
Aerial Seeding: An Effective Forest Restoration Method in Highly Degraded Forest Landscapes of Sub-Tropic Regions

Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Description of Sample Plots

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Age Range (Years) | Average Age (a) | Average DBH (cm) | Average Height (m) | Average BA (m2·ha−1) | Average Stand Density (N·ha−1) |
|---|---|---|---|---|---|---|
| AS | 11–20 | 18 ± 0.3 | 5.4 ± 0.12 | 6.3 ± 0.88 | 5.2 ± 0.66 | 2267 ± 311.0 |
| 21–30 | 24 ± 1.9 | 10.7 ± 1.10 | 12.2 ± 0.87 | 11.9 ± 0.33 | 1408 ± 283.0 | |
| 31–40 | 35 ± 1.2 | 13.4 ± 1.00 | 12.3 ± 0.87 | 17.9 ± 1.54 | 1329 ± 285.4 | |
| 41–50 | 41 ± 0.3 | 16.6 ± 0.32 | 16.6 ± 2.19 | 25.1 ± 4.12 | 1152 ± 168.8 | |
| NR | 11–20 | 18 ± 0.7 | 9.0 ± 0.70 | 9.6 ± 0.93 | 11.3 ± 1.41 | 1808 ± 227.1 |
| 21–30 | 23 ± 0.9 | 11.7 ± 2.16 | 10.9 ± 3.08 | 15.9 ± 4.20 | 1575 ± 343.9 | |
| 31–40 | 36 ± 2.0 | 13.6 ± 0.70 | 12.0 ± 1.00 | 17.6 ± 2.10 | 1221 ± 145.8 | |
| 41–50 | 44 ± 1.2 | 16.5 ± 1.95 | 16.1 ± 0.46 | 20.52 ± 2.57 | 1054 ± 281.6 |
2.3. Tree Biomass
2.4. Understory and Forest Floor Litter
2.5. Soil Sampling
2.6. Carbon Stock Calculations
2.7. Statistical Analysis
3. Results
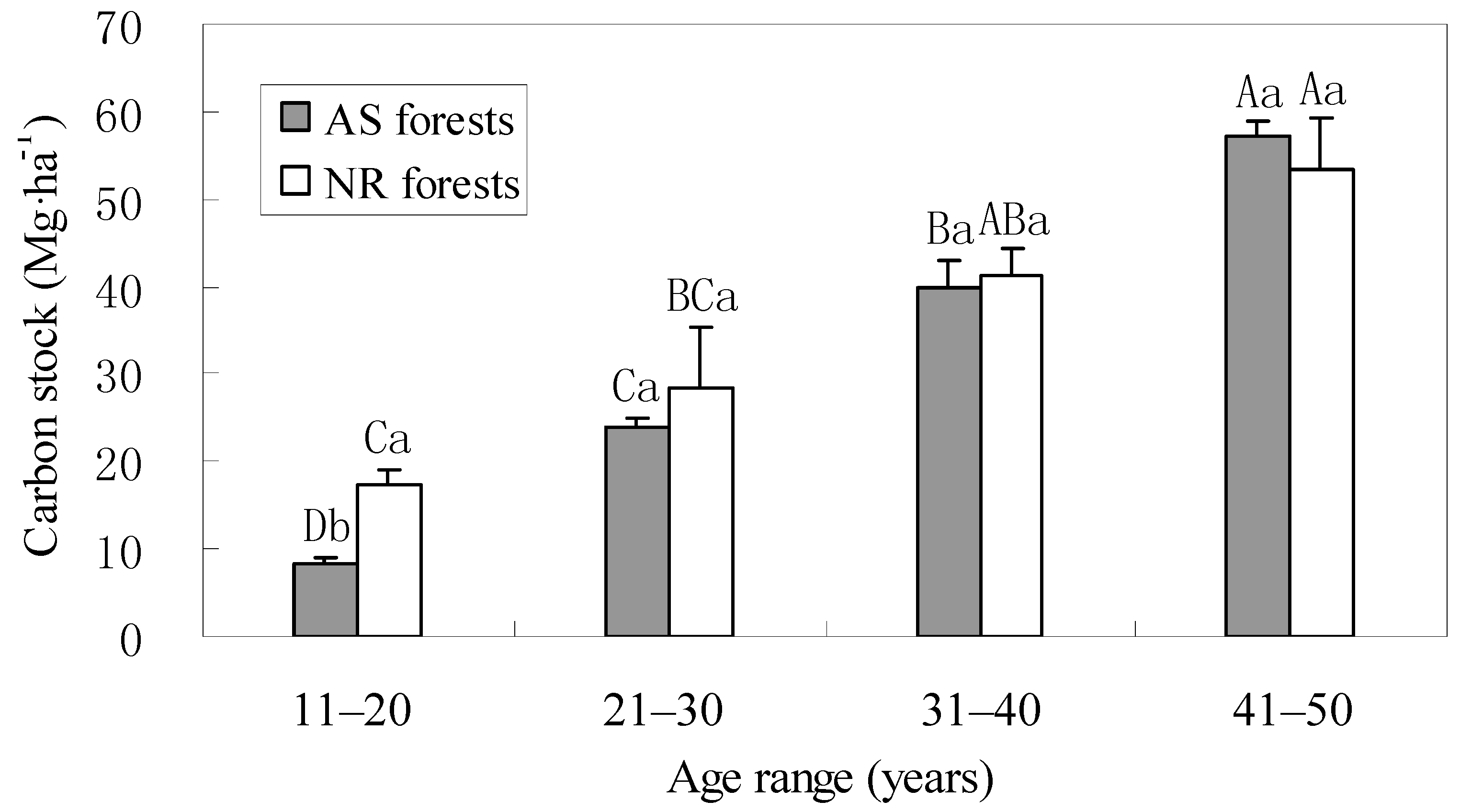
3.1. Carbon Stocks in Trees
| Age Range | Stem | Branch | Leaf | Root | ||||
|---|---|---|---|---|---|---|---|---|
| AS | NR | AS | NR | AS | NR | AS | NR | |
| 11–20 | 3.9 ± 0.34Aa | 9.4 ± 1.04Ab | 2.1 ± 0.27Cb | 3.9 ± 0.50Ca | 1.3 ± 0.97Ca | 2.0 ± 0.29Aa | 0.9 ± 0.15Db | 1.9 ± 0.23Ca |
| (48.0) | (54.8) | (25.4) | (22.2) | (15.5) | (11.7) | (11.1) | (11.1) | |
| 21–30 | 14.2 ± 0.79Bb | 17.8 ± 5.08BCa | 4.7 ± 0.11Ba | 5.1 ± 0.80BCa | 2.1 ± 0.12BCa | 2.4 ± 0.22Aa | 3.1 ± 0.10Ca | 3.3 ± 0.93BCa |
| (58.9) | (62.2) | (19.7) | (18.0) | (8.7) | (8.4) | (12.8) | (11.4) | |
| 31–40 | 24.2 ± 2.14Cb | 26.5 ± 2.03Ca | 6.8 ± 0.94Ba | 6.8 ± 0.68ABa | 2.7 ± 0.43ABa | 2.6 ± 0.35Aa | 5.3 ± 0.46Ba | 5.3 ± 0.67ABa |
| (62.0) | (64.3) | (17.4) | (16.5) | (7.0) | (6.3) | (13.6) | (12.9) | |
| 41–50 | 35.1 ± 2.26Db | 35.4 ± 4.05Ca | 10.1 ± 0.97Aa | 8.3 ± 1.05Aa | 4.1 ± 0.69Aa | 3.0 ± 0.48Aa | 7.8 ± 0.28Aa | 6.8 ± 0.88Aa |
| (61.51) | (66.2) | (17.6) | (15.6) | (7.2) | (5.6) | (13.7) | (12.7) | |
| Mean | 19.4 ± 3.55a | 22.3 ± 3.05a | 5.9 ± 0.93a | 6.0 ± 0.58a | 2.6 ± 0.36a | 2.5 ± 0.18a | 4.3 ± 0.78a | 4.3 ± 0.61a |
| (60.3) | (63.4) | (18.4) | (17.2) | (8.0) | (7.1) | (13.3) | (12.3) | |

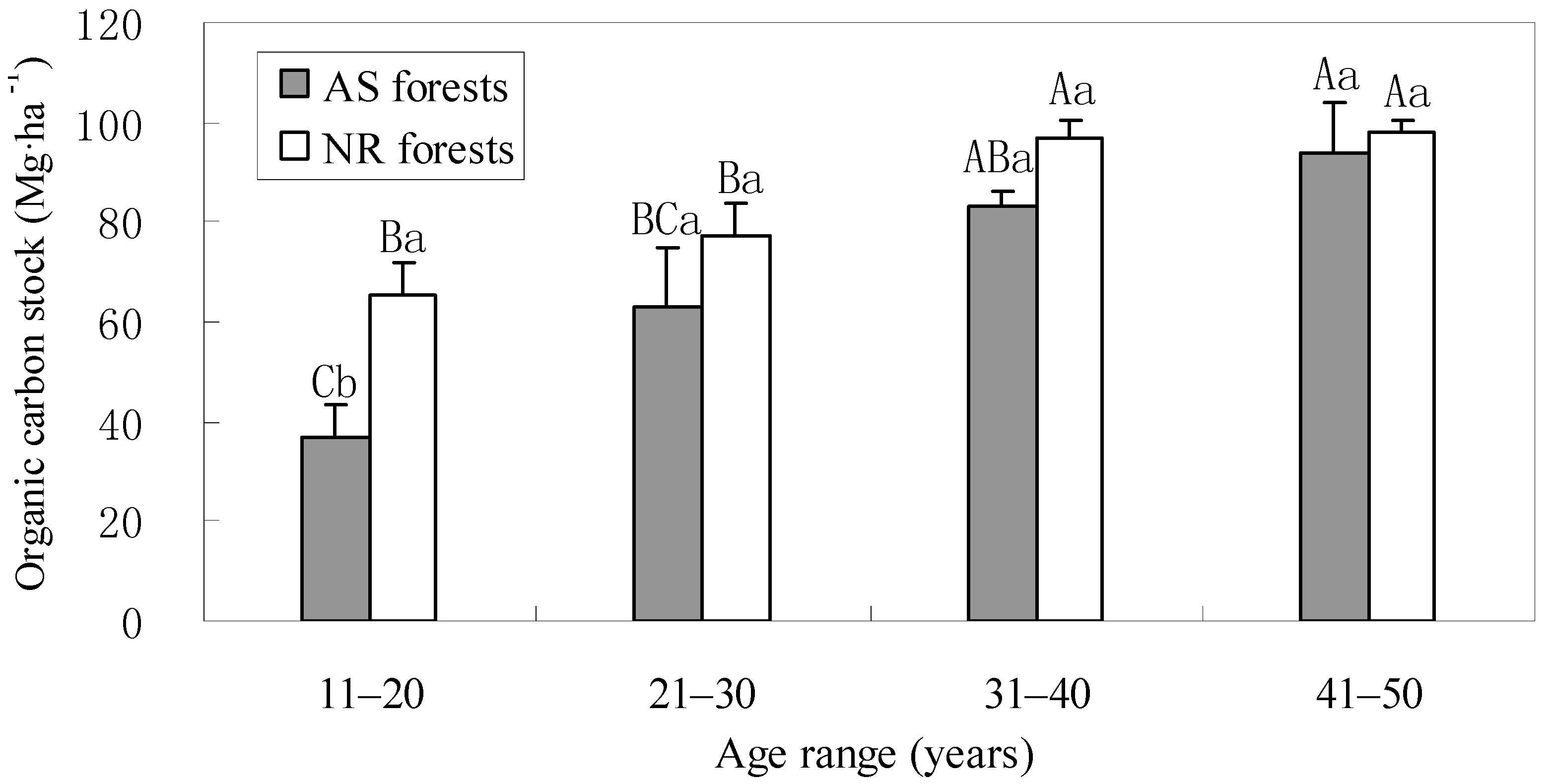
3.2. Carbon Stocks in the Understory and Litter
| Type | Age Range | Mean | ||||
|---|---|---|---|---|---|---|
| 11–20 | 21–30 | 31–40 | 41–50 | |||
| Understory | AS | 3.1 ± 0.33Aa | 4.4 ± 1.00Aa | 3.1 ± 1.13Aa | 4.0 ± 1.07Aa | 3.7 ± 0.52a |
| NR | 8.6 ± 1.85Ab | 6.8 ± 2.76Aa | 5.0 ± 1.70Aa | 5.8 ± 1.98Aa | 6.6 ± 1.11b | |
| Litter | AS | 0.4 ± 0.09Aa | 0.6 ± 0.13Aa | 0.6 ± 0.19Aa | 0.5 ± 0.05Aa | 0.5 ± 0.10a |
| NR | 0.9 ± 0.14Ab | 1.1 ± 0.17Aa | 0.9 ± 0.09Aa | 1.0 ± 0.30Aa | 1.0 ± 0.08b | |
3.3. Carbon Stocks in Soil

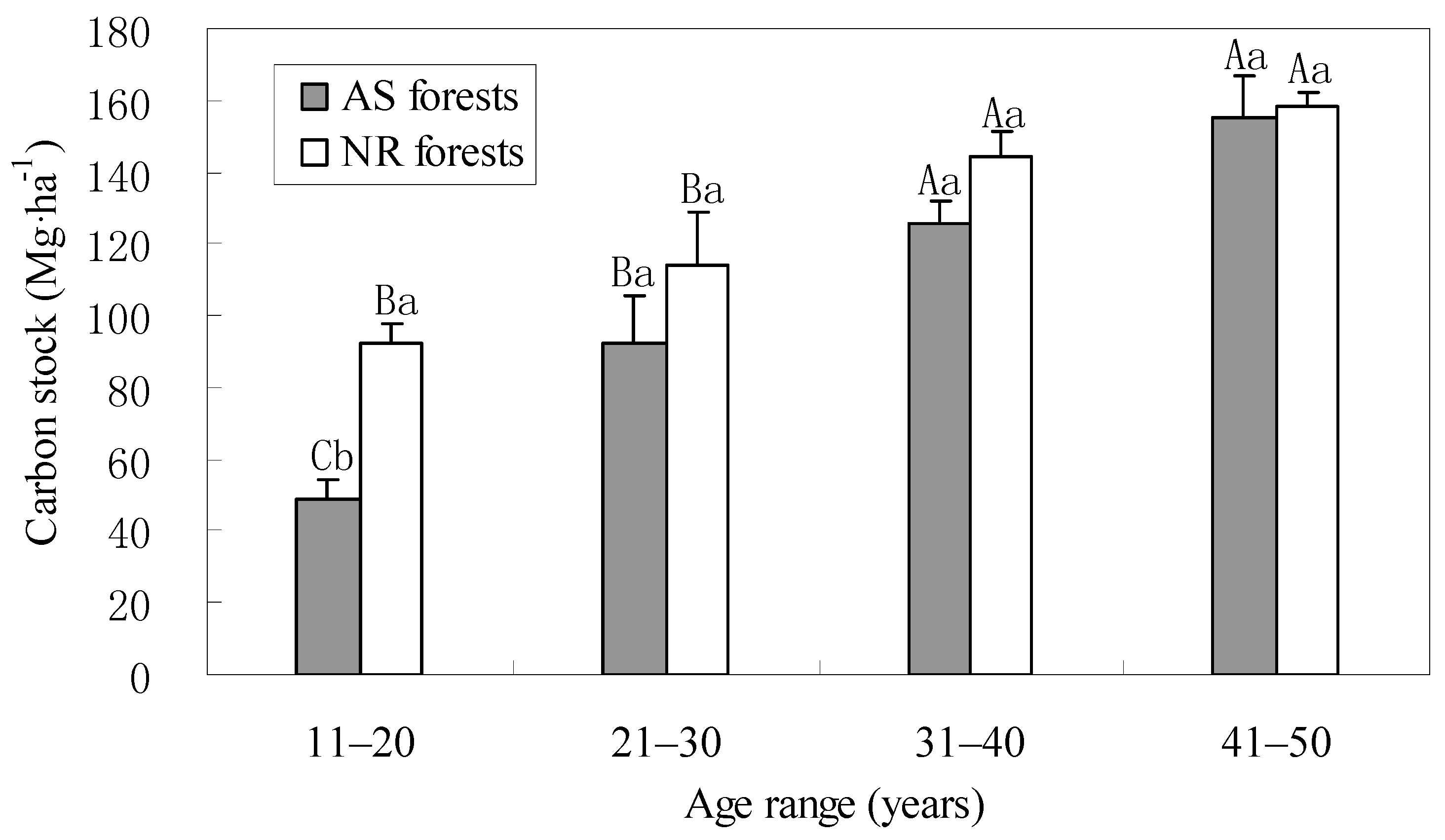
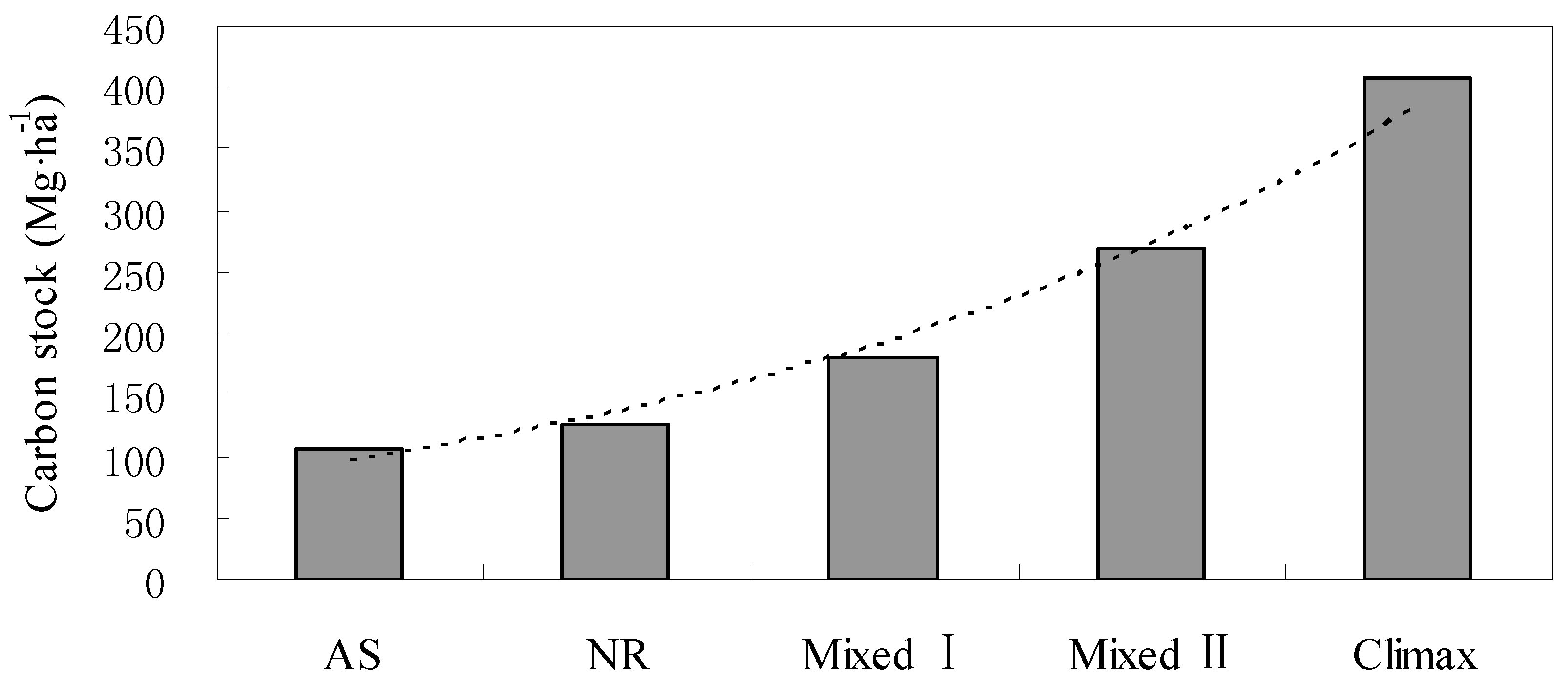
3.4. Total Carbon Stocks

4. Discussion

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, W.H. Degradation and restoration of forest ecosystems in China. For. Ecol. Manag. 2004, 201, 33–41. [Google Scholar] [CrossRef]
- Corbera, E.; Estrada, M.; Brown, K. Reducing greenhouse gas emissions from deforestation and forest degradation in developing countries: Revisiting the assumptions. Clim. Chang. 2010, 100, 355–388. [Google Scholar] [CrossRef]
- Dulal, H.B.; Brodnig, G.; Shah, K.U. Capital assets and institutional constraints to implementation of greenhouse gas mitigation options in agriculture. Mitig. Adapt. Strateg. Glob. Chang. 2011, 16, 1–23. [Google Scholar] [CrossRef]
- Upadhyay, T.P.; Sankhayan, P.L.; Solberg, B. A review of carbon sequestration dynamics in the Himalayan region as a function of land-use change and forest/soil degradation with special reference to Nepal Agriculture. Ecosyst. Environ. 2005, 105, 449–465. [Google Scholar] [CrossRef]
- Guo, L.B.; Gifford, R.M. Soil carbon stocks and land use change: A meta analysis. Glob. Chang. Boil. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Li, G.L.; Liu, Y.; Guo, B.; Xu, Y.; Zhang, K.D.; Zhao, S.R. Advances in the research of the aerial-seeding forest in China. World For. Res. 2006, 19, 45–48. (In Chinese) [Google Scholar]
- Liu, J.; Li, S.; Ouyang, Z.; Tam, C.; Chen, X. Ecological and socioeconomic effects of China’s policies for ecosystem services. Proc. Natl. Acad. Sci. USA 2008, 105, 9477–9482. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.H. Developing Low-carbon Forestry for Low-carbon Economy. World For. Res. 2010, 23, 1–6. (In Chinese) [Google Scholar]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystem. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- The National Forestry Development State Foresty Administration Office. China Forestry Development Division; China Forestry Publishing House: Beijing, China, 2011; pp. 62–92. (In Chinese) [Google Scholar]
- Zhai, M.P. Concept for vegetation rehabilitation and construction in the North-west Region of China. Sci. Soil Water Conserv. 2003, 1, 60–63. (In Chinese) [Google Scholar]
- Tripathi, K.P.; Singh, B. Species diversity and vegetation structure across various strata in natural and plantation forests in Katerniaghat Wildlife Sanctuary, North India. Trop. Ecol. 2009, 50, 191–200. [Google Scholar]
- Shono, K.; Cadaweng, E.A.; Durst, P.B. Application of assisted natural regeneration to restore degraded tropical forestlands. Restor. Ecol. 2007, 15, 620–626. [Google Scholar] [CrossRef]
- Aide, T.M.; Zimmerman, J.K.; Herrera, L.; Rosario, M.; Serrano, M. Forest recovery in abandoned tropical pastures in Puerto Rico. For. Ecol. Manag. 1995, 77, 77–86. [Google Scholar] [CrossRef]
- Orians, G.H.; Pfeiffer, E.W. Ecological effects of the war in Vietnam. Science 1970, 168, 544–554. [Google Scholar] [CrossRef] [PubMed]
- The National Forestry Bureau. China Forestry Development Report 2013; China Forestry Publishing House: Beijing, China, 2013. (In Chinese) [Google Scholar]
- Beyers, J.L. Postfire Seeding for Erosion Control: Effectiveness and impacts on native plant communities. Conserv. Biol. 2004, 18, 947–956. [Google Scholar] [CrossRef]
- Groen, A.H.; Woods, S.W. Effectiveness of aerial seeding and straw mulch for reducing post-wildfire erosion, north-western Montana, USA. Int. J. Wildland Fire 2008, 17, 559–571. [Google Scholar] [CrossRef]
- Greipsson, S.; El-Mayas, H. Large-scale reclamation of barren lands in Iceland by aerial seeding. Land Degrad. Dev. 1999, 10, 185–193. [Google Scholar] [CrossRef]
- Pyke, D.A.; Wirth, T.A.; Beyers, J.L. Does seeding after wildfires in rangelands reduce erosion or invasive species? Restor. Ecol. 2013, 21, 415–421. [Google Scholar] [CrossRef]
- Davies, K.W.; Bates, J.D.; Madsen, M.D.; Nafus, A.M. Restoration of mountain big sagebrush steppe following prescribed burning to control western Juniper. Environ. Manag. 2014, 53, 1015–1022. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, C. Carbon density and distribution of six Chinese temperate forests. Sci. China Life Sci. 2010, 53, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Woodwell, G.M.; Whittaker, R.H.; Reiners, W.A.; Likens, G.E.; Delwiche, C.C.; Botkin, D.B. The biota and the world carbon budget. Science 1978, 199, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Post, W.M.; Emanuel, W.R.; Zinke, P.J.; Stangenberger, A.G. Soil pools and world life zone. Nature 1982, 298, 156–159. [Google Scholar] [CrossRef]
- Cao, M.; Ian Woodward, F. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change. Nature 1998, 393, 249–252. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; del Campo, A.D.; Ceacero, C.J.; Quero, J.L.; de Mena, J.H. On the importance of topography, site quality, stock quality and planting date in a semiarid plantation: Feasibility of using low-density LiDAR. Ecol. Eng. 2014, 67, 25–38. [Google Scholar] [CrossRef]
- Muukkonen, P. Generalized allometric volume and biomass equations for some tree species in Europe. Eur. J. For. Res. 2007, 126, 157–166. [Google Scholar] [CrossRef]
- Pearson, T.; Walker, S.M.; Brown, S. Sourcebook for Land Use, Land-Use Change and Forestry Projects; BioCarbon Fund, Winrock International: Washington, DC, USA, 2005; p. 64. [Google Scholar]
- Rodriguez-Murillo, J.C. Organic carbon content under different types of land use and soil in peninsular Spain. Biol. Fertil. Soils 2001, 33, 53–61. [Google Scholar] [CrossRef]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Wang, B.; Wei, W.J. Carbon storage and density of forests in Jiangxi province. Jiangxi Sci. 2007, 25, 681–687. (In Chinese) [Google Scholar]
- Chen, G.S.; Yang, Y.S.; Xie, J.S.; Guo, J.F.; Gao, R.; Qian, W. Conversion of a natural broad-leafed evergreen forest into pure plantation forests in a subtropical area: Effects on carbon storage. Ann. For. Sci. 2005, 62, 659–668. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Chen, Z.X.; He, Y.J.; Bai, F.M.; Zhang, J.H.; Li, Z.H. Effects of stand density on the biomass and productivity of Pinus massoniana air sowing stands. J. Cent. South For. Univ. 2001, 21, 44–47. (In Chinese) [Google Scholar]
- Pan, P.; Lyu, D.; Ouyang, X.Z.; Wang, X.T. A study on biomass and carbon storage of natural Pinus massoniana forest at different stand growing stages in central Jiangxi Province. Acta Agric. Univ. Jiangxiensis 2014, 36, 131–136. (In Chinese) [Google Scholar]
- Lu, Q.; Zhao, T.S.; Luo, T.X.; Jin, L.Y.; Yin, S.J. Comparative study on biodiversity and stand growth regulation in Pinus taiwanensis from different origin. For. Res. 1996, 9, 273–277. (In Chinese) [Google Scholar]
- Liu, C.H. Community characteristics and growth of natural forest and plantation of Cyclobalanopsis chungii. J. Zhejiang For. Coll. 2005, 22, 56–60. (In Chinese) [Google Scholar]
- Jordan, C.F.; Farnworth, E.G. Natural vs. plantation forests: A case study of land reclamation strategies for the humid tropics. Environ. Manag. 1982, 6, 485–492. [Google Scholar] [CrossRef]
- Zhang, S.W.; Zhao, Y.H.; Hou, M.M.; Xi, L. Comparative study for undergrowth vegetation biomass of Eucalyptus Plantations and natural forest. Resour. Dev. Mark. 2008, 24, 385–387. (In Chinese) [Google Scholar]
- Riegel, G.M.; Miller, R.F.; Krueger, W.C. The effects of aboveground and belowground competition on understory speciescomposition in a Pinus ponderosa forest. For. Sci. 1995, 41, 864–889. [Google Scholar]
- Melillo, J.M.; Aber, J.D.; Linkins, A.E.; Ricca, A.; Fry, B.; Nadelhoffer, K.J. Carbon and nitrogen dynamics along the decay continuum: Plant litter to soil organic matter. Plant Soil 1989, 115, 189–198. [Google Scholar] [CrossRef]
- Carrera, A.L.; Bertiller, M.B. Relationships among plant litter, fine roots, and soil organic C and N across an aridity gradient in Northern Patagonia, Argentina. Ecoscience 2010, 17, 276–286. [Google Scholar] [CrossRef]
- Bassett, O.D.; Prior, L.D.; Slijkerman, C.M.; Jamieson, D.; Bowman, D.M.J.S. Aerial sowing stopped the loss of alpine ash (Eucalyptus delegatensis) forests burnt by three short-interval fires in the Alpine National Park, Victoria, Australia. For. Ecol. Manag. 2015, 342, 39–48. [Google Scholar] [CrossRef]
- Gong, C.; Wang, S.L.; Zeng, Z.Q.; Deng, S.J.; Chen, J.P.; Long, K.S. Carbon storage and its distribution pattern of evergreen broad-leaved forests at different succession stages in mid-subtropical China. Chin. J. Ecol. 2011, 30, 1935–1941. (In Chinese) [Google Scholar]
- Fang, Y.T.; Mo, J.M.; Peng, S.L.; Li, D.J. Role of forest succession on carbon sequestration of forest ecosystems in lower subtropical China. Acta Ecol. Sin. 2003, 23, 1685–1694. (In Chinese) [Google Scholar]
- Cameron, A.D. Building resilience into Sitka spruce (Picea sitchensis (Bong.) Carr.) forests in Scotland in response to the threat of climate change. Forests 2015, 6, 398–415. [Google Scholar] [CrossRef]
- Pretzsch, H. The elasticity of growth in pure and mixed stands of Norway spruce (Picea Abies (L.) Karst.) and common beech (Fagus sylvatica L.). J. For. Sci. 2003, 49, 491–501. [Google Scholar]
- Ma, Q.Y.; Chen, X.L.; Wang, J.; Lin, C.; Kang, F.F.; Cao, W.Q.; Ma, Z.B.; Li, W.Y. Carbon content rate in constructive species of main forest types in northern China. J. Beijing For. Univ. 2002, 24, 96–100. (In Chinese) [Google Scholar]
- Liu, G.H.; Fu, B.J.; Fang, J.Y. Carbon dynamics of Chinese forests and its contribution to global carbon balance. Acta Ecol. Sin. 2000, 20, 733–740. (In Chinese) [Google Scholar]
- Houghton, R.A.; Skole, D.L.; Nobre, C.A.; Hackler, J.L.; Lawrence, K.T.; Chomentowski, W.H. Annual fluxes of carbon from deforestation and regrowth in the Brazilian Amazon. Nature 2000, 403, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Fedrigo, M.; Kasel, S.; Bennett, L.T.; Roxburgh, S.H.; Nitschke, C.R. Carbon stocks in temperate forests of south-eastern Australia reflect large tree distribution and edaphic conditions. For. Ecol. Manag. 2014, 334, 129–143. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Guan, D.S.; Zhang, H.D. Characteristics of storage and decomposition of coarse woody debris (CWD) under three forests in Guangzhou. Acta Ecol. Sin. 2009, 29, 5227–5236. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; Wei, X.; Liu, Y.; Ouyang, X.; Li, Q.; Ning, J. Aerial Seeding: An Effective Forest Restoration Method in Highly Degraded Forest Landscapes of Sub-Tropic Regions. Forests 2015, 6, 1748-1762. https://doi.org/10.3390/f6061748
Xiao X, Wei X, Liu Y, Ouyang X, Li Q, Ning J. Aerial Seeding: An Effective Forest Restoration Method in Highly Degraded Forest Landscapes of Sub-Tropic Regions. Forests. 2015; 6(6):1748-1762. https://doi.org/10.3390/f6061748
Chicago/Turabian StyleXiao, Xin, Xiaohua Wei, Yuanqiu Liu, Xunzhi Ouyang, Qinglin Li, and Jinkui Ning. 2015. "Aerial Seeding: An Effective Forest Restoration Method in Highly Degraded Forest Landscapes of Sub-Tropic Regions" Forests 6, no. 6: 1748-1762. https://doi.org/10.3390/f6061748