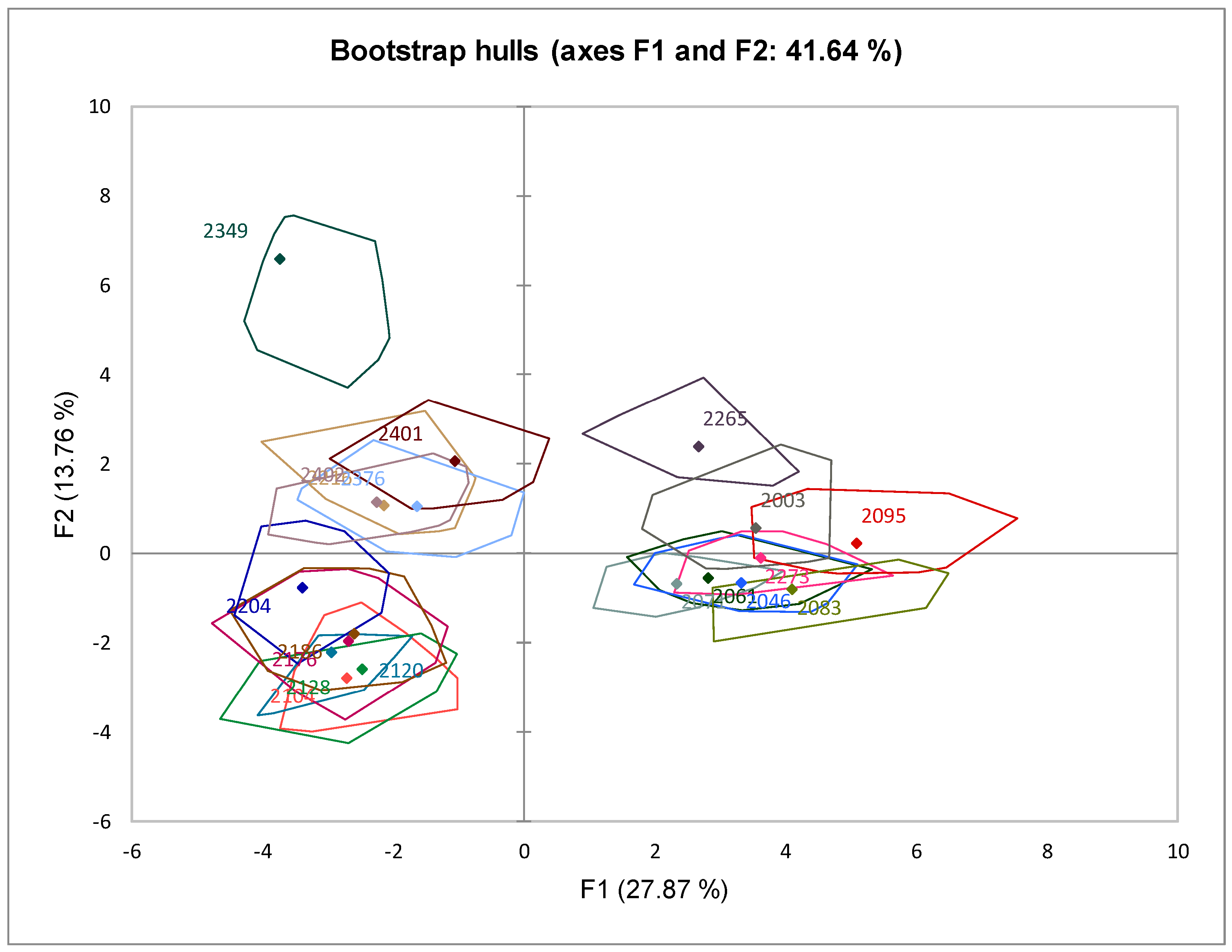
3.3. Behavior of Saccharomyces Strains under Fermentative Conditions
3.4. Volatile Compound Production Profiles of the Saccharomyces Ferments
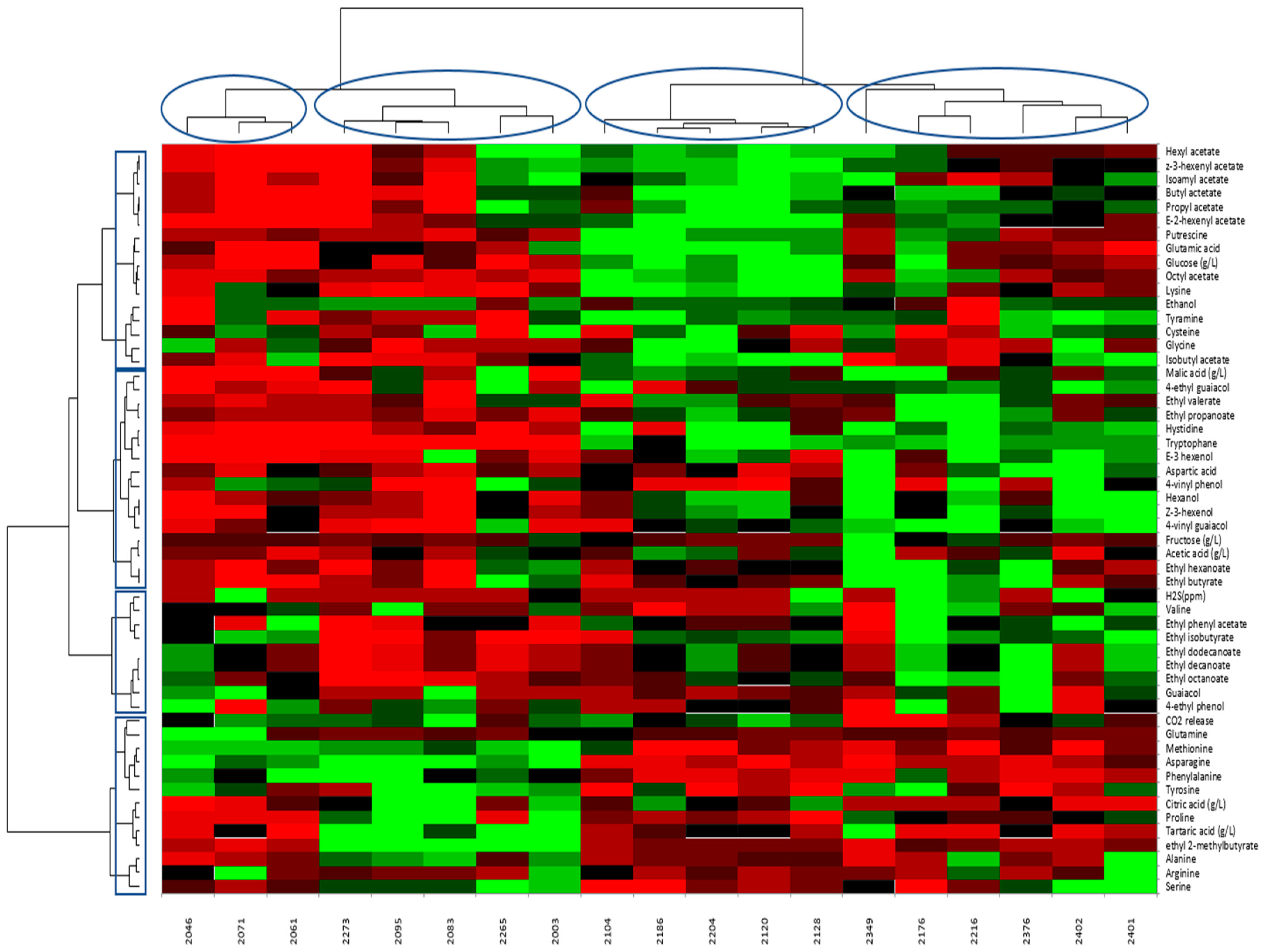
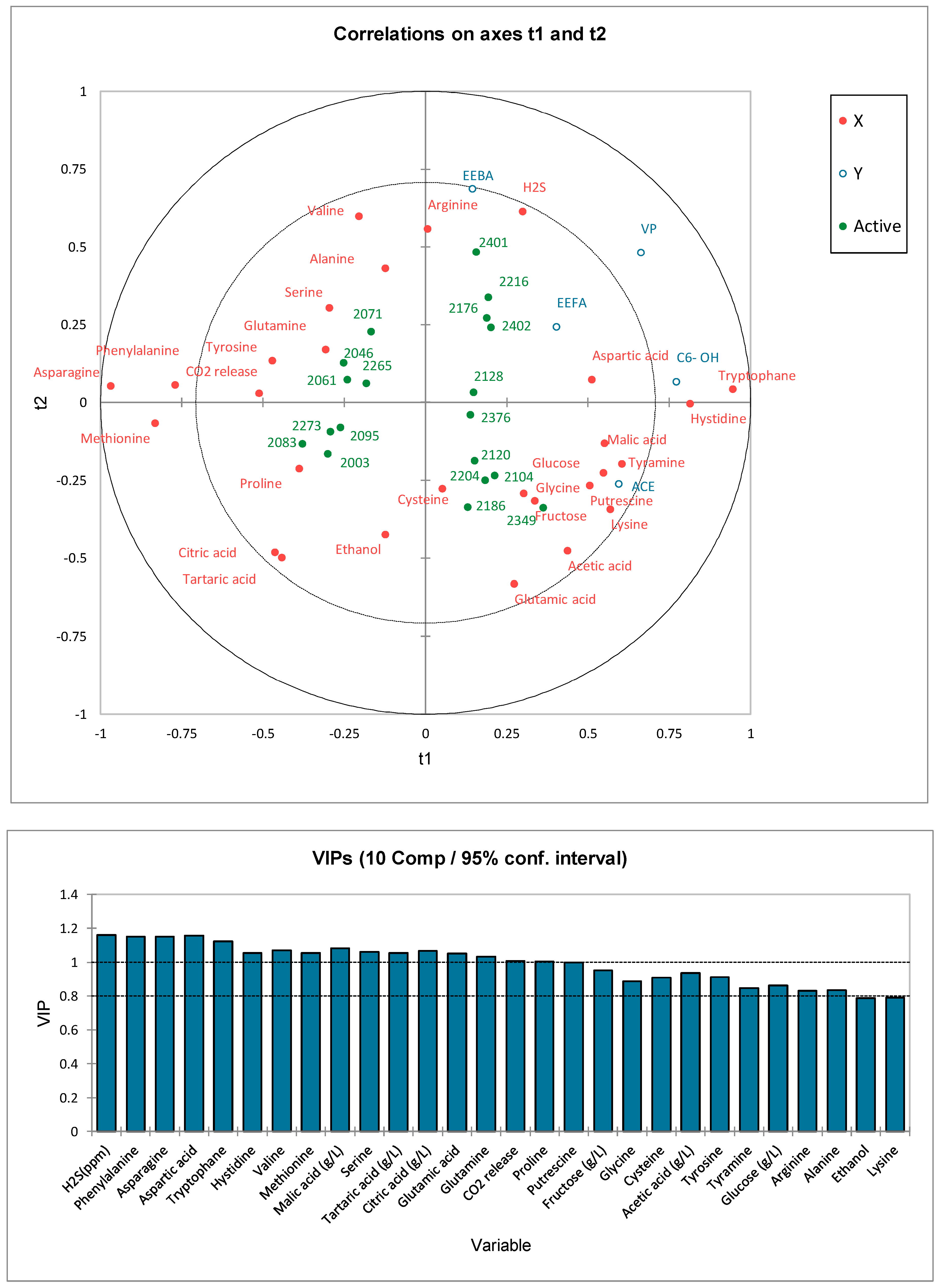
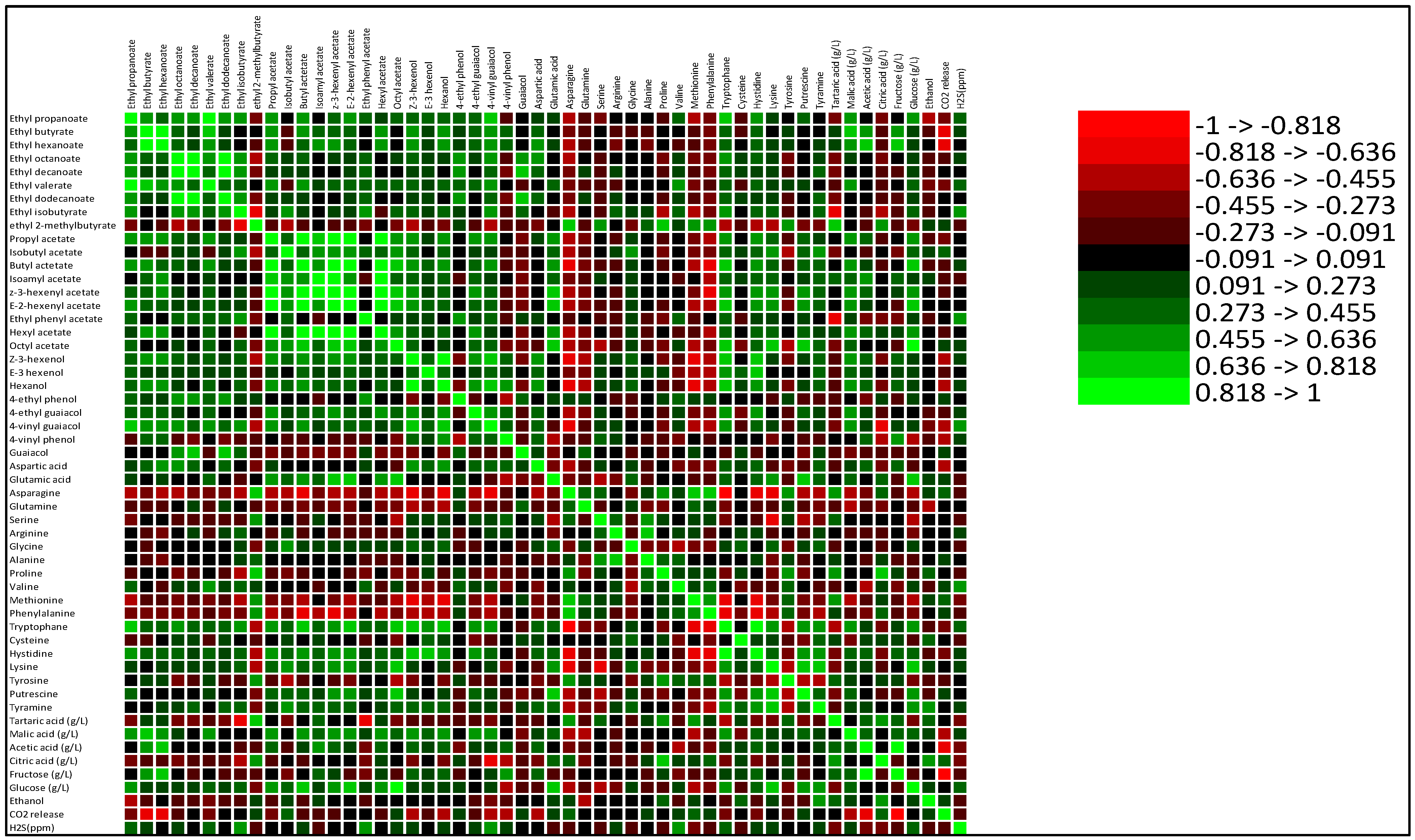
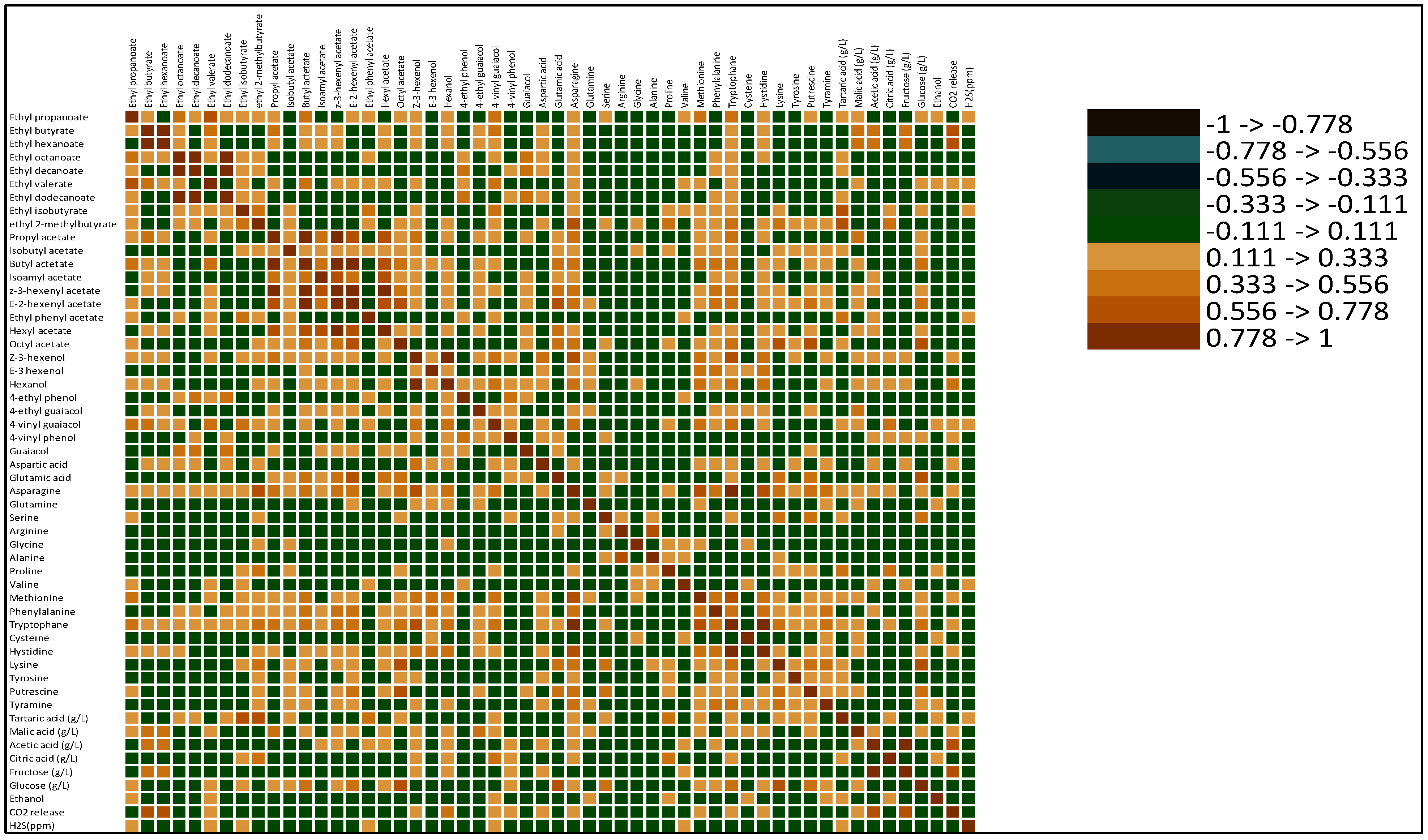
3.5. Correlations of AAs and Physico-Chemical Parameters with Aroma Compound Formation
Data Analysis and Data Configuration
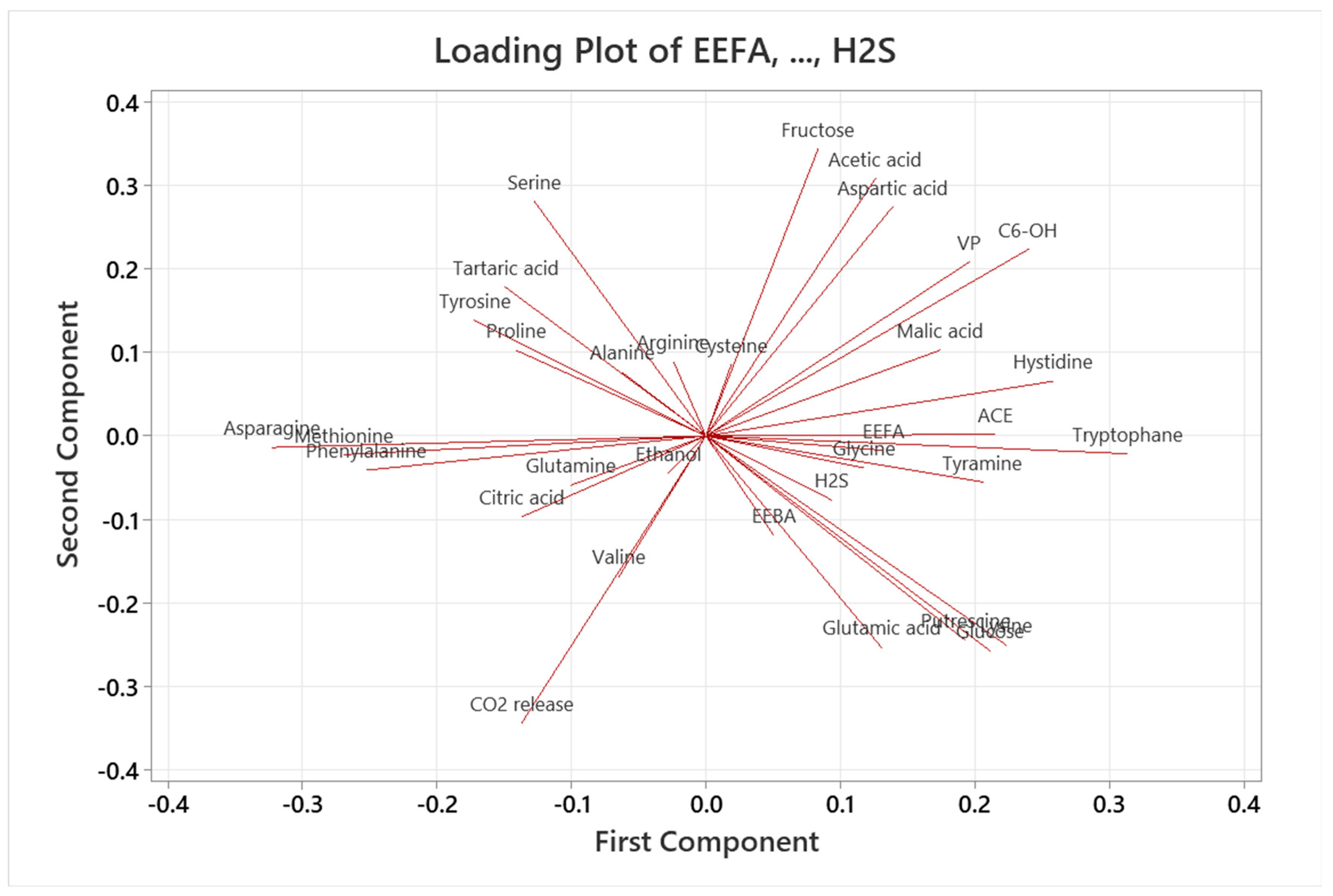
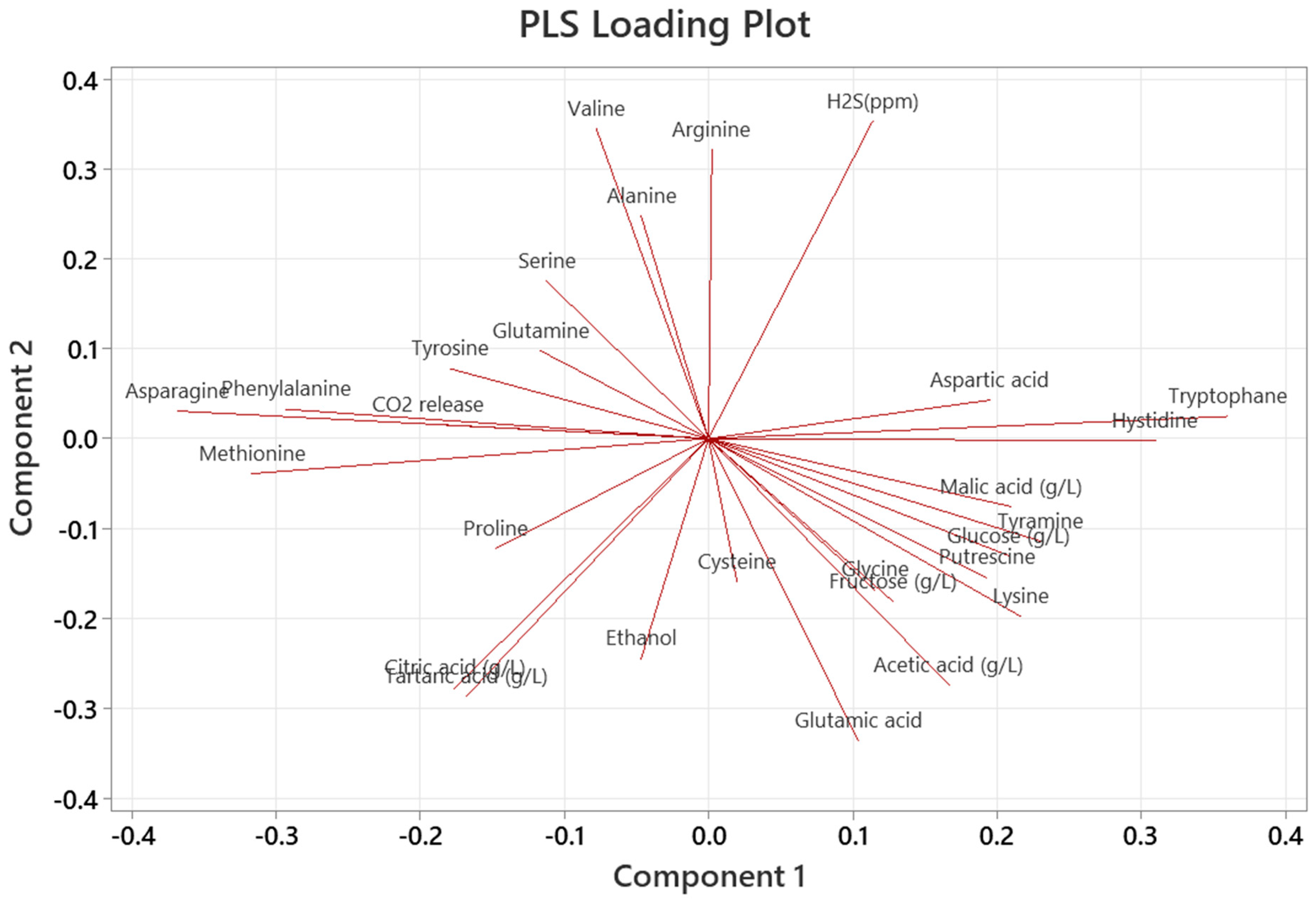
3.6. Relationships between Aroma Compounds and Variables
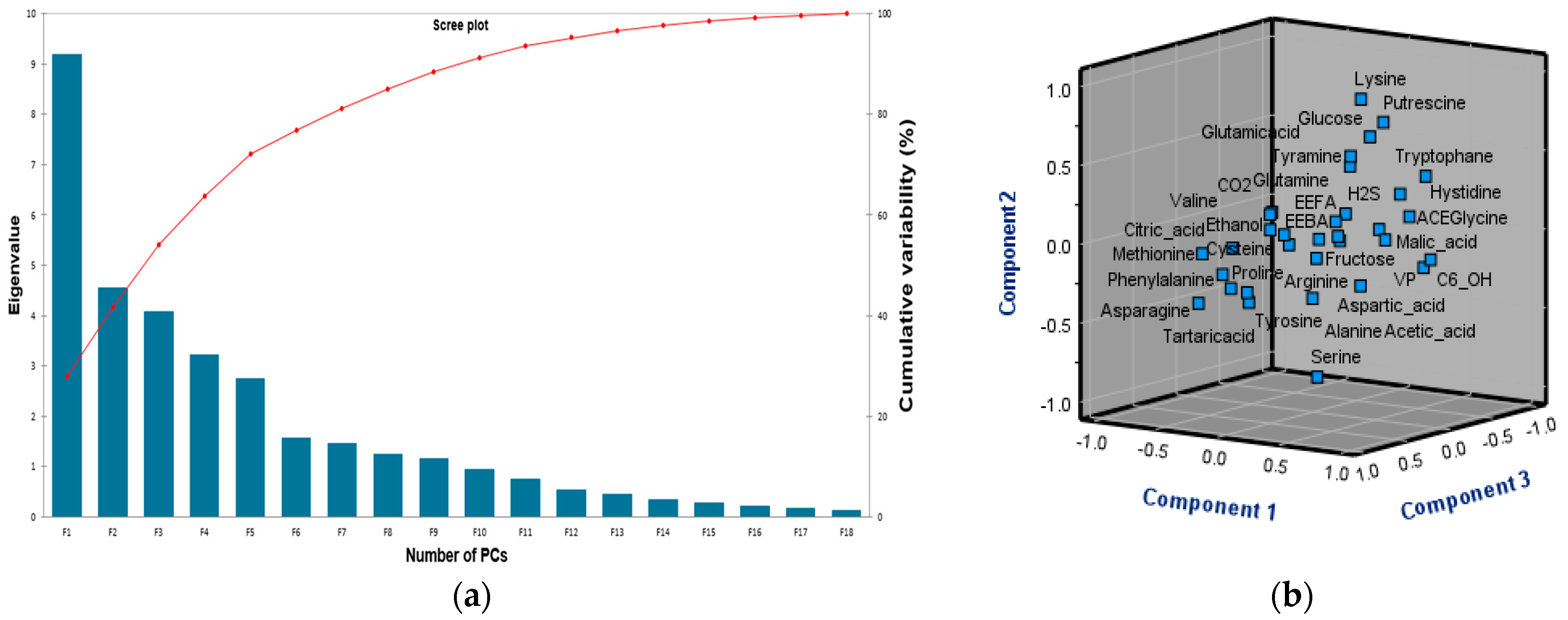
3.6.1. Principal Component Analysis (PCA)
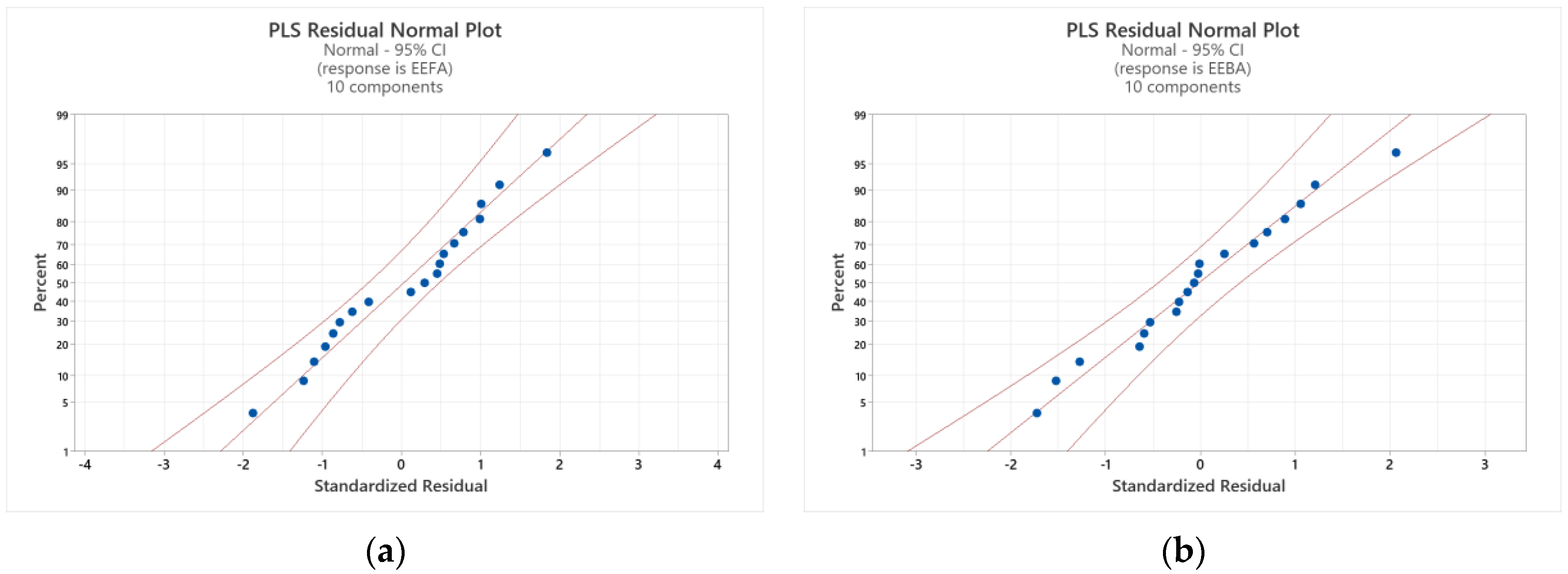
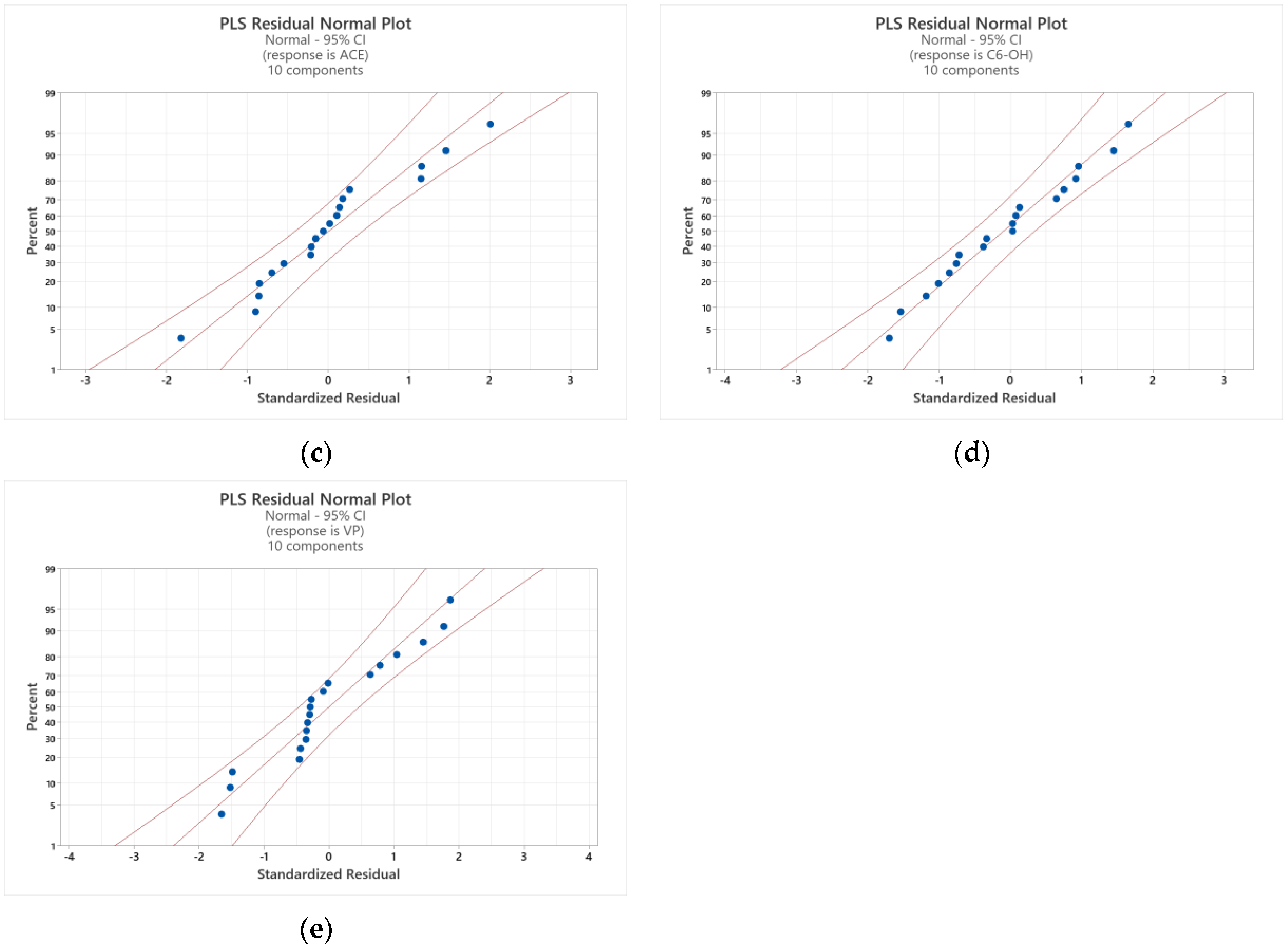
3.6.2. Partial Least Squares (PLS) Regression Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A






References
- Øvsthus, I.; Martenalc, M.; Radovanović Vukajlović, T.; Lesica, M.; Butinar, L.; Mozetič Vodopivec, B.; Antalick, G. Chemical Composition of Apple Cider: A Comparative Study of Norwegian and French Ciders. Acta Hortic. 2023; In press. [Google Scholar]
- Vangdal, E.; Kvamm-Lichtenfeld, K. Ciders Produced from Norwegian Fresh Consumption Apple Cultivars. Acta Hortic. 2018, 1205, 527–532. [Google Scholar] [CrossRef]
- Wicklund, T.; Skottheim, E.R.; Remberg, S.F. Various Factors Affect Product Properties in Apple Cider Production. Int. J. Food Stud. 2020, 9, SI84–SI96. [Google Scholar] [CrossRef]
- Česnik, U.; Øvsthus, I.; Martelanc, M.; Mozetič Vodopivec, B.; Butinar, L. Yeast Diversity from Ciders Produced in Hardanger. Foods, 2023; Article in preparation. [Google Scholar]
- Nguyen, H.; Gaillardin, C. Evolutionary Relationships between the Former Species Saccharomyces Uvarum and the Hybrids Saccharomyces Bayanus and Saccharomyces Pastorianus; Reinstatement of Saccharomyces Uvarum (Beijerinck) as a Distinct Species. FEMS Yeast Res. 2005, 5, 471–483. [Google Scholar] [CrossRef]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe Domestication and the Identification of the Wild Genetic Stock of Lager-Brewing Yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef]
- Almeida, P.; Gonçalves, C.; Teixeira, S.; Libkind, D.; Bontrager, M.; Masneuf-Pomarède, I.; Albertin, W.; Durrens, P.; Sherman, D.J.; Marullo, P.; et al. A Gondwanan Imprint on Global Diversity and Domestication of Wine and Cider Yeast Saccharomyces Uvarum. Nat. Commun. 2014, 5, 4044. [Google Scholar] [CrossRef]
- Marullo, P.; Dubourdieu, D. 12—Yeast Selection for Wine Flavor Modulation. In Managing Wine Quality, 2nd ed.; Reynolds, A.G., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2022; pp. 371–426. ISBN 978-0-08-102065-4. [Google Scholar]
- McCarthy, G.C.; Morgan, S.C.; Martiniuk, J.T.; Newman, B.L.; McCann, S.E.; Measday, V.; Durall, D.M. An Indigenous Saccharomyces Uvarum Population with High Genetic Diversity Dominates Uninoculated Chardonnay Fermentations at a Canadian Winery. PLoS ONE 2021, 16, e0225615. [Google Scholar] [CrossRef]
- Naumov, G.I.; Nguyen, H.-V.; Naumova, E.S.; Michel, A.; Aigle, M.; Gaillardin, C. Genetic Identification of Saccharomyces Bayanus Var. Uvarum, a Cider-Fermenting Yeast. Int. J. Food Microbiol. 2001, 65, 163–171. [Google Scholar] [CrossRef]
- Valles, B.; Bedrinana, R.; Tascon, N.; Simon, A.; Madrera, R. Yeast Species Associated with the Spontaneous Fermentation of Cider. Food Microbiol. 2007, 24, 25–31. [Google Scholar] [CrossRef]
- Blein-Nicolas, M.; Albertin, W.; Valot, B.; Marullo, P.; Sicard, D.; Giraud, C.; Huet, S.; Bourgais, A.; Dillmann, C.; De Vienne, D.; et al. Yeast Proteome Variations Reveal Different Adaptive Responses to Grape Must Fermentation. Mol. Biol. Evol. 2013, 30, 1368–1383. [Google Scholar] [CrossRef] [PubMed]
- McKay, M.; Buglass, A.J.; Gook Lee, C. Cider and Perry. In Handbook of Alcoholic Beverages; Buglass, A.J., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2011; pp. 231–265. ISBN 978-0-470-97652-4. [Google Scholar]
- Wei, J.; Wang, S.; Zhang, Y.; Yuan, Y.; Yue, T. Characterization and Screening of Non-Saccharomyces Yeasts Used to Produce Fragrant Cider. LWT 2019, 107, 191–198. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Wang, Y.; Ju, H.; Niu, C.; Song, Z.; Yuan, Y.; Yue, T. Assessment of Chemical Composition and Sensorial Properties of Ciders Fermented with Different Non-Saccharomyces Yeasts in Pure and Mixed Fermentations. Int. J. Food Microbiol. 2020, 318, 108471. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.T.; Lee, P.-R.; Yu, B.; Liu, S.-Q. Cider Fermentation with Three Williopsis Saturnus Yeast Strains and Volatile Changes. Ann. Microbiol. 2015, 65, 921–928. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Boekhout, T.; Gojkovic, Z.; Katz, M. Evaluation of Non-Saccharomyces Yeasts in the Fermentation of Wine, Beer and Cider for the Development of New Beverages. J. Inst. Brew. 2018, 124, 389–402. [Google Scholar] [CrossRef]
- McKay, M.; Buglass, A.J.; Gook Lee, C. Fermented Beverages: Beers, Ciders, Wines and Related Drinks. In Handbook of Alcoholic Beverages: Technical, Analytical and Nutritional Aspects; John Wiley & Sons, Ltd.: Chichester, UK, 2010; ISBN 978-0-470-51202-9. [Google Scholar]
- Jiranek, V.; Langridge, P.; Henschke, P.A. Validation of Bismuth-Containing Indicator Media for Predicting H2S-Producing Potential of Saccharomyces Cerevisiae Wine Yeasts Under Enological Conditions. Am. J. Enol. Vitic. 1995, 46, 269–273. [Google Scholar] [CrossRef]
- Ugliano, M.; Henschke, P.A. Comparison of Three Methods for Accurate Quantification of Hydrogen Sulfide during Fermentation. Anal. Chim. Acta 2010, 660, 87–91. [Google Scholar] [CrossRef]
- Layne, J.; Lomas, S. Simple Sugars Analysis from Wine by LC-RI Using Luna® Omega SUGAR LC Columns; Phenomenex, Inc.: Torrance, CA, USA, 2018; p. 2. [Google Scholar]
- Martelanc, M. Simultaneous Determination of Chemical Parameters for Wine and Cider Quality Control Using HPLC-UV/RI System Based on Reverse-Phase Separation Capabilities. In Proceedings of the Analytical and Biomedical Applications, Kemomed, Kranj, Slovenia, 20 October 2022. [Google Scholar]
- Nguyen, N.; McGinley, M. Real-Time Response to Bacteria Infection of Bioethanol Fermentation Using a Short RezexTM ROA Column. Appl. Noteb. 2008, 1, 1–4. [Google Scholar]
- Topić Božič, J.; Butinar, L.; Bergant Marušič, M.; Korte, D.; Mozetič Vodopivec, B. Determination of Biogenic Amines Formation by Autochthonous Lactic Acid Bacteria from ‘Refošk’ Grapes Using Different Analytical Methods. LWT 2022, 156, 112908. [Google Scholar] [CrossRef]
- Antalick, G.; Perello, M.-C.; De Revel, G. Esters in Wines: New Insight through the Establishment of a Database of French Wines. Am. J. Enol. Vitic. 2014, 65, 293–304. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of Nitrogen Status in Wine Alcoholic Fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef]
- Ljungdahl, P.O.; Daignan-Fornier, B. Regulation of Amino Acid, Nucleotide, and Phosphate Metabolism in Saccharomyces Cerevisiae. Genetics 2012, 190, 885–929. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Jiang, J.; Ma, Z.; Xiao, M.; Yang, L.; Tian, B.; Yu, Y.; Bi, C.; Fang, A.; Yang, Y. The Role of Hydroxycinnamic Acid Amide Pathway in Plant Immunity. Front. Plant Sci. 2022, 13, 922119. [Google Scholar] [CrossRef] [PubMed]
- Tronchoni, J.; Gamero, A.; Arroyo-López, F.N.; Barrio, E.; Querol, A. Differences in the Glucose and Fructose Consumption Profiles in Diverse Saccharomyces Wine Species and Their Hybrids during Grape Juice Fermentation. Int. J. Food Microbiol. 2009, 134, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Schütz, M.; Gafner, J. Lower Fructose Uptake Capacity of Genetically Characterized Strains of Saccharomyces Bayanus Compared to Strains of Saccharomyces Cerevisiae: A Likely Cause of Reduced Alcoholic Fermentation Activity. Am. J. Enol. Vitic. 1995, 46, 175–180. [Google Scholar] [CrossRef]
- Wang, D.; Xu, Y.; Hu, J.; Zhao, G. Fermentation Kinetics of Different Sugars by Apple Wine Yeast Saccharomyces Cerevisiae. J. Inst. Brew. 2004, 110, 340–346. [Google Scholar] [CrossRef]
- Wicklund, T.; Guyot, S.; Le Quéré, J.-M. Chemical Composition of Apples Cultivated in Norway. Crops 2021, 1, 8–19. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2016; ISBN 978-1-118-62780-8. [Google Scholar]
- Qin, Z.; Petersen, M.A.; Bredie, W.L.P. Flavor Profiling of Apple Ciders from the UK and Scandinavian Region. Food Res. Int. 2018, 105, 713–723. [Google Scholar] [CrossRef]
- He, W.; Liu, S.; Heponiemi, P.; Heinonen, M.; Marsol-Vall, A.; Ma, X.; Yang, B.; Laaksonen, O. Effect of Saccharomyces Cerevisiae and Schizosaccharomyces Pombe Strains on Chemical Composition and Sensory Quality of Ciders Made from Finnish Apple Cultivars. Food Chem. 2021, 345, 128833. [Google Scholar] [CrossRef]
- Medina, S.; Perestrelo, R.; Pereira, R.; Câmara, J.S. Evaluation of Volatilomic Fingerprint from Apple Fruits to Ciders: A Useful Tool to Find Putative Biomarkers for Each Apple Variety. Foods 2020, 9, 1830. [Google Scholar] [CrossRef]
- Satora, P.; Sroka, P.; Duda-Chodak, A.; Tarko, T.; Tuszyński, T. The Profile of Volatile Compounds and Polyphenols in Wines Produced from Dessert Varieties of Apples. Food Chem. 2008, 111, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Vareda, J.; Pereira, R.; Silva, C.; Câmara, J.S.; Perestrelo, R. Geographical Differentiation of Apple Ciders Based on Volatile Fingerprint. Food Res. Int. 2020, 137, 109550. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Li, F.; Cui, L.; Guo, Y. Effects of Fermentation Temperature on Key Aroma Compounds and Sensory Properties of Apple Wine. J. Food Sci. 2015, 80, S2937–S2943. [Google Scholar] [CrossRef]
- Pour Nikfardjam, M.; May, B.; Tschiersch, C. 4-Ethylphenol and 4-Ethylguaiacol Contents in Bottled Wines from the German ‘Württemberg’ Region. Eur. Food Res. Technol. 2009, 230, 333–341. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Species and Strain Code | Colony Color on BiGGy Agar 1 | H2S Detector Tubes 2 (ppm) |
|---|---|---|
| S. uvarum 2046 | white | 0 ± 0.00 E |
| S. uvarum 2071 | white | 35 ± 8.7 BC |
| S. uvarum 2120 | brown | 0 ± 0.00 E |
| S. uvarum 2186 | brown | 0 ± 0.00 E |
| S. uvarum 2401 | white | 13.3 ± 5.8 DE |
| S. cerevisiae 2003 | brown | 10 ± 0.00 DE |
| S. cerevisiae 2095 | brown | 0 ± 0.00 E |
| S. cerevisiae 2265 | white, light brown edge | 0 ± 0.00 E |
| S. cerevisiae 2273 | white | 0 ± 0.00 E |
| S. cerevisiae 2303 | brown | 0 ± 0.00 E |
| S. cerevisiae 2349 | brown | 0 ± 0.00 E |
| S. uvarum 2128 | brown | 36.7 ± 10.4 BC |
| S. uvarum 2204 | brown | 0 ± 0.00 E |
| S. uvarum 2216 | white | 23.3 ± 2.9 CD |
| S. uvarum 2376 | white | 0 ± 0.00 E |
| S. uvarum 2083 | white, light brown edge | 0 ± 0.00 E |
| S. uvarum 2104 | brown | 0 ± 0.00 E |
| S. uvarum 2176 | white | 58.3 ± 18.9 A |
| S. uvarum 2402 | brown | 48.3 ± 2.9 AB |
| S. uvarum 2061 | brown | 0 ± 0.00 E |
| Yeast Species | Strain no. | Acetate Esters(AE) | ||||||||
| Propyl Acetate | Isobutyl Acetate | Butyl Acetate | Isoamyl Acetate | Z-3-Hexenyl Acetate | E-2-Hexenyl Acetate | Ethyl Phenyl Acetate | Hexyl Acetate | OctylAcetate | ||
| S. uvarum | 2104 | 7.72 ± 0.47 ABC | 7.98 ± 1.19 AB | 145.97 ± 3.54 ABC | 69.99 ± 12.52 ABC | 3.71 ± 0.13 AB | 0.35 ± 0.07 ABCD | 1.45 ± 0.75 ABCD | 29.77 ± 0.89 ABC | 2.39 ± 0.29 BC |
| S. uvarum | 2120 | 10.90 ± 0.58 A | 11.11 ± 4.64 A | 220.86 ± 41.45 A | 103.57 ± 52.16 AB | 4.68 ± 0.98 AB | 0.51 ± 0.16 A | 1.03 ± 0.65 BCD | 38.47 ± 9.16 A | 2.76 ± 0.14 AB |
| S. uvarum | 2128 | 9.29 ± 1.32 ABC | 10.01 ± 1.20 ABC | 191.05 ± 25.9 AB | 91.34 ± 11.65 ABC | 4.09 ± 0.10 AB | 0.42 ± 0.07 AB | 1.17 ± 0.07 ABCD | 33.84 ± 1.79 AB | 3.18 ± 0.74 A |
| S. uvarum | 2176 | 9.46 ± 0.43 ABC | 5.28 ± 0.31 ABC | 196.73 ± 5.9 AB | 56.26 ± 9.23 ABC | 3.41 ± 0.02 ABC | 0.35 ± 0.04 ABCD | 2.48 ± 1.02 A | 29.86 ± 1.21 ABC | 2.10 ± 0.09 BCD |
| S. uvarum | 2186 | 9.62 ± 0.99 ABC | 10.07 ± 2.36 AB | 207.93 ± 31.8 AB | 80.29 ± 27.62 ABC | 3.94 ± 0.45 AB | 0.42 ± 0.08 AB | 1.22 ± 0.07 ABCD | 34.65 ± 3.52 AB | 2.15 ± 0.50 BC |
| S. uvarum | 2204 | 10.38 ± 0.63 AB | 9.283 ± 1.86 AB | 213.61 ± 22.2 AB | 90.32 ± 29.43 ABC | 3.84 ± 0.50 AB | 0.40 ± 0.05 ABC | 1.02 ± 0.07 BCD | 31.96 ± 5.61 AB | 2.07 ± 0.38 BCD |
| S. uvarum | 2216 | 9.29 ± 1.02 ABC | 4.84 ± 0.60 ABC | 194.17 ± 48.7 AB | 39.08 ± 8.34 ABC | 3.14 ± 0.60 ABCD | 0.36 ± 0.04 ABCD | 1.23 ± 0.06 ABCD | 23.87 ± 6.13 ABCD | 1.94 ± 0.37 CDE |
| S. cerevisiae | 2349 | 8.94 ± 1.36 ABC | 4.40 ± 0.61 BC | 161.28 ± 18.41 ABC | 99.41 ± 23.76 AB | 3.52 ± 0.41 AB | 0.27 ± 0.04 ABCD | 0.48 ± 0.05 CD | 33.91 ± 4.70 AB | 1.01 ± 0.0.3 F |
| S. uvarum | 2376 | 9.10 ± 2.46 ABC | 6.91 ± 2.59 ABC | 158.67 ± 61.76 ABC | 49.23 ± 24.20 ABC | 2.81 ± 0.84 ABCD | 0.32 ± 0.08 ABCD | 1.36 ± 1.00 ABCD | 22.91 ± 8.88 ABCD | 0.99 ± 0.0.6 F |
| S. uvarum | 2402 | 8.41 ± 1.47 ABC | 9.25 ± 2.37 AB | 166.11 ± 49.54 ABC | 65.48 ± 20.68 ABC | 3.09 ± 0.81 ABCD | 0.31 ± 0.11 ABCD | 1.81 ± 0.42 ABC | 24.66 ± 8.59 ABCD | 1.31 ± 0.0.5 DEF |
| S. uvarum | 2401 | 9.00 ± 1.94 ABC | 10.20 ± 3.26 ABC | 159.13 ± 43.61 ABC | 86.14 ± 28.94 ABC | 3.08 ± 0.92 ABCD | 0.28 ± 0.07 ABCD | 1.27 ± 0.24 ABCD | 22.25 ± 9.79 ABCD | 1.17 ± 0.08 EF |
| S. uvarum | 2071 | 5.82 ± 1.36 BC | 4.98 ± 1.71 ABC | 95.61 ± 39.4 BC | 29.48 ± 18.71 BC | 1.36 ± 0.69 D | 0.15 ± 0.07 D | 0.74 ± 0.16 CD | 8.48 ± 4.83 D | 0.87 ± 0.08 F |
| S. uvarum | 2061 | 5.63 ± 0.99 C | 9.60 ± 3.30 AB | 63.46 ± 53.23 C | 45.96 ± 20.88 ABC | 1.42 ± 0.68 CD | 0.16 ± 0.09 CD | 2.24 ± 0.56 AB | 9.25 ± 6.02 CD | 1.17 ± 0.09 EF |
| S. uvarum | 2046 | 7.29 ± 2.01 ABC | 6.17 ± 2.77 ABC | 124.13 ± 35.94 ABC | 50.56 ± 26.21 ABC | 2.17 ± 0.76 BCD | 0.20 ± 0.09 BCD | 1.21 ± 0.30 ABCD | 17.46 ± 8.16 BCD | 0.90 ± 0.09 F |
| S. cerevisiae | 2265 | 10.43 ± 2.05 AB | 5.82 ± 1.32 ABC | 170.95 ± 51.36 ABC | 88.59 ± 18.02 ABC | 3.66 ± 0.67 AB | 0.33 ± 0.10 ABCD | 1.15 ± 0.06 ABCD | 35.88 ± 8.79 AB | 1.11 ± 0.05 F |
| S. cerevisiae | 2273 | 6.73 ± 1.40 ABC | 2.36 ± 0.60 C | 105.34 ± 32.26 ABC | 12.01 ± 1.02 C | 1.23 ± 0.36 D | 0.21 ± 0.11 BCD | 0.29 ± 0.07 D | 8.98 ± 4.03 D | 1.12 ± 0.03 F |
| S. cerevisiae | 2003 | 9.12 ± 1.88 ABC | 6.94 ± 1.97 ABC | 171.20 ± 50.52 ABC | 118.73 ± 62.59 A | 3.89 ± 0.24 AB | 0.32 ± 0.08 ABCD | 0.68 ± 0.14 CD | 39.72 ± 12.89 A | 0.95 ± 0.01 F |
| S. cerevisiae | 2095 | 7.65 ± 1.14 ABC | 4.93 ± 0.56 ABC | 119.12 ± 21.09 ABC | 59.46 ± 3.74 ABC | 2.54 ± 0.29 BCD | 0.25 ± 0.03 ABCD | 0.66 ± 0.03 CD | 24.27 ± 2.46 ABCD | 1.06 ± 0.16 F |
| S. uvarum | 2083 | 5.99 ± 0.74 BC | 4.93 ± 1.12 ABC | 111.12 ± 30.50 ABC | 35.92 ± 13.09 ABC | 2.17 ± 0.46 BCD | 0.26 ± 0.08 ABCD | 1.20 ± 0.47 ABCD | 19.74 ± 5.42 ABCD | 0.95 ± 0.05 F |
| Yeast Species | Strain no. | Ethyl Esters from Fatty Acids(EEFA) | ||||||||
| Ethyl propanoate | Ethyl butyrate | Ethyl hexanoate | Ethyl octanoate | Ethyl decanoate | Ethyl valerate | Ethyl dodecanoate | ||||
| S. uvarum | 2104 | 57.36 ± 4.95 CDE | 45.16 ± 1.23 FG | 86.53 ± 3.80 EFGH | 291.56 ± 45.08 EFGHI | 247.30 ± 33.52 DEFG | 0.42 ± 0.03 CD | 258.68 ± 32.96 DEFG | ||
| S. uvarum | 2120 | 70.66 ± 16.71 BCD | 57.37 ± 7.25 CDEFG | 121.80 ± 21.89 DEFG | 373.86 ± 67.47 CDEFG | 392.82 ± 90.60 DEF | 0.59 ± 0.13 BCD | 401.77 ± 89.09 DEF | ||
| S. uvarum | 2128 | 53.38 ± 9.49 DE | 52.17 ± 3.14 EFG | 120.43 ± 7.44 DEFG | 425.35 ± 76.54 BCDEF | 488.00 ± 63.42 CD | 0.52 ± 0.7 BCD | 495.37 ± 62.37 CD | ||
| S. uvarum | 2176 | 158.94 ± 10.53 A | 93.79 ± 5.46 AB | 191.53 ± 16.28 BCDEF | 606.22 ± 49.90 B | 887.72 ± 87.30 B | 1.39 ± 0.08 A | 888.43 ± 85.84 B | ||
| S. uvarum | 2186 | 69.13 ± 11.88 CD | 55.23 ± 5.87 DEFG | 123.36 ± 14.0 CDEFG | 359.55 ± 38.35 CDEFGH | 468.80 ± 103.79 CD | 0.82 ± 0.17 B | 476.49 ± 102.06 CD | ||
| S. uvarum | 2204 | 100.91 ± 9.56 B | 59.85 ± 3.87 CDEFG | 110.91 ± 6.62 DEFGH | 452.85 ± 25.5 BCDE | 741.37 ± 50.63 BC | 0.82 ± 0.05 B | 744.51 ± 49.76 BC | ||
| S. uvarum | 2216 | 172.37 ± 10.62 A | 74.33 ± 3.9 BCDE | 133.56 ± 8.04 BCDEF | 548.31 ± 6.95 BC | 463.66 ± 16.62 D | 1.18 ± 0.02 A | 471.43 ± 16.35 D | ||
| S. cerevisiae | 2349 | 41.54 ± 6.13 DEF | 104.21 ± 17.0 A | 256.10 ± 49.27 A | 339.71 ± 77.20 DEFGH | 141.68 ± 27.57 FG | 0.61 ± 0.12 BCD | 154.82 ± 27.11 FG | ||
| S. uvarum | 2376 | 88.12 ± 19.10 BC | 80.15 ± 16.9 ABCD | 185.19 ± 30.43 BC | 973.86 ± 40.27 A | 1905.93 ± 189.17 A | 0.73 ± 0.20 BC | 1889.66 ± 186.02 A | ||
| S. uvarum | 2402 | 45.20 ± 8.91 DEF | 49.15 ± 5.69 EFG | 106.44 ± 11.39 DEFGH | 293.59 ± 93.67 DEFGHI | 147.69 ± 57.34 FG | 0.50 ± 0.05 BCD | 160.72 ± 56.39 FG | ||
| S. uvarum | 2401 | 70.76 ± 12.89 BCD | 55.83 ± 7.83 CDEFG | 83.51 ± 18.44 FGH | 485.70 ± 60.40 BCD | 911.34 ± 145.23 B | 0.57 ± 0.07 BCD | 911.66 ± 142.80 B | ||
| S. uvarum | 2071 | 30.65 ± 7.57 EF | 40.60 ± 5.69 G | 56.81 ± 16.0 H | 320.02 ± 29.51 DEFGH | 434.32 ± 136.17 DE | 0.39 ± 0.03 D | 442.58 ± 133.90 DE | ||
| S. uvarum | 2061 | 31.84 ± 1.64 EF | 40.55 ± 6.28 G | 101.68 ± 25.6 DEFGH | 402.24 ± 151.09 CDEFG | 246.06 ± 90.58 DEFG | 0.45 ± 0.08 CD | 257.46 ± 89.07 DEFG | ||
| S. uvarum | 2046 | 43.61 ± 15.19 DEF | 47.99 ± 8.93 FG | 83.97 ± 21.60 FGH | 459.05 ± 92.84 BCDE | 813.25 ± 137.77 B | 0.46 ± 0.10 CD | 815.20 ± 135.47 B | ||
| S. cerevisiae | 2265 | 40.92 ± 5.91 DEF | 80.59 ± 8.62 ABC | 147.85 ± 16.34 BCDE | 247.14 ± 17.84 FGHI | 67.17 ± 13.19 FG | 0.69 ± 0.06 BCD | 81.55 ± 12.97 G | ||
| S. cerevisiae | 2273 | 30.44 ± 7.57 EF | 45.83 ± 7.78 FG | 63.54 ± 15.44 GH | 113.50 ± 23.76 I | 0.009 ± 0.01 G | 0.45 ± 0.08 CD | 15.50 ± 0.01 G | ||
| S. cerevisiae | 2003 | 21.63 ± 5.0 F | 68.59 ± 11.4 BCDEF | 153.89 ± 26.90 BCD | 345.91 ± 39.18 DEFGH | 168.46 ± 38.26 EFG | 0.73 ± 0.23 BC | 181.15 ± 37.63 EFG | ||
| S. cerevisiae | 2095 | 43.44 ± 10.08 DEF | 51.09 ± 4.62 EFG | 95.96 ± 10.40 DEFGH | 177.46 ± 18.36 HI | 21.61 ± 19.33 G | 0.61 ± 0.10 BCD | 36.75 ± 19.01 G | ||
| S. uvarum | 2083 | 25.70 ± 1.83 EF | 39.03 ± 4.13 G | 70.54 ± 13.79 GH | 223.80 ± 52.53 GHI | 269.30 ± 78.80 DEFG | 0.36 ± 0.07 D | 280.31 ± 77.48 DEFG | ||
| Yeast Species | Strain no. | Ethyl Esters from Branched Acids(EEBA) | C6-Alcohols(C6-OH) | |||||||
| Ethyl isobutyrate | Ethyl 2-methylbutyrate | Z-3-Hexenol | E-3-Hexenol | Hexanol | ||||||
| S. uvarum | 2104 | 2.56 ± 0.34 CDEF | 0.46 ± 0.04 A | 140.19 ± 6.51 ABCD | 4.39 ± 0.46 A | 1731.77 ± 89.15 BCDE | ||||
| S. uvarum | 2120 | 4.92 ± 1.69 ABC | 0.64 ± 0.20 A | 176.86 ± 25.80 ABC | 5.00 ± 0.52 A | 2171.38 ± 229.22 ABC | ||||
| S. uvarum | 2128 | 4.93 ± 0.76 ABC | 0.64 ± 0.09 A | 151.29 ± 15.1 ABCD | 3.94 ± 0.41 A | 1797.3 ± 218.7 BCDE | ||||
| S. uvarum | 2176 | 6.69 ± 0.60 A | 0.74 ± 0.03 A | 148.12 ± 17.15 ABCDE | 4.58 ± 0.38 A | 1869.75 ± 83.91 ABCDE | ||||
| S. uvarum | 2186 | 4.65 ± 0.58 ABCD | 0.57 ± 0.09 A | 154.85 ± 2.82 ABCD | 4.65 ± 0.36 A | 1967.90 ± 44.01 ABCD | ||||
| S. uvarum | 2204 | 4.31 ± 0.21 ABCDE | 0.56 ± 0.05 A | 168.81 ± 12.2 ABC | 5.42 ± 0.40 A | 2208.67 ± 161.80 ABC | ||||
| S. uvarum | 2216 | 5.05 ± 0.34 ABC | 0.66 ± 0.06 A | 181.20 ± 10.05 AB | 5.53 ± 0.92 A | 2234.25 ± 185.44 ABC | ||||
| S. cerevisiae | 2349 | 2.81 ± 0.48 CDEF | 0.36 ± 0.09 A | 191.59 ± 7.24 A | 5.69 ± 0.30 A | 2539.14 ± 96.89 A | ||||
| S. uvarum | 2376 | 4.28 ± 0.83 ABCDE | 0.54 ± 0.14 A | 158.60 ± 10.66 ABCD | 5.08 ± 0.45 A | 1803.84 ± 127.14 BCDE | ||||
| S. uvarum | 2402 | 4.67 ± 1.13 ABCD | 0.52 ± 0.12 A | 187.92 ± 10.84 A | 5.86 ± 0.72 A | 2417.87 ± 172.85 AB | ||||
| S. uvarum | 2401 | 6.58 ± 1.62 A | 0.62 ± 0.14 A | 181.73 ± 11.85 AB | 5.16 ± 0.94 A | 2242.75 ± 140.58 ABC | ||||
| S. uvarum | 2071 | 5.53 ± 1.41 AB | 0.37 ± 0.12 A | 121.63 ± 22.1 CDE | 3.67 ± 0.54 A | 1626.81 ± 308.32 CDE | ||||
| S. uvarum | 2061 | 5.14 ± 1.10 ABC | 0.44 ± 0.09 A | 148.37 ± 29.6 ABCD | 3.87 ± 0.44 A | 1841.51 ± 434.03 ABCDE | ||||
| S. uvarum | 2046 | 4.09 ± 1.01 ABCDE | 0.43 ± 0.15 A | 108.39 ± 14.9 DE | 3.75 ± 0.78 A | 1353.55 ± 238.15 DE | ||||
| S. cerevisiae | 2265 | 2.13 ± 0.24 DEF | 2.28 ± 0.38 A | 147.23 ± 9.61 ABCDE | 4.26 ± 0,97 A | 1883.32 ± 143.94 ABC | ||||
| S. cerevisiae | 2273 | 1.20 ± 0.46 F | 2.17 ± 0.46 A | 129.62 ± 10.8 BCDE | 3.90 ± 0.35 A | 1749.91 ± 106.43 BCDE | ||||
| S. cerevisiae | 2003 | 2.24 ± 0.60 DEF | 2.35 ± 0.31 A | 129.60 ± 28.17 BCDE | 4.02 ± 1.21 A | 1622.19 ± 377.31 CDE | ||||
| S. cerevisiae | 2095 | 1.97 ± 0.51 EF | 2.28 ± 0.35 A | 128.68 ± 36.2 BCDE | 3.96 ± 0.28 A | 1655.67 ± 447.66 CDE | ||||
| S. uvarum | 2083 | 3.23 ± 0.20 BCDEF | 2.26 ± 0.32 A | 92.40 ± 20.39 E | 6.43 ± 4.27 A | 1222.57 ± 292.76 E | ||||
| Yeast Species | Strain no. | VolatilePhenols(VP) | ||||||||
| 4-Ethyl phenol | 4-Ethyl guaiacol | 4-Vinyl guaiacol | 4-Vinyl phenol | Guaiacol | ||||||
| S. uvarum | 2104 | 5.74 ± 0.93 BCD | 0.36 ± 0.15 AB | 609.05 ± 116.3 ABCDE | 212.40 ± 37.09 ABCDE | 1.10 ± 0.09 A | ||||
| S. uvarum | 2120 | 7.35 ± 1.12 ABCD | 0.29 ± 0.09 AB | 1087.30 ± 325.63 E | 29.27 ± 19.12 E | 1.45 ± 0.72 A | ||||
| S. uvarum | 2128 | 6.83 ± 0.75 BCD | 0.29 ± 0.01 AB | 1456.3 ± 17.87 BCDE | 153.97 ± 198.2 BCDE | 2.40 ± 2.10 A | ||||
| S. uvarum | 2176 | 12.91 ± 0.31 A | 0.30 ± 0.02 AB | 2001.80 ± 161.37 E | 39.46 ± 7.96 E | 3.42 ± 4.89 A | ||||
| S. uvarum | 2186 | 5.75 ± 1.31 BCD | 0.23 ± 0.03 AB | 1175.05 ± 273.51 E | 39.72 ± 11.56 E | 2.09 ± 2.22 A | ||||
| S. uvarum | 2204 | 7.29 ± 0.60 ABCD | 0.26 ± 0.02 AB | 1216.44 ± 149.23 DE | 47.71 ± 8.01 DE | 0.91 ± 0.24 A | ||||
| S. uvarum | 2216 | 6.15 ± 0.30 BCD | 0.32 ± 0.01 AB | 2121.76 ± 107.63 ABCD | 413.98 ± 324 ABCD | 1.80 ± 1.32 A | ||||
| S. cerevisiae | 2349 | 4.10 ± 0.96 CD | 0.29 ± 0.05 AB | 1632.24 ± 134.28 A | 561.04 ± 26.01 A | 0.89 ± 0.81 A | ||||
| S. uvarum | 2376 | 12.92 ± 2.33 A | 0.29 ± 0.03 AB | 1199.19 ± 156.97 CDE | 81.33 ± 14.71 CDE | 11.91 ± 17.74 A | ||||
| S. uvarum | 2402 | 5.07 ± 0.78 BCD | 0.34 ± 0.08 AB | 1599.26 ± 121.19 AB | 473.78 ± 33.05 AB | 0.60 ± 0.40 A | ||||
| S. uvarum | 2401 | 7.56 ± 1.09 ABCD | 0.31 ± 0.03 AB | 1793.76 ± 158.55 BCDE | 187.33 ± 259.49 BCDE | 3.38 ± 0.13 A | ||||
| S. uvarum | 2071 | 2.66 ± 0.90 D | 0.23 ± 0.04 AB | 884.52 ± 148.71 ABCDE | 327.35 ± 47.5 ABCDE | 8.99 ± 14.11 A | ||||
| S. uvarum | 2061 | 9.06 ± 5.41 ABC | 0.22 ± 0.03 AB | 1085.67 ± 366.57 ABCDE | 274.44 ± 206 ABCDE | 3.35 ± 0.13 A | ||||
| S. uvarum | 2046 | 10.82 ± 2.52 AB | 0.18 ± 0.03 B | 653.55 ± 113.66 CDE | 82.09 ± 35.37 CDE | 4.82 ± 4.44 A | ||||
| S. cerevisiae | 2265 | 6.57 ± 1.83 BCD | 0.40 ± 0.04 A | 1608.44 ± 145.22 ABC | 418.23 ± 29.5 ABC | 1.08 ± 0.17 A | ||||
| S. cerevisiae | 2273 | 6.46 ± 0.95 BCD | 0.17 ± 0.04 B | 610.93 ± 127.20 ABCDE | 228.56 ± 43.8 ABCDE | 0.93 ± 0.18 A | ||||
| S. cerevisiae | 2003 | 8.12 ± 1.82 ABCD | 0.24 ± 0.07 AB | 680.06 ± 196.04 ABCDE | 246.31 ± 78.99 ABCDE | 1.09 ± 0.48 A | ||||
| S. cerevisiae | 2095 | 8.33 ± 2.80 ABCD | 0.28 ± 0.18 AB | 36.01 ± 10.53 E | 18.18 ± 2.88 E | 0.75 ± 0.02 A | ||||
| S. uvarum | 2083 | 9.58 ± 1.53 ABC | 0.21 ± 0.06 AB | 357.71 ± 34.07 E | 26.92 ± 2.24 E | 6.30 ± 1.32 A | ||||
| PLS Regression: Model Selection and Validation for Different Families of Aroma Attributes. | ||||
|---|---|---|---|---|
| Y-Block (i.e., Aroma Groups of Attributes) (y Variables, n = 5) | Number of Latent Variables (LVs) | R-Sq (R2) | Captured Cumulative X-Block Variance (R2X cum) | Captured Cumulative Y-Block Variance (R2Y cum) |
| Ethyl esters from fatty acids | 10 * | 0.96 | 0.89 | 0.93 |
| Ethyl esters of branched acids | 0.92 | |||
| Acetate esters | 0.95 | |||
| C6-alcohols | 0.86 | |||
| Volatile phenols | 0.94 | |||
| Ethyl esters from fatty acids | 9 | 0.96 | 0.86 | 0.90 |
| Ethyl esters of branched acids | 0.91 | |||
| Acetate esters | 0.87 | |||
| C6-alcohols | 0.86 | |||
| Volatile phenols | 0.92 | |||
| Ethyl esters from fatty acids | 8 | 0.87 | 0.82 | 0.88 |
| Ethyl esters of branched acids | 0.91 | |||
| Acetate esters | 0.86 | |||
| C6-alcohols | 0.86 | |||
| Volatile phenols | 0.92 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Česnik, U.; Martelanc, M.; Øvsthus, I.; Radovanović Vukajlović, T.; Hosseini, A.; Mozetič Vodopivec, B.; Butinar, L. Functional Characterization of Saccharomyces Yeasts from Cider Produced in Hardanger. Fermentation 2023, 9, 824. https://doi.org/10.3390/fermentation9090824
Česnik U, Martelanc M, Øvsthus I, Radovanović Vukajlović T, Hosseini A, Mozetič Vodopivec B, Butinar L. Functional Characterization of Saccharomyces Yeasts from Cider Produced in Hardanger. Fermentation. 2023; 9(9):824. https://doi.org/10.3390/fermentation9090824
Chicago/Turabian StyleČesnik, Urban, Mitja Martelanc, Ingunn Øvsthus, Tatjana Radovanović Vukajlović, Ahmad Hosseini, Branka Mozetič Vodopivec, and Lorena Butinar. 2023. "Functional Characterization of Saccharomyces Yeasts from Cider Produced in Hardanger" Fermentation 9, no. 9: 824. https://doi.org/10.3390/fermentation9090824