
Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications


1
Engineering Research Center of Agricultural Microbiology Technology, Ministry of Education & Heilongjiang Provincial Key Laboratory of Plant Genetic Engineering and Biological Fermentation Engineering for Cold Region & Key Laboratory of Microbiology, College of Heilongjiang Province & School of Life Sciences, Heilongjiang University, Harbin 150080, China
2
State Key Laboratory of Microbial Metabolism, School of Life Sciences and Biotechnology, Shanghai Jiao Tong University, Shanghai 200240, China
*
Authors to whom correspondence should be addressed.
Fermentation 2022, 8(11), 629; https://doi.org/10.3390/fermentation8110629
Submission received: 16 September 2022
/
Revised: 7 November 2022
/
Accepted: 9 November 2022
/
Published: 11 November 2022
(This article belongs to the Section Microbial Metabolism, Physiology & Genetics)
Abstract
:Glucansucrase (GS) belongs to the GH70 family, which not only can synthesize exopolysaccharides (EPSs) with different physicochemical properties through glucosyl transglycosylation (by hydrolyzing sucrose) but can also produce oligosaccharides. Different strains produce different GSs, which catalyze the synthesis of EPS with different glycosidic bond structures; these EPSs have different biological functions. As an important enzymatic tool, GS has great potential in health care medicine, biological materials, ecological protection, the food chemical industry, etc. GS is mainly produced by lactic acid bacteria (LAB), including Leuconostoc, Streptococcus, Lactobacillus, and Weissella species. With the elucidation of the crystal structure of GS and the advancement of genome sequencing technology, its synthesis reaction mechanism and specific structural characteristics are gradually becoming clear. This review summarizes the isolation, purification, physical and chemical properties, detection methods, sources, and applications of GS in order to provide a reference for the research and development of GS.
1. Introduction
Polysaccharide polymers are one of the most valuable natural substances, and they play an important role in the fields of medical devices, food chemicals, and ecological and environmental protection [1]. Among them, the production of microbial exopolysaccharide (EPS) has the characteristics of being unaffected by environmental factors and easier downstream processing compared with plants, algae, fungi, and petroleum-based EPSs, which makes it shine in the field of production [2]. Bacterial enzymes involved in the production of EPS from sucrose, termed glucansucrases (GSs), are extracellular enzymes that can be assisted by branched sucrases (BRSs). GSs belong to the glycoside hydrolase GH70, which are very efficient transglycosylases and do not require expensive nucleotide-activated sugars (NDP-sugars) [3,4,5].
GSs are additionally named glucosyltransferases (GTFs), which catalyze the synthesis of EPSs and oligosaccharides using the glucose unit from the sucrose donor. GSs are applied in the fields of feed, food, medicine, and other engineering fields because of their outstanding physicochemical properties [6,7]. Bacterial GSs are mainly produced by lactic acid bacteria (LAB), a group of Gram-positive bacteria like Leuconostoc, Streptococcus, Lactobacillus, and Weissella species [8,9,10]. According to the glycosidic linkages present in the polymer, these enzymes are classified as (i) the synthesis of dextran by dextransucrases, consisting mainly of α-1,6 linkage and some α-1,2, α-1,3 and α-1,6 branches, (ii) the synthesis of mutan α-1,3, by mutansucrases (iii) forming alternan via alternansucrase, composed of alternating α-1,6 and α-1,3 glucosidic linkages, and (iv) reuteransucrase synthesizing reuteran containing α-1,4 and α-1,6 linkages [5]. These enzymes additionally catalyze the formation of oligosaccharide and glycoconjugate products in the presence of sucrose and nonsucrose receptor substrates [9,11,12].
EPSs and oligosaccharides are being extensively studied for their prospects toward probiotics by stimulating the growth of probiotic strains or beneficial endogenous strains from the gastrointestinal tract [13,14,15]. Although these investigations have touched on a few imperative issues, such as the approximate structure of GS, uncovering the relationship between the structure and activity of GS and its component of activity are still issues to be unraveled.
2. Structure of the GS
Sequence similarity analysis showed that GSs belonged to the GH70 family of glycoside hydrolases, with an average molecular weight of about 160 kDa and an optimum temperature and pH of 30 °C and 5.0–6.0, respectively [16,17]. GSs have been found from Lactobacillus kunkeei H3 and H25 have a mass of 300 kDa [18]. Due to the relatively large molecular weight of enzymes, and structural analysis being very difficult, only the crystal structures of four GSs have been resolved. However, with the advent of the cryoelectron microscopy technology revolution, it is possible to resolve protein structure using a high resolution, which will help to promote the 3D structure exploration and characterization of GS.
Amino acid sequence analysis showed that the GSs from LAB share a frequent structure and are composed of four different domains: (i) a sign peptide, followed by using (ii) an extraordinarily variable stretch, (iii) a quite conserved catalytic or sucrose-binding domain and (iv) a C-terminal glucan binding domain composed of a series of tandem repeats [16,19]. Currently, crystallographic analysis of the 3D structure of Lactobacillus GSs showed that they contain a common domain organization. A truncated enzyme was used for crystallization [5]. With the elucidation of the 3D structure of GTF180-∆N, the hypothesis of the ring arrangement was confirmed [20,21]. Different from the previous prediction of the primary structure of the enzyme, the three-dimensional structure of the truncated GSs formed five domains (domain A, domain B, domain C, domain IV, and domain V) and are arranged as C-A-B-IV-V lines (Figure 1). Except for domain C, these four domains are composed of two discontinuous polypeptide chains at the N-terminal and C-terminal ends. Among them, A, B, and C are catalytic cores, while IV and V are unique to GH70 GS [5]. The active site of the GSs is located at the interface of domain A and domain B [22], providing residues for the active site gap and their possible role in determining the ligation specificity of the product is discussed.
Domain A consists of (β/α)8 barrels and contains a catalytic site with the catalytic residue at the bottom of the domain. A crystal structure analysis of the GTF180-N mutant D1025N bound to sucrose [23,24] confirmed that these three residues constitute the catalytic site of GS. Domain B can form a Ca2+ binding site in domain A, which are used to form the binding site of substrate and receptor [5]. Domain C is located at the U-shaped bottom end of GS. It is the only domain in GH70 GS enzymes that is formed by a continuous polypeptide segment, but its function is still unclear. IV and V are additional domains; the structure of domain IV is different from that of any other known protein and only occurs in GH70 enzymes (Figure 1) [5,24].
Domain V is adjacent to domain IV. It contains several repeats and has been shown to be involved in glucan binding [13] and is a glucan-binding domain that is involved, to a certain extent, in glucan extension and enzymatic processing, providing a high-affinity anchoring platform for the synthesis of high-molecular-weight dextran [25,26]. There may well be subtle interactions between the V domain and the catalytic domain. It can help capture the polymer chain and keep it near the active site to facilitate the extension or branching of the sugar chain. Partial or total truncation of domain V affects the binding capacity of glucan and also changes the size of the synthesized polymer. Structural analysis of domain V revealed the presence of a consensus β-solenoid fold with multiple copies [27]. In the crystal structure, it has a variety of conformations, mainly divided into two types, one is the extension to the active catalytic center, and the other is the extension to the outside of the conformation in Leuconostoc mesenteroides NRRL B-1355-alternating α-1,3/1,6-glucosyltransferase (Asr). Its domain V was found to extend to the catalytic core in Ln. reuteri GTF-180N. Its domain V was found to extend out of the conformation (Figure 1) [5,27].
The exact role of domain IV is unclear. It is speculated that domain IV acts as a hinge to promote the growth of glucan chains by directing the glucan chains bound to domain V towards or away from the catalytic site. The N-terminus of GS contains a signal peptide (36 to 40 amino acids) for its secretion [5,28]. The amino acid fragment between the signal peptide and the GS core region is highly variable in content and dimension (200 to 700 amino acids) [28]. The function of the N-terminal domain remains unknown.
3. Catalytic Mechanism of GS
Both the catalytic mechanism and structure of GH70 are closely related to the GH13 and GH77 families [24]. The newly discovered GH70 subfamily GtfB, GtfC, and GtfD are inactive against sucrose but can catalyze starch and maltodextrin to α-glucan [24]. The common characteristic of GH family enzymes is that they also use a catalytic (β/α)8 barrel domain to break down the α-glycosidic bonds between glucose and other glucose or fructose [29,30]. Robyt found there are two active sites; according to the GS catalytic reaction, one is composed of covalent β-glucose-enzyme intermediates under oxygen-carbon ion-like transformation conditions [5,31]. The β-glucose-enzyme intermediate is catalyzed through the a-retaining double displacement reaction, which involves three important residues: a nucleophile, an acid-base catalyst, and a transition state stabilizer (Figure 2). Aspartic acid, which first acts as a nucleophile, attacks the ectopic C1 carbon of the sucrose–glucose unit. Glutamate acts as an acid-base catalyst to transfer protons to fructose and release fructose. The transition state stabilizes the stable residue dimension and transitions to a covalent β-glucose-enzyme. Finally, the covalent β-glucosyl-enzyme is formed from the transition state stabilizer (Figure 2). The other is composed of chain and enzyme intermediates [30]. The C1 position in the later intermediate attacks the C6 position of the glucosyl group to form a glycosidic bond, thereby increasing the length [31]. Although the determinants of the size distribution of GS products have been broadly studied before, many are still unknown [26,32,33,34,35,36]. The N-terminal variable region and C-terminal glucan-binding domain have been indicated as playing a role in product size distribution [26].
The catalytic mechanism of GS allows for the hydrolysis of sucrose to obtain a glucosyl-enzyme intermediate, and this mechanism is based on a detailed structural analysis of Bacillus circulans 251 CGTase [37]. Due to the different receptors, GS appears to synthesize different products: (i) through hydrolysis, water acts as a receptor and hydrolyses to glucose; (ii) through transglycosylation, the glucosyl moiety is converted to an accepting sugar after the fructose is discharged [27]. When substrate-only sucrose exists, GS hydrolyses sucrose to dextran; furthermore, due to the different amino acid sequences of the GS active center, the glycosidic bond composition and branching structure of the produced glucan is also different. Most of the synthesized glucans are composed of one or two types of glycosidic bonds, in which the composition ratio, branching degree, and branch length are also random, which depends on the amino acid sequences of the GS active center [38,39].
4. Isolation and Purification of GS
GS is an extracellular enzyme for which its structure and catalytic mechanism have not been clarified. In order to understand the structure and function of GS, it is necessary to isolate and purify GS [17]. Various GS purification methods, including salting-out and solvents [40], phase partition [41,42,43], polyethylene glycol fractioning [41,43,44], chromatography column [45,46,47], ultrafiltration [48], and combined processes such as sugaring and gel permeation chromatography [49,50].
Salting out, normally as ammonium sulfate precipitation, is when the ionic strength in a solution and the solubility of different proteins are different; high concentrations of salt ions compete with proteins for water molecules in protein solutions, therefore destroying the hydrated membrane on the surface of the protein, reducing its solubility, and allowing it to precipitate out of the solution. Due to different protein solubility, different concentrations of salt solution can be used to precipitate different proteins. Normally, people use ammonium sulfate precipitation as the first step for purification because of its high solubility. Robyt [51] thought ammonium sulfate would hurt the enzyme activity of GS as the concentration rose, especially to more than 80%.
At present, a variety of techniques have been successfully used for the isolation and purification of Ln. mesenteroides GS, and ultrafiltration and gel filtration chromatography are considered to be the best way to purify GS due to the resulting high recovery of enzyme activity. Miao [52] used freeze-drying ion-exchange chromatography and gel filtration methods to purify the enzyme. The crude enzyme solution was concentrated by freeze-drying and loaded into a DEAE-Sepharose FF 16/10 anion exchange column. Further purification was performed using Sepharose CL-6B gel filtration chromatography. The GS was purified 8.6-fold, and its specific activity was 1.3 IU/mg. Polyethylene glycol (PEG) is an uncharged linear macromolecular polymer, and its strong dehydration ability can destroy the hydration layer on the surface of protein molecules and cause protein precipitation. This method is cheap and easy to perform but it is easily affected by centrifugation temperature and pH, so it is usually used in combination with other methods. Song [16] precipitated the crude GS with 10% (v/v) PEG 2000 and then loaded it into a HiTrap Q FF anion exchange column and Sepharose CL-6B column. The purified fractions were dialyzed, concentrated, and collected, resulting in a specific activity of 1.4 U/mg protein, with 13.2-fold purification. Nigam et al. [53] compared the phase separation and purification effect of polyethylene glycol PEG 6000 and PEG 400 on dextransucrase, and the results showed that the tertiary phase separation effect of PEG 6000 was better; the final recovery rate was 84%, and the specific activity of the enzyme after purification.
The yield from GS separation by traditional separation technology is low, and its catalytic properties have not been thoroughly analyzed. In recent years, the cloning and expression of GS by means of genetic engineering is expected to overcome the shortcomings of low enzyme yield and expand the industrial application of the enzyme [54,55]. Kim et al. [56]. constructed Escherichia coli BL21 (DE3) carrying the Ln. lactis EG001 GS gene. The crude enzyme was mixed with Ni-NTA agarose, and the mixture was loaded onto a chromatography column. The proteins were eluted, and the specific activity of the purified enzyme showed an increase of over 2.3-fold with respect to the crude enzyme. Amari [54] et al. constructed engineered bacteria that could express the dextransucrase gene of W. confusa C39-2. The recombinant enzyme was purified using an affinity chromatography protocol, and the activity was 4-fold higher. At present, GS derived from a variety of LAB has been isolated and purified, and its structure and properties have been continuously analyzed (Table 1).
5. Physiological and Biochemical Properties of GS
Many factors can affect the catalytic activity of GS, including pH, temperature, and some organic solvents and metal ions. According to Kralj [34], the maximum GS activity from L. reuteri was gained at a pH of 4.0–5.5, which is comparable with most papers. Miao [52] found that GS from L. reuteri SK24.003 retained high activity at a low pH, showing better acid-resistance. However, a pH value that is too high or too low is still not conducive to the synthesis of GS. Previous studies have shown that GS has the highest purity and the most stable activity at pH 7 [95], while a low pH can significantly reduce enzyme synthesis. The analysis of the GS produced by Ln. mesenteroides DRP2-19 and isolated from sauerkraut showed that the optimum pH for GS was 5.56, and the synthesis of GS was severely affected at pH < 4.5 or >7, which corroborated the previous point [96]. Because of the different sources and structures of GSs, the optimum reaction temperature for each GS is different. Generally, the optimum range is from 30–40°C in culture. When the temperature exceeds 45 °C, the enzyme activity begins to decrease, and when it exceeds 50 °C, the enzyme loss is more obvious. Most double-charged ions (Mg2+, Mn2+, Ni2+, Co2+, Ca2+, Fe2+, and Zn2+) activated enzyme activity, suggesting it was a metal-activated enzyme [52]. GS has a calcium ion activation site, which can increase enzyme activity. It is concluded that calcium ion can enhance the activity of dextransucrase. The stability of the entire edifice is enhanced by calcium coordination, which likely reinforces the interaction between the two domains [97]. Qader et al. [98] found that when the concentration of CaCl2 was 0.005%, the enzyme activity increased to 108.26 DSU/mL/h, which was 2.03 times higher than that of the control group. When the content of CaCl2 was higher than 0.005%, enzyme activity decreased gradually. Some reports found that when Ca2+ is within a certain concentration range, Ca2+ will preferentially bind to the activation site on the enzyme, and the activation effect is stronger than the inhibitory effect. Furthermore, Hg+, Zn2+, Cu2+, Pb2+, and Fe3+ had a strong inhibitory effect on enzyme activity, which was the same as in previous reports; when copper ions were present, there was no transferase activity [99]. The activity and stability of GS in the presence of organic solvents were related to the solvent concentration and its nature. Chemical inhibitors indicate that the function of amino acid residues is located at the GS-active site [16]. Most chemical inhibitors had an inhibition effect on GS, like sodium dodecyl sulfate (SDS), ethylene diamine, tetraacetic acid (EDTA), and β-Mercaptoethanol (β-ME) [25,100]. Other chemical reagents, including butanol, n-hexane, chloroform, calcium ammonium nitrate (CAN), and ethyl acetate, also inhibited the activity of GS with increasing concentrations, whereas glycerol, formaldehyde, and dimethyl sulfoxide (DMSO) enhanced the activity of GS to a certain extent [101].
6. EPS Synthesis by GS
Glucose can not only form EPS in a targeted manner but can also increase the production of EPS and reduce the production of other products, which has become a research focus in the food industry. Disparate GSs generate various glucans, which contain diverse structures and dissimilar linkage compositions, branching degrees, and size; meanwhile, it has vast applications [102,103]. There are four types of glucan (the enzyme produced), depending on the main glucosidic bonds in their glucan, including dextran, mutan, alternan, and reuteran (Figure 3) [13]. GS was discovered by Pasteur from microbial bacteria because of its positive spinning power and a comparable extracellular enzyme called dextransucrase. Dextran is generally composed of α-1,6 and α-1,3 glycosidic bonds based on specific dextransucrase [102,104]. As mentioned early, the majority of dextran in nature is synthesized from sucrose by dextransucrase, secreted mainly by LAB, like Steptococcus, Leuconostoc and Lactobacillus [7,105]. Guggenheim [106] isolated GTF from S. mutants OMZ176, which can synthesize dextran linked by α-1,3 glucosidic bonds. The corresponding GS is consequently named mutansucrase. Streptococci is the main source for producing the mutant. In order to figure out dextran and alternan, due to their similarity in linkages, Côté and Robyt named an α-glucan polymer as alternan, which also contained α-1,3 and α-1,6 glucosidic linkages [100]. The corresponding GS is named alternansucrase. Alternating α-1,6 and α-1,3 bonds are responsible for the polysaccharide’s resistance to the enzymatic hydrolysis of endoglucanase, and this is considered to be the determinant of its unique physical properties [100]. A new type of glucan was found, which mainly involves the α-1,4 bonds and α-1,6 linkages from L. reuteri 121, one of genus of LAB; it was named reuteran, and its corresponding enzyme is called reuteransucrase [7]. There are large alternating α-1,4 and α-1,6 linkages with no consecutive α-1,6 bonds [36,107]. GS catalyzes the formation of sucrose into EPSs; natural GS has a low yield, and its catalytic properties have not been fully analyzed, which has brought difficulties to the industrial application of EPS.
Previous studies have demonstrated that GS can synthesize high-molecular-weight polysaccharides from sucrose as a substrate. Kajala [79] synthesized dextran in vitro using partially purified GS under the conditions of 100 mL of 20 mM Na-acetate, pH 5.4, containing 2 mM CaCl2, and 146 mM sucrose at 35 °C for 24 h. Following overnight incubation, the glucan produced was precipitated with ethanol; it was freeze-dried and subjected to nuclear magnetic resonance spectroscopy (NMR) analysis [108]. Miao’s report stated that the glucan preparation was incubated at pH 5.2 and 35 °C for 0–48 h with 100 mg/mL sucrose [52]. The 1,6-,1,4-α-D-glucan polymer they obtained was synthesized in vitro from sucrose by GS and had a molecular weight of 2.5 × 107 g/mol. Wang incubating 100 g/L sucrose with wild-type and mutant dextransucrase (2.0 U/mL) in a calcium acetate buffer (50 mM, pH 5.4) at 25 °C for 24 h in a water bath to produce dextran [109]. Polymers were also synthesized by incubating 1 U/mL of GS in a 20 mmol sodium acetate buffer (pH 5.4) containing 100 g/L of sucrose, 0.05 g/L of CaCl2, and 1 g/L of sodium azide at 30 °C for 48 h [18]. Therefore, using the properties of GS to synthesize EPSs with different structures in vitro and increase the yield of the EPSs not only makes it possible to change the EPS (in a certain direction), but also helps to analyze the structure–activity relationship of EPS.
7. Acceptor Reaction
GS mainly catalyzes the production of different kinds of glucans, but when there are acceptor sugars in the system, this triggers the acceptor reaction to transfer part of the sucrose to other acceptor sugars, producing different types of glycosidic bonds and degrees of polymerization of oligosaccharides. There are many kinds of glucan receptors, mainly maltose, mannose, lactose, cellobiose, raffinose, catechol, ronobiose, melibiose, etc. [110,111,112] (Figure 4). These properties are exploited to produce functional oligosaccharides, in particular, isomalto-oligosaccharides [113], oligoalternans (oligosaccharides with alternating α-1,6 and α-1,3 linkages) [6,114,115,116], oligoreuterans with α-1,6, and α-1,4 linkages or α-1,2 branched gluco-oligosaccharides [8,9,11,12]. Two reactions are catalyzed by GSs, depending on the nature of the acceptor: (i) hydrolysis: in which water is used as acceptor, and (ii) glucosyl transfer (transferase). The latter reaction can be divided into (a) polymerization, in which the growing glucan chain is used as acceptor, and (b) oligosaccharide synthesis, in which oligosaccharides (e.g., maltose, isomaltose) are used as an acceptor [34].
The range of glucans and oligosaccharides produced by the GS present in Lactobacilli may potentially act as prebiotics by stimulating the growth of probiotic strains or beneficial endogenic strains of the gastrointestinal tract. Because the receptor reaction of the enzyme can form a prebiotic, the concept of prebiotics is gaining wider realization in the practice of producing healthy foods [117]. Natural and modified glucans also hold great potential in biotechnology, as well as in food and health-related applications, particularly because of their potential prebiotic properties. For example, glucose oligosaccharides containing more α-1,2- or α-1,3-linked glucose residues are thought to be able to induce a broad spectrum of glycolytic enzymes in probiotic bacteria without increasing gas production [117]. The GS contains two catalytic domains and is the second of this type reported to perform an efficient synthesis of branched-chain oligosaccharides with prebiotic potential. These oligosaccharides produced by glycosylation have excellent physiological properties, which gives them a positive role in industries such as food, feed, cosmetics and pharmaceuticals, and business (Figure 5). Some studies have also revealed the potential of certain oligosaccharides as prebiotics. Because of their different kinds of receptors, oligosaccharides with different oligosaccharide components can be produced. Therefore, it has become a hot issue to produce different kinds of oligosaccharides through receptor reactions to serve various fields of life.
7.1. Acceptor Reaction with Maltose
The type of glucosidic linkage formed in the product depends on the acceptor substrate and the specificity of the enzyme, and maltose is the most effective and studied of the acceptors, which produces isomalto-oligosaccharides (IMOs) [76]. Robyt and Eklund [118], in 1983, found that when maltose was an acceptor molecule, the GSs catalyzed a homologous series of IMOs composed mainly by consecutive α-1,6 bonds. Ispirli [108] used different sucrose concentrations together with maltose in the reaction mixture to test the acceptor reaction at different time intervals. The results showed that the GTFA-ΔN was useful in the production of different oligosaccharides when using sucrose as the donor and maltose as the acceptor sugar. According to the acceptor reaction, IMO was synthesized by using GS as the catalyzed acceptor and maltose as an acceptor molecule [104]. Then, the purified fractions were analyzed and identified by thin-layer chromatography (TLC), a liquid chromatograph mass Spectrometer (LC/MS) system, high-performance liquid chromatography (HPLC), and other methods.
7.2. Acceptor Reaction with Mannose
Mannose can be used as a clinical glyconutrient and is widely distributed in body fluids and tissues, and is also involved in immune regulation. It has properties such as being anti-inflammatory, antitumor, anti-infection, and helping to heal wounds [119,120]. In order to demonstrate the potential of GS to produce functional oligosaccharides, İspirli [112] produced mannose-containing oligosaccharides in an acceptor reaction with sucrose and mannose as the donor and acceptor sugars using the glucanase GTFA-DNE81. İspirli added 0.4 mg/mL sucrose and 0.1 mg/mL mannose (as donor sugar and acceptor sugar) to a 20 mM sodium acetate buffer containing 1 mM CaCl2 at pH 5.2 [112]. The oligosaccharides containing mannose were obtained by thin-layer chromatography and ESI-MS/MS analysis. The presence of the (1→6) Glc and (1→3) Glc units in the oligosaccharide was determined by NMR analysis. Cytokine induction was produced in terms of IL-4, IL-10, IL-12, and TNF-α cytokines tested by the addition of HT-29 colon cell lines in vitro [112]. This study revealed that the immune mannose oligosaccharides had a regulating effect. The mannose-containing oligosaccharides have a prebiotic effect on the tested probiotics. It was shown that the oligosaccharides generated by the receptor reaction can be widely used in the fields of food and medicine.
7.3. Acceptor Reaction with Lactose
Different kinds of glucosucrases and the receptor responses of acceptor sugars play an important role in the production of functional oligosaccharides. Lactose is also a commonly used acceptor sugar for GS, and GSs use sucrose and lactose as the donor and acceptor sugars to synthesize lactose derivative hetero-oligosaccharides [3,117]. Using GS GTFA-DNE81 and using sucrose and lactose as the donor and acceptor sugars, oligo-oligosaccharides containing whey components were produced. NMR and LC-MS analysis showed that they contained α-1,2, α-1,3, and α-1,4 units up to DP 7 in length, confirming that lactose is a good receptor for GS [111]. In order to confirm the prebiotic properties of the receptor product, İspirli used the receptor product as the carbon source; there was no growth of pathogenic bacteria, indicating that it contains a certain prebiotic function. The excellent physicochemical properties of lactose-derivative iso-oligosaccharides were revealed, which can be used as functional oligosaccharides for mass production [111]. It can be seen that the receptor reaction mechanism of GS can be used to synthesize oligosaccharides with different functional properties, and these oligosaccharides usually contain various special properties, including prebiotic properties and immune regulation, and are widely used in food and medicine. In the future, we should continue to explore the receptor responses of different GSs to different polysaccharides so as to characterize the physicochemical properties of the receptor products and then optimize the reaction conditions to serve the fields of food and chemical industry [121].
8. Analysis of EPS
EPSs have much diversity in composition due to the different origins of various strains. They consist of glucans, dextran, mutan, reteran, alternan, galactans, and fructans. The structure of EPS affects its physiological and biochemical properties and technical applications, so the properties of EPS and its microstructure, including molecular weight, monosaccharide composition, functional group analysis, sugar chain structure, glycosidic bond type, etc., have been widely studied to determine or characterize EPS-related functions and applications. EPS containing different structural properties exhibited distinct functional and physicochemical properties. EPS containing specific groups will have specific functions, such as the presence of uronic acid, sulfate groups, glucose, and β-glycosidic bonds, which may increase the anticancer effect of EPS [122,123]. Comprehensive analysis of the structure-activity relationship of EPS is a hot research topic at present. The structure of EPS cannot be thoroughly characterized by chemical methods. It usually needs to combine spectroscopy to thoroughly analyze the structure of EPS, usually including ultraviolet spectroscopy, infrared spectroscopy, and NMR spectroscopy. UV can detect whether the polysaccharide contains proteins, peptides, and nucleic acids. The structure of EPS and the type of glycosidic bond can be determined by infrared spectroscopy. The commonly used methods for analyzing EPS structure are shown in Figure 6.
8.1. Determination of Monosaccharide Composition in EPS
Monosaccharide analysis can determine carbohydrate content and verify the purity of the sample, including the configuration (D or L) of monosaccharide residues, their linking pattern, etc. Gas chromatography (GC) is suitable for the analysis and detection of volatile substances, but the high boiling point of monosaccharides does not have volatility, so volatile substances need to be generated first. Usually, trifluoroacetic acid is used for hydrolysis, reduction, and acetylation to obtain volatile monosaccharide and then for monosaccharide composition analysis. Li [122] studied the monosaccharide composition of the EPS from Streptococcus thermophilus 05-34 via gas chromatography-mass spectrometry (GC-MS). Dhanya [124] performed a monosaccharide analysis of EPS (EPS-DR3A) in marine bacterium using GC-MS, and the fragment pattern obtained in the TIC indicated the presence of monosaccharide derivatives. Gupta et al. used GC-MS to analyze the monosaccharide components of Cupriavidus sp. ISTL7 EPS and the results showed that EPS was a hybrid polymer composed of neutral sugars [125]. Compared with GC, HPLC does not require the derivatization step, which saves time on sample preparation. However, HPLC separates molecules based on their charge and polarity. Bejar [126] used an HPLC method to analyze the monosaccharide composition in an ion-exclusion ORH-801 column, which showed the resulting polymer had a dextran-like structure with predominant α-1,6 linkages. However, many monosaccharides have similar polarity or structure, and this method reduces the chromatographic resolution, resulting in inaccurate analysis when processing complex mixtures, so HPLC is often used in conjunction with GC-MS. Moreover, high-performance anion exchange chromatography (HPAEC) has also made an important contribution to monosaccharide analysis, which provides high-resolution analysis of the most common monosaccharides without the need for a derivatization step. Complete sucrose consumption and oligosaccharide formation can be analyzed by thin-layer chromatography (TLC) [108]. Ispirli’s result [107] showed that sucrose and glucose were completely depleted by TCL data, and the HPLC method could be used for further analysis, provided that the relative levels of different oligosaccharides in the mixture are checked.
8.2. Determination of Functional Groups in EPS
EPS has a variety of branch structures and functional groups, so EPS has a lot of biological characteristics and functions. It is of great significance to analyze the structure of EPS. Fourier transform infrared spectroscopy (FT-IR) has emerged as the most effective tool for monitoring structural changes in biopolymers. The peaks at different positions indicate that EPSs have different structures. The broad peak between 3032–3610 cm−1, corresponding to the hydroxyl groups of the polysaccharide, indicates that the polymer is a polysaccharide [127]. The absorption peak at 1153.27 cm−1 is the C-O-C stretching vibration, which is a specific signal of carbohydrates [128]. If there is a specific absorption peak of the α-glycosidic bond at 846 cm−1, this proves that glucan is α-polysaccharide. FT-IR spectroscopy can preliminarily prove the existence of sulfate, phosphate, uronic acid, mannose, protein, α or β configuration, pyranose, or furanose in EPSs [129]. Tyagi [130] confirmed via FT-IR that EPS is produced by Parapoda ISTM3, containing aliphatic groups, amine groups, uronic acid, and sugar groups [130].
8.3. Determination of Morphology in EPS
Scanning electron microscopy (SEM) can study the microstructure and surface topography of the copolymers, which helps to understand their common physical properties [128]. DU et al. used SEM to analyze the structure of dextran produced by W. confuse H2 under different acceleration voltages (2.0 and 3.0 kV). The results showed a smooth surface at 2000×, as well as its branched structure, revealing the ability of glucan to improve the surface structure of food and the rheological properties of food [97]. In addition, SEM analysis of EPS with Ln curvatus SJTUF 62116 showed that it had an irregular, smooth surface structure. Indicating that EPS-1 was a potential candidate to form a plasticized film and hydrated polymer consistent matrix [131].
An atomic force microscope (AFM) is a very useful tool to study and further understand their physical properties, 3D structure, and dynamics. AFM analysis of the structures of EPS extracted from LAB-induced gels of soy protein isolates revealed that the EPSs had rough and uneven surfaces with spider web-like structures, suggesting that EPS has potential as a biological thickening agent and stabilizer and has a strong water-holding capacity [105]. In addition to this, AFM can also show the interaction of EPS with other molecules [132]. The structural analysis of EPS produced by L. curvatus SJTUF 62116 using AFM images indicated that there may be interactions between EPS molecules and water molecules, providing hydrogen binding sites for water molecules. It is a potential stabilizer for the food and cosmetic industries [131].
SEM and atomic AFM are the most commonly used surface analysis methods. However, the working area of AFM is very difficult to select. The working area is very limited, so it is difficult to scan the surface of large samples. At present, researchers combine SEM with AFM, and use the nanoscale resolution and large-scale search ability of SEM to find and lock the target area for AFM research (as quickly as possible), which can greatly improve the efficiency of the AFM. The AFM and SEM are complementary to each other in microanalysis, which can continuously expand human vision, promote the better exploration of the micro world, and promote the progress of human science.
8.4. Determination of Structure in EPS
Methylation can determine the type and position of glycosidic bonds in monosaccharide analysis, thereby determining the structure of EPS. The first step is to convert the free hydroxyl groups of various monosaccharide residues in the polysaccharide into methoxy groups, followed by acid hydrolysis to various methylated monosaccharides. Then, combining GC and GC-MS to analyze the type and quantity of the methylated derivatives. The characteristic peaks were identified and analyzed, the connection mode of the glycosidic bonds was inferred, and the qualitative and quantitative analysis of the connection mode of the monosaccharide residues was obtained. For the methylation analysis of special polysaccharide and oligosaccharide samples, such as polysaccharides containing uronic acid, the connection mode of the uronic acid residues can be judged by comparing the methylation results before and after the reduction of the carboxyl groups [132,133]. Gerwig [134] methylation analysis result showed the presence of terminal Glcp and 2,3-di- O-substituted Glcp in the molar ratio of 3:1 in EPS. Methylation determination is a very important method to analyze the structure of EPS, but skillfully and correctly obtaining methylation data is still a problem. Attention to technical details and the use of the correct procedural steps are the keys to successful methylation analysis. In addition, periodate oxidation and Smith degradation methods can also be used to analyze the type and position of glycosidic bonds.
NMR is the most effective tool to identify the structure of EPS. It can accurately identify the basic structure of EPS, including ring size, isomeric configuration, and the position of glycosidic bonds. EPS glycosidic bond composition and configuration were measured by 1D NMR (1H and 13C NMR). The linked composition of the glycan produced by the mutant enzyme was analyzed by 600 MHz 1H NMR spectroscopy [56,135]. The proton chemical shift of α-pyranose H1 type is greater than 4.95 ppm, and the proton chemical shift of β-pyranose H1 type is less than 4.95 ppm. For example, DU et al. [136] used NMR to analyze the structure of the EPS produced by L. brevis HDE-9 [136]. Moreover, 2D NMR (HSQC, NOESY, and TOCSY) is also the main technique for analyzing the chemical composition of EPSs [52]. The 2D NMR method can detect the spin direction, linkage mode, the proton chemical change information of the residues, etc., of EPS and is the main means to detect the chemical composition of EPS. Feng et al. [128]. analyzed the EPS of L. plantarum AR307 by HSQC and HMBC, and the results showed that the backbone of EPS was composed of 1,4-α-D-Glcp, 1,4-β-D-Glcp, and was 1,4-β-D-Galp interrupted. It seems that NMR helped with the thorough characterization of EPS structure and was used to infer its possible functions. Generally, by combining methylation analysis and NMR spectroscopy, the linking pattern of the glycan will be determined, revealing the three-dimensional structure of EPS.
8.5. Thermal Analysis of EPS
Thermal analysis plays an important role in the detection of the physical and chemical properties of EPS. Generally speaking, the thermal analysis of EPS mainly includes thermogravimetric analysis (TGA), differential-scanning calorimetry (DSC), etc. [137]. Among them, TGA can analyze the weight-related quantity (such as mass, solid residue, or residual rate) of the sample as a function of temperature or time so as to obtain the thermal stability of the EPS, its thermal decomposition, its impurity composition, its thermal decomposition products, and other information [138]. DSC can analyze the pyrolysis mode and thermal stability of EPS. The TGA and DSC analysis of the thermal properties of L. reuteri E81 showed that the EPS content was still 11.77% at 600 °C, indicating the high temperature resistance of E81, which can be widely used in the production of sourdough bread and other high-temperature processes in the food industry [139]. For more complex EPSs, their structure, pyrolysis process, main functional groups, etc. (such as TGA-FI-TR TGA-MS) can also be used for analysis.
9. Conclusions
The above studies have shown that GS catalyzes this reaction and uses sucrose as a substrate to synthesize glucan, oligosaccharide, glucoside, and other products with many applications and a high economic value, which provides a new method for sucrose processing. However, EPSs with different structures have different properties. The production of EPSs with different structures is mainly affected by the structure and properties of GS. However, the structure and properties of GS have not been fully resolved. Biological techniques and chemical methods were used to analyze the structure and properties of EPS, exploring its regulatory mechanism in EPS biosynthesis, and promoting the further development of glycobiology.
Author Contributions
L.Y.: Methodology, Data curation, Writing—original draft. Z.Q.: Supervision. J.G.: Review, Editing. R.D.: Resources, Supervision. All authors have read and agreed to the published version of the manuscript.
Funding
Outstanding Youth Fund Project of Natural Science Foundation of Heilongjiang Province [No. YQ2021C030] (R.D.). Project funded by China Postdoctoral Science Foundation [No. 2022MD713755] (R.D.). Heilongjiang Province Postdoctoral Funding Project [No. LBH-Z21082] (R.D.). Open Funding Project of State Key Laboratory of Microbial Metabolism [No. MMLKF22-07] (R.D.) and Heilongjiang Province provincial colleges and universities basic scientific research business expenses scientific research projects [No. 2021-KYYWF-0045] (R.D.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Xu, Y.; Cui, Y.; Yue, F.; Liu, L.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Exopolysaccharides produced by lactic acid bacteria and Bifidobacteria: Structures, physiochemical functions and applications in the food industry. Food Hydrocoll. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Rana, S.; Upadhyay, L.S.B. Microbial exopolysaccharides: Synthesis pathways, types and their commercial applications. Int. J. Biol. Macromol. 2020, 157, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.; Pijning, T.; Dijkhuizen, L.; van Leeuwen, S.S. Mutational analysis of the role of the glucansucrase Gtf180-ΔN active site residues in product and linkage specificity with lactose as acceptor substrate. J. Agric. Food Chem. 2018, 66, 12544–12554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Pijning, T.; Tietema, M.; Dobruchowska, J.M.; Yin, H.; Gerwig, G.J.; Kralj, S.; Dijkhuizen, L. Characterization of the glucansucrase GTF180 W1065 mutant enzymes producing polysaccharides and oligosaccharides with altered linkage composition. Food Chem. 2017, 217, 81–90. [Google Scholar] [CrossRef]
- Molina, M.; Cioci, G.; Moulis, C.; Séverac, E.; Remaud-Siméon, M. Bacterial α-glucan and branching sucrases from GH70 family: Discovery, structure–function relationship studies and engineering. Microorganisms 2021, 9, 1607. [Google Scholar] [CrossRef]
- Holt, S.M.; Skory, C.; Cote, G. Enzymatic synthesis of artificial polysaccharides. ACS Sustain. Chem. Eng. 2020, 8, 11853–11871. [Google Scholar] [CrossRef]
- Chen, Z.; Ni, D.; Zhang, W.; Stressler, T.; Mu, W. Lactic acid bacteria-derived α-glucans: From enzymatic synthesis to miscellaneous applications. Biotechnol. Adv. 2021, 47, 107708. [Google Scholar] [CrossRef]
- Kim, M.; Jang, J.-K.; Park, Y.-S. Production optimization, structural analysis, and prebiotic-and anti-inflammatory effects of gluco-oligosaccharides produced by Leuconostoc lactis SBC001. Microorganisms 2021, 9, 200. [Google Scholar] [CrossRef]
- Hasselwander, O.; DiCosimo, R.; You, Z.; Cheng, Q.; Rothman, S.C.; Suwannakham, S.; Baer, Z.C.; Roesch, B.M.; Ruebling-Jass, K.D.; Lai, J.P. Development of dietary soluble fibres by enzymatic synthesis and assessment of their digestibility in vitro, animal and randomised clinical trial models. Int. J. Food Sci. Nutr. 2017, 68, 849–864. [Google Scholar] [CrossRef] [Green Version]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- Sarbini, S.R.; Kolida, S.; Naeye, T.; Einerhand, A.W.; Gibson, G.R.; Rastall, R.A. The prebiotic effect of α-1, 2 branched, low molecular weight dextran in the batch and continuous faecal fermentation system. J. Funct. Foods 2013, 5, 1938–1946. [Google Scholar] [CrossRef]
- Sarbini, S.R.; Kolida, S.; Naeye, T.; Einerhand, A.; Brison, Y.; Remaud-Simeon, M.; Monsan, P.; Gibson, G.R.; Rastall, R.A. In vitro fermentation of linear and α-1, 2-branched dextrans by the human fecal microbiota. Appl. Environ. Microbiol. 2011, 77, 5307–5315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daba, G.M.; Elnahas, M.O.; Elkhateeb, W.A. Contributions of exopolysaccharides from lactic acid bacteria as biotechnological tools in food, pharmaceutical, and medical applications. Int. J. Biol. Macromol. 2021, 173, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Korcz, E.; Kerényi, Z.; Varga, L. Dietary fibers, prebiotics, and exopolysaccharides produced by lactic acid bacteria: Potential health benefits with special regard to cholesterol-lowering effects. Food Funct. 2018, 9, 3057–3068. [Google Scholar] [CrossRef]
- Kabli, M.; İspirli, H.; Balubaid, M.; Taylan, O.; Yılmaz, M.T.; Dertli, E. Optimization of lactose derivative hetero-oligosaccharides production using whey as the acceptor molecule by an active glucansucrase. Biocatal. Biotransform. 2022, 40, 9–16. [Google Scholar] [CrossRef]
- Song, L.; Miao, M.; Jiang, B.; Xu, T.; Cui, S.W.; Zhang, T. Leuconostoc citreum SK24.002 glucansucrase: Biochemical characterisation and de novo synthesis of α-glucan. Int. J. Biol. Macromol. 2016, 91, 123–131. [Google Scholar] [CrossRef]
- Guzman, G.Y.F.; Hurtado, G.B.; Ospina, S.A. New dextransucrase purification process of the enzyme produced by Leuconostoc mesenteroides IBUN 91.2. 98 based on binding product and dextranase hydrolysis. J. Biotechnol. 2018, 265, 8–14. [Google Scholar] [CrossRef]
- Vasileva, T.; Bivolarski, V.; Michailova, G.; Salim, A.; Rabadjiev, Y.; Ivanova, I.; Iliev, I. Glucansucrases produced by fructophilic lactic acid bacteria Lactobacillus kunkeei H3 and H25 isolated from honeybees. J. Basic Microbiol. 2017, 57, 68–77. [Google Scholar] [CrossRef]
- Meng, X.; Li, X.; Pijning, T.; Wang, X.; van Leeuwen, S.S.; Dijkhuizen, L.; Chen, G.; Liu, W. Characterization of the (engineered) branching sucrase gtfz-cd2 from Apilactobacillus kunkeei for efficient glucosylation of benzenediol compounds. Appl. Environ. Microbiol. 2022, 88, e01031-22. [Google Scholar] [CrossRef]
- Vujičić-Žagar, A.; Pijning, T.; Kralj, S.; López, C.A.; Eeuwema, W.; Dijkhuizen, L.; Dijkstra, B.W. Crystal structure of a 117 kDa glucansucrase fragment provides insight into evolution and product specificity of GH70 enzymes. Proc. Natl. Acad. Sci. USA 2010, 107, 21406–21411. [Google Scholar] [CrossRef]
- Ito, K.; Ito, S.; Shimamura, T.; Weyand, S.; Kawarasaki, Y.; Misaka, T.; Abe, K.; Kobayashi, T.; Cameron, A.D.; Iwata, S. Crystal structure of glucansucrase from the dental caries pathogen Streptococcus mutans. J. Mol. Biol. 2011, 408, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Stam, M.R.; Danchin, E.G.; Rancurel, C.; Coutinho, P.M.; Henrissat, B. Dividing the large glycoside hydrolase family 13 into subfamilies: Towards improved functional annotations of α-amylase-related proteins. Protein Eng. Des. Sel. 2006, 19, 555–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangoiti, J.; Pijning, T.; Dijkhuizen, L. Biotechnological potential of novel glycoside hydrolase family 70 enzymes synthesizing α-glucans from starch and sucrose. Biotechnol. Adv. 2018, 36, 196–207. [Google Scholar] [CrossRef]
- Meng, X.; Gangoiti, J.; Bai, Y.; Pijning, T.; Van Leeuwen, S.S.; Dijkhuizen, L. Structure–function relationships of family GH70 glucansucrase and 4, 6-α-glucanotransferase enzymes, and their evolutionary relationships with family GH13 enzymes. Cell. Mol. Life Sci. 2016, 73, 2681–2706. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.T.; Zhang, H.B.; Huang, L.J.; Hu, X.Q. Purification and characterization of extracellular dextranase from a novel producer, Hypocrea lixii F1002, and its use in oligodextran production. Process Biochem. 2011, 46, 1942–1950. [Google Scholar] [CrossRef]
- Moulis, C.; Vaca Medina, G.; Suwannarangsee, S.; Monsan, P.; Remaud-Simeon, M.; Potocki-Veronese, G. One-step synthesis of isomalto-oligosaccharide syrups and dextrans of controlled size using engineered dextransucrase. Biocatal. Biotransform. 2008, 26, 141–151. [Google Scholar] [CrossRef]
- Leemhuis, H.; Pijning, T.; Dobruchowska, J.M.; van Leeuwen, S.S.; Kralj, S.; Dijkstra, B.W.; Dijkhuizen, L. Glucansucrases: Three-dimensional structures, reactions, mechanism, α-glucan analysis and their implications in biotechnology and food applications. J. Biotechnol. 2013, 163, 250–272. [Google Scholar] [CrossRef] [Green Version]
- van Hijum, S.A.; Kralj, S.; Ozimek, L.K.; Dijkhuizen, L.; van Geel-Schutten, I.G. Structure-function relationships of glucansucrase and fructansucrase enzymes from lactic acid bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 157–176. [Google Scholar] [CrossRef] [Green Version]
- André, I.; Potocki-Véronese, G.; Morel, S.; Monsan, P.; Remaud-Siméon, M. Sucrose-utilizing transglucosidases for biocatalysis. Carbohydr. Sustain. Dev. I 2010, 294, 25–48. [Google Scholar] [CrossRef]
- Korakli, M.; Vogel, R.F. Structure/function relationship of homopolysaccharide producing glycansucrases and therapeutic potential of their synthesised glycans. Appl. Microbiol. Biotechnol. 2006, 71, 790–803. [Google Scholar] [CrossRef]
- Robyt, J.F.; Yoon, S.H.; Mukerjea, R. Dextransucrase and the mechanism for dextran biosynthesis. Carbohydr. Res. 2008, 343, 3039–3048. [Google Scholar] [CrossRef] [PubMed]
- Falconer, D.J.; Mukerjea, R.; Robyt, J.F. Biosynthesis of dextrans with different molecular weights by selecting the concentration of Leuconostoc mesenteroides B-512FMC dextransucrase, the sucrose concentration, and the temperature. Carbohydr. Res. 2011, 346, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Robyt, J.F.; Lee, S.Y.; Lee, J.H.; Kim, Y.M. Dextran molecular size and degree of branching as a function of sucrose concentration, pH, and temperature of reaction of Leuconostoc mesenteroides B-512FMCM dextransucrase. Carbohydr. Res. 2003, 338, 1183–1189. [Google Scholar] [CrossRef]
- Kralj, S.; van Geel-Schutten, G.H.; Dondorff, M.M.G.; Kirsanovs, S.; Van Der Maarel, M.; Dijkhuizen, L. Glucan synthesis in the genus Lactobacillus: Isolation and characterization of glucansucrase genes, enzymes and glucan products from six different strains. Microbiology 2004, 150, 3681–3690. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Cho, S.K.; Eom, H.J.; Kim, S.Y.; Kim, T.J.; Han, N.S. Optimized substrate concentrations for production of long-chain isomaltooligosaccharides using dextransucrase of Leuconostoc mesenteroides B-512F. J. Microbiol. Biotechnol. 2008, 18, 1141–1145. [Google Scholar]
- Meng, X.; Dobruchowska, J.M.; Pijning, T.; Gerwig, G.J.; Kamerling, J.P.; Dijkhuizen, L. Truncation of domain V of the multidomain glucansucrase GTF180 of Lactobacillus reuteri 180 heavily impairs its polysaccharide-synthesizing ability. Appl. Microbiol. Biotechnol. 2015, 99, 5885–5894. [Google Scholar] [CrossRef]
- Lawson, C.L.; van Montfort, R.; Strokopytov, B.; Rozeboom, H.J.; Kalk, K.H.; de Vries, G.E.; Penninga, D.; Dijkhuizen, L.; Dijkstra, B.W. Nucleotide sequence and X-ray structure of cyclodextrin glycosyltransferase from Bacillus circulans strain 251 in a maltose-dependent crystal form. J. Mol. Biol. 1994, 236, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Kralj, S.; Eeuwema, W.; Eckhardt, T.H.; Dijkhuizen, L. Role of asparagine 1134 in glucosidic bond and transglycosylation specificity of reuteransucrase from Lactobacillus reuteri 121. FEBS J. 2006, 273, 3735–3742. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, S.S.; Kralj, S.; van Geel-Schutten, I.H.; Gerwig, G.J.; Dijkhuizen, L.; Kamerling, J.P. Structural analysis of the α-D-glucan (EPS180) produced by the Lactobacillus reuteri strain 180 glucansucrase GTF180 enzyme. Carbohydr. Res. 2008, 343, 1237–1250. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, S.; Lona, L.; Franco, T. Effect of phosphate concentration on the production of dextransucrase by Leuconostoc mesenteroides NRRL B512F. Bioprocess Biosyst. Eng. 2003, 26, 57–62. [Google Scholar] [CrossRef]
- Otts, D.; Day, D.F. Dextransucrase secretion in Leuconostoc mesenteroides depends on the presence of a transmembrane proton gradient. J. Bacteriol. 1988, 170, 5006–5011. [Google Scholar] [CrossRef] [PubMed]
- Quirasco, M.; Lopez-Munguia, A.; Remaud-Simeon, M.; Monsan, P.; Farres, A. Induction and transcription studies of the dextransucrase gene in Leuconostoc mesenteroides NRRL B-512F. Appl. Environ. Microbiol. 1999, 65, 5504–5509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, A.; Katiyar, S.S. Fractionation of Leuconostoc mesenteroides NRRL B-512F dextran sucrase by polyethylene glycol: A simple and effective method purification. J. Microbiol. Methods 1994, 20, 225–231. [Google Scholar] [CrossRef]
- Purama, R.K.; Goyal, A. Identification, effective purification and functional characterization of dextransucrase from Leuconostoc mesenteroides NRRL B-640. Bioresour. Technol. 2008, 99, 3635–3642. [Google Scholar] [CrossRef]
- Kobayashi, M.; Matsuda, K. Electrophoretic analysis of the multiple forms of dextransucrase from Leuconostoc mesenteroides. J. Biochem. 1986, 100, 615–621. [Google Scholar] [CrossRef]
- Miller, A.W.; Eklund, S.H.; Robyt, J.F. Milligram to gram scale purification and characterization of dextransucrase from Leuconostoc mesenteroides NRRL B-512F. Carbohydr. Res. 1986, 147, 119–133. [Google Scholar] [CrossRef]
- Miller, A.W.; Robyt, J.F. Stabilization of dextransucrase from Leuconostoc mesenteroides NRRL B-512F by nonionic detergents, poly(ethylene glycol) and high-molecular-weight dextran. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1984, 785, 89–96. [Google Scholar] [CrossRef]
- Kitaoka, M.; Robyt, J.F. Use of a microtiter plate screening method for obtaining Leuconostoc mesenteroides mutants constitutive for glucansucrase. Enzym. Microb. Technol. 1998, 22, 527–531. [Google Scholar] [CrossRef]
- Monsan, P.; Lopez, A. On the production of dextran by free and immobilized dextransucrase. Biotechnol. Bioeng. 1981, 23, 2027–2037. [Google Scholar] [CrossRef]
- Neubauer, H.; Bauche, A.; Mollet, B. Molecular characterization and expression analysis of the dextransucrase DsrD of Leuconostoc mesenteroides Lcc4 in homologous and heterologous Lactococcus lactis cultures. Microbiology 2003, 149, 973–982. [Google Scholar] [CrossRef]
- Robyt, J.F.; Walseth, T.F. Production, purification, and properties of dextransucrase from Leuconostoc mesenteroides NRRL B-512F. Carbohydr. Res. 1979, 68, 95–111. [Google Scholar] [CrossRef]
- Miao, M.; Ma, Y.; Jiang, B.; Cui, S.W.; Jin, Z.; Zhang, T. Characterisations of Lactobacillus reuteri SK24. 003 glucansucrase: Implications for α-gluco-poly and oligosaccharides biosynthesis. Food Chem. 2017, 222, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Nigam, M.; Goyal, A.; Katiyar, S.S. High yield purification of dextransucrase from Leuconostoc mesenteroides NRRL B-512F by phase partitioning. J. Food Biochem. 2006, 30, 12–20. [Google Scholar] [CrossRef]
- Amari, M.; Arango, L.; Gabriel, V.; Robert, H.; Morel, S.; Moulis, C.; Gabriel, B.; Remaud-Siméon, M.; Fontagné-Faucher, C. Characterization of a novel dextransucrase from Weissella confusa isolated from sourdough. Appl. Microbiol. Biotechnol. 2013, 97, 5413–5422. [Google Scholar] [CrossRef]
- Wang, C.; Chen, S.; Zhang, H.B.; Li, Y.; Hu, X.Q. Characterization of the inserted mutagenesis dextransucrases from Leuconostoc mesenteroides 0326 to produce hyperbranched dextran. Int. J. Biol. Macromol. 2018, 112, 584–590. [Google Scholar] [CrossRef]
- Kim, Y.M.; Yeon, M.J.; Choi, N.S.; Chang, Y.H.; Jung, M.Y.; Song, J.J.; Kim, J.S. Purification and characterization of a novel glucansucrase from Leuconostoc lactis EG001. Microbiol. Res. 2010, 165, 384–391. [Google Scholar] [CrossRef]
- Rühmkorf, C.; Bork, C.; Mischnick, P.; Rübsam, H.; Becker, T.; Vogel, R.F. Identification of Lactobacillus curvatus TMW 1.624 dextransucrase and comparative characterization with Lactobacillus reuteri TMW 1.106 and Lactobacillus animalis TMW 1.971 dextransucrases. Food Microbiol. 2013, 34, 52–61. [Google Scholar] [CrossRef]
- Vidal, R.F.; Martínez, A.; Moulis, C.; Escalier, P.; Morel, S.; Remaud-Simeon, M.; Monsan, P. A novel dextransucrase is produced by Leuconostoc citreum strain B/110-1-2: An isolate used for the industrial production of dextran and dextran derivatives. J. Ind. Microbiol. Biotechnol. 2011, 38, 1499–1506. [Google Scholar] [CrossRef]
- Yi, A.R.; Lee, S.R.; Jang, M.U.; Park, J.M.; Eom, H.J.; Han, N.S.; Kim, T.J. Cloning of dextransucrase gene from Leuconostoc citreum HJ-P4 and its high-level expression in E. coli by low temperature induction. J. Microbiol. Biotechnol. 2009, 19, 829–835. [Google Scholar] [CrossRef]
- Ko, J.A.; Jeong, H.J.; Ryu, Y.B.; Park, S.J.; Wee, Y.J.; Kim, D.; Kim, Y.M.; Lee, W.S. Large increase in Leuconostoc citreum KM20 dextransucrase activity achieved by changing the strain/inducer combination in an E. coli expression system. J. Microbiol. Biotechnol. 2012, 22, 510–515. [Google Scholar] [CrossRef]
- Passerini, D.; Vuillemin, M.; Ufarté, L.; Morel, S.; Loux, V.; Fontagné-Faucher, C.; Monsan, P.; Remaud-Siméon, M.; Moulis, C. Inventory of the GH70 enzymes encoded by Leuconostoc citreum NRRL B-1299-identification of three novel α-transglucosylases. FEBS J. 2015, 282, 2115–2130. [Google Scholar] [CrossRef] [PubMed]
- Monchois, V.; Remaud-Simeon, M.; Monsan, P.; Willemot, R.M. Cloning and sequencing of a gene coding for an extracellular dextransucrase (DSRB) from Leuconostoc mesenteroides NRRL B-1299 synthesizing only a alpha (1-6) glucan. FEMS Microbiol. Lett. 1998, 159, 307–315. [Google Scholar] [CrossRef]
- Bozonnet, S.; Dols-Laffargue, M.; Fabre, E.; Pizzut, S.; Remaud-Simeon, M.; Monsan, P.; Willemot, R.M. Molecular characterization of DSR-E, an alpha-1,2 linkage-synthesizing dextransucrase with two catalytic domains. J. Bacteriol. 2002, 184, 5753–5761. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hu, Y.; Zhu, C.; Zhu, B.; Wang, Y. Cloning, sequencing and expression of a dextransucrase gene (dexYG) from Leuconostoc mesenteroides. Biotechnol. Lett. 2008, 30, 1441–1446. [Google Scholar] [CrossRef]
- Kang, H.K.; Kim, Y.M.; Kim, D.M. Functional, genetic, and bioinformatic characterization of dextransucrase (DSRBCB4) gene in Leuconostoc mesenteroides B-1299 CB4. J. Microbiol. Biotechnol. 2008, 18, 1050–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Fulton, D.B.; Robyt, J.F. Enzymatic synthesis of L-DOPA alpha-glycosides by reaction with sucrose catalyzed by four different glucansucrases from four strains of Leuconostoc mesenteroides. Carbohydr. Res. 2010, 345, 1730–1735. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Bruce Fulton, D.; Robyt, J.F. Enzymatic synthesis of two salicin analogues by reaction of salicyl alcohol with Bacillus macerans cyclomaltodextrin glucanyltransferase and Leuconostoc mesenteroides B-742CB dextransucrase. Carbohydr. Res. 2004, 339, 1517–1529. [Google Scholar] [CrossRef]
- Chellapandian, M.; Larios, C.; Sanchez-Gonzalez, M.; Lopez-Munguia, A. Production and properties of a dextransucrase from Leuconostoc mesenteroides IBT-PQ isolated from ‘pulque’, a traditional Aztec alcoholic beverage. J. Ind. Microbiol. Biotechnol. 1998, 21, 51–56. [Google Scholar] [CrossRef]
- Siddiqui, N.N.; Aman, A.; Qader, S.A. Mutational analysis and characterization of dextran synthesizing enzyme from wild and mutant strain of Leuconostoc mesenteroides. Carbohydr. Polym. 2013, 91, 209–216. [Google Scholar] [CrossRef]
- Yalin, Y.; Jin, L.; Jianhua, W.; Da, T.; Zigang, T. Expression and characterization of dextransucrase gene dsrX from Leuconostoc mesenteroides in Escherichia coli. J. Biotechnol. 2008, 133, 505–512. [Google Scholar] [CrossRef]
- Monchois, V.; Remaud-Simeon, M.; Russell, R.R.; Monsan, P.; Willemot, R.M. Characterization of Leuconostoc mesenteroides NRRL B-512F dextransucrase (DSRS) and identification of amino-acid residues playing a key role in enzyme activity. Appl. Microbiol. Biotechnol. 1997, 48, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Funane, K.; Ishii, T.; Matsushita, M.; Hori, K.; Mizuno, K.; Takahara, H.; Kitamura, Y.; Kobayashi, M. Water-soluble and water-insoluble glucans produced by Escherichia coli recombinant dextransucrases from Leuconostoc mesenteroides NRRL B-512F. Carbohydr. Res. 2001, 334, 19–25. [Google Scholar] [CrossRef]
- Gilmore, K.S.; Russell, R.R.; Ferretti, J.J. Analysis of the Streptococcus downei gtfS gene, which specifies a glucosyltransferase that synthesizes soluble glucans. Infect. Immun. 1990, 58, 2452–2458. [Google Scholar] [CrossRef] [Green Version]
- Hanada, N.; Fukushima, K.; Nomura, Y.; Senpuku, H.; Hayakawa, M.; Mukasa, H.; Shiroza, T.; Abiko, Y. Cloning and nucleotide sequence analysis of the Streptococcus sobrinus gtfU gene that produces a highly branched water-soluble glucan. Biochim. Biophys. Acta 2002, 1570, 75–79. [Google Scholar] [CrossRef]
- Tsumori, H.; Minami, T.; Kuramitsu, H.K. Identification of essential amino acids in the Streptococcus mutans glucosyltransferases. J. Bacteriol. 1997, 179, 3391–3396. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, A.; Nakano, Y.J.; Mukasa, H.; Kuramitsu, H.K. Identification of amino acid residues in Streptococcus mutans glucosyltransferases influencing the structure of the glucan product. J. Bacteriol. 1994, 176, 4845–4850. [Google Scholar] [CrossRef] [Green Version]
- Bounaix, M.S.; Robert, H.; Gabriel, V.; Morel, S.; Remaud-Siméon, M.; Gabriel, B.; Fontagné-Faucher, C. Characterization of dextran-producing Weissella strains isolated from sourdoughs and evidence of constitutive dextransucrase expression. FEMS Microbiol. Lett. 2010, 311, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Shi, Q.; Maina, N.H.; Juvonen, M.; Goyal, A. Weissella confusa Cab3 dextransucrase: Properties and in vitro synthesis of dextran and glucooligosaccharides. Carbohydr. Polym. 2014, 101, 554–564. [Google Scholar] [CrossRef]
- Kajala, I.; Shi, Q.; Nyyssölä, A.; Maina, N.H.; Hou, Y.; Katina, K.; Tenkanen, M.; Juvonen, R. Cloning and characterization of a Weissella confusa dextransucrase and its application in high fibre baking. PLoS ONE 2015, 10, e0116418. [Google Scholar] [CrossRef] [Green Version]
- Wangpaiboon, K.; Padungros, P.; Nakapong, S.; Charoenwongpaiboon, T.; Rejzek, M.; Field, R.A.; Pichyangkura, R. An α-1,6-and α-1,3-linked glucan produced by Leuconostoc citreum ABK-1 alternansucrase with nanoparticle and film-forming properties. Sci. Rep. 2018, 8, 8340. [Google Scholar] [CrossRef]
- Miao, M.; Ma, Y.; Jiang, B.; Huang, C.; Li, X.; Cui, S.W.; Zhang, T. Structural investigation of a neutral extracellular glucan from Lactobacillus reuteri SK24.003. Carbohydr. Polym. 2014, 106, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Argüello-Morales, M.A.; Remaud-Simeon, M.; Pizzut, S.; Sarçabal, P.; Willemot, R.; Monsan, P. Sequence analysis of the gene encoding alternansucrase, a sucrose glucosyltransferase from Leuconostoc mesenteroides NRRL B-1355. FEMS Microbiol. Lett. 2000, 182, 81–85. [Google Scholar] [CrossRef]
- Kralj, S.; Stripling, E.; Sanders, P.; van Geel-Schutten, G.H.; Dijkhuizen, L. Highly hydrolytic reuteransucrase from probiotic Lactobacillus reuteri strain ATCC 55730. Appl. Environ. Microbiol. 2005, 71, 3942–3950. [Google Scholar] [CrossRef] [Green Version]
- Miao, M.; Ma, Y.; Huang, C.; Jiang, B.; Cui, S.W.; Zhang, T. Physicochemical properties of a water soluble extracellular homopolysaccharide from Lactobacillus reuteri SK24.003. Carbohydr. Polym. 2015, 131, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Côté, G.L.; Skory, C.D. Cloning, expression, and characterization of an insoluble glucan-producing glucansucrase from Leuconostoc mesenteroides NRRL B-1118. Appl. Microbiol. Biotechnol. 2012, 93, 2387–2394. [Google Scholar] [CrossRef] [PubMed]
- Shiroza, T.; Ueda, S.; Kuramitsu, H.K. Sequence analysis of the gtfB gene from Streptococcus mutans. J. Bacteriol. 1987, 169, 4263–4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, S.; Shiroza, T.; Kuramitsu, H.K. Sequence analysis of the gtfC gene from Streptococcus mutans GS-5. Gene 1988, 69, 101–109. [Google Scholar] [CrossRef]
- Gangoiti, J.; van Leeuwen, S.S.; Gerwig, G.J.; Duboux, S.; Vafiadi, C.; Pijning, T.; Dijkhuizen, L. 4,3-α-Glucanotransferase, a novel reaction specificity in glycoside hydrolase family 70 and clan GH-H. Sci. Rep. 2017, 7, 39761. [Google Scholar] [CrossRef] [Green Version]
- Dobruchowska, J.M.; Gerwig, G.J.; Kralj, S.; Grijpstra, P.; Leemhuis, H.; Dijkhuizen, L.; Kamerling, J.P. Structural characterization of linear isomalto-/malto-oligomer products synthesized by the novel GTFB 4,6-α-glucanotransferase enzyme from Lactobacillus reuteri 121. Glycobiology 2012, 22, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Leemhuis, H.; Dijkman, W.P.; Dobruchowska, J.M.; Pijning, T.; Grijpstra, P.; Kralj, S.; Kamerling, J.P.; Dijkhuizen, L. 4,6-α-Glucanotransferase activity occurs more widespread in Lactobacillus strains and constitutes a separate GH70 subfamily. Appl. Microbiol. Biotechnol. 2013, 97, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Pijning, T.; Gangoiti, J.; Te Poele, E.M.; Börner, T.; Dijkhuizen, L. Insights into Broad-Specificity Starch Modification from the Crystal Structure of Limosilactobacillus Reuteri NCC 2613 4,6-α-Glucanotransferase GtfB. J. Agric. Food Chem. 2021, 69, 13235–13245. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Oh, J.S.; Kim, D. Molecular characterization and expression analysis of the glucansucrase DSRWC from Weissella cibaria synthesizing a alpha (1→6) glucan. FEMS Microbiol. Lett. 2009, 292, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangoiti, J.; van Leeuwen, S.S.; Vafiadi, C.; Dijkhuizen, L. The Gram-negative bacterium Azotobacter chroococcum NCIMB 8003 employs a new glycoside hydrolase family 70 4,6-α-glucanotransferase enzyme (GtfD) to synthesize a reuteran like polymer from maltodextrins and starch. Biochim. Biophys. Acta 2016, 1860, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Gangoiti, J.; Pijning, T.; Dijkhuizen, L. The Exiguobacterium sibiricum 255-15 GtfC Enzyme Represents a Novel Glycoside Hydrolase 70 Subfamily of 4,6-α-Glucanotransferase Enzymes. Appl. Environ. Microbiol. 2016, 82, 756–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, H.; Koepsell, H.; Corman, J.; Bryant, G.; Bogard, M.; Feger, V.; Jackson, R. The effect of certain cultural factors on production of dextransucrase by Leuconostoc mesenteroides. J. Bacteriol. 1952, 64, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, R.; Zhao, F.; Pan, L.; Han, Y.; Xiao, H.; Zhou, Z. Optimization and purification of glucansucrase produced by Leuconostoc mesenteroides DRP2-19 isolated from Chinese Sauerkraut. Prep. Biochem. Biotechnol. 2018, 48, 465–473. [Google Scholar] [CrossRef]
- Molina, M.; Moulis, C.; Monties, N.; Pizzut-Serin, S.; Guieysse, D.; Morel, S.; Cioci, G.; Remaud-Simeéon, M. Deciphering an undecided enzyme: Investigations of the structural determinants involved in the linkage specificity of alternansucrase. ACS Catal. 2019, 9, 2222–2237. [Google Scholar] [CrossRef]
- Qader, S.A.U.; Aman, A.; Bano, S.; Syed, N.; Azhar, A. The effect of calcium ions and temperature on the production, activity and stability of dextransucrase from the newly isolated strain Leuconostoc mesenteroides PCSIR-4. Rom. J. Biochem. 2008, 45, 159–168. [Google Scholar] [CrossRef]
- Bai, Y.; van der Kaaij, R.M.; Leemhuis, H.; Pijning, T.; van Leeuwen, S.S.; Jin, Z.; Dijkhuizen, L. Biochemical characterization of the Lactobacillus reuteri glycoside hydrolase family 70 GTFB type of 4, 6-α-glucanotransferase enzymes that synthesize soluble dietary starch fibers. Appl. Environ. Microbiol. 2015, 81, 7223–7232. [Google Scholar] [CrossRef] [Green Version]
- Côté, G.L.; Robyt, J.F. Isolation and partial characterization of an extracellular glucansucrase from Leuconostoc mesenteroides NRRL B-1355 that synthesizes an alternating (1→6), (1→3)-α-D-glucan. Carbohydr. Res. 1982, 101, 57–74. [Google Scholar] [CrossRef]
- Du, R.; Qiao, X.; Wang, Y.; Zhao, B.; Han, Y.; Zhou, Z. Determination of glucansucrase encoding gene in Leuconostoc mesenteroides. Int. J. Biol. Macromol. 2019, 137, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Su, L.; Wang, L.; Wu, J.; Chen, S. Alpha-glucanotransferase from the glycoside hydrolase family synthesizes α (1–6)-linked products from starch: Features and synthesis pathways of the products. Trends Food Sci. Technol. 2022, 128, 160–172. [Google Scholar] [CrossRef]
- Díaz-Montes, E. Dextran: Sources, structures, and properties. Polysaccharides 2021, 2, 554–565. [Google Scholar] [CrossRef]
- Kothari, D.; Goyal, A. Structural characterization of enzymatically synthesized dextran and oligosaccharides from Leuconostoc mesenteroides NRRL B-1426 dextransucrase. Biochemistry 2013, 78, 1164–1170. [Google Scholar] [CrossRef]
- Wu, J.; Han, X.; Ye, M.; Li, Y.; Wang, X.; Zhong, Q. Exopolysaccharides synthesized by lactic acid bacteria: Biosynthesis pathway, structure-function relationship, structural modification and applicability. Crit. Rev. Food Sci. Nutr. 2022, 25, 1–22. [Google Scholar] [CrossRef]
- Guggentheim, B. Enzymatic hydrolysis and structure of water-insoluble glucan produced by glucosyltransferase from a strain of Streptococcus mutans. Helv. Odontol. Acta 1970, 5, 89–108. [Google Scholar]
- Kralj, S.; van Geel-Schutten, G.; Rahaoui, H.; Leer, R.; Faber, E.; Van Der Maarel, M.; Dijkhuizen, L. Molecular characterization of a novel glucosyltransferase from Lactobacillus reuteri strain 121 synthesizing a unique, highly branched glucan with α-(1→4) and α-(1→6) glucosidic bonds. Appl. Environ. Microbiol. 2002, 68, 4283–4291. [Google Scholar] [CrossRef] [Green Version]
- İspirli, H.; Yüzer, M.O.; Skory, C.; Colquhoun, I.J.; Sağdıç, O.; Dertli, E. Characterization of a glucansucrase from Lactobacillus reuteri E81 and production of malto-oligosaccharides. Biocatal. Biotransform. 2019, 37, 421–430. [Google Scholar] [CrossRef]
- Cui, L.; Hu, J.; Li, C.C.; Wang, C.M.; Zhang, C.Y. An electrochemical biosensor based on the enhanced quasi-reversible redox signal of prussian blue generated by self-sacrificial label of iron metal-organic framework. Biosens. Bioelectron. 2018, 122, 168–174. [Google Scholar] [CrossRef]
- İspirli, H.; Bowman, M.J.; Skory, C.D.; Dertli, E. Synthesis and characterization of Bifidogenic raffinose-derived oligosaccharides via acceptor reactions of glucansucrase E81. LWT 2021, 147, 111525. [Google Scholar] [CrossRef]
- İspirli, H.; Dertli, E. Production of lactose derivative hetero-oligosaccharides from whey by glucansucrase E81 and determination of prebiotic functions. LWT 2021, 137, 110471. [Google Scholar] [CrossRef]
- İspirli, H.; Dertli, E. Production of mannose-containing oligosaccharides by glucansucrase E81 and determination of their functional characteristics. Biocatal. Biotransform. 2020, 38, 202–209. [Google Scholar] [CrossRef]
- Gangoiti, J.; Corwin, S.F.; Lamothe, L.M.; Vafiadi, C.; Hamaker, B.R.; Dijkhuizen, L. Synthesis of novel α-glucans with potential health benefits through controlled glucose release in the human gastrointestinal tract. Crit. Rev. Food Sci. Nutr. 2020, 60, 123–146. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.M.; Teresi, J.; Cote, G. Influence of alternansucrase-derived oligosaccharides and other carbohydrates on α-galactosidase and α-glucosidase activity in Bifidobacterium adolescentis. Lett. Appl. Microbiol. 2008, 46, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Miller-Fosmore, C.; Côté, G. Growth of various intestinal bacteria on alternansucrase-derived oligosaccharides. Lett. Appl. Microbiol. 2005, 40, 385–390. [Google Scholar] [CrossRef]
- Sanz, M.L.; Côté, G.L.; Gibson, G.R.; Rastall, R.A. Prebiotic properties of alternansucrase maltose-acceptor oligosaccharides. J. Agric. Food Chem. 2005, 53, 5911–5916. [Google Scholar] [CrossRef]
- Bivolarski, V.; Vasileva, T.; Gabriel, V.; Iliev, I. Synthesis of glucooligosaccharides with prebiotic potential by glucansucrase URE 13–300 acceptor reactions with maltose, raffinose and lactose. Eng. Life Sci. 2018, 18, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Robyt, J.F.; Eklund, S.H. Relative, quantitative effects of acceptors in the reaction of Leuconostoc mesenteroides B-512F dextransucrase. Carbohydr. Res. 1983, 121, 279–286. [Google Scholar] [CrossRef]
- Dahiya, D.K.; Puniya, M.; Shandilya, U.K.; Dhewa, T.; Kumar, N.; Kumar, S.; Puniya, A.K.; Shukla, P. Gut microbiota modulation and its relationship with obesity using prebiotic fibers and probiotics: A review. Front. Microbiol. 2017, 8, 563. [Google Scholar] [CrossRef]
- Chen, X.Y.; Gänzle, M.G. Lactose and lactose-derived oligosaccharides: More than prebiotics? Int. Dairy J. 2017, 67, 61–72. [Google Scholar] [CrossRef]
- CK Rajendran, S.R.; Okolie, C.L.; Udenigwe, C.C.; Mason, B. Structural features underlying prebiotic activity of conventional and potential prebiotic oligosaccharides in food and health. J. Food Biochem. 2017, 41, e12389. [Google Scholar] [CrossRef]
- Li, W.; Xia, X.; Tang, W.; Ji, J.; Rui, X.; Chen, X.; Jiang, M.; Zhou, J.; Zhang, Q.; Dong, M. Structural characterization and anticancer activity of cell-bound exopolysaccharide from Lactobacillus helveticus MB2-1. J. Agric. Food Chem. 2015, 63, 3454–3463. [Google Scholar] [CrossRef] [PubMed]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. Technol. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Dhanya, B.E.; Prabhu, A.; Rekha, P.D. Extraction and characterization of an exopolysaccharide from a Marine bacterium. Int. Microbiol. 2022, 25, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; Rathour, R.; Singh, R.; Thakur, I.S. Production and characterization of extracellular polymeric substances (EPS) generated by a carbofuran degrading strain Cupriavidus sp. ISTL7. Bioresour. Technol. 2019, 282, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Bejar, W.; Gabriel, V.; Amari, M.; Morel, S.; Mezghani, M.; Maguin, E.; Fontagné-Faucher, C.; Bejar, S.; Chouayekh, H. Characterization of glucansucrase and dextran from Weissella sp. TN610 with potential as safe food additives. Int. J. Biol. Macromol. 2013, 52, 125–132. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Liu, P.; Ahmed, Z.; Xiao, P.; Bai, X. Physical characterization of exopolysaccharide produced by Lactobacillus plantarum KF5 isolated from Tibet Kefir. Carbohydr. Polym. 2010, 82, 895–903. [Google Scholar] [CrossRef]
- Feng, F.; Zhou, Q.; Yang, Y.; Zhao, F.; Du, R.; Han, Y.; Xiao, H.; Zhou, Z. Characterization of highly branched dextran produced by Leuconostoc citreum B-2 from pineapple fermented product. Int. J. Biol. Macromol. 2018, 113, 45–50. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, X.; Min, W.; Yang, Z. Characterization and immunomodulatory activity of an exopolysaccharide produced by Lactobacillus plantarum JLK0142 isolated from fermented dairy tofu. Int. J. Biol. Macromol. 2018, 115, 985–993. [Google Scholar] [CrossRef]
- Tyagi, B.; Gupta, B.; Thakur, I.S. Biosorption of Cr (VI) from aqueous solution by extracellular polymeric substances (EPS) produced by Parapedobacter sp. ISTM3 strain isolated from Mawsmai cave, Meghalaya, India. Environ. Res. 2020, 191, 110064. [Google Scholar] [CrossRef]
- Zhu, J.; Tan, Z.; Zhang, Z.; Shi, X. Characterization on structure and bioactivities of an exopolysaccharide from Lactobacillus curvatus SJTUF 62116. Int. J. Biol. Macromol. 2022, 210, 504–517. [Google Scholar] [CrossRef]
- Sims, I.M.; Carnachan, S.M.; Bell, T.J.; Hinkley, S.F. Methylation analysis of polysaccharides: Technical advice. Carbohydr. Polym. 2018, 188, 1–7. [Google Scholar] [CrossRef]
- Ciucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Gerwig, G.J.; te Poele, E.M.; Dijkhuizen, L.; Kamerling, J.P. Structural analysis of rebaudioside A derivatives obtained by Lactobacillus reuteri 180 glucansucrase-catalyzed trans-alpha-glucosylation. Carbohydr. Res. 2017, 440–441, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Dertli, E.; Colquhoun, I.J.; Côté, G.L.; Le Gall, G.; Narbad, A. Structural analysis of the α-D-glucan produced by the sourdough isolate Lactobacillus brevis E25. Food Chem. 2018, 242, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Du, R.; Yu, L.; Yu, N.; Ping, W.; Song, G.; Ge, J. Characterization of exopolysaccharide produced by Levilactobacillus brevis HDE-9 and evaluation of its potential use in dairy products. Int. J. Biol. Macromol. 2022, 217, 303–311. [Google Scholar] [CrossRef]
- Zhao, D.; Cao, H.Y.; Sun, M.; Yu, L.S.; Du, R.P. Isolation, purification and structural properties analysis of exopolysaccharide from Leuconostoc pseudointestinalis HDL-3. Sci. Technol. Food Ind. 2022, 43, 115–122. [Google Scholar] [CrossRef]
- Chen, Z.; Ni, D.; Cheng, M.; Zhu, Y.; Mu, W. Comparative study of physicochemical properties of dextran and reuteran synthesised by two glucansucrases that are highly similar in amino acid sequence. Int. J. Food Sci. Technol. 2021, 56, 6674–6684. [Google Scholar] [CrossRef]
- İspirli, H.; Sagdic, O.; Yılmaz, M.T.; Dertli, E. Physicochemical characterisation of an α-glucan from Lactobacillus reuteri E81 as a potential exopolysaccharide suitable for food applications. Process Biochem. 2019, 79, 91–96. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of the 3D structure of GS. (a) Schematic representation of the U-shape fold formed by the various domains of GS. (b) Schematic diagram of the 3D structure of Ln. reuteri GTF-180N (PDB: 3HZ3) and sucrose binding site. Sucrose molecules are shown in red, residues in domain A are shown in green carbon, and residues in domain B are shown in yellow at the bottom right. (c) Schematic diagram of the 3D structure of Ln. mesenteroides NRRL B-1355 Asr (PDB: 6HVG) and Ca2+ binding sites. The green circles represent Ca2+.
Figure 1.
Schematic diagram of the 3D structure of GS. (a) Schematic representation of the U-shape fold formed by the various domains of GS. (b) Schematic diagram of the 3D structure of Ln. reuteri GTF-180N (PDB: 3HZ3) and sucrose binding site. Sucrose molecules are shown in red, residues in domain A are shown in green carbon, and residues in domain B are shown in yellow at the bottom right. (c) Schematic diagram of the 3D structure of Ln. mesenteroides NRRL B-1355 Asr (PDB: 6HVG) and Ca2+ binding sites. The green circles represent Ca2+.

Figure 2.
General reaction mechanism for GH70 glucansucrases; D is the nucleophilic asparte residue; E is the general acid/base residue [27].
Figure 2.
General reaction mechanism for GH70 glucansucrases; D is the nucleophilic asparte residue; E is the general acid/base residue [27].

Figure 3.
GSs catalytic reaction mechanism.

Figure 4.
The main receptor of dextransucrase.

Figure 5.
Related applications of oligosaccharides.

Figure 6.
Common detection methods of EPSs, as well as the related properties of oligosaccharides.


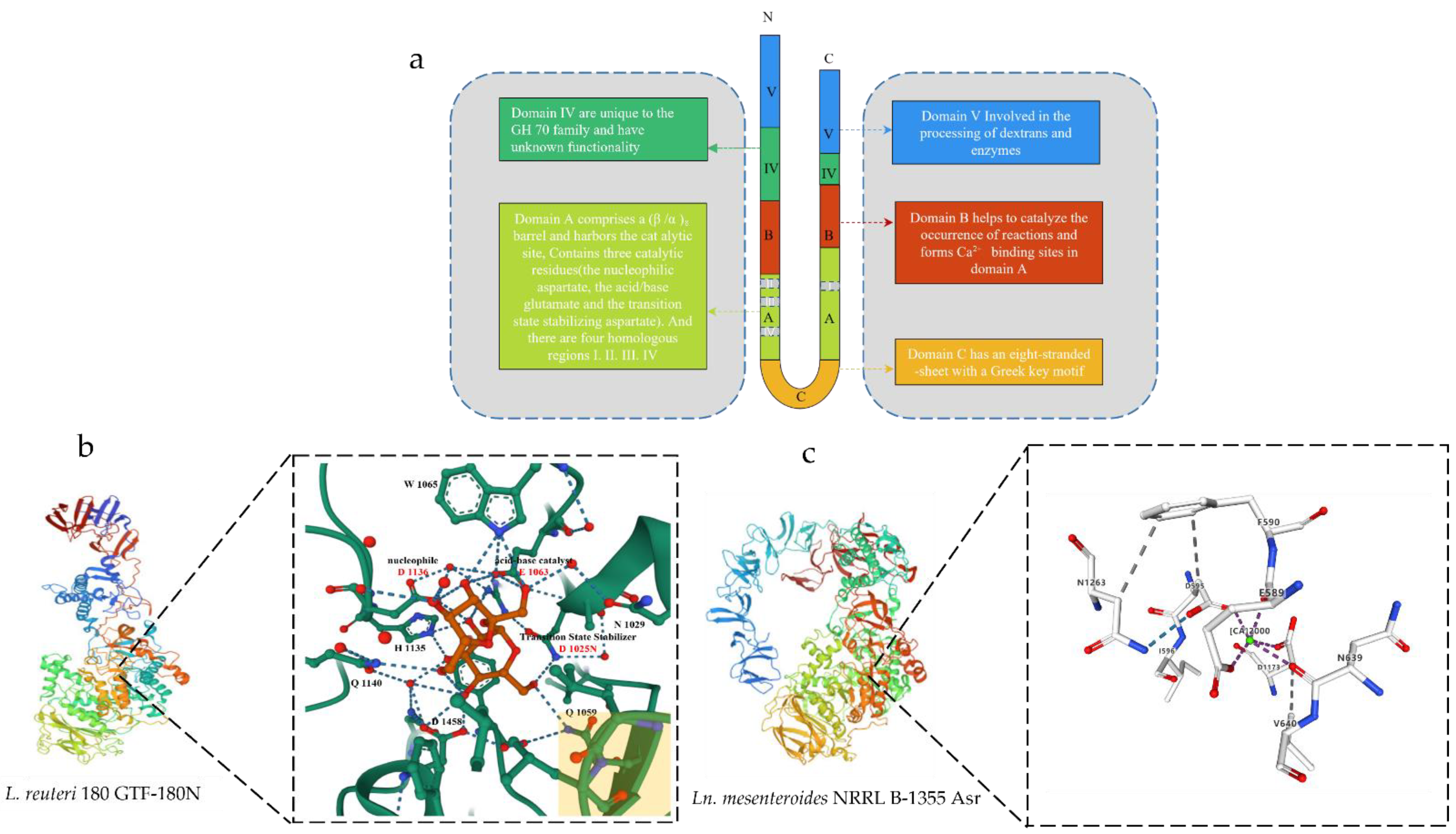{kind=link}
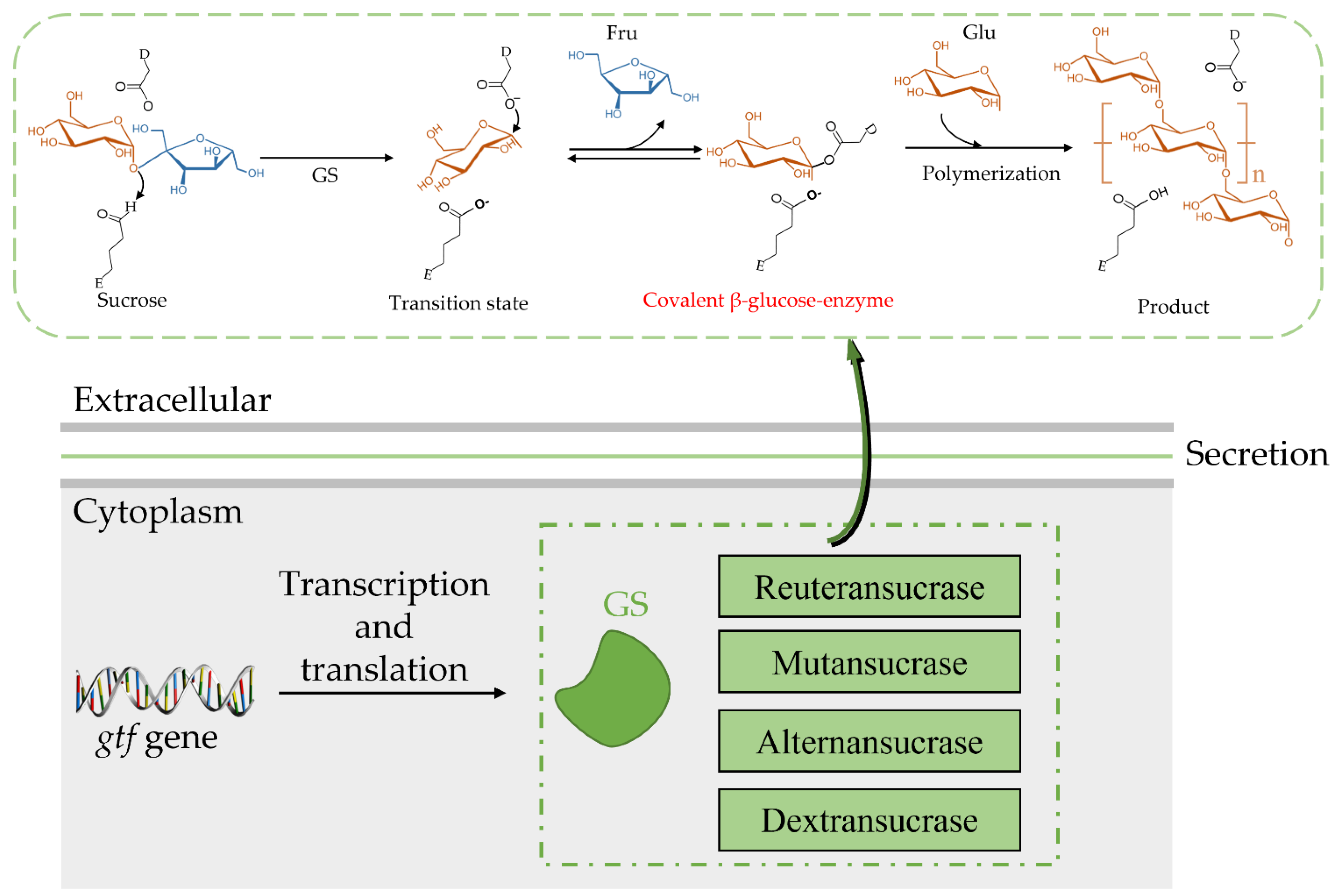{kind=link}
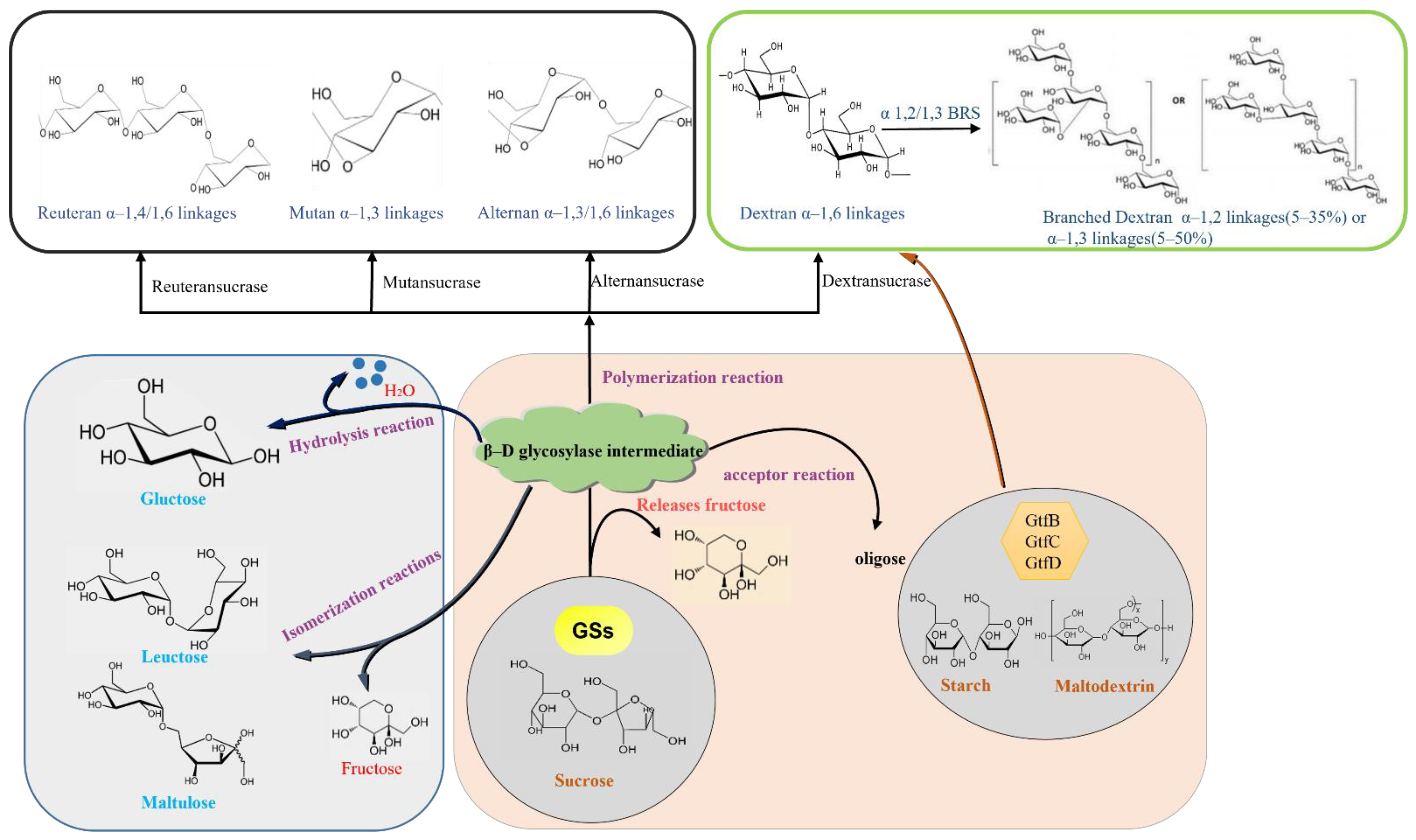{kind=link}
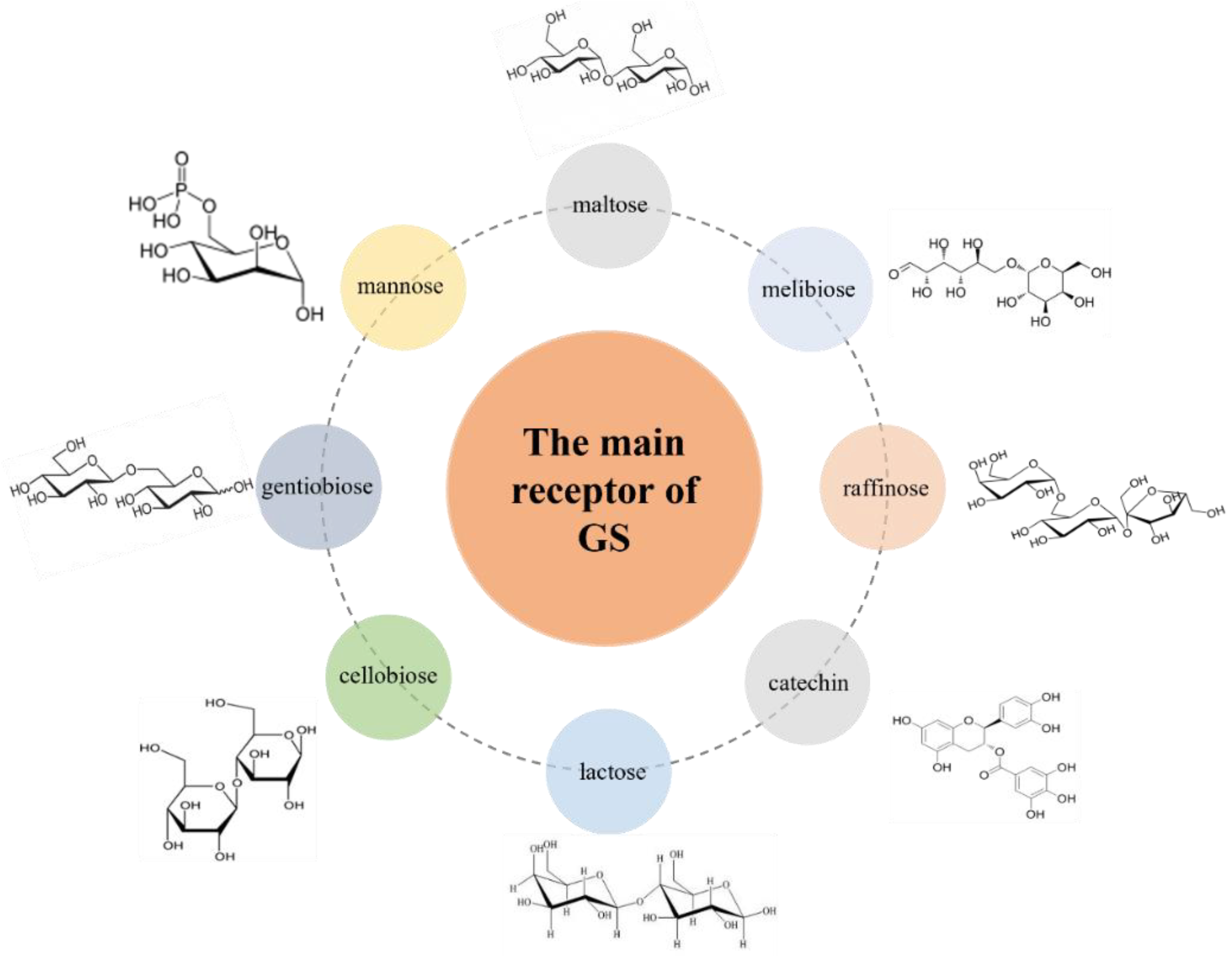{kind=link}
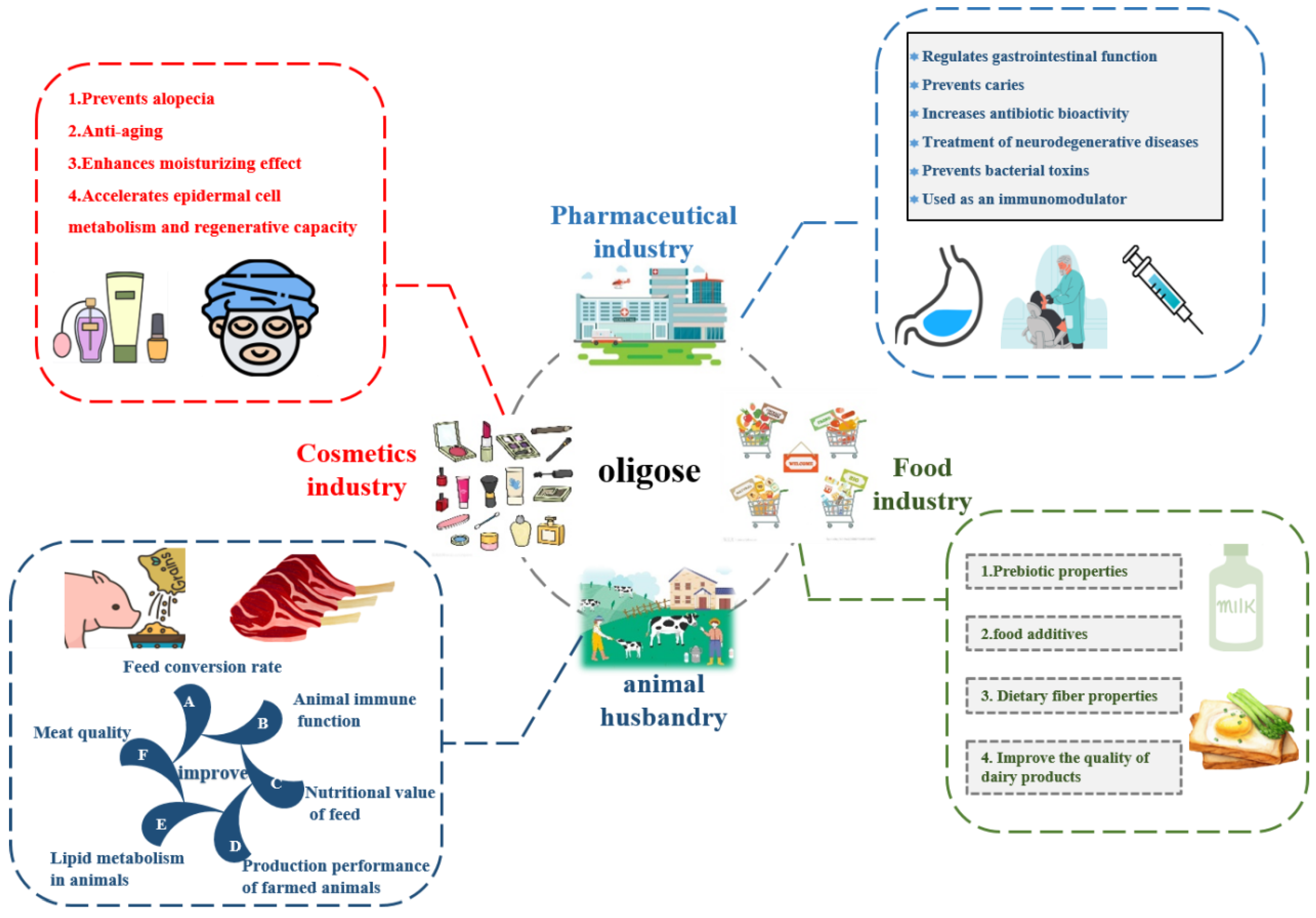{kind=link}
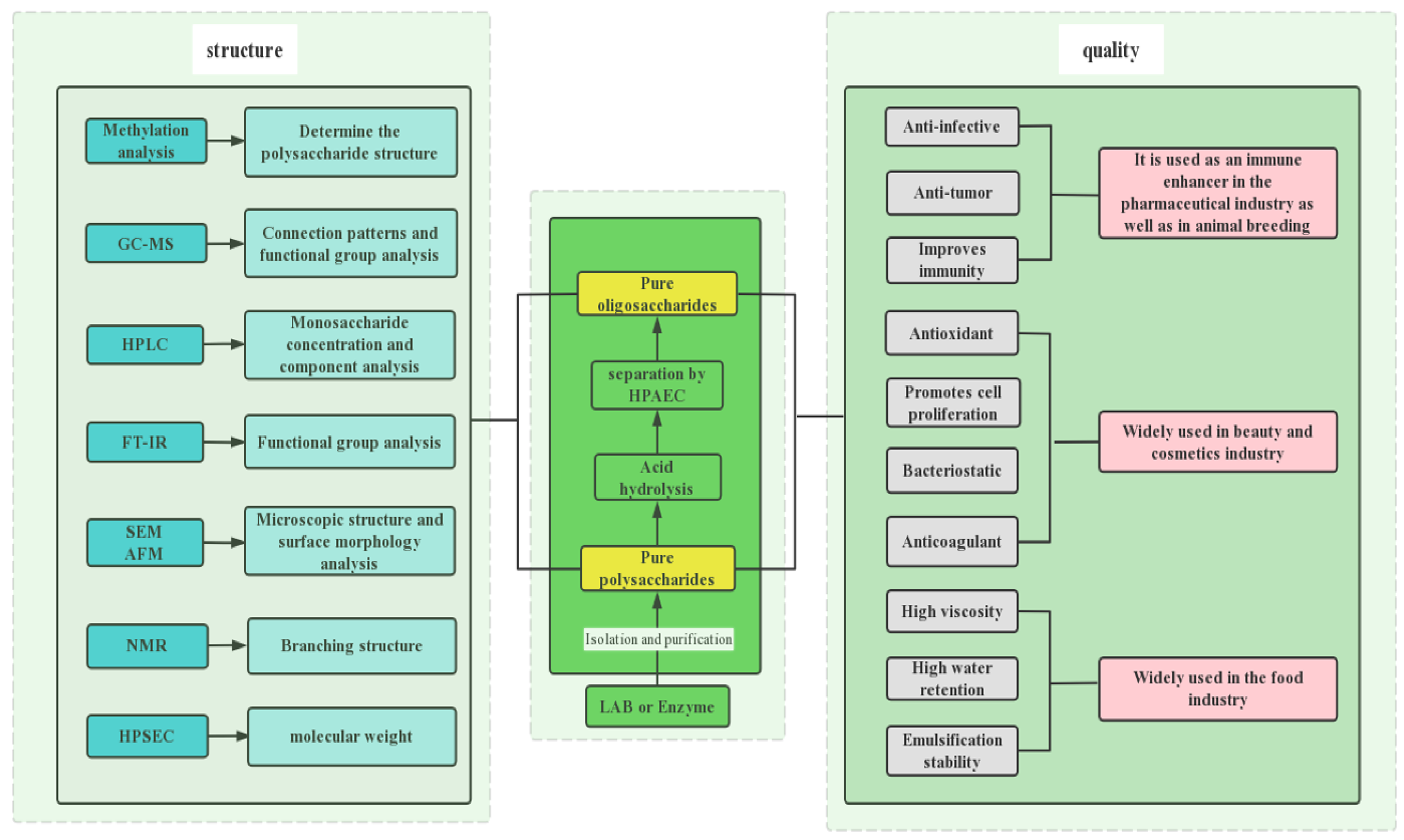{kind=link}
Table 1.
Information on GS from different strains and their related properties in recent years was analyzed and collated using the CAZy database.
Table 1.
Information on GS from different strains and their related properties in recent years was analyzed and collated using the CAZy database.
| Enzyme | MASS | Organism | Genebank | Length | Reference |
|---|---|---|---|---|---|
| Gtf1624 | 183 kDa | Latilactobacillus curvatus TMW 1.624 | CCK33643.1 | 1697 aa | [57] |
| DSR-F | 170 kDa | Ln. citreum B/110-1-2 | ACY92456.2 | 1527 aa | [58] |
| LcDS | 165 kDa | Ln. citreum HJ-P4 | BAF96719.1 | 1477 aa | [59] |
| DexT | 167 kDa | Ln citreum KM20 | ACA83218.1 | 1495 aa | [60] |
| DSR-A | 145 kDa | Ln. citreum NRRL B-1299 | CDX67012.1 | 1290 aa | [61] |
| DSR-B | 168 kDa | Ln. citreum NRRL B-1299 | AAB95453.1 | 1508 aa | [62] |
| DSR-E | 313 kDa | Ln. citreum NRRL B-1299 | CDX66820.1 | 2836 aa | [63] |
| DSR-DP | - | Ln. citreum NRRL B-1299 | CDX66641.1 | 1278 aa | [63] |
| DSR-M | 144 kDa | Ln. citreum NRRL B-1299 | CDX66895.1 | 1293 aa | [61] |
| DexYG | 170 kDa | Ln. mesenteroides 0326 | ABC75033.1 | 1527 aa | [64] |
| DsrBCB4 | 168 kDa | Ln. mesenteroides B-1299CB4 | ABF85832.1 | 1505 aa | [65] |
| DsrC | 165 kDa | Ln. mesenteroides B-1355 | CAB76565.1 | 1477 aa | [66] |
| Dsrb74 | 169 kDa | Ln. mesenteroides B-742CB | AAG38021.1 | 1508 aa | [67] |
| DsrP | 161 kDa | Ln. mesenteroides IBT-PQ | AAS79426.1 | 1454 aa | [68] |
| DsrN | 169 kDa | Ln. mesenteroides KIBGE-IB-22 | AFP53921.1 | 1527 aa | [69] |
| DsrX | 169 kDa | Ln. mesenteroides L0309 | AAQ98615.2 | 1522 aa | [70] |
| DsrD | 169 kDa | Ln. mesenteroides LCC4 | AAG61158.1 | 1527 aa | [50] |
| DSR-S | 169 kDa | Ln. mesenteroides NRRL B-512F | AAD10952.1 | 1527 aa | [71] |
| DSR-T | 110 kDa | Ln. mesenteroides NRRL B-512F | BAA90527.1 | 1016 aa | [72] |
| Gtf1971 | 178 kDa | Ligilactobacillus animalis TMW 1.971 | CCK33644.1 | 1585 aa | [57] |
| Gtf106A | 199 kDa | L. reuteri TMW 1.106 | ABP88726.1 | 1782 aa | [57] |
| GTF-S | 151 kDa | Streptococcus downei MFE 28 | AAA26898.1 | 1365 aa | [73] |
| GTF-U | 176 kDa | Streptococcus sobrinus | BAA14241.1 | 1592 aa | [74] |
| GTF-B | 166 kDa | Streptococcus mutans GS 5 | AAA88588.1 | 1476 aa | [75] |
| GTF-D | 163 kDa | S. mutans GS 5 | AAA26895.1 | 1462 aa | [76] |
| DsrK39 | 158 kDa | Weissella cibaria LBAE-K39 | ADB43097.3 | 1445 aa | [77] |
| WcCab3-DSR | 154 kDa | Weissella confusa Cab3 | AKE50934.1 | 1401 aa | [78] |
| DSR-C39-2 | 155 kDa | W. confusa LBAE C39-2 | CCF30682.1 | 1412 aa | [54] |
| DSR | 156 kDa | W. confusa VTT E-90392 | AHU88292.1 | 1418 aa | [79] |
| LcALT | 229 kDa | Ln. citreum ABK-1 | AIM52834.1 | 2057 aa | [80] |
| GtfB-SK2 | - | Ln. citreum SK24.002 | - | - | [81] |
| ASR | 229 kDa | Ln. mesenteroides NRRL B-1355 | CAB65910.2 | 2057 aa | [82] |
| GtfO | 197 kDa | L. reuteri ATCC 55730 | AAY86923.1 | 1781 aa | [83] |
| Gtf-SK3 | - | L. reuteri SK24.003 | - | - | [84] |
| GtfML1 | - | L. reuteri ML1 | - | - | [34] |
| DSRI | - | Ln. mesenteroides NRRL B-1118 | - | - | [85] |
| GtfB | - | S. mutans GS5 | - | - | [86] |
| GtfC | - | S. mutans GS5 | - | - | [87] |
| GTF-Kg15 | 174 kDa | Latilactobacillus sakei KG15 | AAU08011.1 | 1595 aa | [34] |
| GTF-33 | 172 kDa | Lentilactobacillus parabuchneri 33 | AAU08006.1 | 1561 aa | [34] |
| - | 164 kDa | Leuconostoc lactis EG001 | ACT20911.1 | 1500 aa | [56] |
| GTF-Kg3 | 161 kDa | Limosilactobacillus fermentum KG3 | AAU08008.1 | 1463 aa | [34] |
| GtfB | 179 kDa | L. f ermentum NCC2970 | AOR73699.1 | 1593 aa | [88] |
| GtfB | 179 kDa | L. reuteri 121 | AAU08014.2 | 1619 aa | [89] |
| GtfML1 | 196 kDa | L. reuteri ML1 | AAU08004.1 | 1772 aa | [34] |
| GtfML4 | 180 kDa | L. reuteri ML1 | AAU08003.2 | 1620 aa | [90] |
| GtfB | 196 kDa | L. reuteri NCC2613 | ASA47879.1 | 1662 aa | [91] |
| GtfC | 163 kDa | S. mutans | BAA26114.1 | 1455 aa | [89] |
| DsrwC | 162 kDa | W. cibaria CMU | ACK38203.1 | 1472 aa | [92] |
| GtfD | 87 kDa | Azotobacter chroococcum NCIMB 8003 | AJE22990.1 | 780 aa | [93] |
| GtfC | 99 kDa | Exiguobacterium sibiricum 255-15 | ACB62096.1 | 893 aa | [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, L.; Qian, Z.; Ge, J.; Du, R. Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications. Fermentation 2022, 8, 629. https://doi.org/10.3390/fermentation8110629
AMA Style
Yu L, Qian Z, Ge J, Du R. Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications. Fermentation. 2022; 8(11):629. https://doi.org/10.3390/fermentation8110629
Chicago/Turabian StyleYu, Liansheng, Zhigang Qian, Jingping Ge, and Renpeng Du. 2022. "Glucansucrase Produced by Lactic Acid Bacteria: Structure, Properties, and Applications" Fermentation 8, no. 11: 629. https://doi.org/10.3390/fermentation8110629
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.