
Wood Modification as a Tool to Understand Moisture in Wood


1
Bioresource Chemistry & Technology, Department of Geosciences and Natural Resource Management, University of Copenhagen, Rolighedsvej 23, DK-1958 Frederiksberg C, Denmark
2
Building Materials, Department of Building and Environmental Technology, Lund University, P.O. Box 118, SE-22100 Lund, Sweden
*
Author to whom correspondence should be addressed.
Forests 2021, 12(3), 372; https://doi.org/10.3390/f12030372
Submission received: 16 February 2021
/
Revised: 16 March 2021
/
Accepted: 18 March 2021
/
Published: 20 March 2021
(This article belongs to the Special Issue Wood Modification: Physical Properties and Biological Efficacy)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Moisture plays a central role in the performance of wood products because it affects important material properties such as the resistance to decomposition, the mechanical properties, and the dimensions. To improve wood performance, a wide range of wood modification techniques that alter the wood chemistry in various ways have been described in the literature. Typically, these modifications aim to improve resistance to decomposition, dimensional stability, or, to introduce novel functionalities in the wood. However, wood modification techniques can also be an important tool to improve our understanding of the interactions between wood and moisture. In this review, we describe current knowledge gaps in our understanding of moisture in wood and how modification has been and could be used to clarify some of these gaps. This review shows that introducing specific chemical changes, and even controlling the distribution of these, in combination with the variety of experimental methods available for characterization of moisture in wood, could give novel insights into the interaction between moisture and wood. Such insights could further contribute to applications in several related fields of research such as how to enhance the resistance to decomposition, how to improve the performance of moisture-induced wooden actuators, or how to improve the utilization of wood biomass with challenging swelling anisotropy.
Keywords:
moisture; wood; modification; sorption; equilibrium; swelling; hysteresis; kinetics; diffusion; cell walls1. Introduction
Wood is a hygroscopic material that can take up water molecules from its surroundings. This moisture within the material plays a central role in the performance of wooden products because it affects important material characteristics such as resistance to decomposition [1,2], the mechanical properties [3,4], and the dimensions [3,5]. Hence, predicting how moisture affects the performance of wood is relevant for important applications of wood in society, such as the service life of wooden building components. Alteration of the wood chemistry, i.e., wood modification, can be used to enhance the performance of wood, for instance, by increasing the resistance to decomposition or dimensional stability [6,7]. Key to both of these effects is how modification reduces the moisture content [7,8,9], although the exact mechanisms behind the improved resistance to decomposition are still unknown. A wide range of wood modification techniques have been described in the literature, with various alterations of the wood chemistry. Many of these modifications are aimed at enhancing specific material properties [10,11], while others introduce novel functionalities to the wood material, e.g., magnetic properties [12] or stimuli-responsiveness to pH or temperature [13].
In this review, we focus on how wood modification has been and could be used to increase our fundamental understanding of the interactions between wood and moisture. First, we briefly describe the current state of knowledge on fundamental aspects of moisture in wood and highlight knowledge gaps, organized into four different themes. Second, we point out ways in which wood modifications could be used as a tool within each of these four themes to clarify some of these knowledge gaps. The aim is, thus, not an exhaustive compilation of all of the literature covering the effects of modification on wood-moisture relations or to describe different modification processes, because such reviews already exist; see, for example, [14,15,16,17]. Instead, we cover a focused selection of literature with the aim to inspire future research in the important field of wood-moisture research in order to advance the state-of-the-art about moisture in wood. With a better knowledge about moisture in wood and the effects of wood chemistry, it would be possible to develop and optimize modification processes and chemistries to further enhance the performance of wood.
2. Moisture in Wood: A Brief Overview of Knowledge Gaps
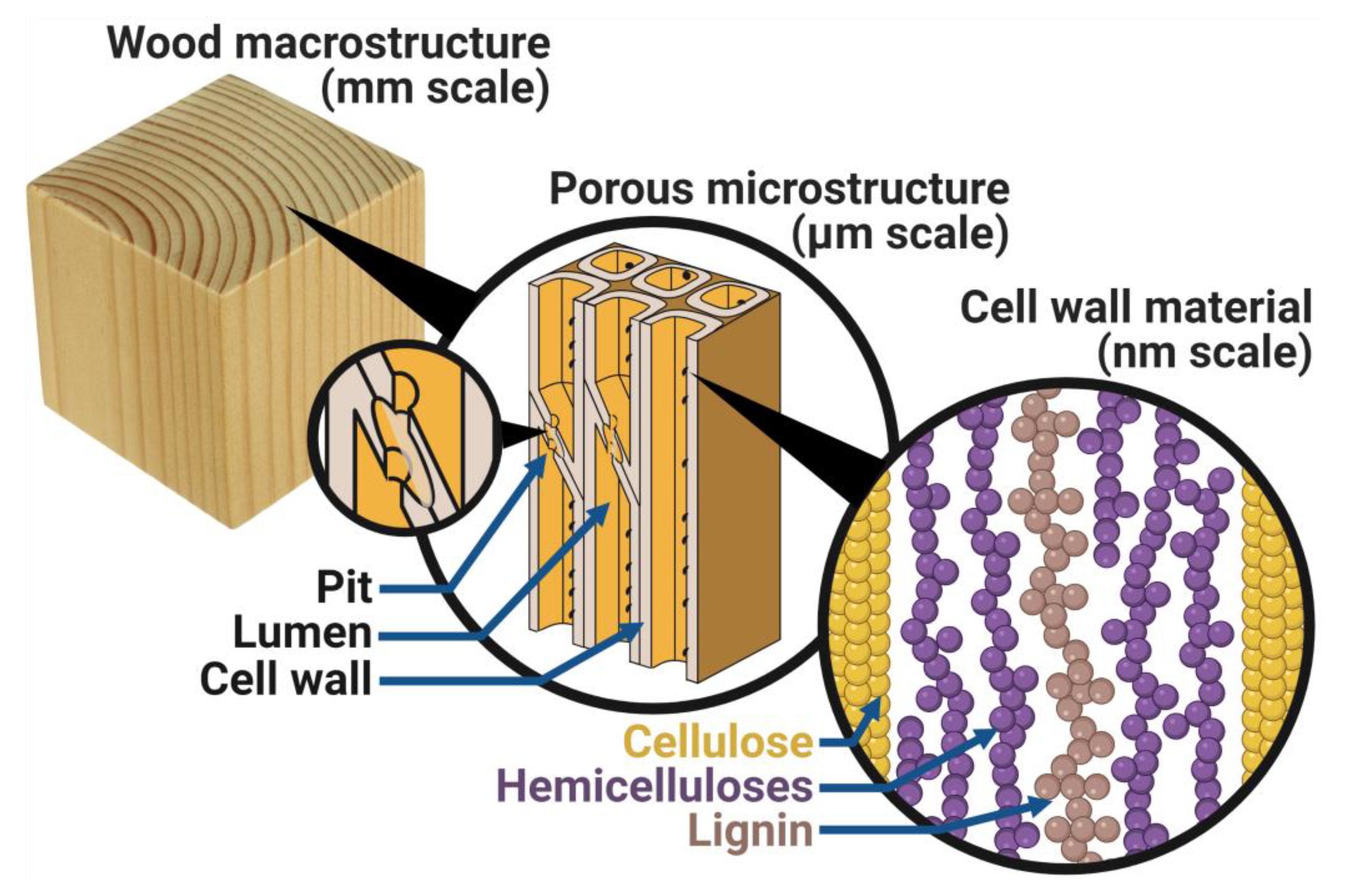
Moisture in wood has been studied for more than a century [3,18,19,20]. Despite a century of research efforts and scientific advances, significant knowledge gaps remain in our fundamental understanding of how moisture is taken up by wood and how it affects material behavior. One reason for this is the complexity of the wood material architecture and the properties of its building blocks. The porous and ordered structure of wood is schematically illustrated in Figure 1. Water molecules can be taken up in the variously sized macro-voids or in the solid cell walls where they interact with the three main constituents: cellulose, hemicelluloses, and lignin. How the water molecules interact with the wood, where they are located, and how the wood constituents and the material architecture respond to this are central to understand moisture in wood. As the following brief overview describes, much knowledge has been gained over the last century, but important questions remain unanswered.
2.1. Equilibrium Moisture States
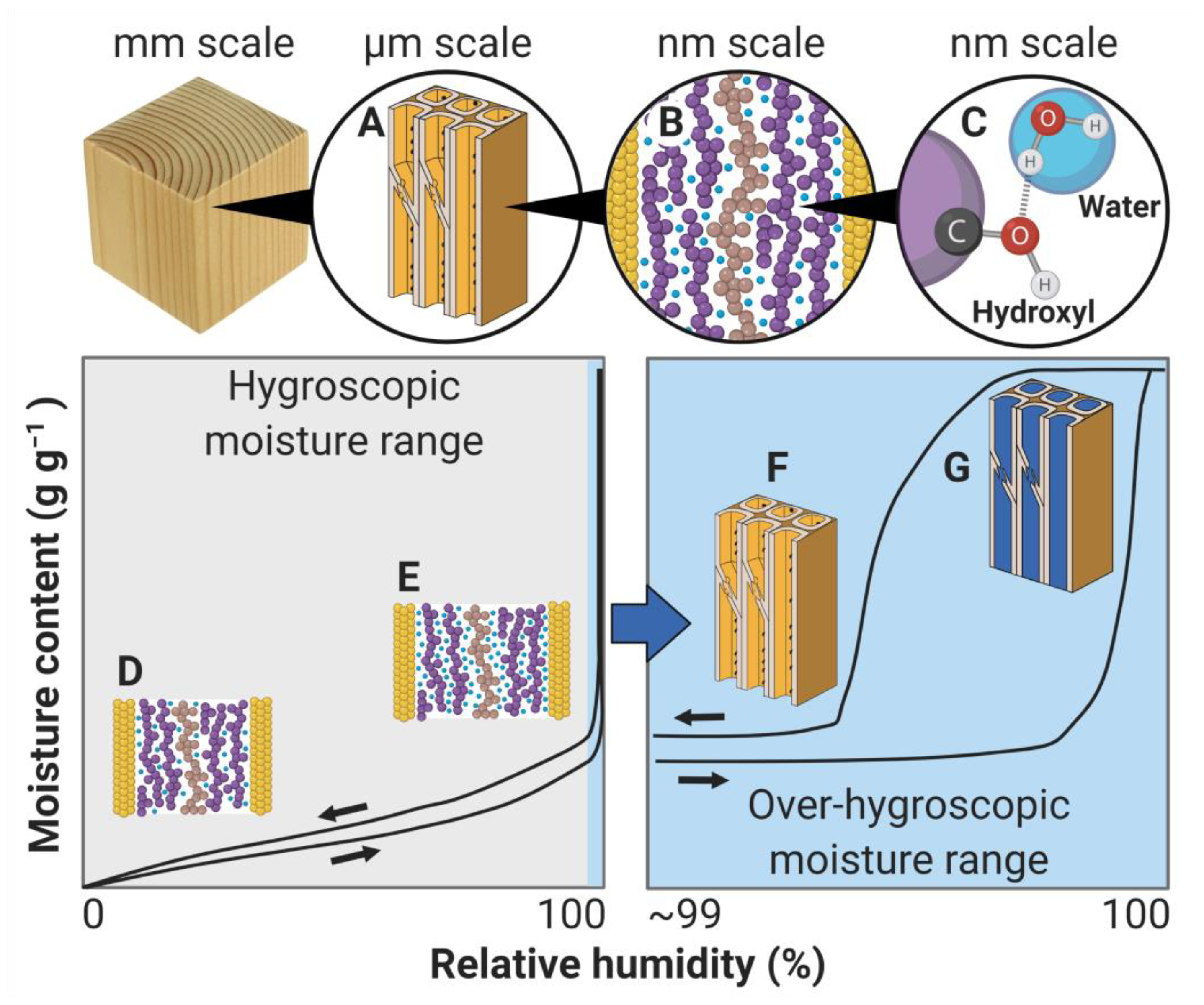
Moisture in wood can be present both within and outside of cell walls; see Figure 2. In the hygroscopic moisture range, generally defined as the range between 0 and 97–98% relative humidity (RH), moisture is predominately found within cell walls. This moisture interacts with the hydroxyl groups of the wood polymers by hydrogen bonding. Because the cell walls are virtually non-porous in the dry state [21,22], sorption of water in them results in swelling of the cell wall as well as of the bulk wood; see Section 2.2. The main mechanism of sorption for water outside of cell walls (in macro-voids such as lumina, pit chambers) is capillary condensation. That is, in confined spaces such as small macro-voids and pores within materials, water vapor condenses at a lower vapor pressure than the saturation vapor pressure [23]. The relative humidity at which capillary condensation occurs depends on pore size and geometry [24]. Because of the large dimensions of the macro-voids outside of cell walls, capillary condensation to a significant extent only occurs at very high humidity levels, above about 99% RH [25,26,27]. Previously, it was hypothesized that cell walls must be saturated before significant amounts of capillary condensed water are present outside of cell walls [3]. However, recent research has shown that the cell walls are not saturated until the whole piece of wood is completely saturated with water [28]. In the over-hygroscopic moisture range, moisture uptake within and outside of cell walls thus occurs simultaneously [28], although the uptake outside of cell walls is generally dominant in this range.
Sorption hysteresis exists in both the hygroscopic and the over-hygroscopic moisture ranges; see Section 2.3. For this reason, capillary water can exist at lower relative humidity levels than the relative humidity that corresponds to the limit for capillary condensation for a specific void. The reason for this is anatomical features, for example, vestured pits that act as ink-bottle necks and hinder the emptying of these macro-voids in desorption; see Section 2.3. In desorption, capillary water has therefore been detected at relative humidity levels down to 76–90% RH for some wood species [29,30,31]. In addition, water outside of cell walls is present if the wood is placed in contact with liquid water, e.g., in rain exposed structures.
The equilibrium moisture content, e.g., the mass of water relative to the dry mass, as a function of relative humidity, is generally presented in sorption isotherms; see Figure 2. In the over-hygroscopic range, sorption isotherms are typically shown as a function of pore water pressure or water potential instead of relative humidity; see, e.g., [28,32]. Substantial efforts have been devoted to understand the factors that control the amount of water within cell walls. The water molecules here interact with hydroxyl groups, and the amount of hydroxyl groups accessible to water molecules—the hydroxyl accessibility—has been correlated to the moisture content [33], but it is not the only factor that controls the moisture content of cell walls [34,35]. Additionally, the availability of space within cell walls has been suggested to be a highly contributing factor [35]. The internal space is controlled by the swelling of cell walls and, therefore, by the mechanics related to this; see Section 2.2. The complex interplay between the various factors controlling the equilibrium moisture content can be characterized by the thermodynamic state of water within cell walls. By determining the difference in enthalpy between water in the vapor phase or liquid state and water within cell walls, information about the interaction and state of the latter can be gained. The difference in enthalpy between the liquid state and the cell wall state of water has been denoted as mixing enthalpy by Nopens et al. [36]. This quantity can be thought of as the excess enthalpy, i.e., how much more heat is released when an infinitesimal amount of water is absorbed in wood compared to how much heat is released when the same amount of water condenses. The mixing enthalpy is therefore a state variable that describes the interaction between water and wood at a given moisture state. In the dry state of wood, the mixing enthalpy is around −1000 J/gwater, which is similar to the difference found in the dry state for a range of biochemical substances [37]. However, the mixing enthalpy is moisture dependent and decreases as the moisture content increases [36,38,39]. This indicates that, at higher moisture contents, the interaction energy between wood and the last added water molecules decreases.
While moisture equilibrium in the hygroscopic moisture range has been widely studied [40,41,42,43,44,45], the over-hygroscopic range has been less investigated. Because water is simultaneously taken up by cell walls and macro-voids in this range, the characterization of moisture states requires experimental techniques that can distinguish between water in different environments (e.g., cell walls and macro-voids). Several methods are able to partition the moisture content into different fractions based on the environment, such as Differential Scanning Calorimetry (DSC) [28,46,47] and low-field Nuclear Magnetic Resonance (NMR) [29,48,49,50], as well as the solute technique [50,51,52,53], although the latter can be used only in the water-saturated state. These methods are based on different principles of measurement and are associated with different types of errors and, therefore, give different results [50]. For example, low-field NMR performed above 0 °C have been shown to underestimate the moisture content within cell walls [49,50,54], which points out the need of measuring the same parameter with several different methods when possible and assessing the uncertainties of each method. Besides partitioning moisture in different locations, the solute exclusion technique and low-field NMR can provide further insights about the state of moisture in wood. The solute exclusion technique can, for instance, detail the accessibility of cell walls to probes of different sizes [55,56]. Although the results do not represent the absolute size distribution of water-filled nano-pores in cell walls [57,58], they provide a window into the size range of these nano-pores. The low-field NMR method can be used to determine location and state of water. Low-field NMR enables differentiation between water within and outside of cell walls as well as water in differently sized macro-voids [29,48]. While the method is commonly used to characterize moisture in water-saturated wood [48,49,50,54], it can also be used to study wood after moisture conditioning [29,59]. Conditioning, especially at high moisture contents is, however, often done by drying samples down to a specific moisture content. This introduces moisture gradients within the sample that can make the interpretation of results challenging due to non-uniform moisture distributions [60]. Increased knowledge on the location and state of water at different water potentials therefore requires careful conditioning procedures in order to get well-defined results.
Most commonly, 1D low-field NMR is used to partition moisture based on the relaxation time, T2, of the different moisture fractions [29,48,49,54]. However, the obtained T2 is affected by both the chemical and physical environment. Instead, two-dimensional (2D) low-field NMR offers the possibility to separate the effects of chemical and physical environments. Studies using this technique have found that there are two distinct populations of moisture within cell walls [59,61,62,63], which has also been observed with Quasielastic Neutron Scattering (QENS) [64]. It has been hypothesized that one of these water populations is related to water bound to wood polymers while the other is related to free water clusters [61], or that these water populations are present in different locations in different chemical environments within the cell wall material [59,62,64]. However, it is not yet clear what the two water populations observed with 2D low-field NMR and QENS represent. Older studies using DSC also determined two populations of moisture within cell walls: freezable and non-freezable water. However, Zelinka et al. [65] showed that the freezable water fraction was actually located in small macro-voids outside the cell walls, so this is most likely not related to the two water populations observed by 2D low-field NMR and QENS.
In the end, much remains to be discovered about the factors controlling the equilibrium moisture content in the full moisture range as well as the interplay between water in different physical locations (cell walls, macro-voids) and chemical environments within these. Central to gaining new knowledge will be the use of complimentary experimental techniques to characterize the moisture as well as appropriate moisture conditioning to ensure a uniform distribution of moisture states within the wood.
2.2. Swelling and Shrinkage
Dimensional change as a result of a change in moisture content is one of the obvious effects of moisture on wood. In the dry state, the solid cell walls of wood are virtually non-porous [21,22]. Therefore, water molecules entering the cell walls result in the creation of transient nano-pores [66] and an increase in dimensions of the cell wall material (swelling). Similarly, these nano-pores collapse, causing the dimensions to decrease (shrinkage) when water molecules are removed from the cell walls upon drying the wood. The volumetric changes of bulk wood are approximately linear in the moisture content range from dry to 25–30% [67,68,69]. Above this range the volumetric change per change in moisture content gradually decreases to zero in the 40–45% moisture content range [67,68,69]. The maximum moisture content for cell walls of many wood species is in the range 26–50% [46,47,49,50,70], and capillary water becomes increasingly dominant in the over-hygroscopic range. It remains to be seen whether the decline in volumetric changes corresponds with increasing amounts of capillary water that does not contribute to shrinkage and swelling. Another possibility could be a difference in the contribution from the two water populations found within cell walls, because the proportion of these changes with moisture content [59,64]. No matter the cause of the decreasing volumetric changes, more detailed information can be obtained from measurements on a scale closer to that of the cell walls than measurements on bulk wood specimens. For instance, X-ray tomography has been used to characterize swelling behavior in the hygroscopic moisture range of wood tissue [71,72,73] and on micro-pillars cut out of cell wall material [74]. Furthermore, developments in experimental setups for moisture conditioning in the over-hygroscopic range during X-ray tomography [75] could potentially also be used for swelling measurements.
The majority of the cellulose microfibrils are aligned in the same direction in the dominant S2 layer of the secondary cell wall, which restricts dimensional changes in this direction. Moreover, other cell wall layers with different microfibril orientations also affect the swelling of the cell walls [76]. These constraints contribute to the marked swelling anisotropy in wood, i.e., that shrinkage and swelling differ widely between the various anatomical directions of wood. At the same time, the constraint from the microfibrils results in significant mechanical stress in the cell wall as it shrinks and swells, as is evident from the deformation of the microfibrils themselves [77]. Swelling anisotropy can be seen on several length scales: from isolated micro-pillars of cell wall material [74] to isolated tissues of early- and latewood [71,72,78] and bulk wood [79,80]. On all these scales, the dimensional changes of the wood material may be affected by mechanical interactions between structural elements (cell wall biopolymers, cell wall layers, cells, tissues, etc.). This complexity makes the study of moisture induced dimensional changes and cell wall mechanics challenging [81], and so far the interaction between internal stress and moisture in equilibrium has not been resolved.
2.3. Moisture History Dependence
The equilibrium state of moisture in wood exhibits history dependence, i.e., the equilibrium state reached with specific external conditions does not only depend on these conditions, but also on the previous moisture states. This is evident in the sorption isotherms, where the equilibrium moisture content at a specific relative humidity depends on whether equilibrium was reached by moisture uptake (absorption) or release (desorption). The moisture history dependence of the sorption isotherms is known as sorption hysteresis. Figure 2 illustrates sorption hysteresis for wood in equilibrium reached by absorption from the dry state and by desorption from the water-saturated state. The curves illustrating these equilibrium states are known as the absorption isotherm and desorption isotherm, respectively. Between these curves, there is a wide range of equilibrium states that can be reached by absorption from non-dry equilibrium states or desorption from non-saturated equilibrium states [82,83,84,85].
Sorption hysteresis exists in both the hygroscopic and over-hygroscopic moisture ranges; see Figure 2. Because the mechanism of moisture sorption differs between these two ranges, as described in Section 2.1, the physical background for sorption hysteresis is also different. In the hygroscopic range, dominated by moisture in cell walls, sorption hysteresis decreases with increasing temperature and seemingly vanishes above 75 °C [40]. This has spawned the idea that sorption hysteresis is linked with softening of the hemicelluloses [86,87,88], which increases the mobility of these polymers. Hence, the cell walls become more pliable and relaxation of mechanical stresses is faster. This hints at a connection between sorption hysteresis and the mechanical stresses arising from shrinkage and swelling. However, the exact mechanisms behind sorption hysteresis in cell walls remain unresolved.
In the over-hygroscopic moisture range, moisture is predominantly found as capillary water, and sorption hysteresis is even more pronounced than in the hygroscopic range; see Figure 2. The sorption mechanism for capillary water relates to the wood anatomy, i.e., the sizes of macro-voids found in the wood structure, as described in Section 2.1. Thus, capillary water held in smaller macro-voids exists at equilibrium with lower levels of relative humidity than capillary water in larger macro-voids. As a result, smaller macro-voids will absorb capillary water before larger pores when wood is conditioned to successively higher levels of relative humidity in the over-hygroscopic range. Some of the smallest macro-voids of significant total volume in the porous wood structure are the pits connecting adjacent cells. During desorption, these pits are thought to act as bottlenecks that prevent emptying of larger macro-voids such as the cell lumina. Thus, sorption hysteresis in the over-hygroscopic range is hypothesized to be caused by these different mechanisms in absorption and desorption; capillary condensation of water in successively larger macro-voids during absorption while emptying is controlled by the small bottleneck pits in desorption [27,89]. On top of this, a small part of sorption hysteresis is related to sorption hysteresis in cell walls, which persists also in the over-hygroscopic range [28].
Shrinkage and swelling are caused by changes in the cell wall moisture content. Because the moisture content in equilibrium with specific climate conditions can vary as a result of sorption hysteresis, the swelling of wood at these conditions can vary. However, studies on bulk wood have shown that the dimensions of the same piece of wood at similar moisture content can be slightly higher if equilibrium is reached by desorption rather than by absorption [90,91,92,93,94]. This swelling hysteresis is, however, small and sometimes not found [4], which is also the case in studies of the swelling on the tissue scale using X-ray tomography [72,73].
2.4. Sorption Kinetics and Diffusion
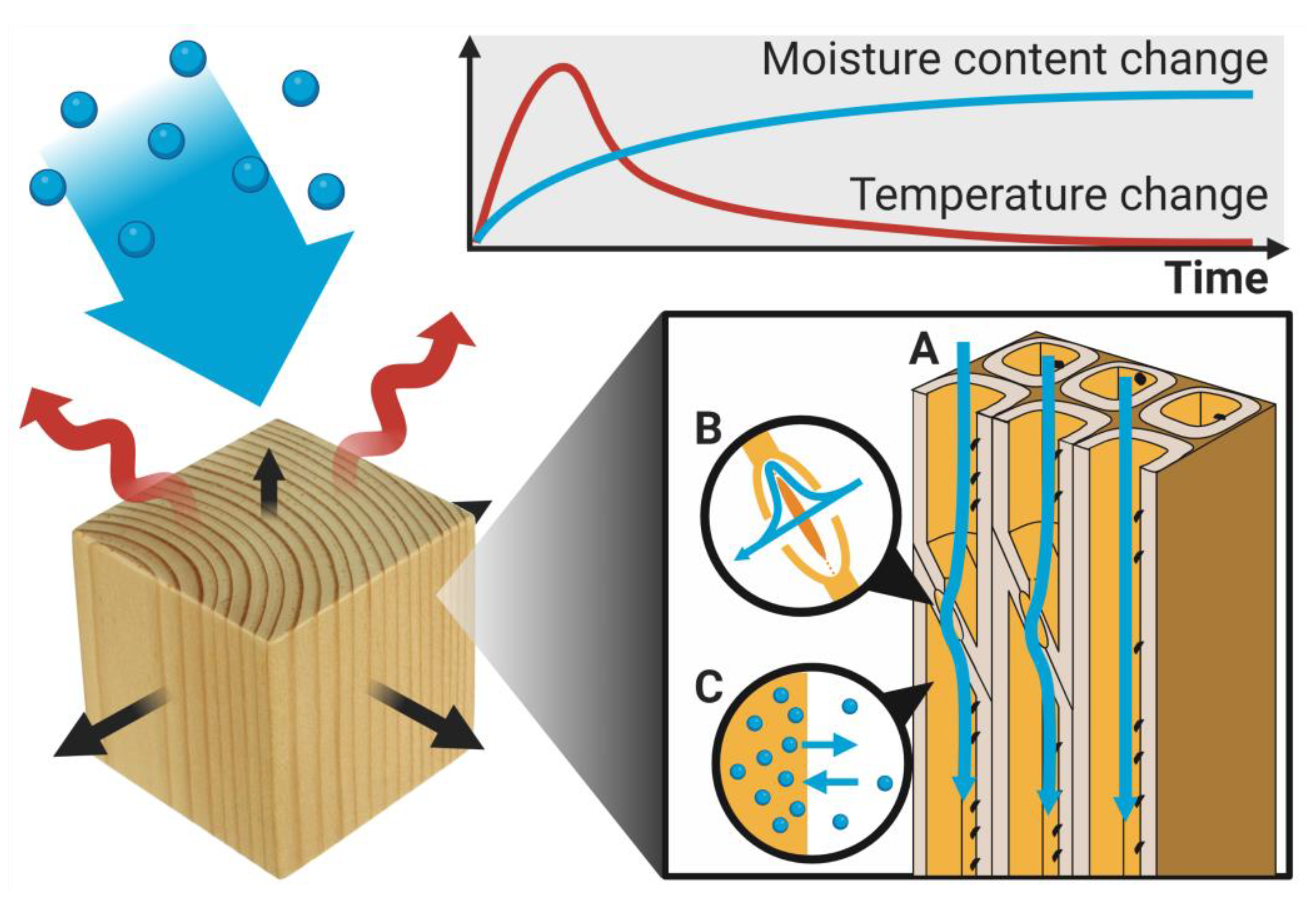
The time-dependent process of moisture being taken up or released when wood approaches equilibrium is known as sorption kinetics. This involves the same physical factors discussed previously for the equilibrium state, such as the cell wall mechanics during shrinkage and swelling with, however, additional contributions from other phenomena [95]; see Figure 3. These include temperature changes arising from the phase change of water (difference in enthalpy), diffusion into cell walls, and resistance to this from stagnant air at the surfaces. Moreover, for wood specimens without all surfaces in direct contact with the external climate, the moisture uptake process also involves diffusion into the porous wood structure and gradients in moisture and temperature on the bulk scale. A complicating factor in analyzing sorption experiments is the effect of moisture diffusion in the porous wood structure as well as contributions from the diffusion of cell wall water in relation to moisture gradients in the bulk wood sample. This effect has been investigated by using wood specimens of different (thin) thicknesses [96,97] to generate different lengths of the diffusion pathways into the wood. The diffusion and sorption processes hereafter needs to be disentangled by careful analysis. The reader is referred to [95] for a more detailed discussion of the various phenomena involved in sorption kinetics.
Research on moisture sorption kinetics in wood has increased over the last decades as a result of the increased use of automated sorption balances in wood research laboratories. This equipment allows the monitoring of mass changes during moisture sorption under controlled climatic conditions with a high temporal and gravimetric resolution. The experimental data hereby obtained, however, usually lacks complementary information on the other phenomena occurring simultaneously with mass change such as temperature changes, with notable exceptions [98]. Decades ago, sorption studies on wood used (non-automated) vacuum sorption balances with instrumentation to record the surface temperature simultaneously with mass change [99,100].
Several theoretical models have been developed to describe sorption kinetics [101,102], but one of the most popular was, for years, the Parallel Exponential Kinetics (PEK) model. This model divides the moisture sorption process into two components with different characteristic times (“fast” and “slow” moisture) based on fitting to experimental sorption kinetic data. However, the PEK model was shown by Thybring et al. [101] to mischaracterize sorption kinetics because more than two components with different time constants were often observed in the data [102]. The other theoretical models described in the literature include the thermally limited moisture transfer (TLMT) model by Willems [103,104]. Although this model has been criticized by Thybring et al. [95] on the same grounds as the PEK model, i.e., describing sorption kinetics by only two process components, Willems [104] argues that the TLMT behavior is a contribution to the overall sorption kinetics. The lack of detailed experimental data for other physical properties than mass is, however, a current barrier to a deeper understanding of sorption kinetics and to testing hypotheses and model predictions.
As described previously, the uptake or release of moisture from bulk wood specimens involves diffusion of water vapor in the porous wood structure. Furthermore, diffusion of cell wall moisture also contributes to the overall transport of moisture in bulk wood. However, the dominant contribution to the moisture diffusivity for bulk wood, i.e., the material property describing the correlation between moisture flux and moisture gradient in the bulk material, comes from the transport of water vapor. This is evident from the significantly larger moisture diffusivity (approx. 4–20 times) in the longitudinal direction of bulk wood than in the transverse (radial and tangential) directions [105]. The vast majority of cells in the wood are oriented along the longitudinal direction, where their lumina provide longer pathways for water vapor diffusion than in the across-the-cell direction; typical cell length-to-width ratio of softwood tracheids is about 70:1 to 100:1 [106,107]. Given the dominant contribution of water vapor diffusion to moisture transport in bulk wood, the contribution from moisture diffusion in cell walls is often assumed to be negligible [108]. The moisture diffusivity of cell walls has been estimated to a magnitude around 10−12 m2/s, with moisture content as the driving potential [88]. This is closely similar to the diffusivity found by molecular dynamics simulations of moisture diffusion in amorphous cellulose [109] when the results are transformed to the same driving potential. The moisture diffusivity of cell walls remains scarcely experimentally characterized. In materials science, moisture diffusivity is regularly determined from sorption experiments on specimens with a simple geometry [110]. From these experiments, the diffusivity can be calculated by modelling the change in moisture content using Fick’s laws of diffusion. For wood, the sorption kinetics is affected by more phenomena than just diffusion, and sorption experiments are therefore unable to provide insights to the moisture diffusivity. For bulk wood specimens, the diffusivity can be determined from measurements of the flux through a specimen exposed to a constant moisture gradient [105], i.e., a steady-state experiment with an unchanging moisture distribution in the specimen such as by using the cup method. While the traditional cup method uses bulk specimens, a similar setup has been developed for measurements in a sorption balance using much smaller specimens [111]. However, using this setup for small wood specimens does not provide information about the moisture diffusivity of cell walls, because the measurement involves a dominant contribution from water vapor diffusion in the porous wood structure.
3. Wood Modifications and Their Effect on Moisture
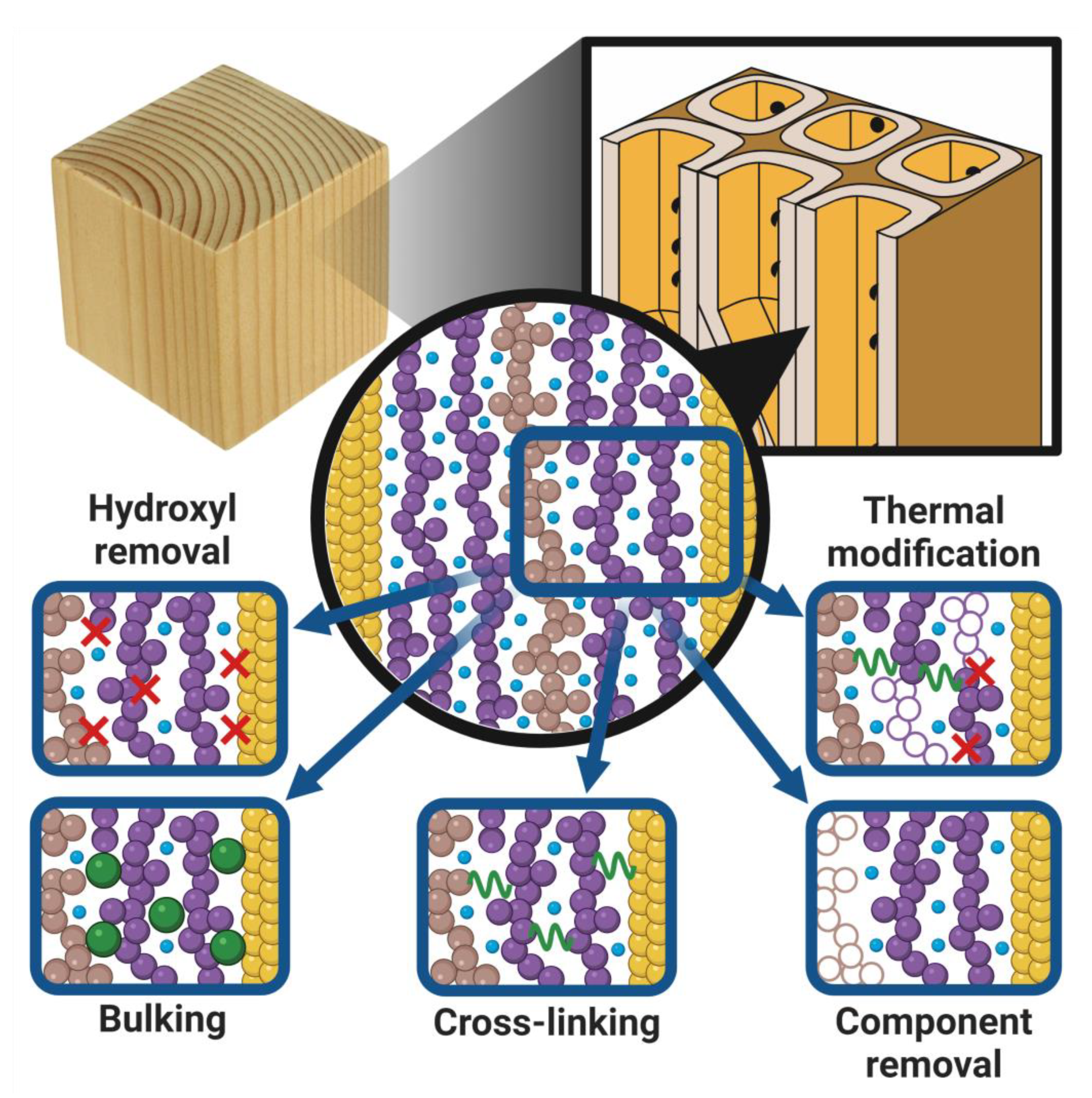
Literature is abundant with modifications for altering the chemistry of wood, and new modification techniques continue to be developed in laboratories worldwide; see, for instance, [6,16,112]. In this section, we focus on a handful of effects of modification on wood chemistry that are relevant to moisture in wood. These effects are of course simplifications of the complexity of real modifications on the wood chemistry, but they help illustrate some common traits among wood modifications. Figure 4 shows a schematic illustration of the five effects described in more detail below.
- Hydroxyl removal: Many modifications involve chemical reaction with hydroxyl groups on the wood components. Because hydroxyls are the main functional group interacting with moisture, hydroxyl removal can affect the number of functional groups interacting with moisture [34]. Hydroxyl removal occurs, for instance, by chemical reaction where hydroxyls are substituted for another type of functional group [113]. However, these functional groups can contain hydroxyl groups themselves, and the hydroxyl concentration after modification may therefore be decreased, preserved, or increased. Examples: Acetylation [114], DMDHEU treatment [17], hexamethylene diisocyanate treatment [115].
- Bulking: Modifications that add molecular moieties inside the wood cell walls and reduce the available space for moisture [35]. Moieties can either be added through reaction with functional groups on wood components, e.g., hydroxyls, or by in-situ polymerisation reactions. Examples: Acetylation [114], furfurylation [116], DMDHEU treatment [17].
- Cross-linking: Modifications involving chemicals that can react with at least two functional groups may create molecular moieties that form a covalently bonded bridge between adjacent wood components. This restricts the mobility of these adjacent components and reduces the capacity for swelling [17]. Examples: Formaldehyde treatment [117], DMDHEU treatment [17], hexamethylene diisocyanate treatment [115].
- Thermal modification: A variety of thermal modification processes has been described in the literature. In these, the wood is heated to around 150–240 °C [14], whereby primarily hemicelluloses are thermally degraded and partially removed. Depending on the atmosphere, including the amount of moisture in the treatment process, a variety of chemical reactions can occur, e.g., cross-linking reactions [14]. Thus, thermal modification can have multiple effects on the moisture in the modified wood, depending on the process conditions. Examples: Open and closed processes in wet or dry moisture conditions; see more in [14].
Apart from using different types of modifications, as described above, a possible variation is to use targeted modification, i.e., tuning the spatial distribution of modification reactions in the wood structure. Multiple examples exist of modifications that are limited to the macro-void structure or have either little or full penetration into cell walls. However, Keplinger et al. [120] pioneered the deliberate tuning of the spatial distribution to either full cell wall modification or limited modification reactions to the cell wall surfaces through selection of the chemical route. Targeted modification has also been further explored by Digaitis et al. [121] for acetylation.
3.1. Modifying the Equilibrium Moisture States
The moisture in cell walls interacts with hydroxyl groups within small internal spaces (nano-pores) created by the water molecules by swelling the cell wall material. All of these factors can be altered by a range of modification techniques that may reduce the hydroxyl accessibility [14,122] and the available space within cell walls [56], as well as the cell wall moisture content [35]. For example, thermal treatment can give a similar decrease in hydroxyl accessibility but reduce the moisture content to different extents, depending on the treatment process [123]. Modification with DMDHEU restricts swelling by crosslinking but simultaneously increases the hydroxyl accessibility [124]. Acetylation reduces the hydroxyl accessibility with an increased degree of modification [122,125] and limits the available space within cell walls [56]. Alternatively, the internal space and swelling behavior can be altered by selective component removal. For example, hemicellulose extraction reduces the moisture content [126], while delignification increases the moisture content in the hygroscopic range [118], except when performed to such a high degree that it causes collapse of the cell wall [118]. In the latter case, both absorption and desorption has been seen to decrease but, interestingly, the desorption isotherm above 90% RH was found to increase [118]. Moreover, component removal gives additional possibilities for changing the factors affecting moisture equilibrium by a subsequent treatment such as a bulking or cross-linking modification [119]. There are, thus, multiple ways in which different modifications can be used and combined to achieve certain properties of the wood cell walls. For example, a recent study [35] has shown that it is possible to independently alter hydroxyl accessibility and available space within cell walls.
By deliberate tuning of the factors described above, including the chemical composition of the cell walls, knowledge gaps about the equilibrium moisture state could be addressed. For example, by studies of changes in equilibrium moisture content as function of these many factors. Additionally, altering the hydroxyl accessibility and the mechanical properties of the cell walls are expected to affect the strength of the interaction with water and the mechanical work required to swell the cell wall material. This would undoubtedly affect the thermodynamics of moisture sorption, and characterization of the enthalpy of water within cell walls by calorimetric measurements could hereby be a novel window into insights about moisture in wood.
Complementary information about the equilibrium state at different moisture contents could be gained from studies of the two water populations within cell walls observed with 2D low-field NMR and QENS; see Section 2.1. Thermally treated oak [63] and sulfur dioxide treated spruce [62] have previously been characterized with 2D low-field NMR. However, a carefully designed study using wood with altered chemical composition by component removal and subsequent tuning of the internal space and hydroxyl accessibility by modification could be helpful in the understanding of these two water populations and their chemical environments.
Modification does not only affect moisture within cell walls. For instance, acetylation and furfurylation have been shown to affect the interaction between wood and capillary water [54,119,127]. This could be caused by a change in contact angle, which has been observed for both acetylated and furfurylated wood [128]. If such a change in contact angle also occurs within cell lumina, it would presumably affect at which relative humidity capillary condensation occurs within the macro-voids because this also depends on the contact angle [24]. Targeted modification of the cell wall-lumen interface can therefore be used to change the contact angle and thus control the interaction with capillary water independent from the interaction with water in cell walls. This would enable studies of the interplay between capillary water in different macro-voids and moisture within cell walls. Alternatively, the latter can be isolated completely for studies in the over-hygroscopic moisture range by filling up lumina and other macro-voids, as will be described in Section 3.4.
3.2. Modifying the Swelling and Shrinkage
One of the original targets of wood modification was dimensional stabilization of the material [11], i.e., to limit the shrinkage and swelling. Thus, many of the available wood modification technologies reduce the dimensional changes, partly because less moisture is taken up by the wood cell walls. However, modifications affect the physical properties of wood in different ways, as described in the beginning of Section 3. Selective removal of cell wall components loosens the structure and creates space within cell walls for more water [129]. On the other hand, cross-linking modifications may increase the cell wall stiffness and limit stress relaxation [130]. Certain cross-linking modifications even contain hydroxyls, which enables the modification to simultaneously reduce the cell wall moisture content and increase hydroxyl accessibility of the modified cell walls [124]. Thermal modification removes mainly hemicelluloses, and cross-linking reactions may occur depending on the moisture conditions during modification. As a result, thermally modified wood can be processed to either increase or decrease the swelling [123]. By tuning the internal space within cell walls for moisture and the stiffness of the cell wall material, it might be possible to elucidate the interaction between moisture induced swelling and cell wall mechanics. Furthermore, the potential difference in swelling contribution between the two water populations found within cell walls could be studied in this way [63]. A deeper understanding of this interaction will, however, require a shift away from measurements on the bulk wood scale to detailed investigations on smaller length scales. A variety of non-destructive characterization techniques offers possibilities for investigation of wood swelling at the micro-scale, such as X-ray tomography [71,72,74] or deformations and moisture states at even smaller scales by scattering techniques [76,81,131].
3.3. Modifying the Moisture History Dependence
Many wood modifications reduce the cell wall moisture content, as described in Section 3.1. This generally leads to a reduction in the absolute difference between the absorption and desorption isotherms for bulking and cross-linking modifications [125,132,133]. However, modifications that remove material from the cell walls, such as thermal treatment or delignification, have been observed to increase the sorption hysteresis at least temporarily or for certain segments of the moisture range [118,134,135]. This variability in the effect of modification on sorption hysteresis presents an opportunity for investigating the fundamental mechanisms behind this phenomenon.
Another possibility to control the extent of sorption hysteresis could be by changing the stiffness of the cell wall. The mechanical properties of the cell walls can, for instance, be altered by cross-linking modifications that reduce the polymer mobility and increase the stiffness. This would not only reduce the cell wall moisture content, but also restrain softening of the cell wall polymers [136]. Softening may also be hindered by modifications that reduce the cell wall moisture content without cross-linking polymers, e.g., bulking, simply because too few water molecules are present to promote polymer mobility [137]. Thus, acetylation has been shown to increase the softening point of water-saturated wood [138]. However, it is also possible that bulking may facilitate softening by acting as a plasticizer, similar to the effect of impregnating wood with polyethylene glycols [139]. The effect of modification on the softening and cell wall mechanics may require both advanced experimental techniques and new analysis methods [137,140].
3.4. Modifying the Sorption Kinetics and Diffusion
Wood modification provides several opportunities for manipulating the sorption process in order to elucidate the fundamental mechanisms. One strategy for disentangling the simultaneous diffusion and sorption processes occurring in bulk wood samples is to remove the diffusion of water vapor. Hereby, the moisture uptake process is restricted to sorption and diffusion of cell wall moisture. This was done by Stamm [141] by impregnating wood samples with a metal alloy of low melting temperature. After cooling, the impregnated samples had most of the cell lumina filled with stiff metal. The disadvantage of this approach is that the impregnated metal might limit swelling and thereby generate stresses within the wood, and cracks might form at the interfaces between cell wall surfaces and the metal. Instead, the porous wood structure could be filled with hydrophobic, flexible polymers that are grafted onto the cell wall surfaces, as done previously by Keplinger et al. [120,142] for other purposes. Moreover, the effect of the temperature change associated with moisture sorption could be manipulated with a similar methodology by the inclusion of thermally conductive fillers in lumen-filling polymers [143]. Irrespective of how the thermal properties of the material are manipulated by modification, it is necessary to characterize changes in both mass and temperature in order to gain a deeper understanding of the sorption kinetics of wood.
Filling up the macro-voids in the wood structure with hydrophobic polymers, grafted to cell wall surfaces, could also be used to investigate moisture diffusion in cell walls. Modifications that reduce the moisture content of cell walls can be expected to decrease the diffusivity in agreement with results on bulk wood moisture diffusivity after thermal modification [144]. Subsequent modification of cell walls after lumen filling could isolate moisture diffusion to cell walls. A potential challenge with the incomplete filling of lumina could be circumvented by using small, thin specimens and characterizing their diffusivity using an adapted cup method setup in a sorption balance [111].
4. Perspectives
Introducing specific chemical changes and even controlling the spatial distribution of these, in combination with detailed characterization of moisture in wood, could give novel insights into the interaction between moisture and wood. Such novel insights could further contribute to applications in several related fields of research. For example, it could give new insights into how to enhance the resistance of wood to fungal decomposition in moist environments. Water is an essential parameter in decomposition processes, but the exact role of water in the decomposition processes is not yet fully understood, neither in untreated wood nor in modified wood [1]. A more detailed understanding of wood-moisture interactions could give important knowledge on the role of water in decomposition processes. This would in turn give important information on how to design modification techniques and processes to further enhance the performance of wood in outdoor structures in a sustainable and cost-effective way.
Additionally, an in-depth understanding of the interaction between moisture and wood could give new insights into how to further improve the performance of moisture-induced wooden actuators by altering the swelling and/or swelling anisotropy. Such wooden actuators can be used in architectural and constructional elements that adapt when the ambient climate changes [145,146], with the advantage that they do not need a power supply, which is valuable in large-scale applications [146]. Because of the anisotropic structure of wood, several different types of moisture-induced actuation, e.g., bending or torsional actuation, are possible. Detailed knowledge on how to control and predict these moisture induced deformations would thus be beneficial for research within this field. Furthermore, novel insights into wood-moisture interactions and moisture induced deformations could give valuable input on how to improve the utilization of wood biomass with problematic swelling anisotropy, for example, wood species with spiral grain or juvenile wood in the upper parts of the stem. This could enable new applications for some wood species or a more effective use of the whole wood stem.
5. Conclusions
This review shows that there are multiple interesting pathways in which wood modification could be used as a tool to learn more about the interactions between moisture and wood, native as well as modified. By using already existing modification techniques, combining different modification techniques, and/or controlling the spatial distribution of the modification, several of the existing knowledge gaps within the field could be addressed.
Author Contributions
Conceptualization, E.E.T.; investigation, E.E.T. and M.F.; writing—original draft preparation, E.E.T. and M.F.; writing—review and editing, E.E.T. and M.F.; visualization, E.E.T.; project administration, E.E.T. and M.F.; funding acquisition, M.F and E.E.T. All authors have read and agreed to the published version of the manuscript.
Funding
Funding from Interreg Öresund-Kattegat-Skagerrak, SNS Forest Research and the Swedish Research Council FORMAS (grant no. 2018-00387) is gratefully acknowledged.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
Figures created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brischke, C.; Alfredsen, G. Wood-water relationships and their role for wood susceptibility to fungal decay. Appl. Microbiol. Biotechnol. 2020, 104, 3781–3795. [Google Scholar] [CrossRef]
- Boddy, L. Effect of temperature and water potential on growth-rate of wood-rotting basidiomycetes. Trans. Br. Mycol. Soc. 1983, 80, 141–149. [Google Scholar] [CrossRef]
- Tiemann, H.D. Effect of Moisture upon the Strength and Stiffness of Wood; US Department of Agriculture, Forest Service: Washington, DC, USA, 1906.
- Ishimaru, Y.; Arai, K.; Mizutani, M.; Oshima, K.; Iida, I. Physical and mechanical properties of wood after moisture conditioning. J. Wood Sci. 2001, 47, 185–191. [Google Scholar] [CrossRef]
- Sargent, R. Evaluating dimensional stability in solid wood: A review of current practice. J. Wood Sci. 2019, 65, 36. [Google Scholar] [CrossRef]
- Gérardin, P. New alternatives for wood preservation based on thermal and chemical modification of wood—A review. Ann. For. Sci. 2016, 73, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Thybring, E.E. The decay resistance of modified wood influenced by moisture exclusion and swelling reduction. Int. Biodeterior. Biodegrad. 2013, 82, 87–95. [Google Scholar] [CrossRef]
- Rowell, R.M.; Ibach, R.E.; McSweeny, J.; Nilsson, T. Understanding decay resistance, dimensional stability and strength changes in heat-treated and acetylated wood. Wood Mater. Sci. Eng. 2009, 4, 14–22. [Google Scholar] [CrossRef]
- Ringman, R.; Beck, G.; Pilgård, A. The importance of moisture for brown rot degradation of modified wood: A critical discussion. Forests 2019, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Luković, M.; Mendoza, M.; Schlepütz, C.M.; Griffa, M.; Xu, B.; Gaan, S.; Herrmann, H.; Burgert, I. Bioinspired struvite mineralization for fire-resistant wood. ACS Appl. Mater. Interfaces 2019, 11, 5427–5434. [Google Scholar] [CrossRef] [PubMed]
- Stamm, A.J.; Burr, H.K.; Kline, A.A. Staybwood... Heat-stabilized wood. Ind. Eng. Chem. 1946, 38, 630–634. [Google Scholar] [CrossRef]
- Merk, V.; Chanana, M.; Gierlinger, N.; Hirt, A.M.; Burgert, I. Hybrid wood materials with magnetic anisotropy dictated by the hierarchical cell structure. Acs Appl. Mater. Interfaces 2014, 6, 9760–9767. [Google Scholar] [CrossRef]
- Cabane, E.; Keplinger, T.; Merk, V.; Hass, P.; Burgert, I. Renewable and functional wood materials by grafting polymerization within cell walls. ChemSusChem 2014, 7, 1020–1025. [Google Scholar] [CrossRef]
- Hill, C.; Altgen, M.; Rautkari, L. Thermal modification of wood—A review: Chemical changes and hygroscopicity. J. Mater. Sci. 2021, 56, 6581–6614. [Google Scholar] [CrossRef]
- Sandberg, D.; Kutnar, A.; Mantanis, G. Wood modification technologies—A review. Iforest Biogeosciences For. 2017, 10, 895–908. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Wang, K.; Li, J.; Zhang, S.; Shi, S.Q. Environmentally benign wood modifications: A review. Acs Sustain. Chem. Eng. 2020, 8, 3532–3540. [Google Scholar] [CrossRef]
- Emmerich, L.; Bollmus, S.; Militz, H. Wood modification with DMDHEU (1.3-dimethylol-4.5-dihydroxyethyleneurea)—State of the art, recent research activities and future perspectives. Wood Mater. Sci. Eng. 2017, 14, 13–18. [Google Scholar] [CrossRef]
- Volbehr, B.F.K.J. Untersuchungen Über Die Quellung der Holzfaser [Investigations of the Swelling of Wood Fibres]; Universität Kiel: Kiel, Germany, 1896. [Google Scholar]
- Zeller, S.M. Humidity in relation to moisture imbibition by wood and to spore germination on wood. Ann. Mo. Bot. Gard. 1920, 7, 51–74. [Google Scholar] [CrossRef]
- Carrington, H. The elastic constants of spruce as influenced by moisture content. Aeronaut. J. 1922, 26, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Salmén, L.; Fahlén, J. Reflections on the ultrastructure of softwood fibers. Cellul. Chem. Technol. 2006, 40, 181–185. [Google Scholar]
- Papadopoulos, A.N.; Hill, C.A.S.; Gkaraveli, A. Determination of surface area and pore volume of holocellulose and chemically modified wood flour using the nitrogen adsorption technique. Holz Als Roh- Und Werkst. 2003, 61, 453–456. [Google Scholar] [CrossRef]
- Thomson, W. On the equilibrium of vapour at a curved surface of liquid. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1871, 42, 448–452. [Google Scholar] [CrossRef]
- Sing, K.S.W.; Rouquerol, F.; Rouquerol, J.; Llewellyn, P. 8—Assessment of mesoporosity. In Adsorption by Powders and Porous Solids, 2nd ed.; Rouquerol, F., Rouquerol, J., Sing, K.S.W., Llewellyn, P., Maurin, G., Eds.; Academic Press: Oxford, UK, 2014; pp. 269–302. [Google Scholar] [CrossRef]
- Thygesen, L.G.; Engelund, E.T.; Hoffmeyer, P. Water sorption in wood and modified wood at high values of relative humidity. Part I: Results for untreated, acetylated, and furfurylated Norway spruce. Holzforschung 2010, 64, 315–323. [Google Scholar] [CrossRef]
- Engelund, E.T.; Thygesen, L.G.; Hoffmeyer, P. Water sorption in wood and modified wood at high values of relative humidity. Part 2: Appendix. Theoretical assessment of the amount of capillary water in wood microvoids. Holzforschung 2010, 64, 325–330. [Google Scholar] [CrossRef]
- Fredriksson, M. On wood–water interactions in the over-hygroscopic moisture range—Mechanisms, methods, and influence of wood modification. Forests 2019, 10, 779. [Google Scholar] [CrossRef] [Green Version]
- Fredriksson, M.; Thybring, E.E. On sorption hysteresis in wood: Separating hysteresis in cell wall water and capillary water in the full moisture range. PLoS ONE 2019, 14, e0225111. [Google Scholar] [CrossRef]
- Almeida, G.; Gagne, S.; Hernandez, R.E. A NMR study of water distribution in hardwoods at several equilibrium moisture contents. Wood Sci. Technol. 2007, 41, 293–307. [Google Scholar] [CrossRef]
- Passarini, L.; Malveau, C.; Hernández, R. Water state study of wood structure of four hardwoods below fiber saturation point with nuclear magnetic resonance. Wood Fiber Sci. 2014, 46, 480–488. [Google Scholar]
- Hernandez, R.E.; Caceres, C.B. Magnetic resonance microimaging of liquid water distribution in sugar maple wood below fiber saturation point. Wood Fiber Sci. 2010, 42, 259–272. [Google Scholar]
- Almeida, G.; Hernandez, R.E. Influence of the pore structure of wood on moisture desorption at high relative humidities. Wood Mater. Sci. Eng. 2007, 2, 33–44. [Google Scholar] [CrossRef]
- Chow, S.Z. Hydroxyl accessibility, moisture-content, and biochemical activity in cell-walls of Douglas-fir trees. Tappi 1972, 55, 539–544. [Google Scholar]
- Rautkari, L.; Hill, C.A.S.; Curling, S.; Jalaludin, Z.; Ormondroyd, G.A. What is the role of the accessibility of wood hydroxyl groups in controlling moisture content? J. Mater. Sci. 2013, 48, 6352–6356. [Google Scholar] [CrossRef]
- Thybring, E.E.; Piqueras, S.; Tarmian, A.; Burgert, I. Water accessibility to hydroxyls confined in solid wood cell walls. Cellulose 2020, 27, 5617–5627. [Google Scholar] [CrossRef]
- Nopens, M.; Wadsö, L.; Ortmann, C.; Fröba, M.; Krause, A. Measuring the heat of interaction between lignocellulosic materials and water. Forests 2019, 10, 674. [Google Scholar] [CrossRef] [Green Version]
- Kocherbitov, V. Salt-saturated salt solution as a standard system for sorption calorimetry. Thermochim. Acta 2004, 421, 105–110. [Google Scholar] [CrossRef]
- Kelsey, K.E.; Clarke, L.N. The heat of sorption of water by wood. Aust. J. Appl. Sci. 1956, 7, 160–175. [Google Scholar]
- Hearmon, R.F.S.; Burcham, J.N. Specific heat and heat of wetting of wood. Nature 1955, 176, 978. [Google Scholar] [CrossRef]
- Weichert, L. Investigations on sorption and swelling of spruce, beech and compressed beech wood at temperatures between 20 °C and 100 °C. Holz Als Roh- Und Werkst. 1963, 21, 290–300. [Google Scholar] [CrossRef]
- Kelsey, K.E. The sorption of water vapour by wood. Aust. J. Appl. Sci. 1957, 8, 42–54. [Google Scholar]
- Djolani, B. Hysteresis and second order effects of the moisture sorption in the wood at temperatures of 5, 21, 35 and 50 celsius. Ann. Sci. For. 1972, 29, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Christensen, G.N.; Hergt, H.F.A. Effect of previous history on kinetics of sorption by wood cell walls. J. Polym. Sci. A1 1969, 7, 2427–2430. [Google Scholar] [CrossRef]
- Hedlin, C.P. Sorption isotherms of twelve woods at subfreezing temperatures. For. Prod. J. 1967, 17, 43–48. [Google Scholar]
- Pidgeon, L.M.; Maass, O. The adsorption of water by wood. J. Am. Chem. Soc. 1930, 52, 1053–1069. [Google Scholar] [CrossRef]
- Simpson, L.A.; Barton, A.F.M. Determination of the fibre saturation point in whole wood using differential scanning calorimetry. Wood Sci. Technol. 1991, 25, 301–308. [Google Scholar] [CrossRef]
- Passarini, L.; Zelinka, S.L.; Glass, S.V.; Hunt, C.G. Effect of weight percent gain and experimental method on fiber saturation point of acetylated wood determined by differential scanning calorimetry. Wood Sci. Technol. 2017, 51, 1291–1305. [Google Scholar] [CrossRef]
- Fredriksson, M.; Thygesen, L.G. The states of water in Norway spruce (Picea abies (L.) Karst.) studied by low-field nuclear magnetic resonance (LFNMR) relaxometry: Assignment of free-water populations based on quantitative wood anatomy. Holzforschung 2017, 71, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Telkki, V.V.; Yliniemi, M.; Jokisaari, J. Moisture in softwoods: Fiber saturation point, hydroxyl site content, and the amount of micropores as determined from NMR relaxation time distributions. Holzforschung 2013, 67, 291–300. [Google Scholar] [CrossRef]
- Thybring, E.E.; Digaitis, R.; Nord-Larsen, T.; Beck, G.; Fredriksson, M. How much water can wood cell walls hold? A triangulation approach to determine the maximum cell wall moisture content. PLoS ONE 2020, 15, e0238319. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.E.; Scallan, A.M. Effect of component removal upon porous structure of cell wall of wood. 2. Swelling in water and fiber saturation point. Tappi 1967, 50, 496–501. [Google Scholar]
- Ahlgren, P.A.; Wood, J.R.; Goring, D.A.I. Fiber saturation point of various morphological subdivisions of Douglas-fir and aspen wood. Wood Sci. Technol. 1972, 6, 81–84. [Google Scholar] [CrossRef]
- Feist, W.C.; Tarkow, H. A new procedure for measuring fiber saturation points. For. Prod. J. 1967, 17, 65–68. [Google Scholar]
- Beck, G.; Thybring, E.E.; Thygesen, L.G.; Hill, C. Characterization of moisture in acetylated and propionylated radiata pine using low-field nuclear magnetic resonance (LFNMR) relaxometry. Holzforschung 2018, 72, 225. [Google Scholar] [CrossRef]
- Flournoy, D.S.; Kirk, T.K.; Highley, T.L. Wood decay by brown-rot fungi—Changes in pore structure and cell-wall volume. Holzforschung 1991, 45, 383–388. [Google Scholar] [CrossRef]
- Hill, C.A.S.; Forster, S.C.; Farahani, M.R.M.; Hale, M.D.C.; Ormondroyd, G.A.; Williams, G.R. An investigation of cell wall micropore blocking as a possible mechanism for the decay resistance of anhydride modified wood. Int. Biodeterior. Biodegrad. 2005, 55, 69–76. [Google Scholar] [CrossRef]
- Alince, B. Comments on porosity or swollen pulp fibers analyzed by solute-exclusion. Tappi 1991, 74, 200–202. [Google Scholar]
- Day, J.C.; Alince, B.; Robertson, A.A. The characterization of pore systems by macromolecular penetration. Cellul. Chem. Technol. 1979, 13, 317–326. [Google Scholar]
- Bonnet, M.; Courtier-Murias, D.; Faure, P.; Rodts, S.; Care, S. NMR determination of sorption isotherms in earlywood and latewood of Douglas fir. Identification of bound water components related to their local environment. Holzforschung 2017, 71, 481. [Google Scholar] [CrossRef]
- Fredriksson, M.; Thybring, E.E.; Zelinka, S.L. Artifacts in electrical measurements on wood caused by non-uniform moisture distributions. Holzforschung 2020. [Google Scholar] [CrossRef]
- Cox, J.; McDonald, P.J.; Gardiner, B.A. A study of water exchange in wood by means of 2D NMR relaxation correlation and exchange. Holzforschung 2010, 64, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Jeoh, T.; Karuna, N.; Weiss, N.D.; Thygesen, L.G. Two-dimensional 1H-Nuclear Magnetic Resonance relaxometry for understanding biomass recalcitrance. Acs Sustain. Chem. Eng. 2017, 5, 8785–8795. [Google Scholar] [CrossRef]
- Rostom, L.; Courtier-Murias, D.; Rodts, S.; Care, S. Investigation of the effect of aging on wood hygroscopicity by 2D 1H NMR relaxometry. Holzforschung 2020, 74, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Plaza Rodriguez, N.Z. Neutron Scattering Studies of Nano-Scale Wood-Water Interactions; University of Wisconsin—Madison: Madison, WI, USA, 2017. [Google Scholar]
- Zelinka, S.L.; Lambrecht, M.J.; Glass, S.V.; Wiedenhoeft, A.C.; Yelle, D.J. Examination of water phase transitions in Loblolly pine and cell wall components by differential scanning calorimetry. Thermochim. Acta 2012, 533, 39–45. [Google Scholar] [CrossRef]
- Kulasinski, K.; Guyer, R.; Derome, D.; Carmeliet, J. Water adsorption in wood microfibril-hemicellulose system: Role of the crystalline–amorphous interface. Biomacromolecules 2015, 16, 2972–2978. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.E.; Bizon, M. Changes in shrinkage and tangential compression strength of sugar maple below and above the fiber saturation point. Wood Fiber Sci. 1994, 26, 360–369. [Google Scholar]
- Almeida, G.; Hernandez, R.E. Changes in physical properties of yellow birch below and above the fiber saturation point. Wood Fiber Sci. 2006, 38, 74–83. [Google Scholar]
- Almeida, G.; Hernández, R.E. Changes in physical properties of tropical and temperate hardwoods below and above the fiber saturation point. Wood Sci. Technol. 2006, 40, 599–613. [Google Scholar] [CrossRef]
- Zauer, M.; Kretzschmar, J.; Großmann, L.; Pfriem, A.; Wagenführ, A. Analysis of the pore-size distribution and fiber saturation point of native and thermally modified wood using differential scanning calorimetry. Wood Sci. Technol. 2014, 48, 177–193. [Google Scholar] [CrossRef]
- Patera, A.; Derome, D.; Griffa, M.; Carmeliet, J. Hysteresis in swelling and in sorption of wood tissue. J. Struct. Biol. 2013, 182, 226–234. [Google Scholar] [CrossRef]
- Patera, A.; Van den Bulcke, J.; Boone, M.N.; Derome, D.; Carmeliet, J. Swelling interactions of earlywood and latewood across a growth ring: Global and local deformations. Wood Sci. Technol. 2018, 52, 91–114. [Google Scholar] [CrossRef]
- Derome, D.; Zillig, W.; Carmeliet, J. Variation of measured cross-sectional cell dimensions and calculated water vapor permeability across a single growth ring of spruce wood. Wood Sci. Technol. 2012, 46, 827–840. [Google Scholar] [CrossRef]
- Rafsanjani, A.; Stiefel, M.; Jefimovs, K.; Mokso, R.; Derome, D.; Carmeliet, J. Hygroscopic swelling and shrinkage of latewood cell wall micropillars reveal ultrastructural anisotropy. J. R. Soc. Interface 2014, 11. [Google Scholar] [CrossRef] [PubMed]
- Engqvist, J.; Hall, S.; Fredriksson, M. A miniaturised pressure plate cell for in-situ X-ray imaging of water distribution in wood. In Proceedings of the Annual Meeting of the Northern European Network on Wood Science and Engineering (WSE), Lund, Sweden, 9–10 October 2019; pp. 130–131. [Google Scholar]
- Plaza, N.Z.; Pingali, S.V.; Qian, S.; Heller, W.T.; Jakes, J.E. Informing the improvement of forest products durability using small angle neutron scattering. Cellulose 2016, 23, 1593–1607. [Google Scholar] [CrossRef]
- Zabler, S.; Paris, O.; Burgert, I.; Fratzl, P. Moisture changes in the plant cell wall force cellulose crystallites to deform. J. Struct. Biol. 2010, 171, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Masuda, M. Microscopic observation of transverse swelling of latewood tracheid: Effect of macroscopic/mesoscopic structure. J. Wood Sci. 2006, 52, 283–289. [Google Scholar] [CrossRef]
- Noack, D.; Schwab, E.; Bartz, A. Characteristics for a judgment of sorption and swelling behavior of wood. Wood Sci. Technol. 1973, 7, 218–236. [Google Scholar] [CrossRef]
- Keylwerth, R. Untersuchungen über freie und behinderte Quellung von Holz—Erste Mitteilung: Freie Quellung [Investigations on free and restraint swelling of wood—Part I: Free swelling]. Holz Als Roh- Und Werkst. 1962, 20, 252–259. [Google Scholar] [CrossRef]
- Arzola-Villegas, X.; Lakes, R.; Plaza, N.Z.; Jakes, J.E. Wood moisture-induced swelling at the cellular scale—Ab intra. Forests 2019, 10, 996. [Google Scholar] [CrossRef] [Green Version]
- Fredriksson, M.; Thybring, E.E. Scanning or desorption isotherms? Characterising sorption hysteresis of wood. Cellulose 2018, 25, 4477–4485. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Avramidis, S. Water sorption hysteresis in wood: I review and experimental patterns—Geometric characteristics of scanning curves. Holzforschung 2017, 71, 307–316. [Google Scholar] [CrossRef]
- Peralta, P.N.; Bangi, A.P. Modeling wood moisture sorption hysteresis based on similarity hypothesis. Part 1. Direct approach. Wood Fiber Sci. 1998, 30, 48–55. [Google Scholar]
- Ahlgren, L. Moisture Fixation in Porous Building Materials; Division of Building Technology, Lund Institute of Technology: Lund, Sweden, 1972. [Google Scholar]
- Salmén, L.; Larsson, P.A. On the origin of sorption hysteresis in cellulosic materials. Carbohydr. Polym. 2018, 182, 15–20. [Google Scholar] [CrossRef]
- Keating, B.A.; Hill, C.A.S.; Sun, D.; English, R.; Davies, P.; McCue, C. The water vapor sorption behavior of a galactomannan cellulose nanocomposite film analyzed using parallel exponential kinetics and the Kelvin–Voigt viscoelastic model. J. Appl. Polym. Sci. 2013, 129, 2352–2359. [Google Scholar] [CrossRef] [Green Version]
- Engelund, E.T.; Thygesen, L.G.; Svensson, S.; Hill, C.A.S. A critical discussion of the physics of wood-water interactions. Wood Sci. Technol. 2013, 47, 141–161. [Google Scholar] [CrossRef] [Green Version]
- Fortin, Y. Moisture Content-Matric Potential Relationship and Water Flow Properties of Wood at High Moisture Contents; University of British Columbia: Vancouver, BC, Canada, 1979. [Google Scholar] [CrossRef]
- Arevalo, R.; Hernandez, R.E. Influence of moisture sorption on swelling of mahogany (Swietenia macrophylla King) wood. Holzforschung 2001, 55, 590–594. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Aggarwal, P. Effect of moisture sorption state on transverse dimensional changes in wood. Holz Als Roh- Und Werkst. 2004, 62, 50–55. [Google Scholar] [CrossRef]
- Hartley, I.D.; Avramidis, S. Static dimensional changes of Sitka spruce and Western hemlock influenced by sorption conditions. J. Inst. Wood Sci. 1996, 14, 83–88. [Google Scholar]
- Hernandez, R.E. Influence of moisture sorption history on the swelling of Sugar maple wood and some tropical hardwoods. Wood Sci. Technol. 1993, 27, 337–345. [Google Scholar] [CrossRef]
- Seifert, J. On the sorption and swelling of wood and wood base materials – part 2: Swelling behavior of wood and wood base materials. Holz Als Roh- Und Werkst. 1972, 30, 294–303. [Google Scholar] [CrossRef]
- Thybring, E.E.; Glass, S.V.; Zelinka, S.L. Kinetics of water vapor sorption in wood cell walls: State of the art and research needs. Forests 2019, 10, 704. [Google Scholar] [CrossRef] [Green Version]
- Christensen, G.N. Kinetics of sorption of water vapour by wood. Aust. J. Appl. Sci. 1960, 11, 295–304. [Google Scholar]
- Murr, A. The relevance of water vapour transport for water vapour sorption experiments on small wooden samples. Transp. Porous Media 2019, 128, 385–404. [Google Scholar] [CrossRef] [Green Version]
- Thorell, A.; Wadsö, L. Determination of external mass transfer coefficients in dynamic sorption (DVS) measurements. Dry. Technol. 2018, 36, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.W.; Hart, C.A. Water vapor sorption rates by wood cell walls. Wood Fiber Sci. 1970, 1, 270–282. [Google Scholar]
- Christensen, G.N.; Kelsey, K.E. The rate of sorption of water vapor by wood. Holz Als Roh- Und Werkst. 1959, 17, 178–188. [Google Scholar] [CrossRef]
- Thybring, E.E.; Boardman, C.R.; Glass, S.V.; Zelinka, S.L. The parallel exponential kinetics model is unfit to characterize moisture sorption kinetics in cellulosic materials. Cellulose 2019, 26, 723–735. [Google Scholar] [CrossRef]
- Glass, S.V.; Zelinka, S.L.; Thybring, E.E. Exponential decay analysis: A flexible, robust, data-driven methodology for analyzing sorption kinetic data. Cellulose 2021, 28, 153–174. [Google Scholar] [CrossRef]
- Willems, W. Thermally limited wood moisture changes: Relevance for dynamic vapour sorption experiments. Wood Sci. Technol. 2017, 51, 751–770. [Google Scholar] [CrossRef]
- Willems, W. Heuristic study on the interaction between heat exchange and slow relaxation processes during wood moisture content changes. Holzforschung 2020. [Google Scholar] [CrossRef]
- Sonderegger, W.; Vecellio, M.; Zwicker, P.; Niemz, P. Combined bound water and water vapour diffusion of Norway spruce and European beech in and between the principal anatomical directions. Holzforschung 2011, 65, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Nylinder, P.; Hägglund, E. Ståndorts- och trädegenskapers inverkan på utbyte och kvalitet vid framställning av sulfitmassa av gran [The influence of stand and tree properties on yield and quality of sulphite pulp of Swedish spruce (Picea excelsa)]. Medd. Från Statens Skogsforskningsinstitut 1954, 44, 1–184. [Google Scholar]
- Bannan, M.W. The length, tangential diameter, and length/width ratio of conifer tracheids. Can. J. Bot. 1965, 43, 967–984. [Google Scholar] [CrossRef]
- Hozjan, T.; Svensson, S. Theoretical analysis of moisture transport in wood as an open porous hygroscopic material. Holzforschung 2011, 65, 97–102. [Google Scholar] [CrossRef]
- Kulasinski, K.; Keten, S.; Churakov, S.V.; Guyer, R.; Carmeliet, J.; Derome, D. Molecular mechanism of moisture-induced transition in amorphous cellulose. ACS Macro Lett. 2014, 1037–1040. [Google Scholar] [CrossRef]
- Crank, J. Diffusion coefficients in solids, their measurement and significance. Discuss. Faraday Soc. 1957, 99–104. [Google Scholar] [CrossRef]
- Zhan, T.; Sun, F.; Lv, C.; He, Q.; Wang, X.; Xu, K.; Zhang, Y.; Cai, L. Evaluation of moisture diffusion in lignocellulosic biomass in steady and unsteady states by a dynamic vapor sorption apparatus. Holzforschung 2019, 73, 1113–1119. [Google Scholar] [CrossRef]
- Chen, C.; Kuang, Y.; Zhu, S.; Burgert, I.; Keplinger, T.; Gong, A.; Li, T.; Berglund, L.; Eichhorn, S.J.; Hu, L. Structure–property–function relationships of natural and engineered wood. Nat. Rev. Mater. 2020, 5, 642–666. [Google Scholar] [CrossRef]
- Rowell, R.M. Chemical modification of wood. In Handbook of Wood Chemistry and Wood Composites, 1st ed.; Rowell, R.M., Ed.; CRC Press: London, UK, 2005; pp. 381–420. [Google Scholar]
- Hill, C.A.S.; Jones, D.; Strickland, G.; Cetin, N.S. Kinetic and mechanistic aspects of the acetylation of wood with acetic anhydride. Holzforschung 1998, 52, 623–629. [Google Scholar] [CrossRef]
- Williams, F.C.; Hale, M.D. The resistance of wood chemically modified with isocyanates: The role of moisture content in decay suppression. Int. Biodeterior. Biodegrad. 2003, 52, 215–221. [Google Scholar] [CrossRef]
- Thygesen, L.G.; Ehmcke, G.; Barsberg, S.; Pilgård, A. Furfurylation result of Radiata pine depends on the solvent. Wood Sci. Technol. 2020, 54, 929–942. [Google Scholar] [CrossRef]
- Tarkow, H.; Stamm, A.J. Effect of formaldehyde treatments upon the dimensional stabilization of wood. J. For. Prod. Res. Soc. 1953, 3, 33–37. [Google Scholar]
- Grönquist, P.; Frey, M.; Keplinger, T.; Burgert, I. Mesoporosity of delignified wood investigated by water vapor sorption. ACS Omega 2019, 4, 12425–12431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Thybring, E.E.; Fredriksson, M.; Ma, E.; Cao, J.; Digaitis, R.; Thygesen, L.G. Effects of changes in biopolymer composition on moisture in acetylated wood. Forests 2020, 11. [Google Scholar] [CrossRef]
- Keplinger, T.; Cabane, E.; Chanana, M.; Hass, P.; Merk, V.; Gierlinger, N.; Burgert, I. A versatile strategy for grafting polymers to wood cell walls. Acta Biomater. 2015, 11, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Digaitis, R.; Thygesen, L.G.; Thybring, E.E.; Fredriksson, M. Targeted acetylation of Norway spruce tissue and its effect on moisture states in wood. In Proceedings of the Annual Meeting of the Northern European Network on Wood Science and Engineering (WSE), Lund, Sweden, 9–10 October 2019; pp. 13–15. [Google Scholar]
- Beck, G.; Strohbusch, S.; Larnøy, E.; Militz, H.; Hill, C. Accessibility of hydroxyl groups in anhydride modified wood as measured by deuterium exchange and saponification. Holzforschung 2017, 72, 17–23. [Google Scholar] [CrossRef]
- Altgen, M.; Hofmann, T.; Militz, H. Wood moisture content during the thermal modification process affects the improvement in hygroscopicity of Scots pine sapwood. Wood Sci. Technol. 2016, 50, 1181–1195. [Google Scholar] [CrossRef]
- Emmerich, L.; Altgen, M.; Rautkari, L.; Militz, H. Sorption behavior and hydroxyl accessibility of wood treated with different cyclic N-methylol compounds. J. Mater. Sci. 2020, 55, 16561–16575. [Google Scholar] [CrossRef]
- Popescu, C.M.; Hill, C.A.S.; Curling, S.; Ormondroyd, G.A.; Xie, Y. The water vapour sorption behaviour of acetylated birch wood: How acetylation affects the sorption isotherm and accessible hydroxyl content. J. Mater. Sci. 2014, 49, 2362–2371. [Google Scholar] [CrossRef]
- Elder, T.; Houtman, C. Time-domain NMR study of the drying of hemicellulose extracted aspen (Populus tremuloides Michx.). Holzforschung 2013, 67, 405. [Google Scholar] [CrossRef]
- Thygesen, L.G.; Elder, T. Moisture in untreated, acetylated, and furfurylated Norway spruce studied during drying using Time Domain NMR. Wood Fiber Sci. 2008, 40, 309–320. [Google Scholar]
- Bryne, L.E.; Wålinder, M.E.P. Ageing of modified wood. Part 1: Wetting properties of acetylated, furfurylated, and thermally modified wood. Holzforschung 2010, 64, 295–304. [Google Scholar] [CrossRef]
- Stone, J.E.; Scallan, A.M. A structural model for the cell wall of water-swollen wood pulp fibres based on their accessibility to macromolecules. Cellul. Chem. Technol. 1968, 2, 343–358. [Google Scholar]
- Akitsu, H.; Norimoto, M.; Morooka, T.; Rowell, R.M. Effect of humidity on vibrational properties of chemically modified wood. Wood Fiber Sci. 1993, 25, 250–260. [Google Scholar]
- Plaza, N.Z. On the experimental assessment of the molecular-scale interactions between wood and water. Forests 2019, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Himmel, S.; Mai, C. Water vapour sorption of wood modified by acetylation and formalisation—Analysed by a sorption kinetics model and thermodynamic considerations. Holzforschung 2016, 70, 203–213. [Google Scholar] [CrossRef]
- Xie, Y.J.; Hill, C.A.S.; Xiao, Z.F.; Mai, C.; Militz, H. Dynamic water vapour sorption properties of wood treated with glutaraldehyde. Wood Sci. Technol. 2011, 45, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Olek, W.; Majka, J.; Czajkowski, L. Sorption isotherms of thermally modified wood. Holzforschung 2013, 67, 183–191. [Google Scholar] [CrossRef]
- Majka, J.; Czajkowski, Ł.; Olek, W. Effects of cyclic changes in relative humidity on the sorption hysteresis of thermally modified spruce wood. BioResources 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Östberg, G.; Salmén, L.; Terlecki, J. Softening temperature of moist wood measured by differential scanning calorimetry. Holzforschung 1990, 44, 223–225. [Google Scholar] [CrossRef]
- Jakes, J.E.; Hunt, C.G.; Zelinka, S.L.; Ciesielski, P.N.; Plaza, N.Z. Effects of moisture on diffusion in unmodified wood cell walls: A phenomenological polymer science approach. Forests 2019, 10, 1084. [Google Scholar] [CrossRef] [Green Version]
- Björkman, A.; Salmén, L. Studies on solid wood. II. The influence of chemical modifications on viscoelastic properties. Cellul. Chem. Technol. 2000, 34, 7–20. [Google Scholar]
- Wagner, L.; Almkvist, G.; Bader, T.K.; Bjurhager, I.; Rautkari, L.; Gamstedt, E.K. The influence of chemical degradation and polyethylene glycol on moisture-dependent cell wall properties of archeological wooden objects: A case study of the Vasa shipwreck. Wood Sci. Technol. 2016, 50, 1103–1123. [Google Scholar] [CrossRef]
- Jäger, A.; Bader, T.; Hofstetter, K.; Eberhardsteiner, J. The relation between indentation modulus, microfibril angle, and elastic properties of wood cell walls. Compos. Part A 2011, 42, 677–685. [Google Scholar] [CrossRef]
- Stamm, A.J. Bound water diffusion into wood in the fiber direction. For. Prod. J. 1959, 9, 27–32. [Google Scholar]
- Keplinger, T.; Cabane, E.; Berg, J.K.; Segmehl, J.S.; Bock, P.; Burgert, I. Smart hierarchical bio-based materials by formation of stimuli-responsive hydrogels inside the microporous structure of wood. Adv. Mater. Interfaces 2016, 3, 1600233. [Google Scholar] [CrossRef]
- Chen, H.; Ginzburg, V.V.; Yang, J.; Yang, Y.; Liu, W.; Huang, Y.; Du, L.; Chen, B. Thermal conductivity of polymer-based composites: Fundamentals and applications. Prog. Polym. Sci. 2016, 59, 41–85. [Google Scholar] [CrossRef]
- Rousset, P.; Perre, P.; Girard, P. Modification of mass transfer properties in poplar wood (P-robusta) by a thermal treatment at high temperature. Holz Als Roh- Und Werkst. 2004, 62, 113–119. [Google Scholar] [CrossRef]
- Rüggeberg, M.; Burgert, I. Bio-inspired wooden actuators for large scale applications. PLoS ONE 2015, 10, e0120718. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.; Vailati, C.; Menges, A.; Rüggeberg, M. Hygroscopically actuated wood elements for weather responsive and self-forming building parts—Facilitating upscaling and complex shape changes. Constr. Build. Mater. 2018, 165, 782–791. [Google Scholar] [CrossRef]
Figure 1.
Simplified overview of wood structure on different length scales: Wood macrostructure with annual rings; porous microstructure with pits, which are pathways through cell walls that connect lumina (singular: lumen) of adjacent cells; and cell wall material made from cellulose, hemicelluloses, and lignin.
Figure 1.
Simplified overview of wood structure on different length scales: Wood macrostructure with annual rings; porous microstructure with pits, which are pathways through cell walls that connect lumina (singular: lumen) of adjacent cells; and cell wall material made from cellulose, hemicelluloses, and lignin.

Figure 2.
Moisture in wood can be found in both the porous capillary structure (A) and the solid cell wall material (B), where the water molecules interact with hydroxyls on the cell wall polymers (C). Diagrams depict equilibrium moisture content as a function of relative humidity (sorption isotherms) in both the hygroscopic (left) and over-hygroscopic (right) moisture ranges. In the hygroscopic moisture range, changes in moisture content are dominated by changes in moisture content (blue spheres) within the cell wall material (D,E), whereas changes in moisture content (blue filling in macro-voids) in the capillary structure (F,G) becomes dominant in the over-hygroscopic moisture range. Moisture history dependence is indicated by arrows indicating equilibrium reached by increasing relative humidity (absorption isotherm) and decreasing relative humidity (desorption isotherm).
Figure 2.
Moisture in wood can be found in both the porous capillary structure (A) and the solid cell wall material (B), where the water molecules interact with hydroxyls on the cell wall polymers (C). Diagrams depict equilibrium moisture content as a function of relative humidity (sorption isotherms) in both the hygroscopic (left) and over-hygroscopic (right) moisture ranges. In the hygroscopic moisture range, changes in moisture content are dominated by changes in moisture content (blue spheres) within the cell wall material (D,E), whereas changes in moisture content (blue filling in macro-voids) in the capillary structure (F,G) becomes dominant in the over-hygroscopic moisture range. Moisture history dependence is indicated by arrows indicating equilibrium reached by increasing relative humidity (absorption isotherm) and decreasing relative humidity (desorption isotherm).

Figure 3.
Sorption kinetics in wood involves a change in moisture content and temperature over time. Here it is illustrated by water vapor absorption, which results in an increase in mass (moisture content, blue curve), dimensions (swelling, black arrows), and temperature (red arrows and curve) in the approach to equilibrium. During the absorption process, water vapor diffuses through the porous wood microstructure (blue arrows), where it goes through lumina (A), pits (B), and cell walls, while constantly exchanging water molecules (C) between the water vapor and the moisture within the cell walls.
Figure 3.
Sorption kinetics in wood involves a change in moisture content and temperature over time. Here it is illustrated by water vapor absorption, which results in an increase in mass (moisture content, blue curve), dimensions (swelling, black arrows), and temperature (red arrows and curve) in the approach to equilibrium. During the absorption process, water vapor diffuses through the porous wood microstructure (blue arrows), where it goes through lumina (A), pits (B), and cell walls, while constantly exchanging water molecules (C) between the water vapor and the moisture within the cell walls.

Figure 4.
Types of wood modifications affecting the cellulose (yellow), hemicelluloses (purple), and lignin (brown) of the cell wall material as well as the interactions with water (blue spheres). Hydroxyl removal reduces the number of hydroxyls (OH); bulking adds volume within cell walls; cross-linking covalently joins adjacent wood components; component removal selectively removes or reduces a specific component (here illustrated by delignification, i.e., removal of lignin); and thermal modification involves both component removal (primarily hemicelluloses) and hydroxyl removal, and cross-linking during the treatment.
Figure 4.
Types of wood modifications affecting the cellulose (yellow), hemicelluloses (purple), and lignin (brown) of the cell wall material as well as the interactions with water (blue spheres). Hydroxyl removal reduces the number of hydroxyls (OH); bulking adds volume within cell walls; cross-linking covalently joins adjacent wood components; component removal selectively removes or reduces a specific component (here illustrated by delignification, i.e., removal of lignin); and thermal modification involves both component removal (primarily hemicelluloses) and hydroxyl removal, and cross-linking during the treatment.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Thybring, E.E.; Fredriksson, M. Wood Modification as a Tool to Understand Moisture in Wood. Forests 2021, 12, 372. https://doi.org/10.3390/f12030372
AMA Style
Thybring EE, Fredriksson M. Wood Modification as a Tool to Understand Moisture in Wood. Forests. 2021; 12(3):372. https://doi.org/10.3390/f12030372
Chicago/Turabian StyleThybring, Emil Engelund, and Maria Fredriksson. 2021. "Wood Modification as a Tool to Understand Moisture in Wood" Forests 12, no. 3: 372. https://doi.org/10.3390/f12030372
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.