
Effects of Replacing Alfalfa Hay with Oat Hay in Fermented Total Mixed Ration on Growth Performance and Rumen Microbiota in Lambs

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Preparation of FTMR
2.3. Feed Composition Analysis
2.4. Animals, Diets, and Experimental Design
2.5. Feed Intake, Growth Performance, and Rumen Samples Collection
2.6. Bacterial DNA Extraction, Polymerase Chain Reaction Amplification, and 16S rDNA Sequencing
2.7. Bioinformatics Analysis
2.8. Statistical Analysis
3. Results
3.1. Animal Performance
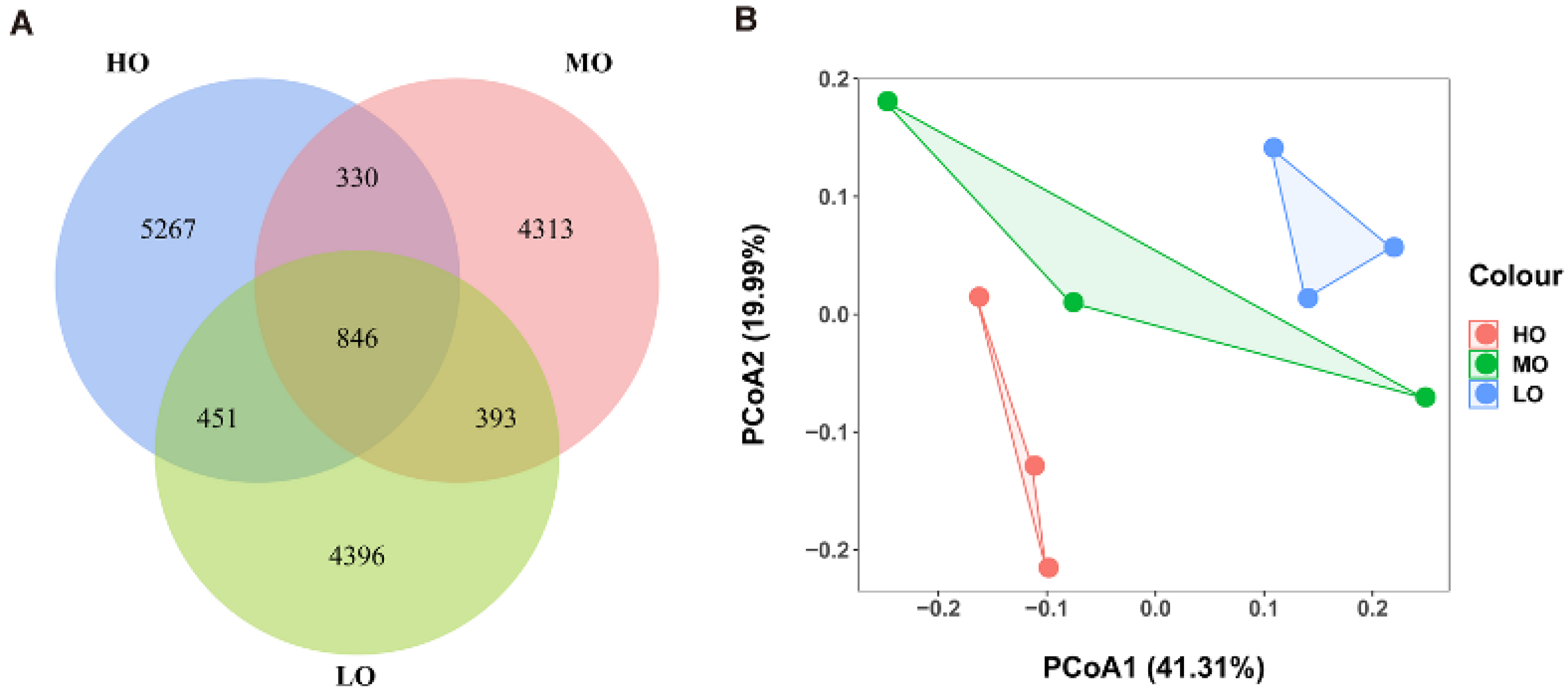
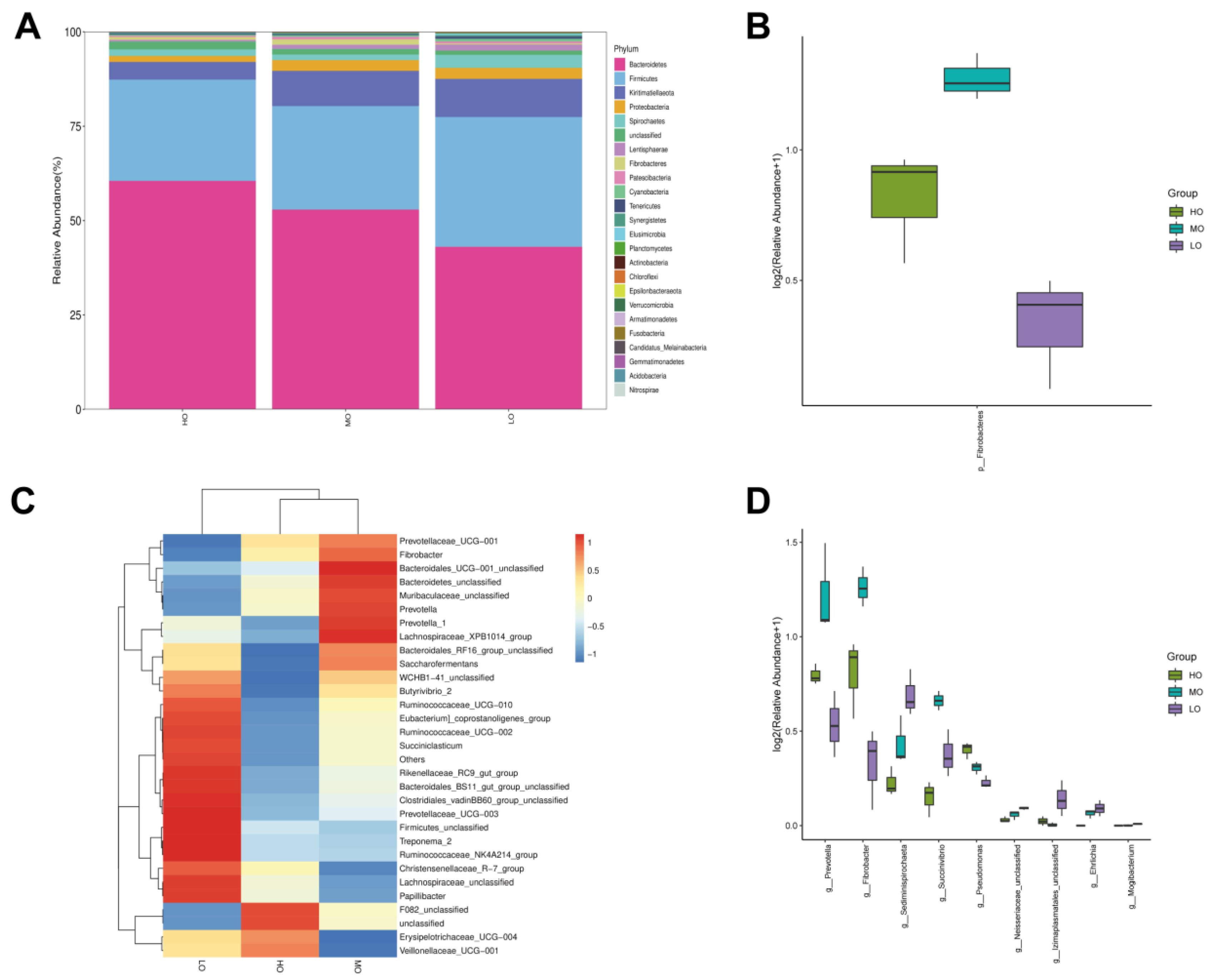
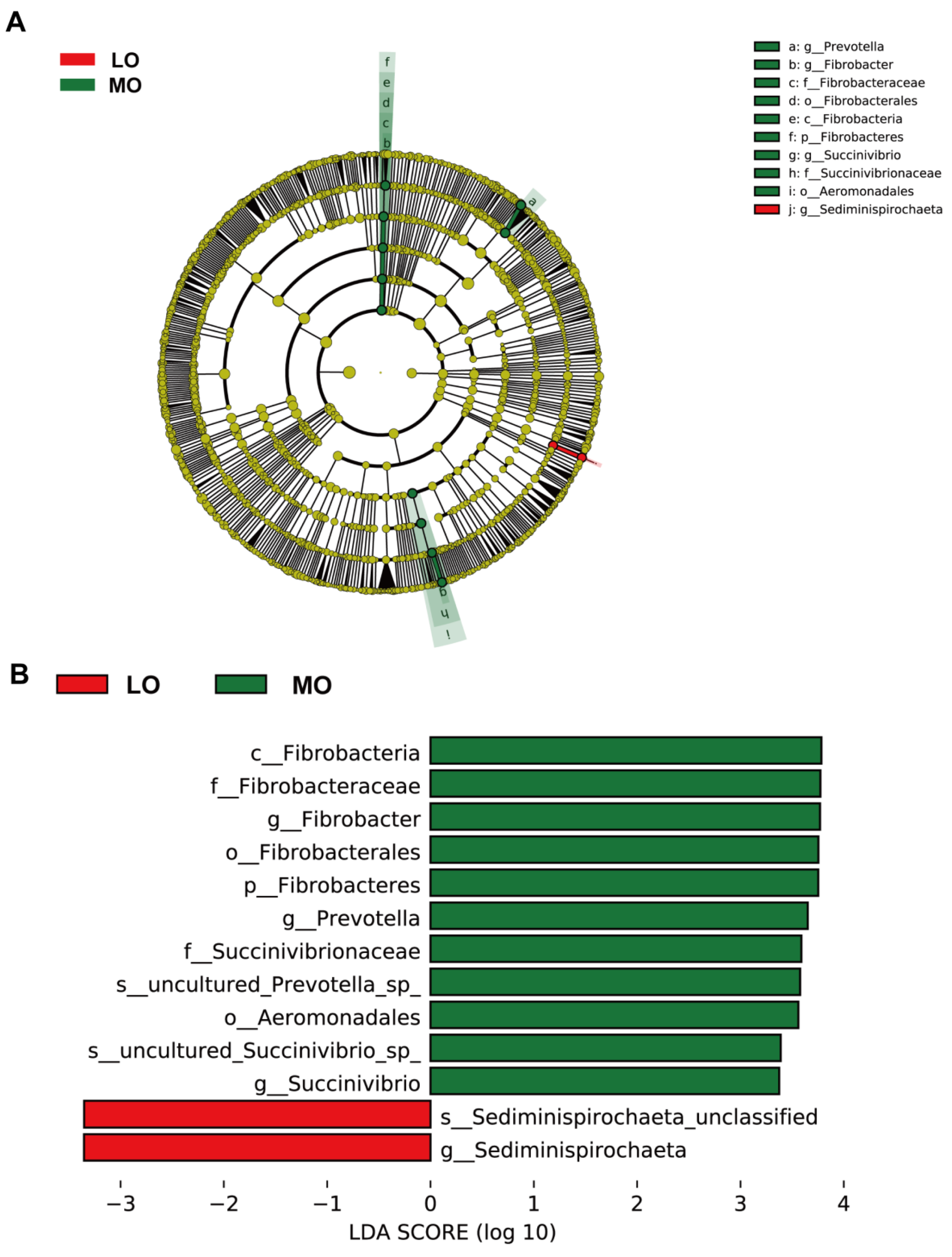
3.2. Rumen Bacterial Communities
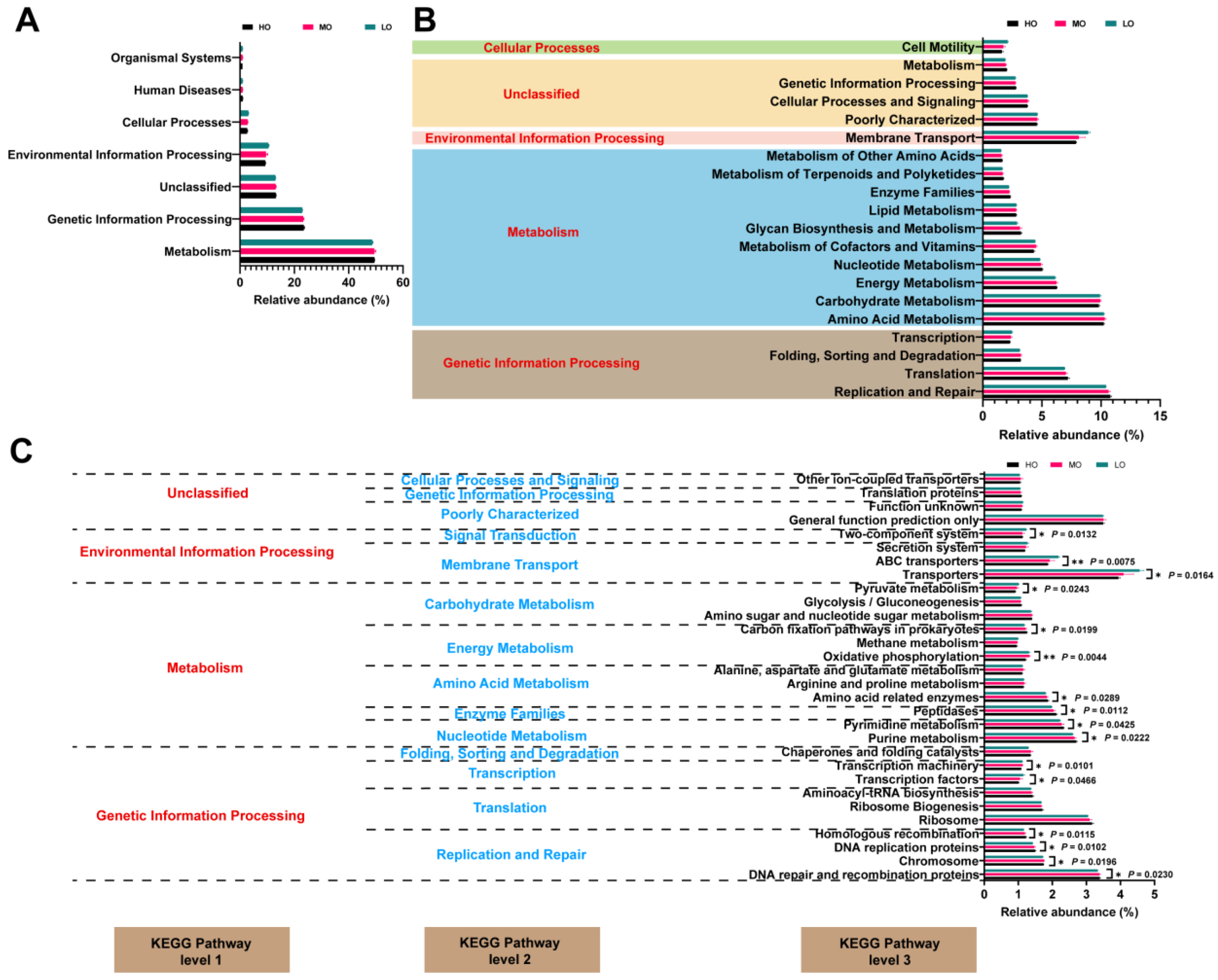
3.3. Predicted Metabolic Pathways and Functions of Rumen Bacterial Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, X.; Ma, Y.; Khan, M.Z.; Xiao, J.; Alugongo, G.M.; Li, S.; Wang, Y.; Cao, Z. A Combination of Lactic Acid Bacteria and Molasses Improves Fermentation Quality, Chemical Composition, Physicochemical Structure, In Vitro Degradability and Rumen Microbiota Colonization of Rice Straw. Front. Vet. Sci. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhang, Y.; Yu, Z.; Xu, Q.; Zheng, N.; Zhao, S.; Huang, G.; Wang, J. Ruminal microbiota–host interaction and its effect on nutrient metabolism. Anim. Nutr. 2021, 7, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, M.; Wang, X.; Yu, Z.; Na, R. Ensiling alfalfa with whole crop corn improves the silage quality and in vitro digestibility of the silage mixtures. Grassl. Sci. 2017, 63, 211–217. [Google Scholar] [CrossRef]
- Chávez, C.C.M.; Ramírez, P.N.; Hoyos, L.R.; Durán, M.G.R.; Madrigal, J.F.; Sánchez, M.C.C.; Cisneros, J.M.A.; Palacios, C.A.M. Retos y perspectivas del sector acuícola rumbo al 2050. Cienc. Nicolaita 2022. [Google Scholar] [CrossRef]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Fermentation quality and microbial community of alfalfa and stylo silage mixed with Moringa oleifera leaves. Bioresour. Technol. 2019, 284, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yin, Q.; Gentu, G.; Xue, Y.; Hou, M.; Liu, L.; Jia, Y. Feeding forage mixtures of alfalfa hay and maize stover optimizes growth performance and carcass characteristics of lambs. Anim. Sci. J. 2017, 89, 359–366. [Google Scholar] [CrossRef]
- Wang, M.; Franco, M.; Cai, Y.; Yu, Z. Dynamics of fermentation profile and bacterial community of silage prepared with alfalfa, whole-plant corn and their mixture. Anim. Feed Sci. Technol. 2020, 270. [Google Scholar] [CrossRef]
- Doran, M.; Laca, E.; Sainz, R. Total tract and rumen digestibility of mulberry foliage (Morus alba), alfalfa hay and oat hay in sheep. Anim. Feed Sci. Technol. 2007, 138, 239–253. [Google Scholar] [CrossRef]
- Du, W.; Hou, F.; Tsunekawa, A.; Kobayashi, N.; Peng, F.; Ichinohe, T. Effects of oat hay and leguminous forage mixture feeding on enteric methane emission, energy utilization, and feed conversion efficiency in male crossbred Simmental beef cattle. Anim. Sci. J. 2020, 91, e13472. [Google Scholar] [CrossRef]
- Zou, Y.; Zou, X.; Li, X.; Guo, G.; Ji, P.; Wang, Y.; Li, S.; Wang, Y.; Cao, Z. Substituting oat hay or maize silage for portion of alfalfa hay affects growth performance, ruminal fermentation, and nutrient digestibility of weaned calves. Asian-Australas. J. Anim. Sci. 2018, 31, 369–378. [Google Scholar] [CrossRef]
- Horcada, A.; Polvillo, O.; Juárez, M.; Avilés, C.; Martínez, A.; Peña, F. Influence of feeding system (concentrate and total mixed ration) on fatty acid profiles of beef from three lean cattle breeds. J. Food Compos. Anal. 2016, 49, 110–116. [Google Scholar] [CrossRef]
- McAuliffe, S.; Mee, J.F.; Lewis, E.; Galvin, N.; Hennessy, D. Feeding System Effects on Dairy Cow Rumen Function and Milk Production. Animals 2022, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, C.; Armentano, L. Short Communication: Feed Selection by Dairy Cows Fed Individually in a Tie-Stall or as a Group in a Free-Stall Barn. J. Dairy Sci. 2007, 90, 2386–2389. [Google Scholar] [CrossRef] [Green Version]
- Hao, W.; Wang, H.L.; Ning, T.T.; Yang, F.Y.; Xu, C.C. Aerobic Stability and Effects of Yeasts during Deterioration of Non-Fermented and Fermented Total Mixed Ration with Different Moisture Levels. Asian-Australas. J. Anim. Sci. 2015, 28, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Zheng, M.; Jiang, D.; Tian, P.; Zheng, M.; Xu, C. Replacing Alfalfa with Paper Mulberry in Total Mixed Ration Silages: Effects on Ensiling Characteristics, Protein Degradation, and In Vitro Digestibility. Animals 2021, 11, 1273. [Google Scholar] [CrossRef]
- Chaosap, C.; Lukkananukool, A.; Polyorach, S.; Sommart, K.; Sivapirunthep, P.; Limsupavanich, R. Effects of Dietary Energy Density in a Fermented Total Mixed Ration Formulated with Different Ratios of Rice Straw and Cassava Pulp on 2- or 14-Day-Aged Meat Quality, Collagen, Fatty Acids, and Ribonucleotides of Native Thai Cattle Longissimus Muscle. Foods 2022, 11, 2046. [Google Scholar] [CrossRef]
- Li, Y.; Lv, J.; Wang, J.; Zhou, S.; Zhang, G.; Wei, B.; Sun, Y.; Lan, Y.; Dou, X.; Zhang, Y. Changes in Carbohydrate Composition in Fermented Total Mixed Ration and Its Effects on In Vitro Methane Production and Microbiome. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Meenongyai, W.; Pattarajinda, V.; Stelzleni, A.M.; Sethakul, J.; Duangjinda, M. Effects of forage ensiling and ration fermentation on total mixed ration pH, ruminal fermentation and performance of growing Holstein-Zebu cross steers. Anim. Sci. J. 2017, 88, 1372–1379. [Google Scholar] [CrossRef]
- Zhang, G.; Li, Y.; Fang, X.; Cai, Y.; Zhang, Y. Lactation performance, nitrogen utilization, and profitability in dairy cows fed fermented total mixed ration containing wet corn gluten feed and corn stover in combination replacing a portion of alfalfa hay. Anim. Feed Sci. Technol. 2020, 269. [Google Scholar] [CrossRef]
- Trabi, E.B.; Seddik, H.-E.; Xie, F.; Lin, L.; Mao, S. Comparison of the rumen bacterial community, rumen fermentation and growth performance of fattening lambs fed low-grain, pelleted or non-pelleted high grain total mixed ration. Anim. Feed Sci. Technol. 2019, 253, 1–12. [Google Scholar] [CrossRef]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Zened, A.; Combes, S.; Cauquil, L.; Mariette, J.; Klopp, C.; Bouchez, O.; Troegeler-Meynadier, A.; Enjalbert, F. Microbial ecology of the rumen evaluated by 454 GS FLX pyrosequencing is affected by starch and oil supplementation of diets. FEMS Microbiol. Ecol. 2013, 83, 504–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Guo, H.M.; Tang, Z.H.; Cao, K.; Wang, Y.P.; Ye, J.A. Effects of pelletedrape straw feed on productive performance, rumen fermentation parameters, and blood biochemical indexes of Hu sheep. Acta Pratacult. Sin. 2016, 25, 171–179. [Google Scholar] [CrossRef]
- Liu, C.; Wu, H.; Liu, S.; Chai, S.; Meng, Q.; Zhou, Z. Dynamic Alterations in Yak Rumen Bacteria Community and Metabolome Characteristics in Response to Feed Type. Front. Microbiol. 2019, 10, 1116. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Liu, B.; Xiao, J.; Guo, M.; Zhao, S.; Hu, M.; Cui, Y.; Li, D.; Wang, C.; Ma, S.; et al. Effects of Different Roughage Diets on Fattening Performance, Meat Quality, Fatty Acid Composition, and Rumen Microbe in Steers. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef]
- AOAC. Oficial Methods of Analysis; Association of Oficial Analytical Chemists: Arlington, TX, USA, 1995. [Google Scholar]
- Murphy, R.P. A method for the extraction of plant samples and the determination of total soluble carbohydrates. J. Sci. Food Agric. 1958, 9, 714–717. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Freer, M.; Dove, H.; Nolan, J.V. Nutrient Requirements of Domesticated Ruminants; CSIRO Publishing: Melbourne, Australia, 2007. [Google Scholar]
- Guo, X.; Zheng, P.; Zou, X.; Chen, X.; Zhang, Q. Influence of Pyroligneous Acid on Fermentation Parameters, CO2 Production and Bacterial Communities of Rice Straw and Stylo Silage. Front. Microbiol. 2021, 12, 701434. [Google Scholar] [CrossRef]
- Hosoda, K.; Matsuyama, H.; Park, W.-Y.; Nishida, T.; Ishida, M. Supplementary effect of peppermint (Mentha × piperita) on dry matter intake, digestibility, ruminal fermentation and milk production in early lactating dairy cows. Anim. Sci. J. 2006, 77, 503–509. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated Simultaneous Determination of Ammonia and Total Amino Acids in Ruminal Fluid and In Vitro Media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Ma, J.; Shah, A.M.; Shao, Y.; Wang, Z.; Zou, H.; Hu, R.; Peng, Q.; Kang, K.; Wanapat, M. Effects of yeast cell wall on the growth performance, ruminal fermentation, and microbial community of weaned calves. Livest. Sci. 2020, 239, 104170. [Google Scholar] [CrossRef]
- Logue, J.B.; Stedmon, C.; Kellerman, A.M.; Nielsen, N.J.; Andersson, A.F.; Laudon, H.; Lindström, E.; Kritzberg, E.S. Experimental insights into the importance of aquatic bacterial community composition to the degradation of dissolved organic matter. ISME J. 2015, 10, 533–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 2016, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Zhang, T.; Tian, Q.; Cheng, Y.; Gebeyew, K.; Liu, G.; Tan, Z.; He, Z. Supplementing Mannan Oligosaccharide Reduces the Passive Transfer of Immunoglobulin G and Improves Antioxidative Capacity, Immunity, and Intestinal Microbiota in Neonatal Goats. Front. Microbiol. 2022, 12, 795081. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Su, X.; Li, J.; Yang, Y.; Wang, P.; Yan, F.; Yao, J.; Wu, S. Real-time monitoring of ruminal microbiota reveals their roles in dairy goats during subacute ruminal acidosis. Npj Biofilm. Microbiomes 2021, 7, 45. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, Y.; Wu, L.; Xun, W.; Liu, Q.; Cao, T.; Hou, G.; Zhou, H. Moderate Coconut Oil Supplement Ameliorates Growth Performance and Ruminal Fermentation in Hainan Black Goat Kids. Front. Vet. Sci. 2020, 7, 622259. [Google Scholar] [CrossRef]
- Kim, T.-I.; Mayakrishnan, V.; Lim, D.-H.; Yeon, J.-H.; Baek, K.-S. Effect of fermented total mixed rations on the growth performance, carcass and meat quality characteristics of Hanwoo steers. Anim. Sci. J. 2017, 89, 606–615. [Google Scholar] [CrossRef]
- Kim, D.; Jung, J.-S.; Choi, K.-C. A Preliminary Study on Effects of Fermented Feed Supplementation on Growth Performance, Carcass Characteristics, and Meat Quality of Hanwoo Steers during the Early and Late Fattening Period. Appl. Sci. 2021, 11, 5202. [Google Scholar] [CrossRef]
- Huhtanen, P.; Khalili, H.; Nousiainen, J.; Rinne, M.; Jaakkola, S.; Heikkilä, T. Prediction of the relative intake potential of grass silage by dairy cows. Livest. Prod. Sci. 2002, 73, 111–130. [Google Scholar] [CrossRef]
- Torres-Pitarch, A.; Gardiner, G.E.; Cormican, P.; Rea, M.; Crispie, F.; O’Doherty, J.V.; Cozannet, P.; Ryan, T.; Cullen, J.; Lawlor, P.G. Effect of cereal fermentation and carbohydrase supplementation on growth, nutrient digestibility and intestinal microbiota in liquid-fed grow-finishing pigs. Sci. Rep. 2020, 10, 13716. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.M.; Chang, S.S.; Cho, Y.; Kim, H.C.; Kwon, E.G.; Yang, S.H.; Paek, B.H. Effects of Forage Source and Shipping Time on Growth Performance and Carcass Characteristics of Hanwoo Steers. J. Korean Soc. Grassl. Forage Sci. 2009, 29, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Mushi, D.; Safari, J.; Mtenga, L.; Kifaro, G.; Eik, L. Effects of concentrate levels on fattening performance, carcass and meat quality attributes of Small East African×Norwegian crossbred goats fed low quality grass hay. Livest. Sci. 2009, 124, 148–155. [Google Scholar] [CrossRef]
- Wang, X.; Tsai, T.; Wei, X.; Zuo, B.; Davis, E.; Rehberger, T.; Hernandez, S.; Jochems, E.; Maxwell, C.; Zhao, J. Effect of Lactylate and Bacillus subtilis on Growth Performance, Peripheral Blood Cell Profile, and Gut Microbiota of Nursery Pigs. Microorganisms 2021, 9, 803. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Qi, M.; Wang, S.; Diao, Q.; Zhang, N. Dietary energy and protein levels influenced the growth performance, ruminal morphology and fermentation and microbial diversity of lambs. Sci. Rep. 2019, 9, 16612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Huang, X.; Zhang, Z.; Duan, Z. Effect of Caragana korshinskii Kom. as a partial substitution for sheep forage on intake, digestibility, growth, carcass features, and the rumen bacterial community. Trop. Anim. Heal. Prod. 2022, 54, 190. [Google Scholar] [CrossRef]
- Yang, B.; Le, J.; Wu, P.; Liu, J.; Guan, L.L.; Wang, J. Alfalfa Intervention Alters Rumen Microbial Community Development in Hu Lambs During Early Life. Front. Microbiol. 2018, 9, 574. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Sun, X.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Effects of Herbal Tea Residue on Growth Performance, Meat Quality, Muscle Metabolome, and Rumen Microbiota Characteristics in Finishing Steers. Front. Microbiol. 2022, 12, 821293. [Google Scholar] [CrossRef]
- Zebeli, Q.; Tafaj, M.; Weber, I.; Steingass, H.; Drochner, W. Effects of dietary forage particle size and concentrate level on fermentation profile, in vitro degradation characteristics and concentration of liquid- or solid-associated bacterial mass in the rumen of dairy cows. Anim. Feed Sci. Technol. 2008, 140, 307–325. [Google Scholar] [CrossRef]
- Wang, J.; Cheng, L.; Chaudhry, A.S.; Khanaki, H.; Abbasi, I.H.R.; Ma, Y.; Abbasi, F.; Guo, X.; Zhang, S. Silage Mixtures of Alfalfa with Sweet Sorghum Alter Blood and Rumen Physiological Status and Rumen Microbiota of Karakul Lambs. Animals 2022, 12, 2591. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, H.; Gao, Z.; Xu, J.; Liu, B.; Guo, M.; Yang, X.; Niu, J.; Zhu, X.; Ma, S.; et al. Whole-plant corn silage improves rumen fermentation and growth performance of beef cattle by altering rumen microbiota. Appl. Microbiol. Biotechnol. 2022, 106, 4187–4198. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; You, S.; Sun, L.; Wang, X.; Jia, Y.; Zhou, Y. Effects of Replacing Alfalfa Hay With Native Grass Hay in Pelleted Total Mixed Ration on Physicochemical Parameters, Fatty Acid Profile, and Rumen Microbiota in Lamb. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Chai, J.; Lv, X.; Diao, Q.; Usdrowski, H.; Zhuang, Y.; Huang, W.; Cui, K.; Zhang, N. Solid diet manipulates rumen epithelial microbiota and its interactions with host transcriptomic in young ruminants. Environ. Microbiol. 2021, 23, 6557–6568. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census Collaborators; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Li, Z.; Yu, Z.; Zhu, W. Effects of dietary replacement of soybean meal with dried distillers grains with solubles on the microbiota occupying different ecological niches in the rumen of growing Hu lambs. J. Anim. Sci. Biotechnol. 2020, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Oskoueian, E.; Jahromi, M.; Jafari, S.; Shakeri, M.; Le, H.; Ebrahimi, M. Manipulation of Rice Straw Silage Fermentation with Different Types of Lactic Acid Bacteria Inoculant Affects Rumen Microbial Fermentation Characteristics and Methane Production. Vet. Sci. 2021, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Júnior, R.T.A.; De Souza, E.J.O.; De Melo, A.A.S.; Silva, D.K.D.A.; Torres, T.R.; Perreira, G.F.C. Mesquite Extract as Phytogenic Additive to Improve the Nutrition of Sheep. J. Agric. Sci. 2017, 9, 164. [Google Scholar] [CrossRef] [Green Version]
- Eun, J.-S.; Beauchemin, K. Assessment of the efficacy of varying experimental exogenous fibrolytic enzymes using in vitro fermentation characteristics. Anim. Feed Sci. Technol. 2007, 132, 298–315. [Google Scholar] [CrossRef]
- Zhao, Y.; Xie, B.; Gao, J.; Zhao, G. Dietary Supplementation with Sodium Sulfate Improves Rumen Fermentation, Fiber Digestibility, and the Plasma Metabolome through Modulation of Rumen Bacterial Communities in Steers. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Ellison, M.; Conant, G.; Lamberson, W.; Cockrum, R.; Austin, K.; Rule, D.; Cammack, K. Diet and feed efficiency status affect rumen microbial profiles of sheep. Small Rumin. Res. 2017, 156, 12–19. [Google Scholar] [CrossRef]
- Huang, J.; Wu, T.; Sun, X.; Zou, C.; Yang, Y.; Cao, Y.; Yang, Y.; Iqbal, M.W.; Lin, B. Effect of replacing conventional feeds with tropical agricultural by-products on the growth performance, nutrient digestibility and ruminal microbiota of water buffaloes. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Ran, T.; Tang, S.; Yu, X.; Hou, Z.; Hou, F.; Beauchemin, K.; Yang, W.; Wu, D. Diets varying in ratio of sweet sorghum silage to corn silage for lactating dairy cows: Feed intake, milk production, blood biochemistry, ruminal fermentation, and ruminal microbial community. J. Dairy Sci. 2021, 104, 12600–12615. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.; Lelis, A.; Fan, P.; Agustinho, B.C.; Lobo, R.; Arce-Cordero, J.; Dai, X.; Jeong, K.; Faciola, A. Effects of lactic acid-producing bacteria as direct-fed microbials on the ruminal microbiome. J. Dairy Sci. 2022, 105, 2242–2255. [Google Scholar] [CrossRef]
- Wang, S.R.; Guo, G.; Li, J.F.; Chen, L.; Dong, Z.H.; Shao, T. Improvement of fermentation profile and structural carbohydrate compositions in mixed silages ensiled with fibrolytic enzymes, molasses and Lactobacillus plantarum MTD-1. Ital. J. Anim. Sci. 2018, 18, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Mickdam, E.; Khiaosa-Ard, R.; Metzler-Zebeli, B.; Humer, E.; Harder, H.; Khol-Parisini, A.; Zebeli, Q. Modulation of ruminal fermentation profile and microbial abundance in cows fed diets treated with lactic acid, without or with inorganic phosphorus supplementation. Anim. Feed Sci. Technol. 2017, 230, 1–12. [Google Scholar] [CrossRef]
- Ren, Z.; Yao, R.; Liu, Q.; Deng, Y.; Shen, L.; Deng, H.; Zuo, Z.; Wang, Y.; Deng, J.; Cui, H.; et al. Effects of antibacterial peptides on rumen fermentation function and rumen microorganisms in goats. PLoS ONE 2019, 14, e0221815. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zeng, X.; Zeng, Y.; Liu, X.; Wang, Y.; Wang, X.; Li, J.; Wang, Y.; Feng, Z.; Huang, P.; et al. Effects of Phragmites australis Shoot Remainder Silage on Growth Performance, Blood Biochemical Parameters, and Rumen Microbiota of Beef Cattle. Front. Vet. Sci. 2022, 9, 778654. [Google Scholar] [CrossRef]
- Shivani, Y.; Subhash, Y.; Sasikala, C.; Ramana, C.V. Characterisation of a newly isolated member of a candidatus lineage, Marispirochaeta aestuarii gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 3929–3936. [Google Scholar] [CrossRef]
- Borhami, B.E.A.; El-Shazly, K.; Akkada, A.R.A. Effect of ruminal infusion of acetic acid and sodium acetate on the concentrations of ciliate protozoa. J. Agric. Sci. 1972, 78, 239–244. [Google Scholar] [CrossRef]
- Kim, H.; Park, T.; Kwon, I.; Seo, J. Specific inhibition of Streptococcus bovis by endolysin LyJH307 supplementation shifts the rumen microbiota and metabolic pathways related to carbohydrate metabolism. J. Anim. Sci. Biotechnol. 2021, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A. Altering ruminal nitrogen metabolism to improve protein utilization. Introduction. J. Nutr. 1996, 126, 1324S–1325S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramšak, A.; Peterka, M.; Tajima, K.; Martin, J.C.; Wood, J.; Johnston, M.E.A.; Aminov, R.I.; Flint, H.J.; Avguštin, G. Unravelling the genetic diversity of ruminal bacteria belonging to the CFB phylum. FEMS Microbiol. Ecol. 2000, 33, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.; Ilina, L.; Laptev, G.; Filippova, V.; Brazhnik, E.; Dunyashev, T.; Dubrovin, A.; Novikova, N.; Tiurina, D.; Tarlavin, N.; et al. The structure and functional profile of ruminal microbiota in young and adult reindeers (Rangifer tarandus) consuming natural winter-spring and summer-autumn seasonal diets. PeerJ 2021, 9, e12389. [Google Scholar] [CrossRef]
- Wang, Z.; He, Y.; Wang, C.; Ao, H.; Tan, Z.; Xing, K. Variations in Microbial Diversity and Metabolite Profiles of Female Landrace Finishing Pigs With Distinct Feed Efficiency. Front. Vet. Sci. 2021, 8, 702931. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | LO | MO | HO |
|---|---|---|---|
| Ingredient (g/kg DM) | |||
| Oat hay | 200 | 300 | 400 |
| Alfalfa hay | 400 | 300 | 200 |
| Natural forage | 30 | 30 | 30 |
| Corn stalk | 20 | 20 | 20 |
| Corn | 220 | 200 | 180 |
| Soybean meal | 90 | 110 | 130 |
| Wheat bran | 20 | 20 | 20 |
| Calcium hydrogen phosphate | 3 | 3 | 3 |
| NaCl | 2 | 2 | 2 |
| NaHCO3 | 5 | 5 | 5 |
| Premix | 10 | 10 | 10 |
| Chemical compositions | |||
| DM (g/kg FW) | 45.73 ± 0.46 | 45.90 ± 0.40 | 45.77 ± 0.49 |
| CP (g/kg DM) | 13.33 ± 0.12 | 13.28 ± 0.24 | 13.12 ± 0.17 |
| NDF (g/kg DM) | 43.20 ± 0.02 | 50.29 ± 0.01 | 55.30 ± 0.01 |
| ADF (g/kg DM) | 29.53 ± 0.02 | 31.83 ± 0.01 | 33.77 ± 0.01 |
| WSC (g/kg DM) | 4.66 ± 0.50 | 4.91 ± 0.52 | 4.37 ± 0.73 |
| OM (g/kg DM) | 34.13 ± 0.31 | 34.65 ± 0.46 | 33.93 ± 0.53 |
| ME | 10.80 ± 0.01 | 10.70 ± 0.01 | 10.59 ± 0.01 |
| Fermentation profile | |||
| pH | 4.55 ± 0.06 | 4.52 ± 0.04 | 4.49 ± 0.02 |
| Lactic acid (g/kg DM) | 10.28 ± 0.08 | 9.86 ± 1.29 | 9.69 ± 0.93 |
| Acetic acid (g/kg DM) | 1.15 ± 0.01 | 0.88 ± 0.14 | 0.76 ± 0.18 |
| Propionic acid (g/kg DM) | 1.24 ± 0.01 | 0.63 ± 0.20 | 0.94 ± 0.14 |
| Ammonia-N (g/kg DM) | 3.03 ± 0.01 | 2.65 ± 0.16 | 4.39 ± 0.03 |
| Microbial counts | |||
| Lactic acid bacteria (Log10 cfu/g FM) | 5.90 ± 0.04 | 7.81 ± 0.01 | 5.91 ± 0.26 |
| Aerobic bacteria (Log10 cfu/g FM) | 6.05 ± 0.18 | 4.48 ± 0.13 | 6.74 ± 0.50 |
| Items | LO | MO | HO | SEM | p-Value |
|---|---|---|---|---|---|
| Initial BW (kg) | 28.00 | 29.33 | 29.17 | 0.333 | 0.2205 |
| Final BW (kg) | 42.83 | 45.50 | 42.17 | 0.764 | 0.1715 |
| TWG (kg) | 14.83 ab | 16.17 a | 13.00 b | 0.601 | 0.0423 |
| ADG (g/day) | 247.22 ab | 269.44 a | 216.67 b | 10.015 | 0.0423 |
| DMI (kg/day) | 1.17 b | 1.18 a | 1.13 c | 0.008 | <0.0001 |
| Items | LO | MO | HO | SEM | p-Value |
|---|---|---|---|---|---|
| OTUs | 2446 | 2276 | 2627 | 110.723 | 0.5390 |
| Chao1 | 2448.61 | 2282.45 | 2633.22 | 110.220 | 0.5359 |
| Simpson | 1.00 | 0.99 | 0.97 | 0.007 | 0.1790 |
| Shannon | 9.48 | 8.98 | 8.68 | 0.192 | 0.3059 |
| Goods’ coverage | 99.88 | 99.92 | 99.93 | 0.001 | 0.5582 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Wang, Y.; Wang, Z.; Ge, G.; Jia, Y.; Du, S. Effects of Replacing Alfalfa Hay with Oat Hay in Fermented Total Mixed Ration on Growth Performance and Rumen Microbiota in Lambs. Fermentation 2023, 9, 9. https://doi.org/10.3390/fermentation9010009
Liu M, Wang Y, Wang Z, Ge G, Jia Y, Du S. Effects of Replacing Alfalfa Hay with Oat Hay in Fermented Total Mixed Ration on Growth Performance and Rumen Microbiota in Lambs. Fermentation. 2023; 9(1):9. https://doi.org/10.3390/fermentation9010009
Chicago/Turabian StyleLiu, Mingjian, Yu Wang, Zhijun Wang, Gentu Ge, Yushan Jia, and Shuai Du. 2023. "Effects of Replacing Alfalfa Hay with Oat Hay in Fermented Total Mixed Ration on Growth Performance and Rumen Microbiota in Lambs" Fermentation 9, no. 1: 9. https://doi.org/10.3390/fermentation9010009