
Allometric Equations for Estimating Biomass of Euterpe precatoria, the Most Abundant Palm Species in the Amazon

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Euterpe precatoria
2.3. Data Collection
2.4. Models and Statistics
3. Results and Discussion
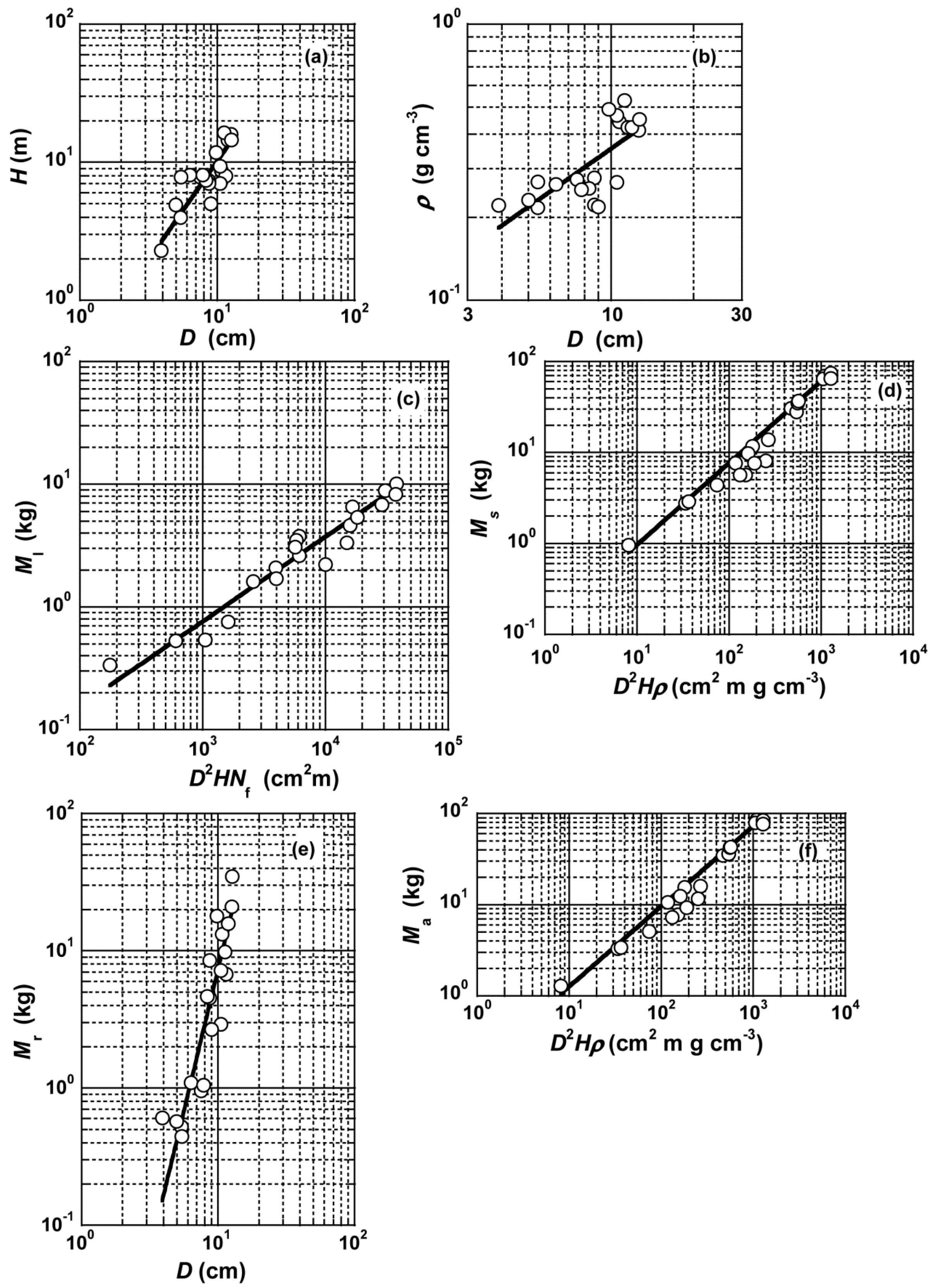
3.1. Stem Height and Specific Gravity

{kind=link}
{kind=link}
| y | x | Lng | h | Radj2 | ||
|---|---|---|---|---|---|---|
| H | D | −0.97 | (−1.86–−0.08) | 1.43 | (1.07–1.90) | 0.658 |
| ρ | D | −0.36 | (−0.60–0.12) | 0.32 | (0.23–0.45) | 0.515 |
| Ml | D2HNf | −5.04 | (−5.80–−4.28) | 0.69 | (0.61–0.78) | 0.937 |
| Ms | D2Hρ | −2.11 | (−2.43–−1.79) | 0.90 | (0.84–0.96) | 0.982 |
| Mr | D2Hρ | −3.97 | (−5.21–−2.72) | 1.01 | (0.81–1.27) | 0.787 |
| Ma | D2Hρ | −1.79 | (−2.10–−1.48) | 0.88 | (0.83–0.94) | 0.983 |

3.2. Allometric Models for Estimating Leaf Mass
3.3. Allometric Models for Estimating Stem Mass
3.4. Allometric Models for Estimating Root Mass

3.5. Allometric Models for Estimating Above-Ground Mass
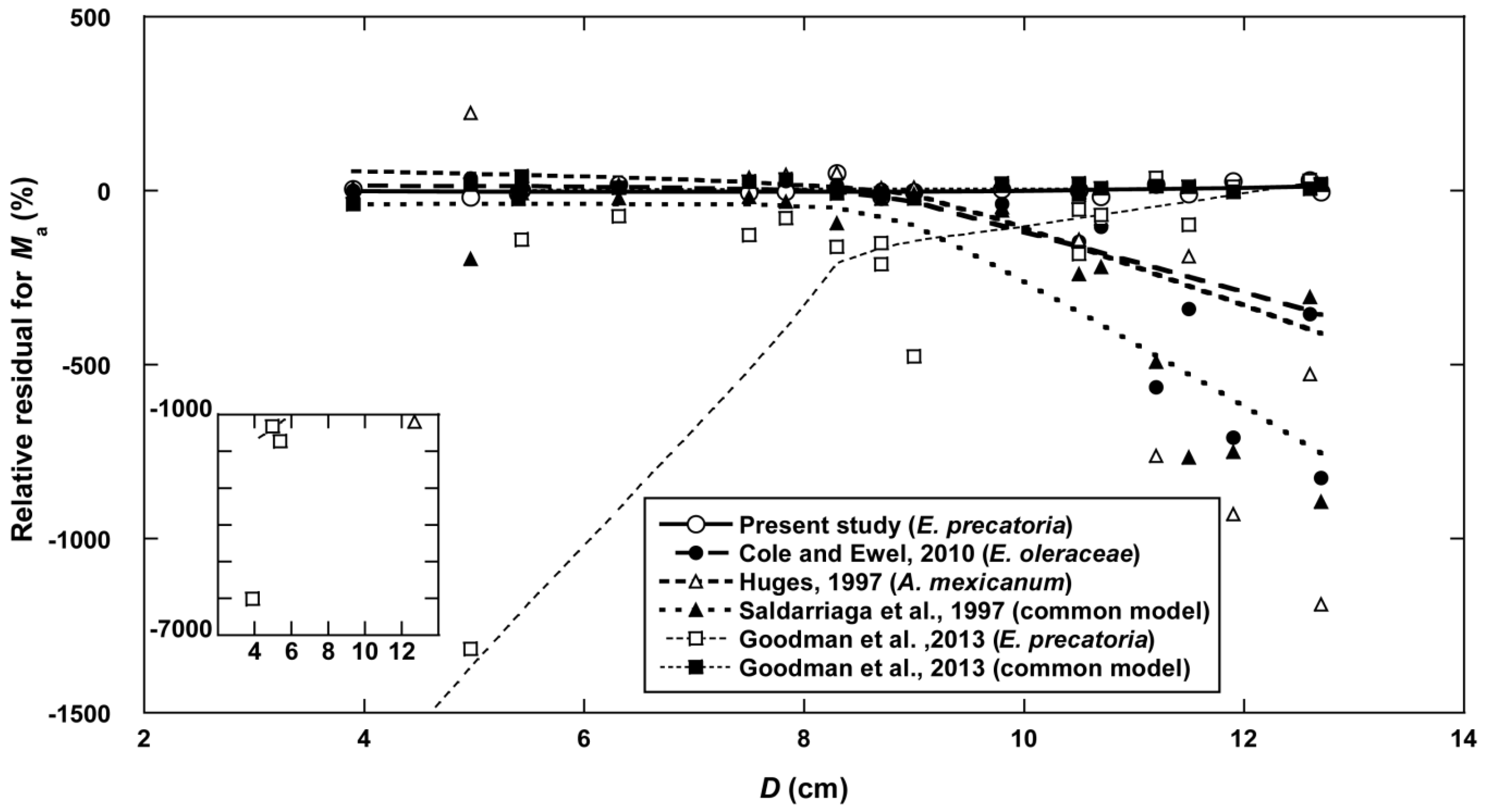
3.6. Comparison of Models for Estimating Above-Ground Mass in Palm Species
| Equations for Estimating Ma | Focal Species | RSS kg2 | References |
|---|---|---|---|
| Ma = Ms + Ml + Mrachis, | E. oleraceae | 5823 | Cole and Ewel [16] * |
| where Ms = 0.0314 × (D2H)0.917 × 1.040 | |||
| Ml = 0.0237 × (D2HNf)0.512 × 1.036 | |||
| Mrachis = 0.0458 × (D2HNf)0.388 × 1.036 | |||
| Ma = 0.3060 × D1.837 × 1.035 | A. inexicanwn | 10,642 | Hughes [13] |
| Ma = 1.697 × 10−3 × D1.754 × H2.151 | Common | 7606 | Saldarriaga et al. [9] |
| Ma = 13.59H − 108.8 | E. precatoria | 236,389 | Goodman et al. [12] |
| Ma = 0.0950 (dmf D2H) | Common | 486 | Goodman et al. [12] ** |
| Ma = 0.167 × (D2Hρ) 0.883 | E. precatoria | 397 | The present study |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chambers, J.Q.; dos Santos, J.; Ribeiro, R.J.; Higuchi, N. Tree damage, allometric relationships, and above-ground net primary production in central Amazon forest. For. Ecol. Manag. 2001, 152, 73–84. [Google Scholar] [CrossRef]
- Houghton, R.A. Aboveground forest biomass and the global carbon balance. Glob. Chang. Biol. 2005, 11, 945–958. [Google Scholar] [CrossRef]
- Lima, J.N.A.; Suwa, R.; Ribeiro, H.P.G.M.; Kajimoto, T.; Santos, J.; Silva, P.R.; Souza, S.A.C.; Barros, C.P.; Noguchi, H.; Ishizuka, M.; et al. Allometric models for estimating above- and below-ground biomass of tropical rainforests at São Gabriel da Cachoeira in upper Rio Negro, Brazilian Amazon. For. Ecol. Manag. 2012, 277, 163–172. [Google Scholar] [CrossRef]
- Kunert, N.; Barros, P.; Higuchi, N. Do palm water use characteristics explain the spatial distribution of palms in the Central Amazon? Acta Hortic. 2013, 991, 197–204. [Google Scholar]
- Fearnside, P.M. Biomassa das florestas Amazônicas brasileiras. In Emissão × Seqüestro de CO2: Uma Oportunidade de Negócios Para o Brasil; Companhia Vale do Rio Doce: Rio de Janeiro, Brazil, 1994; pp. 95–124. [Google Scholar]
- Henderson, A. The Palms of the Amazon; Oxford University Press: Oxford, UK, 1995; p. 380. [Google Scholar]
- Castilho, C.V.; Magnusson, W.E.; de Araujo, R.N.O.; Luizao, R.C.C.; Luizao, F.J.; Lima, A.P.; Higuchi, N. Variation in aboveground tree live biomass in a central Amazonian Forest: Effects of soil and topography. For. Ecol. Manag. 2006, 234, 85–96. [Google Scholar] [CrossRef]
- Laurance, W.F.; Fearnside, P.M.; Laurance, S.G.; Delamonica, P.; Lovejoy, T.E.; Rankin-de-Merona, J.M.; Chambers, J.; Gascon, C. Relationship between soils and Amazon forest biomass: A landscape-scale study. For. Ecol. Manag. 1999, 118, 127–138. [Google Scholar] [CrossRef]
- Saldarriaga, J.G.; West, D.C.; Tharp, M.L.; Uhl, C. Long-term chronosequence of forest succession in the upper Rio Negro of Colombia and Venezuela. J. Ecol. 1988, 76, 938–958. [Google Scholar] [CrossRef]
- Cummings, D.L.; Kauffman, J.B.; Perry, D.A.; Hughes, R.F. Aboveground biomass and structure of rainforests in the southwestern Brazilian Amazon. For. Ecol. Manag. 2002, 163, 293–307. [Google Scholar] [CrossRef]
- Avalos, G.; Sylvester, O. Allometric estimation of total leaf area in the neotropical palm Euterpe oleracea at La Selva, Costa Rica. Trees 2010, 24, 969–974. [Google Scholar] [CrossRef]
- Goodman, C.R.; Phillips, L.O.; del Castillo, T.D.; Freitas, L.; Cortese, T.S.; Monteagudo, A.; Baker, R.T. Amazon palm biomass and allometry. For. Ecol. Manag. 2013, 310, 994–1004. [Google Scholar] [CrossRef]
- Hughes, R.F. Effects of Deforestation and Land Use on Biomass, Carbon and Nutrient Polls in the Los Tuxtlas Region, Mexico. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1997. [Google Scholar]
- Rich, P.M. Mechanical architecture of arborescent rain forest palms. Principes 1986, 30, 117–131. [Google Scholar]
- Ter Steege, H.; Nigel, C.A.; Sabatier, D.; Baraloto, C.; Salomao, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342. [Google Scholar] [CrossRef]
- Cole, T.G.; Ewel, J.J. Allometric equations for four valuable tropical tree species. For. Ecol. Manag. 2006, 229, 351–360. [Google Scholar] [CrossRef]
- Rich, P.M. Mechanical structure of the stem of arborescent palms. Bot. Gaz. 1987, 148, 42–50. [Google Scholar] [CrossRef]
- Kunert, N.; Laboratory for Forest Management, Manaus, Brazil. Micrometeorological Tower Measurements from the ZF-2 site at km 32, Digital Media, Large-Scale Biosphere—Atmosphere Experiment in Amazonia INPA Manaus. Unpublished data. 2013. [Google Scholar]
- Bravard, S.; Righi, D. Geochemical differences in an Oxisol—Spodosol toposequence of Amazonia, Brazil. Geoderma 1989, 44, 29–42. [Google Scholar] [CrossRef]
- Ferraz, J.; Ohta, S.; Salles, P.C. Distribuição dos Solos ao Longo de Dois Transectos em Floresta Primária ao Norte de INPA, Manaus, AM. In Pesquisas Florestais para Conservação da Floresta e Reabilitação de Áreas Degradadas da Amazônia; Higuchi, N., Campos, M.A.A., Sampaio, P.T.B., dos Santos, J., Eds.; CT-INPA/JICA: Manaus, Brazil, 1998; pp. 111–143. [Google Scholar]
- Lorenzi, H.; Bacher, L.; Lacerida, M.; Sartori, S. Frutas Brasileiras e Exóticas Cultivadas (de Consumo in Natura); Instituto Plantarum de Estudos das Flora: Nova Odessa, Brazil, 2006; p. 81. [Google Scholar]
- Avalos, G.; Otárola, M.F. Allometry and stilt root structure of the neotropical palm Euterpe precatoria (Arecaceae) across sites and successional stages. Am. J. Bot. 2010, 97, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Avalos, G.; Schneider, A.R. Quantification of ramet production in the neotropical palm Euterpe precatoria (Areacaceae) in Costa Rica. Ecotropica 2011, 17, 31–38. [Google Scholar]
- Clay, J.W.; Clement, C.R. Selected Species and Strategies to Enhance Income Generation from Amazonian Forests; Forestry Working Paper FO: Misc/93/6; FAO: Rome, Italy, 1993. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2014. Available online: http://www.R-project.org/ (accessed on 10 April 2014).
- McMahon, T.A.; Kronauer, R.E. Tree structures: Deducing the principle of mechanical design. J. Theor. Biol. 1976, 59, 443–466. [Google Scholar] [CrossRef] [PubMed]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Brown, A.C.S.; Cairns, M.A.; Cahmbers, J.Q.; Eamus, D.; Fölster, H.; Fromad, F.; Higuchi, N.; Kira, T.; Lescure, J.P.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.F.; Martins, F.R.; Santos, F.A.M. Allometry of a neotropical palm, Euterpe edulis Mart. Acta Bot. Bras. 2004, 18, 369–374. [Google Scholar] [CrossRef]
- Cairns, M.A.; Brown, S.; Helmer, E.H.; Baumgardner, G.A. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef]
- Houghton, R.A.; Laurence, K.T.; Hackers, J.L.; Brown, S. The spatial distribution of forest biomass in Brazilian Amazon: A comparison of estimates. Glob. Chang. Biol. 2001, 7, 731–746. [Google Scholar] [CrossRef]
- Brown, S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Malhi, Y.; Wood, D.; Baker, T.R.; Wright, J.; Phillips, O. Regional variation of above-ground live biomass in old-growth Amazonian forests. Glob. Chang. Biol. 2006, 12, 1–32. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Nogueira, E.M.; Fearnside, P.M.; Nelson, B.W.; Barbosa, R.I.; Keizer, E.W.H. Estimates of forest biomass in the Brazilian Amazon: New allometric equations and adjustments to biomass from wood-volume inventories. For. Ecol. Manag. 2008, 256, 1853–1867. [Google Scholar] [CrossRef]
- Avalos, G.; Salazar, D.; Araya, L.A. Stilt root structure in the neotropical palms Iriartea deltoidea and Socratea exorrhiza. Biotropica 2005, 37, 44–53. [Google Scholar] [CrossRef]
- Khan, M.N.I.; Suwa, R.; Hagihara, A. Carbon and nitrogen pools in a mangrove stand of Kandelia obovata (S., L.) Yong: Vertical distribution in the soil-vegetation system. Wetl. Ecol. Manag. 2007, 15, 141–153. [Google Scholar] [CrossRef]
- Zianis, D.; Mencuccini, M. On simplifying allometric analysis of forest biomass. For. Ecol. Manag. 2004, 187, 311–332. [Google Scholar] [CrossRef]
- Todd, M.C. The Role of Nutrient Cycling in the Sustainability of Young Plant Communities on Mined Sites. Ph.D. Thesis, University of Western Australia, Perth, WA, Australia, 2000. [Google Scholar]
- Simpson, W.T. Specific Gravity, Moisture Content, and Density Relationship for Wood; General Technical Report, FPL-GTR-76; USDA Forest Service, Forest Products Laboratory: Madison, WI, USA, 1993. [Google Scholar]
- Colgan, S.M.; Swemmer, T.; Asner, P.G. Structural relationships between form factor, wood density, and biomass in African savanna woodlands. Trees 2014, 28, 91–102. [Google Scholar] [CrossRef]
- Botânico, M.P.; Angyalossy, V. Is the secondary thickening in palms always diffuse? An. Acad. Bras. Cienc. 2014, 85, 1461–1472. [Google Scholar] [CrossRef]
- Tomlinson, P.B. The uniqueness of palms. Bot. J. Linn. Soc. 2006, 151, 5–14. [Google Scholar] [CrossRef]
- Rich, P.M. Developmental anatomy of the stem of Welfia georgii, Iriartea gigantea, and other arborescent palms—Implications for mechanical support. Am. J. Bot. 1987, 74, 792–802. [Google Scholar] [CrossRef]
- Brown, S. Estimating Biomass and Biomass Change of Tropical Forests: A Primer; FAO Forestry Paper 134; FAO: Rome, Italy; p. 55.
- Ngomanda, A.; Obiang, L.E.N.; Lebamba, J.; Mavouroulou, M.Q.; Gomat, H.; Mankou, S.G.; Loumeto, J.; Iponga, M.D.; Ditsouga, K.F.; Koumba, Z.R.; et al. Site-specific versus pantropical allometric equations: Which option to estimate the biomass of a moist central African forest? For. Ecol. Manag. 2014, 312, 1–9. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Silva, F.; Suwa, R.; Kajimoto, T.; Ishizuka, M.; Higuchi, N.; Kunert, N. Allometric Equations for Estimating Biomass of Euterpe precatoria, the Most Abundant Palm Species in the Amazon. Forests 2015, 6, 450-463. https://doi.org/10.3390/f6020450
Da Silva F, Suwa R, Kajimoto T, Ishizuka M, Higuchi N, Kunert N. Allometric Equations for Estimating Biomass of Euterpe precatoria, the Most Abundant Palm Species in the Amazon. Forests. 2015; 6(2):450-463. https://doi.org/10.3390/f6020450
Chicago/Turabian StyleDa Silva, Fernando, Rempei Suwa, Takuya Kajimoto, Moriyoshi Ishizuka, Niro Higuchi, and Norbert Kunert. 2015. "Allometric Equations for Estimating Biomass of Euterpe precatoria, the Most Abundant Palm Species in the Amazon" Forests 6, no. 2: 450-463. https://doi.org/10.3390/f6020450