1. Introduction
There is growing interest in efficient year-round production of high-quality forest tree seedlings under semi-controlled or controlled conditions prior to planting in the field. Recent years’ developments in light-emitting diode (LED) technology have increased the efforts in tailoring the light climate for production of plants with desirable physiological traits. However, the production of high-quality forest tree seedlings using such lighting systems requires improved knowledge about the responses of the plants to the light and temperature climate during the growth–dormancy cycling.
For a wide range of tree species of the boreal and temperate zone, phenological transitions marking the end of the growing season occur in response to shifting light conditions. Shortening of the photoperiod (short days; SD) to a day length shorter than a critical one, has long been known to induce growth cessation and formation of dormant winter buds [
1,
2,
3,
4,
5]. The critical photoperiod is also well known to increase with higher northern latitude of origin [
1,
2,
3,
4,
5].
Furthermore, a range of studies has shown that light quality (spectral composition) influences autumn phenology in temperate and boreal trees. In seedlings of Norway spruce (
Picea abies (L.) H. Karst.), silver birch
(Betula pendula Roth) and bay willow (
Salix pentandra L.), sustained growth depends on the presence of sufficient FR photons (700–800 nm) in the light spectrum [
6,
7,
8,
9,
10,
11]. The requirement for FR light depends on latitudinal origin, and northern provenances require higher irradiances of FR than more southern ones for sustained shoot elongation [
10,
11,
12,
13].
Other parts of the light spectrum have been shown to affect tree growth, including interactions between different wavelengths. A study using narrow-bandwidth LEDs with wavelengths corresponding to the absorption maxima of the relevant photoreceptors, indicated that a red R: FR light ratio of 1 (peaking at 660 and 730 nm, respectively) provided as day extension was more efficient in preventing bud set in seedlings of northern provenances of Norway spruce than FR only (provenances from 59 °N (Halden), 64 °N (Snåsa) and 66 °N (Rana); all from Norway) [
10]. Also, day extension with R light at 0.6–18 µmol m
−2 s
−1 (0.1–3.3 W m
−2; lower not tested) or B light (peaking at 460 nm) at 6 and 13 µmol m
−2 s
−1 (1.5 and 3.3 W m
−2; lower not tested) delayed bud set, but did not prevent it, with more delay in southern than northern provenances [
10,
11]. The different provenances in these studies showed different sensitivity to different irradiances during day extension with R light [
10,
11]. When the R light was provided for 12 h following the main 12 h light period from fluorescent tubes, the southernmost Halden provenance showed delayed bud set from 0.6 µmol m
−2 s
−1 (0.1 W m
−2) of R light and the intermediate Snåsa provenance from 9 µmol m
−2 s
−1 (1.7 W m
−2), while all plants of the northernmost Rana provenance developed terminal buds [
10]. When the R light was given both during the 12 h main light period and as a 12 h day extension, even the Rana plants showed delayed bud set from 10 µmol m
−2 s
−1 (1.8 W m
−2) [
10]. Furthermore, a night interruption with R light at 22 µmol m
−2 s
−1 delayed or reduced formation of terminal buds in Norway spruce seedlings with more delay in a more southern (61 °N) than northern (64 °N) provenance [
14]. Also, a night break with combined R and FR light (22 µmol m
−2 s
−1 and 3.3 µmol m
−2 s
−1, respectively) was more efficient in preventing bud set than the R light only [
14].
In Subalpine fir (
Abies lasiocarpa (Hook.) Nutt.), day extension with FR light at 42 µmol m
−2 s
−1 (7 W m
−2) increased shoot elongation and delayed bud set, although it was not generally prevented [
15]. In Scots pine (
Pinus sylvestris L.) seedlings, FR-enriched light spectra enhanced shoot and needle elongation, increased shoot dry mass and stem diameter, and stimulated secondary needle formation [
16,
17]. Furthermore, addition of FR light to supplementary B and R light increased the number of branches in Norway spruce seedlings but not in Scots pine, but the effect of FR light on shoot elongation was less clear in Norway spruce than Scots pine [
17].
Although the photoperiod and light quality are considered the primary environmental cues controlling phenological transitions in trees, temperature has been shown to modulate their timing references in [
4,
5,
18,
19,
20]. In a number of deciduous and coniferous tree species grown under controlled conditions, higher temperature under SD was shown to accelerate bud set and development of deep dormancy, as compared to lower temperature [
21,
22,
23,
24,
25,
26]. Oppositely, field studies of
Populus have shown a delay in bud development at higher compared to lower autumn temperature [
27,
28,
29]. The inconsistency of temperature effects between field experiments and experiments under controlled conditions may be due to climatic differences, including different light spectra. Under field conditions, photoperiod, light quality, irradiance and temperature shift gradually, while under controlled conditions, shifts in light parameters and temperature are typically more abrupt or constant conditions are used.
In Norway spruce, bud set in response to SD is associated with strong upregulation of the transcript level of the
FLOWERING LOCUS T-TERMINAL FLOWER I-LIKE 2 (
PaFTL2) gene, and bud burst with decreased
FTL2 expression [
11,
30,
31,
32]. Similar results were found for Scots pine [
33]. Furthermore, for Norway spruce, very low transcript levels were reported under day extension with growth-sustaining FR light [
11]. That study also demonstrated some accumulation of the
PaFTL2 transcript under day extension with R or B light treatments at 18 and 13 µmol m
−2 s
−1, respectively (3.3 W m
−2). Consistent with delayed growth cessation and bud set under these conditions compared to SD, the
FTL2 transcript levels then accumulated more slowly and to lower levels than under SD. These results, and the notion that
FTL2-overexpression in Norway spruce causes early bud set [
32], indicate that
FTL2 acts as a growth inhibitor. The degree and rate of its accumulation seem to depend on the strength of the signal leading to growth cessation and bud set [
10,
11,
30,
31,
32].
In
Populus species, a
CONSTANS (
CO)-
FLOWERING LOCUS T (
FT)-regulon is involved in control of winter bud formation [
34]. A high
CO-expression at the end of a long day induces
FT-expression and thus sustained elongation growth in such species [
34]. In Norway spruce, no such CO-FT regulon has been found, but two
CO-LIKE (
COL) genes were characterised;
PaCOL1 and
PaCOL2 [
35]. Furthermore, a homologous gene to a floral integrator in
Arabidopsis thaliana ((L.) Heynh.),
SUPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (
PaSOC1), was identified in Norway spruce [
11,
36]. The
PaCOL1,
PaCOL2 and
PaSOC1 genes showed decreased transcript levels under SD-induced growth cessation, as compared to long days (LD) [
11,
35]. These genes also had reduced expression under day-extension treatments with R or B light, which delay bud formation, as compared to growth-sustaining LD and FR light treatments [
11]. Similarly, two
SOC1-like genes in white spruce (
Picea glauca (Moench) Voss) showed reduced expression under SD but another
SOC1-like gene showed increased transcript levels in developing buds [
37].
To contribute to improved knowledge about responses of forest tree seedlings to light and temperature conditions, we aimed to investigate interactive effects of temperature and light quality during day-extension treatments on growth cessation, bud set and subsequent bud burst in Norway spruce seedlings. In this respect, day-extension treatments with B, R or FR light or different R:FR light ratios were provided to plants grown at 18 °C or 22/24 °C. Furthermore, to shed light on effects of the irradiance of the day-extension treatments, higher irradiances were used than those tested previously in the same provenance (Halden, 59 °N) [
10,
11]. In addition, we aimed to investigate the sensitivity of this and a more northern provenance (Rana, 66 °N) to interactive effects of temperature and photoperiod (SD, LD). The control of the winter bud formation by light quality and temperature was also addressed by analyses of the transcript levels of
PaFTL2,
PaCOL1-2 and
PaSOC1, which were previously shown to change under bud set.
2. Materials and Methods
2.1. Growth Chambers, Recordings of Environmental Conditions and Plant Fertilization
All experiments (pregrowing and experimental treatments) were performed in closed growth chambers manufactured by the Centre for plant research in controlled climate (SKP) at the Norwegian University of Life Sciences (NMBU, Ås, Norway). The temperature and relative air humidity (RH) were recorded and controlled using a Priva computer system (Priva, De Lier, The Netherlands). In all experiments, the %RH was adjusted to correspond to a water vapour deficit (vpd) of 0.5 kPa (detailed %RH below). The variation in temperature and %RH was ≤ 1 °C and ≤ 5%, respectively. The photosynthetic photon flux density (PPFD) at 400–740 nm during the main light phase was measured at the top of the plant canopy using a LI-COR quantum sensor model Li-250 (LI-COR, Lincoln, NE, USA) and the R:FR light ratio was measured by a Skye R:FR-sensor (Skye Instruments, Llandrindod Wells, UK). The variation was ≤ 5% for both the PPFD and R:FR ratio. Details about the day-extension treatments with different light qualities are described below.
The plants were watered as required and fertilized twice a week with a complete nutrition solution. This consisted of a mixture of Calcinit and Kristalon Indigo (Yara, Oslo, Norway) and contained 14.3 mM nitrogen, 1 mM phosphorus, 5.1 mM potassium and the other essential macro- and microelements, at an electrical conductivity (EC) of 1.5 mS cm−1, as measured with a ScanGrow EC meter (ScanGrow, Slangerup, Denmark).
2.2. Plant Materials and Pre-Growing Conditions
Seeds of Norway spruce from the provenance CØ1, originating from 59 °N latitude, in Halden, Norway (seed lot 98063; Skogfrøverket, Hamar, Norway) and the provenance P1, originating from 66 °N latitude, in Rana, Norway (seed lot 4145; Skogfrøverket) were used. Two seeds were sown in each pot (Vefi, Drammen, Norway) of 5.5 × 5.5 × 4.5 cm to ensure the presence of at least one plant since the germination rate was about 60%. The seeds were sown in a 3:1 mixture of fertilised peat (pH 6; S-soil, Hasselfors, Oslo, Norway) and perlite. After germination, thinning to one plant per pot was done using scissors to avoid root disturbance. The seedlings were grown for 7 weeks before starting the experimental treatments.
The plants were grown at a constant temperature of 18 °C and RH of 76%. LD of 24 h was used to ensure a photoperiod longer that the critical one, allowing continued growth for the northern provenances. In experiment 1–4, a PPFD of 200 μmol m−2 s−1 from high pressure sodium (HPS) lamps (Gavita 400W HPS GAN 400 AL, Gavita, Aalsmeer, Netherlands) was used, with the R:FR light ratio adjusted to 1.7 with low-intensity light from incandescent lamps (Narva 60W, Germany and Philips Electronics, Amsterdam, Netherlands). In experiment 5–6, a PPFD of 250 μmol m−2 s−1 μmol was applied with quartz metal halide lamps (Master HPI-T Plus 400W/645 E40 1SL, Phillips, Amsterdam, Netherlands). In these two experiments, the plants were placed on 50 cm × 50 cm trolleys with the HPI-T lamps mounted at 1 m distance above the trolleys. A R:FR ratio of 1.8 was then achieved using incandescent lamps.
2.3. Experimental Design and Conditions
2.3.1. The Main Light Phase Conditions during the Experimental Period
Table 1 provides an overview of the experimental conditions, including the pregrowing conditions, in the six different experiments performed. Experiment 1–2 investigated interactive effects of temperature and photoperiod in the Halden and Rana provenances. Experiment 3–6 tested combined effects of temperature and light quality in the Halden provenance.
In all experiments, the main light period was provided for 12 h daily from HPS lamps (Lucalox 400W, General electric, New York, NY, USA). In experiment 1–2, the main light period had a PPFD of 200 μmol m−2 s−1, whereas in experiment 3–4, a PPFD of 160 μmol m−2 s−1 was used. The R:FR light ratio during this period was adjusted with incandescent lamps to 1.7 and 2 in experiments 1–2 and 3–4, respectively. The reasons for these differences were the following: Experiment 1–2 was performed in small (not walk-in) growth chambers enabling 200 μmol m−2 s−1 and a R:FR ratio of 1.7. Experiment 3 and 4 were performed in larger walk-in chambers, where day extension with different light qualities was provided on 50 cm × 50 cm trollies enclosed by reflecting plastic curtains (described below). In both chamber types, the lamps used for the main photoperiod in experiment 1–4 were mounted in the ceiling of the chambers. The highest irradiance that could be obtained at the top of the canopy in each plastic-enclosed trolley in experiment 3–4 was 160 μmol m−2 s−1 and the lowest possible R:FR ratio obtained was 2.0 (incandescent lamps above the plastic curtains). In experiments 5 and 6, the HPS lamps were mounted 1 m above each trolley to enable higher irradiance than possible with lamps at the top of the growth chamber. Thus, in experiment 5–6, a PPFD of 260 μmol m−2 s−1 was used. To ensure sufficient FR light in the spectrum of the main light phase for the northern provenance used, the R:FR ratio was modified to 2.5 (lower was not possible) using five FR light-emitting diode panels (LEDs) with 5 diodes in each for each trolley (1 m above the trolleys), with a band width between 680 nm and 750 nm and a maximum at 725 nm. Since the main light was closer to the plants in these two experiments, heat-generating incandescent lamps could not be used for the adjustment of the R:FR ratio during the main light phase.
2.3.2. Temperature and Day-Extension Treatments during the Experimental Period
In experiments 1 and 2, seedlings from the Halden and Rana provenances were exposed to either SD of 12 h or LD of 24 h photoperiod using a main light period of 12 h, followed by 12 h darkness or day extension to 24 h with low-intensity incandescent light, respectively (light sources described above). For each photoperiod, three subsets of seedlings, with 20 plants in each, were exposed to 12, 18 or 24 °C. Due to chamber capacity, the experiments with the two provenances were performed in sequence, using the same 6 chambers for both experiments.
In experiment 3–6, different subsets of seedlings, with 18–20 plants from the Halden provenance in each treatment, were exposed to day extensions with different light qualities from LEDs (Philips GreenPower LED research module, Phillips, Amsterdam, Netherlands) at 18 °C or 22 °C in experiment 3, and at 18 °C or 24 °C in experiment 4–6. The idea was to test temperatures commonly occurring during the summer in nursery greenhouses in Scandinavia. The different light treatments were given on individual trolleys properly isolated with non-transparent, reflecting plastic curtains. The airflow in the chambers came from below, ensuring equal temperature in all trolleys. The different temperature treatments in each experiment were given in separate growth chambers.
The different light qualities were applied as a 12 h extension of the 12 h main daily light period. In addition, an overlap between the LED light and the main light phase of 30 min at the start and end of the main photoperiod was used to ensure that there was no gap in the lighting. A SD treatment with 12 h darkness following the 12 h main light period was also established as a control treatment (no day extension). For all light quality treatments, an equal amount of energy, i.e., 7 W m
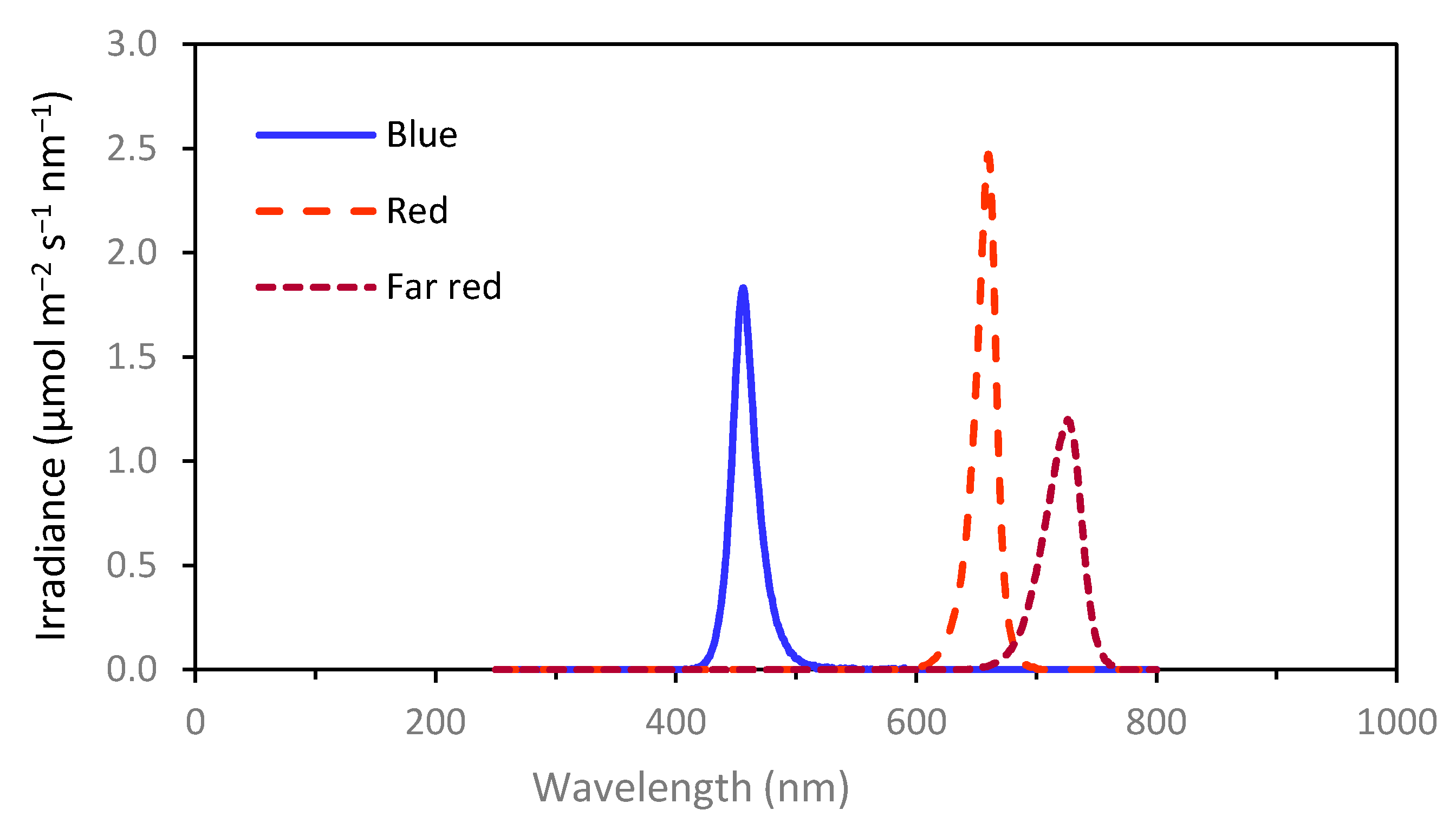
−2, was used, as measured at the top of the canopy with a SD2000 Miniature Fiber Optic Spectrometer (Ocean Optics, Orlando, FL, USA). This was calculated to correspond to 27, 38 and 42 µmol m
−2 s
−1 for the B, FR and R LED light, respectively. These calculations employed conversion factors obtained by later measurements of the LED spectra (
Figure 1) in W m
−2 and µmol m
−2 s
−1, using an Optronic model 756 spectroradiometer (Optronic laboratories, Orlando, FL, USA). The peak wavelengths were 460 nm for the B light, 660 nm for the R light and 730 nm for the FR light.
Experiments 3 and 4 included day-extension treatments with B, R or FR light. Experiment 5 employed day extension with FR light (phytochrome photostationary state; PPS = 0.24), R: FR ratio 0.5, 1 or 2 (PPS = 0.72, 0.78 and 0.81, respectively) or R light (PPS = 0.88). In experiment 6, day extension with the FR light, R: FR ratio 0.1, 0,2 or 0.5 (PPS = 0.4, 0.56 and 0.72, respectively) or the R light was used. The PPS value is defined as the ratio of the FR light-absorbing form of phytochrome (Pfr) to the total phytochrome, i.e., the R light-absorbing form of phytochrome (Pr) + Pfr at equilibrium. This was calculated according to [
38] by multiplying the irradiance at each wavelength with the relative absorption for each phytochrome form, with the absorption data derived from [
38]. In experiment 6, the idea was to provide a more equally distributed range of PPS values (orthogonal treatments) than in experiment 5.
In experiment 1–2, the RH values used were 64%, 76%, and 83% for 12 °C, 18 °C and 24 °C, respectively. In experiment 3–6, the RH values were 76%, 81% and 83% for 18 °C, 22 °C and 24 °C, respectively.
2.3.3. Environmental Conditions during Forcing to Bud Burst
After 6–8 weeks of exposure to the different temperature and light quality/photoperiod treatments in experiments 1, 2 and 4, seedlings were retransferred to growth chambers with environmental conditions as during the pregrowth; 18 °C, vpd 0.5 kPa, and 24 photoperiod (12 h main light period + 12 h low-intensity light from incandescent lamps) with a PPFD of 200 μmol m−2 s−1 and a R:FR ratio of 1.7 during the 12 h main light period.
2.4. Recordings of Growth and Stages of Bud Set and Bud Burst
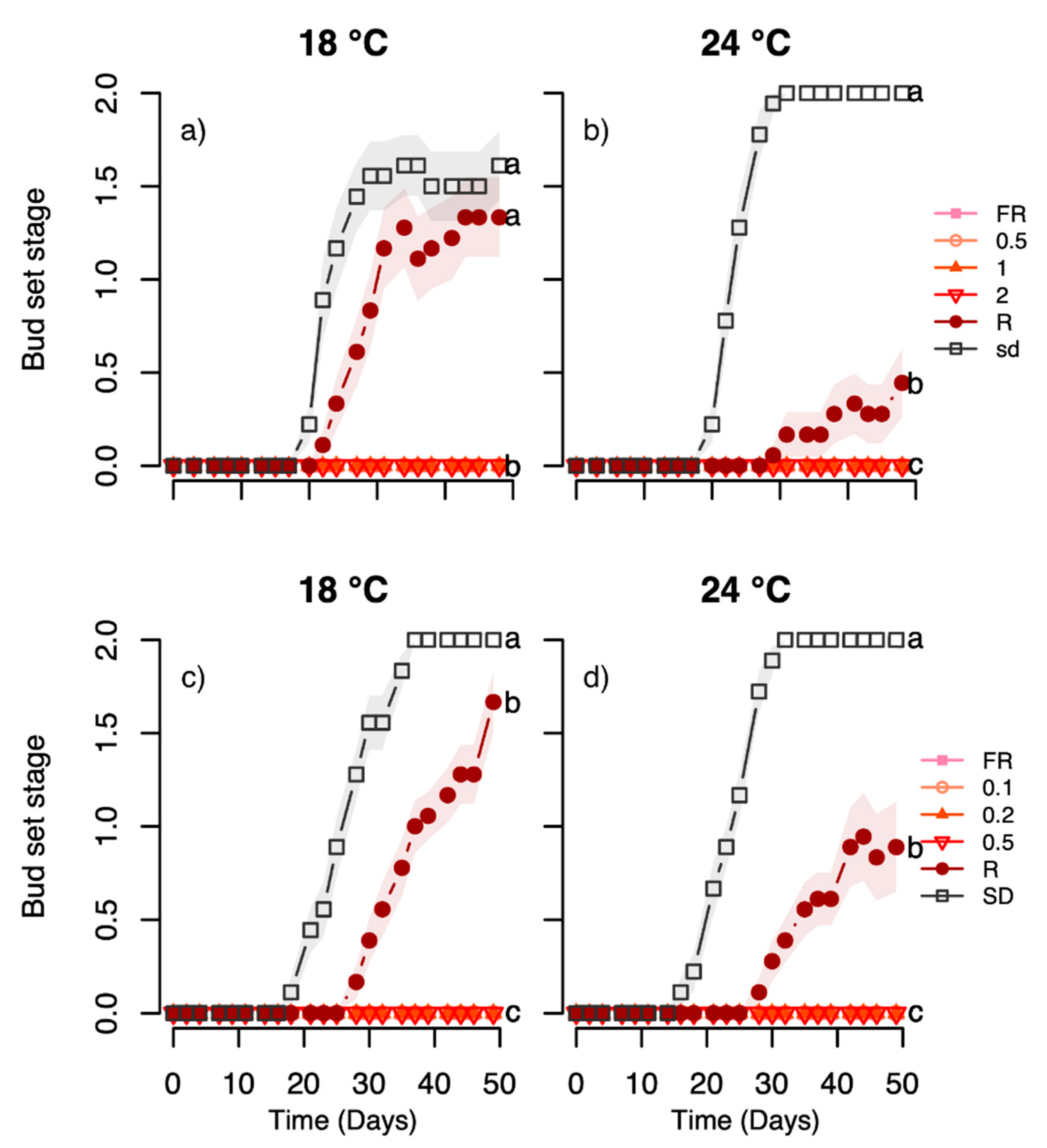
For each treatment, the height of each plant was measured once a week in all experiments, as total height from the base of the stem to the shoot apical meristem. The increase in height (cumulative shoot elongation) was calculated. Since height measurements for the day of transfer to the different treatments (day 0) are lacking for the Rana provenance in experiment 2, plant height is shown for both studied provenances in experiments 1 and 2. The winter bud development was recorded three times per week using codes where growing plants without buds were coded as 0, light green buds as 1 and brown buds as 2 (
Figure S1). It should be noted that in seedlings of Norway spruce, only vegetative buds are formed and that the generation time of this conifer species is about 25 years. In all experiments, 18–20 plants were used per treatment. After transfer of 18–20 plants per bud-inducing treatment to the forcing conditions, bud burst was recorded with the following bud stages; 2: closed bud, 1: opening in bud, 0: regrowth with needles protruding from the bud (
Figure S2).
2.5. Studies of Gene Expression
2.5.1. Sample Collection
To assess the effect of light quality/photoperiod and temperature treatments on the transcript levels of PaFTL2, PaCOL1, PaCOL2 and PaSOC1, 4 and 3 repeated samples were harvested per treatment in time courses from experiments 1–2 and 5, respectively. Each sample consisted of about 5 mm of the shoot tips without elongated needles from 3 pooled plants. In experiment 1–2, samples were harvested from all treatments and in experiment 5, from the R and FR light, the R: FR ratio 1 and the SD treatment. The samples were collected at the start of the experimental treatments (day 0), at day 5 (experiment 1–2) or 7 (experiment 5) and at day 51, 49 and 48 in experiments 1, 2 and 5, respectively. The samples were harvested in the middle of the photoperiod into liquid nitrogen (N2) and kept at −80 °C until analysis.
2.5.2. RNA Extraction and Purification and Analyses of Transcript Levels
Total RNA was extracted and purified using Masterpure Plant RNA Purification Kit (Epicentre, Madison, WI, USA) and PureLink RNA Mini kit (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s instructions. RNA quality was assessed using an Agilent 2100 Bioanalyzer with an RNA 144000 NanoKit (Agilent technologies, Palo Alto, CA, USA) and the quantity of isolated RNA was measured with a NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, Wilmington, DE, USA). cDNA was synthesized from 1 µg RNA in a 20 μL reaction volume using reverse transcriptase Superscript VILO cDNA Synthesis (Life Technologies, Thermo Fisher Scientific).
Gene-specific primers for the target genes
PaFTL2,
PaCOL1,
PaCOL2 and
PaSOC1 and the genes used as internal reference genes,
Paα-TUBULIN,
PaACTIN (
PaACT) and
PaELONGATION FACTOR 1α (
PaEF1α), were designed using the Primer3 software and the melting points and product lengths were assessed as described previously [11,39; primers in 11]. The transcript levels were analysed using a 7500 Fast Real-Time PCR system (Applied Biosystems, Foster City, CA, USA) and the relative transcript levels quantified on basis of the threshold cycle (CT) values from the quantitative PCR analyses, all according to [
39]. All samples were analysed in triplicate and the transcript levels of the target genes were normalised to the mean values of the internal reference genes that all had stable transcript levels under the experimental conditions, and thereafter related to the day 0 samples. A non-template control was run for each primer pair.
2.6. Statistical Analyses
To evaluate the effect of the photoperiod, temperature and provenance in experiments 1 and 2 on shoot height increase in the overall experiments (entire time courses), a three-way analysis of variance (ANOVA) was performed. To evaluate the effect of temperature and provenance on bud set stages and subsequent bud burst stages in these experiments, a two-way ANOVA was done. Furthermore, to evaluate the effect of light quality provided as day-extension and temperature on shoot elongation, bud set and bud burst stages in experiment 3–6, a two-way ANOVA was used. A linear model for the shoot elongation and a generalized linear model for the bud set and bud burst stages were used. Plant and time were treated as random effects according to [
40], because the recordings were performed on the same plants in time courses, which violates the assumption of independence of the data. To evaluate the effects of photoperiod, temperature and provenance on the transcript levels of
PaFTL2,
PaSOC1,
PaCOL1 and
PaCOL2 at the end of experiment 1–2, a three-way ANOVA was used. To assess the effects of light quality and temperature on transcript levels at the end of experiment 5, a two-way ANOVA was used.
To facilitate the visual comparison of the effects of the different treatments on shoot elongation, bud set and bud burst as well as transcript levels additional ANOVAs were performed for the values at the end of the experiments within one figure, followed by a Tukey’s post hoc test.
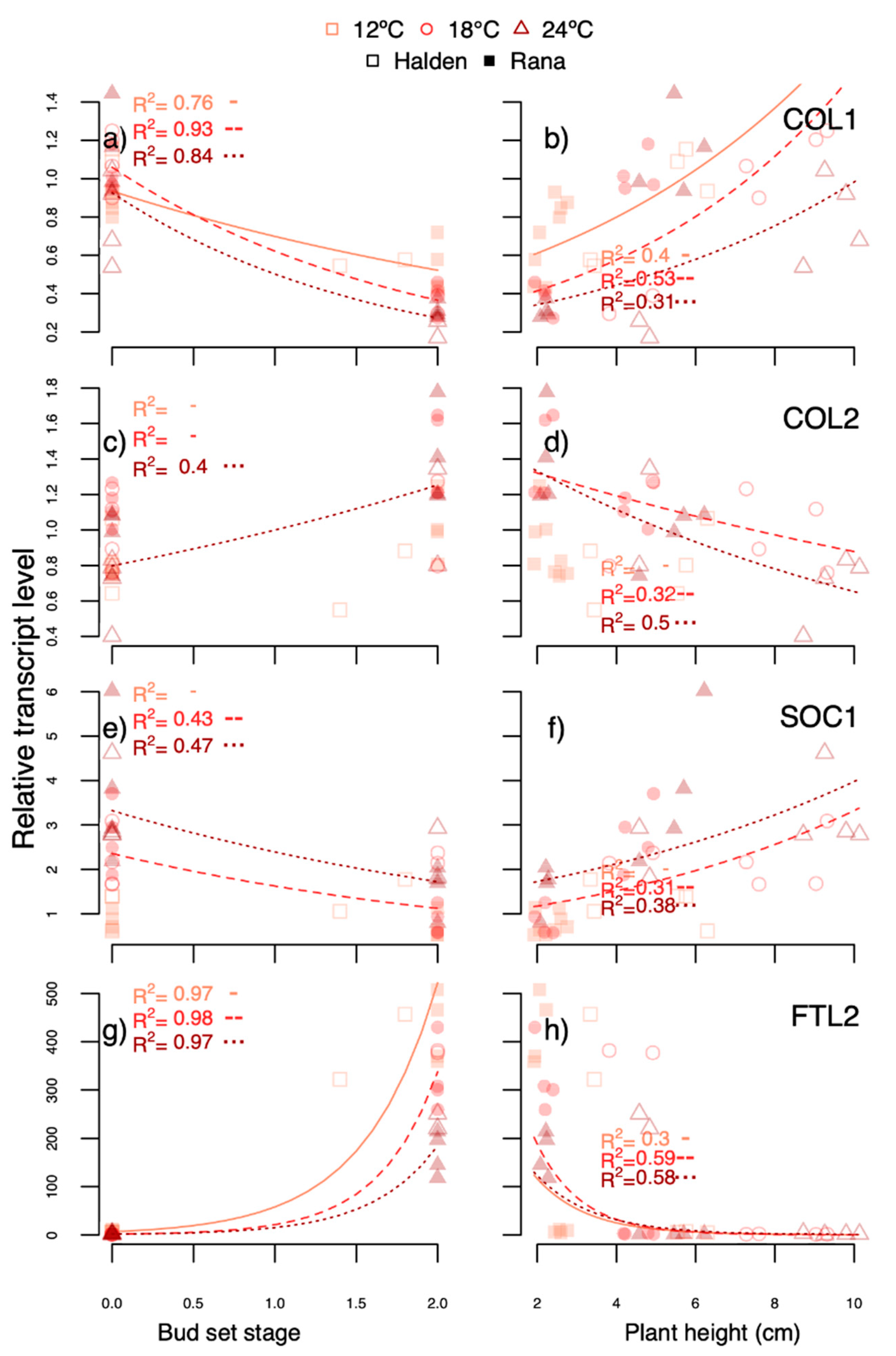
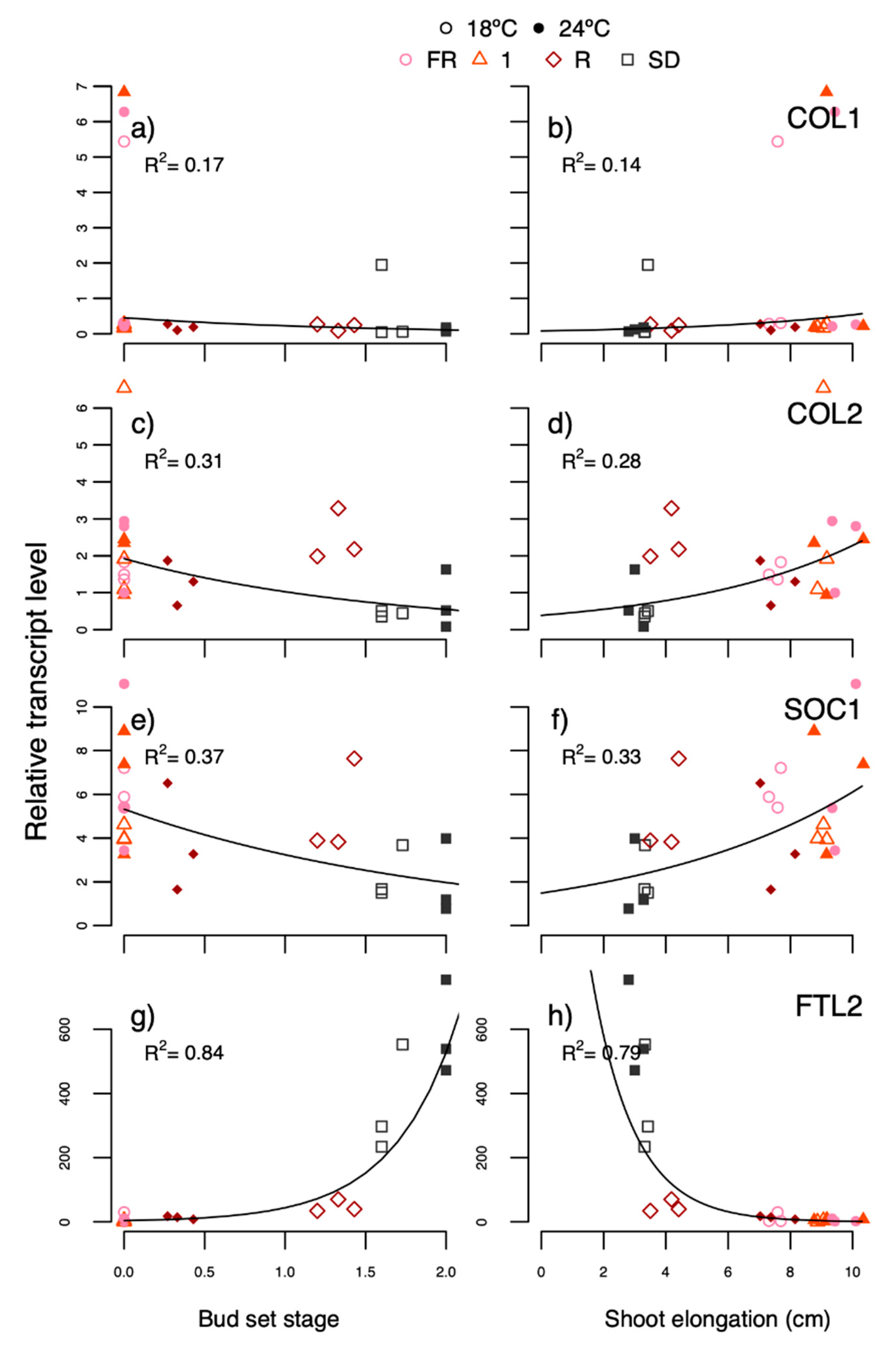
An exponential model was fitted using the bud set stage or shoot elongation for the different treatments to predict the transcript levels of the previously mentioned genes at the end of the experiments. Although the bud set category or shoot elongation in several cases were either not the best fit or not the only variable explaining the best fit for each regression, these were used for the visual representation, except that models with an
R2 value lower than 0.2 were not added. All statistical analyses were done using the R software, version 3.6 (
www.r-project.org; accessed on 24 December 2020).
4. Discussion
The light climate, particularly photoperiod, is well known to affect the growth–dormancy cycling in seedlings of forest trees. However, climatic regulation of coniferous tree growth has been little studied in terms of interactive effects between climatic parameters. Here we provide information about interactive effects of temperature and light quality under day extensions, as compared to SD, on growth and terminal, vegetative bud development as well as subsequent bud burst in young seedlings of Norway spruce.
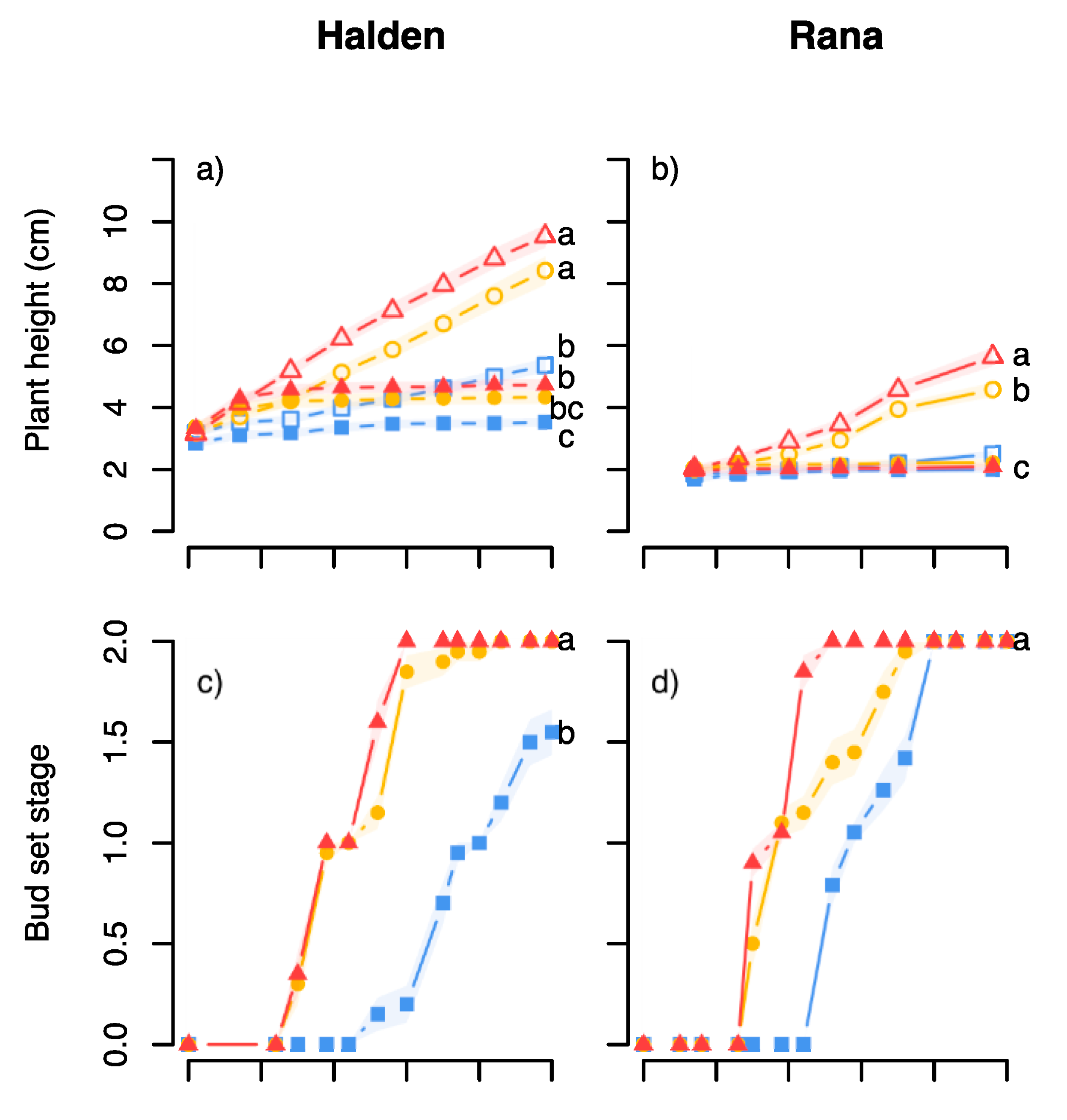
Our trials evaluating sensitivity to temperature under SD in the Halden and Rana provenances from 59 °N and 66 °N, respectively, revealed that these provenances had different responsiveness to temperature for bud phenology and height growth. Compared to the northern provenance, the more southern one appeared more sensitive to temperature, both in terms of stronger height growth and the duration of terminal bud development (
Figure 2a–d). The more rapid bud development and less growth in the northern provenance at all temperatures, even under LD of a 24 h photoperiod, may reflect that “Rana” as one of the world’s northernmost Norway spruce provenances, shows adaptation to the short, usually cool growing season at high northern latitudes by investing more resources to survival rather than growth. More rapid bud set in northern than southern provenances of Norway spruce is consistent with previous studies under controlled conditions, although it could be noted that most such studies did generally not report duration of bud development (bud set and bud burst stages), but percentage of plants with buds only [
8,
10,
14,
25].
Similar to previous studies of Norway spruce and white spruce seedlings under controlled conditions [
21,
25,
26], growth cessation and bud development under SD were generally more rapid at higher compared to lower temperature (
Figure 2a–d). The slow bud development in plants exposed to SD at 12 °C is likely to be due to a slow metabolic rate at such low temperature. This is supported by temperature having a similar effect on the first appearance of terminal buds as well as the further development towards mature buds with brown, well-developed bud scales. Furthermore, the Rana plants tended to have a more rapid subsequent bud burst, while both provenances broke buds faster after being exposed to the lowest temperature during SD (
Figure 2e,f). Taken together, these responses demonstrate that the depth of dormancy not only depends on the specific temperature regime under bud set but also the latitudinal origin, which is similar to the situation reported in [
23,
25].
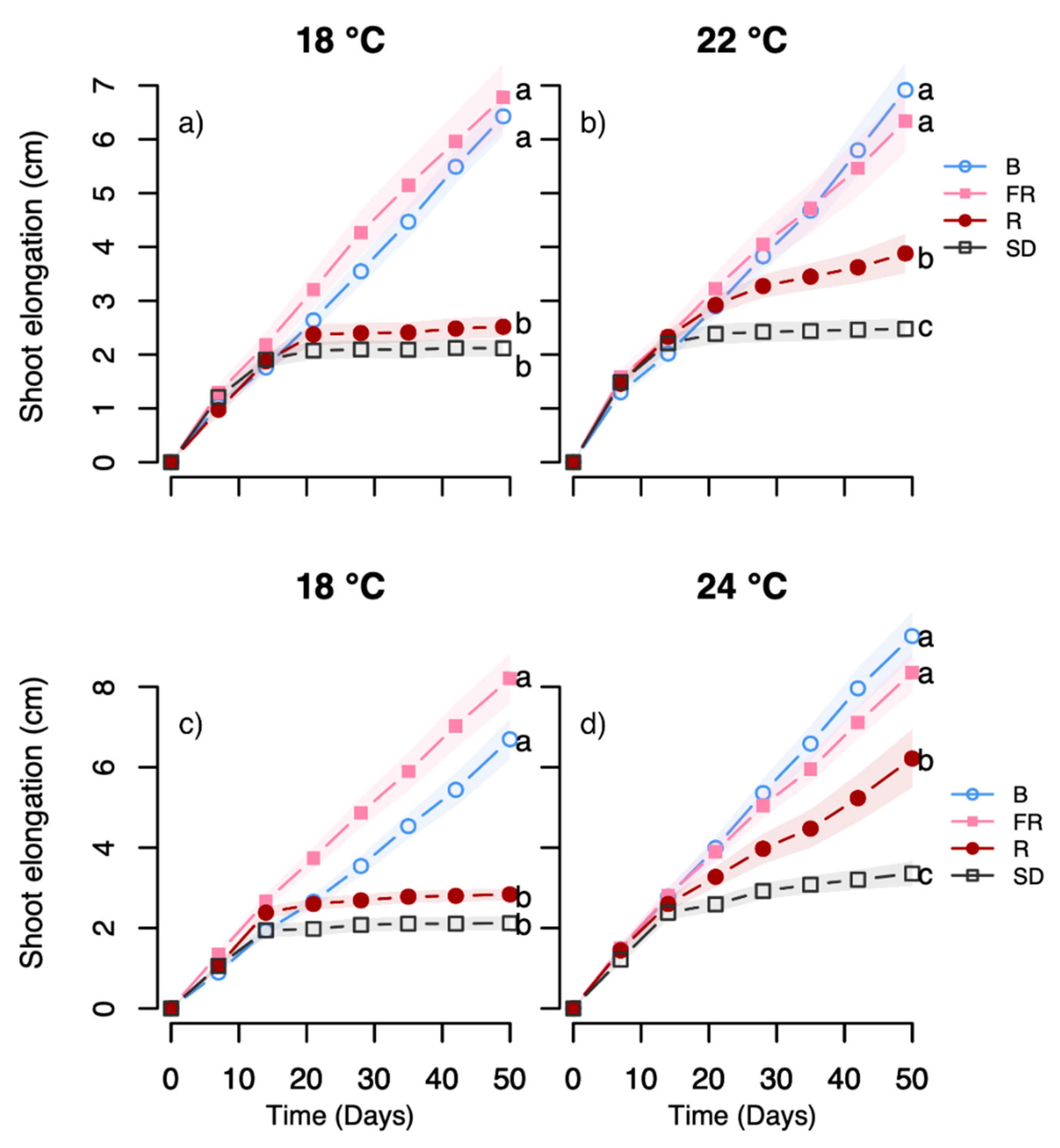
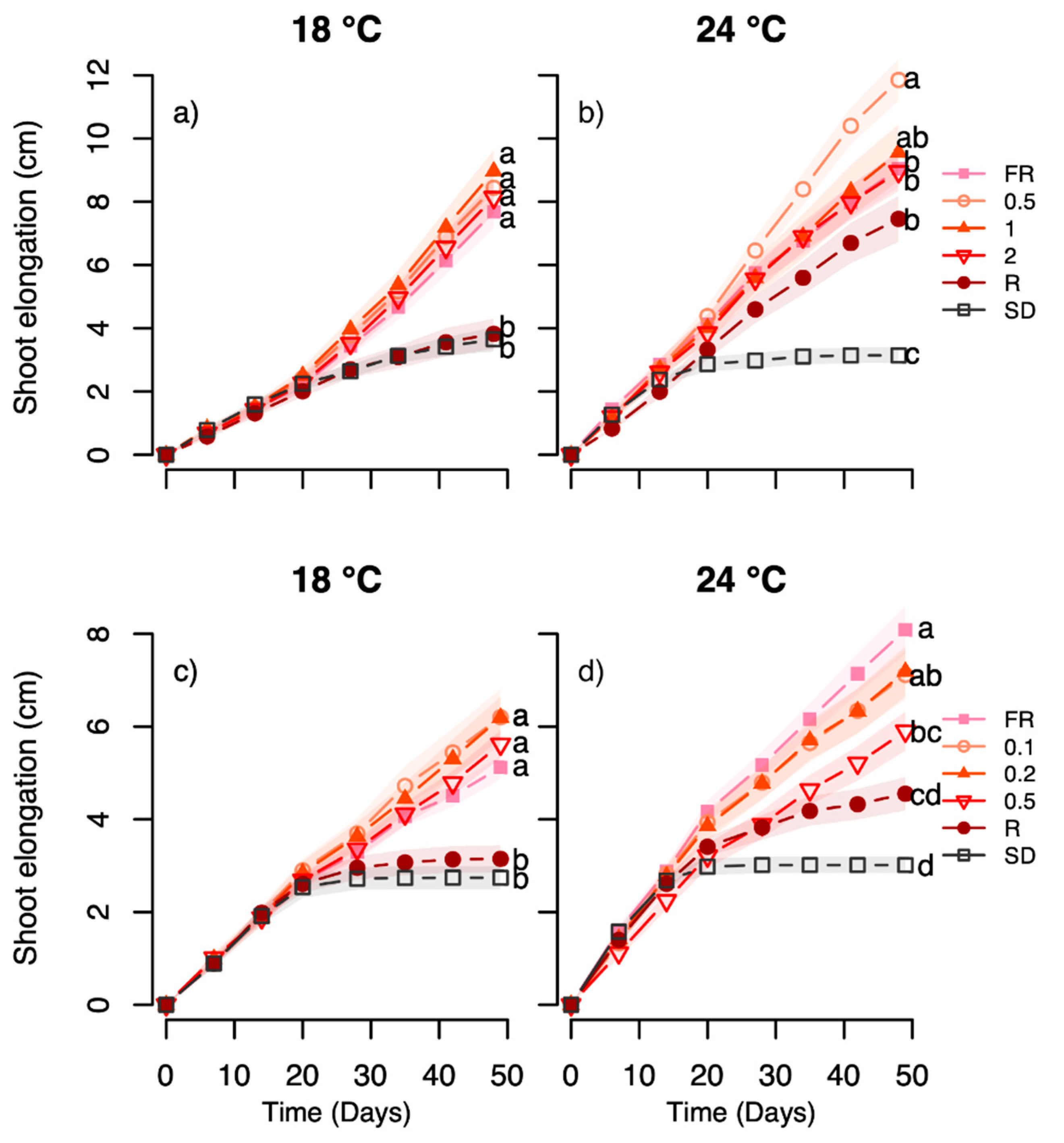
The light-quality experiments employing day extensions with narrow bandwidth LEDs indicate an interplay between temperature and light quality, particularly with respect to effect of R light, as studied in the Halden provenance. The growth and terminal bud development in response to such an R light treatment exhibited strong temperature-dependence, with substantially increased growth and delayed bud set at 24 °C but not 18 °C, as compared to under SD (
Figure 3,
Figure 4,
Figure 5 and
Figure 6). For the Halden provenance, which has a critical photoperiod for growth between 16 and 18 h [
41], SD of 12 h clearly acts as a strong signal inducing rapid bud set in the vast majority of the plants at 18 °C and 22/24 °C (
Figure 3,
Figure 4,
Figure 5 and
Figure 6). In this provenance, bud set was also previously shown to be delayed by R light day extensions at 0.6–18 µmol m
−2 s
−1 (0.1–3.3 W m
−2)) at 18 °C as compared to SD [
10,
11], while our data here for R light at 38 µmol m
−2 s
−1 (7 W m
−2) show that the response is modified at 24 °C compared with 18 °C. Namely, the combination of 24 °C and R light resulted in substantially delayed bud development or even prevention of bud set in large portions of the plants, i.e., 59%, 73% and 56% in experiments 4, 5 and 6, respectively. Thus, at an elevated temperature such as 24 °C, the day extension with R light apparently acts as a far weaker bud-inducing signal than SD alone, or a somewhat weaker growth-sustaining signal than a LD treatment of a wider wavelength area. The interactive effect of temperature and day extension with R light on bud set in experiment 3–6 can also be hypothesised to be due to possible signalling associated with the peak of the growing season, considering the higher proportion of R light during late summer (July–August) in field conditions [
42]. However, it should also be noted that responses under controlled conditions employing day-extension treatments may differ from in the field [
43].
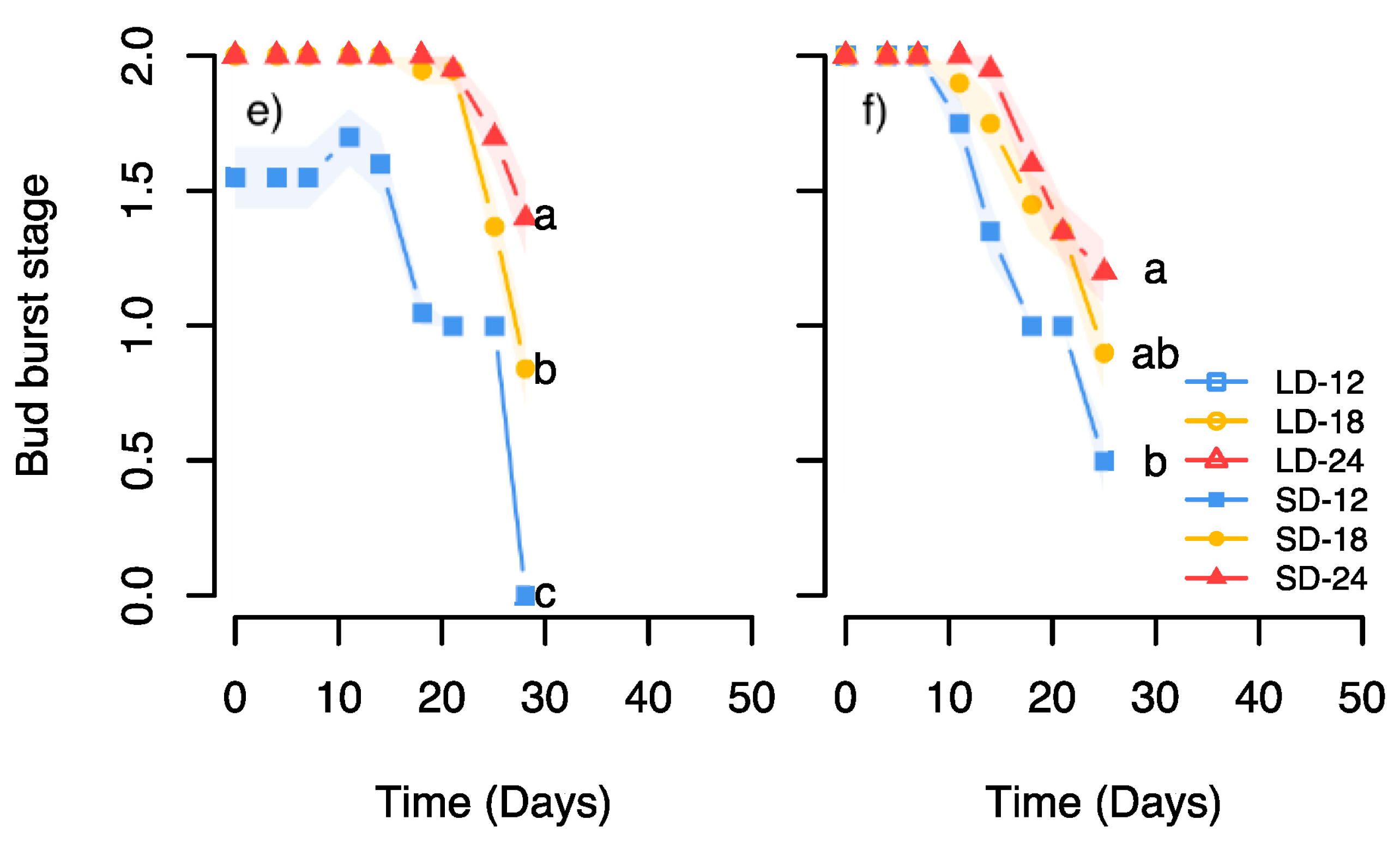
Furthermore, the subsequent advancing effect of bud burst of the R light treatment at 24 °C during bud set compared to SD, but not at 18 °C, and the slightly advanced bud burst after the R light at 24 °C relative to 18 °C, suggest less deep dormancy in buds developed under this light quality combined with elevated temperature (
Figure 4e,f). This is opposite to the situation under SD in this and previous studies of Norway spruce seedlings where the most rapid bud burst was observed after lower compared to higher temperature under bud development (
Figure 2e,f and
Figure 4e,f) [
23,
25]. Differences in depth of dormancy, in addition to cold hardiness, may have important practical implications for seedling production in nurseries since a shallow dormancy may increase the risk of freezing damage due to early spring bud break or fluctuating temperatures during winter [
44].
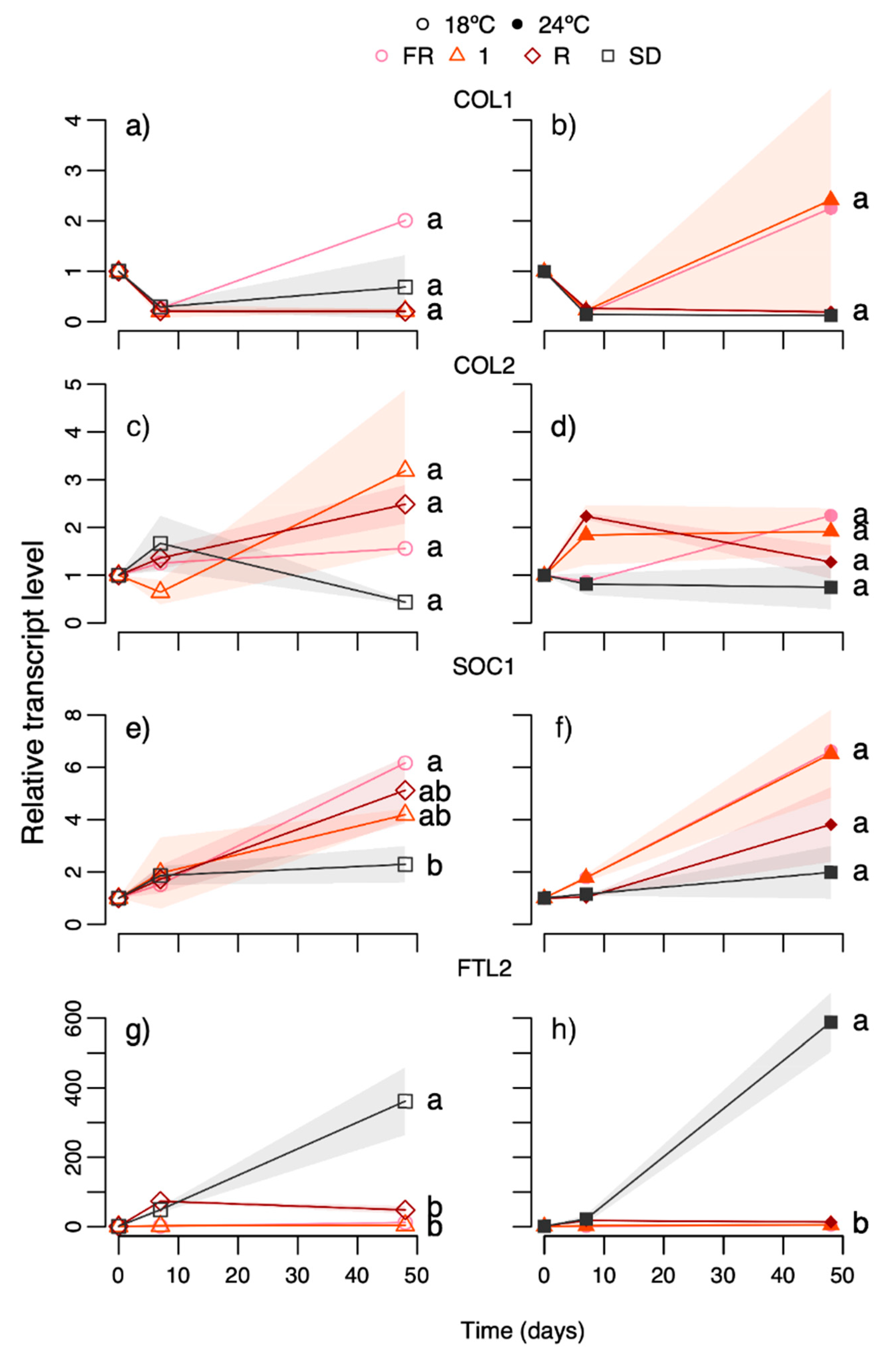
As expected, based on previous studies [
8,
10,
11,
14], the FR light day extension sustained shoot elongation and prevented terminal, vegetative bud development. Here we show that this occurred independently of temperature (
Figure 3,
Figure 4,
Figure 5 and
Figure 6). Previously, day extension with a combination of R and FR light (R:FR ratio 1) was found to be more efficient in preventing bud set than FR light only [
10]. However, in the current study, the FR light and all the tested R:FR ratios (0.1–2) sustained shoot elongation, often with increased growth at 24 °C compared to 18 °C, and no winter bud formation occurred (
Figure 3,
Figure 4,
Figure 5 and
Figure 6). The reason for the difference between this and the previous study remains elusive, but using different lamp types providing different spectral distribution and irradiances during the main light phase in [
10] compared to this study may have influenced.
The sustained growth and prevention of bud set at all temperatures under day extension with B light at 27 µmol m
−2 s
−1 (7 W m
−2) (
Figure 3 and
Figure 4), in contrast to the previously tested 6 and 13 µmol m
−2 s
−1 (1.5 and 3.3 W m
−2) [
10,
11], indicate an irradiance-dependence for such a B light treatment to sustain growth. It may be hypothesised that the B light day extension provided here may increase the photosynthetic capacity and the leaf formation rate [
45], and thus trigger continued growth instead of terminal bud development. In Norway spruce seedlings, increased proportion of B light during the main light period (30% versus 5%) was recently shown to improve growth and result in thicker stems, more branches and higher dry matter content in needles and roots [
46]. Such responses are consistent with that an increased proportion of B light under a relatively low irradiance may induce anatomical and morphological characteristics normally associated with higher irradiances, as shown in
Rosa x hybrida [
45]. Possibly the day extension with B light employed in the current study may have a similar effect. However, the responses to B light may differ between woody species. In Subalpine fir seedlings, increased B light proportion during the day and a day extension at 27 µmol m
−2 s
−1 increased/promoted terminal bud development [
15,
46]. It could also be noted that B light was recently shown to promote bud burst in field-collected dormant branches of several deciduous tree species, implying a role of B light as an important environmental cue in phenological transitions [
47].
In general, photoperiod is known to be sensed by the R:FR light-absorbing phytochromes in trees and possibly the B light-sensing cryptochromes, and these interact with the production of growth-regulating and growth-inhibiting plant hormones [
4,
11,
25,
48] (and references therein). Information about the mechanistic interaction between such light receptors and a time-keeping clock mechanism underlying growth rhythms, as well as its output pathways, is limited for conifers. Nevertheless, the rhythmically expressed
FTL2 gene is known to be involved in control of bud set [
32]. Consistent with
FTL2 being upregulated under bud-inducing conditions and its predicted role as a transcription factor regulating growth inhibition or/and promotion of bud set in conifers [
11,
30,
31,
32], the transcript level of
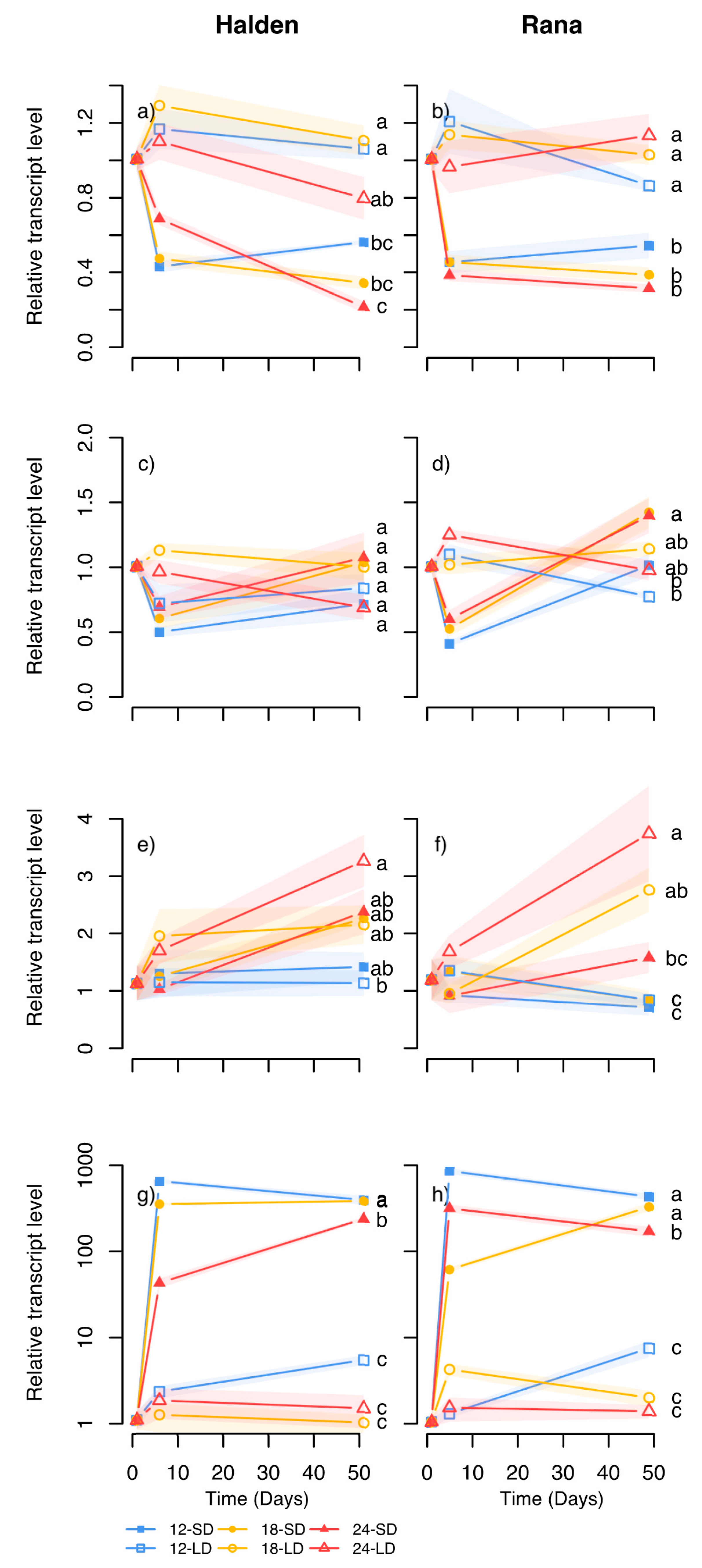
PaFTL2 was higher in the R light-treated plants at 18 °C (at least day 7) where bud set was far more advanced than at 24 °C but less than under SD. Overall, the
PaFTL2 transcript levels correlated well with bud development (
Figure 8,
Figure 9 and
Figure 10;
R2 0.84–0.98), and were far higher in SD-exposed plants than under R light (day 48, but not day 7, experiment 5) and growth-sustaining treatments (FR light, R:FR ratio 1 and LD;
Figure 7g,h and
Figure 9g,h). The transcript levels in shoot tips and apical needles of the B light exposed plants at 18 °C was similar with those under FR light, consistent with the sustained growth (data not shown; B light at 24 °C not analysed). Furthermore, 7 days after retransfer of the plants from SD at 12 °C, 18 °C or 24 °C to LD at 18 °C (experiment 1–2), the
FTL2-transcript levels had decreased to levels close to those of LD in all cases in both provenances (data not shown). These responses are consistent with a role of FTL2 in phenological transitions [
32].
Of the other genes studied,
PaCOL1 showed high correlation between transcript levels and bud set in response to SD (
Figure 8a). Also, consistent with previous studies [
11,
34], the levels of this as well as
PaCOL2 and
PaSOC1 tended to be lower under SD than the growth-sustaining treatments, but there was no clear-cut, directional effect of temperature. Although
CO in
Populus seedlings is involved in controlling growth rhythms [
49] and both
CO and
COL-genes such as
COL1 are determinants of sensitivity to photoperiod and
SOC1 is a floral integrator in herbaceous plants [
36,
49], the function of
COL1-2 and
SOC1 in conifers such as Norway spruce remains to be elucidated by functional studies employing altered gene expression.


 and
and 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}