Herbivore Gender Effects on Volatile Induction in Aspen and on Olfactory Responses in Leaf Beetles


1
Terrestrial Ecology Section, Department of Biology, University of Copenhagen, Universitetsparken 15, DK-2100 Copenhagen E, Denmark
2
Department of Environmental and Biological Sciences, University of Eastern Finland, Kuopio Campus, PO Box 1627, FI-70211 Kuopio, Finland
*
Author to whom correspondence should be addressed.
Forests 2020, 11(6), 638; https://doi.org/10.3390/f11060638
Submission received: 30 April 2020
/
Revised: 30 May 2020
/
Accepted: 3 June 2020
/
Published: 4 June 2020
(This article belongs to the Section Forest Ecology and Management)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Hybrid aspen (Populus tremula × tremuloides Michx.) is a fast-growing tree species used for short-rotation forestry in northern latitudes. Aspen species have a rich herbivore fauna, including defoliating leaf beetles that induce emissions of volatile organic compounds (VOCs) when feeding on aspen leaves. We investigated the differential induction of VOCs by male and female Phratora laticollis leaf beetles feeding on hybrid aspen and the differences in the orientation of beetles in response to gender-specific induced VOCs. The hypotheses for the study were (1) the VOCs in the headspace of plants infested with beetles of the two genders individually and in mixed aggregates would vary subtly, and (2) foraging adult beetles would be able to detect differences in VOC blends and use them to fine-tune their orientation choices. In Y-tube bioassays, both females and males preferred VOCs from leaves damaged by one gender (females or males) over undamaged leaves. However, if leaves were damaged by a two-gender population, neither females nor males indicated a preference over volatiles of undamaged leaves. Leaves damaged by both beetle genders simultaneously had significantly increased green leaf volatile (GLV), benzenoid and homoterpene emissions compared to undamaged leaves. Emissions of these compounds possibly indicate higher herbivore pressure and a higher risk of attack by parasitoids and predators and could thus be the cause of the lack of beetle preference. Our findings provide new basic information on gender-based host plant selection by herbivores and may be helpful in the development of sustainable biogenic VOC-based herbivore-control methods for intensive short-rotation hybrid aspen production.
1. Introduction
When plants are consumed by herbivores, they release a bouquet of volatile organic compounds (VOCs) that reflects the extent and temporal dynamics of the physical damage inflicted and the actions of chemical elicitors released by the herbivores into the wound site [1,2]. Taken together, the physical wounding and chemical elicitation lead to a defense response and induced VOC blend that can be distinct to the plant species and the herbivore species inducing the damage [1,2,3]. It has been well documented that plants can release VOCs from sites localized to the damage and also from undamaged parts of the plant in what is known as a systemic response [4,5,6]. Furthermore, various plant species, including woody and herbaceous plants, have been shown to coordinate systemic defenses in response to herbivore-induced VOCs released by neighbouring plants [7,8,9,10]. Several Populus species, and some cultivated hybrids, have been studied as models for plant defense induction and emission of herbivore-induced plant volatiles (HIPVs) [11,12,13,14]. Some Populus species have shown a lack [14,15] and some species a presence [16] of systemic defense between damaged and intact ramets or leaves suggesting that localized wounds may constitute the dominant defense response in mature trees. Moreover, hybrid Populus saplings have been observed to both enhance secretion of extra-floral nectar (EFN) and prime volatile emissions in response to HIPVs emitted by conspecific neighbours [7] and also to coordinate systemic defenses through within-plant signalling by HIPVs [8,17,18].
Populus nigra saplings have been shown to emit distinct VOC blends in response to damage by several different herbivore species of the Lepidoptera and Coleoptera families [13,16]. The most distinct differences were observed between the different orders, with differences between species within each order less clearly marked [6]. For the Lepidoptera species Lymantria dispar (gypsy moth) and Laothoe populi (poplar hawkmoth), the developmental stage of the herbivore had a more profound influence on HIPVs than the herbivore species, with early instar larvae inducing nitrogenous volatiles and terpenoids more strongly than later instar larvae [14]. Thus far, the effects of gender on herbivore-induced VOC emissions have only been studied with plant-sucking insects on agricultural plants [19,20].
In numerous studies, VOCs emitted in response to herbivore-feeding have been shown to provide cues to foraging insects [21,22]. These cues may repel herbivorous insects in a process known as direct defense [23], or attract natural enemies of the feeding herbivores in a process known as indirect defense [24]. The emission of herbivore-induced VOCs by plants is thought to play a role in negotiating the reliability–detectability problem of finding inconspicuous prey by predatory and parasitic insects [25]. However, in recent years, plant-emitted VOCs have also been shown to play key roles in attracting herbivores [23], whereas insect-emitted pheromones may act in combination with HIPVs in the process of mate location by natural enemies [26]. It is becoming clearer that subtle differences in induction of volatiles by insects of different species and different developmental stages can provide distinct informative cues to other organisms. It is also clear that combinations of plant and insect derived cues can be involved in synergy to elicit mate-finding behaviours. For example, males of the plant bug Nezara viridula preferred to orientate towards host plants damaged by conspecific females than male-damaged plants [20]. Therefore, it is important to develop a greater understanding of how herbivores respond to HIPVs, and whether there is a difference in the behaviours of the different sexes to the plant-herbivore complexes.
Hybrid aspen P. tremula × tremuloides is a hybrid of European and American aspen and a fast-growing tree species that has the capacity to sequester large amounts of carbon in northern latitudes [27]. It has been widely planted in Nordic and Baltic countries for short rotation bioenergy biomass, pulpwood and timber production [27,28,29]. Hybrid aspen stands may be affected by a range of pathogenic infections and herbivore infestations. Serious outbreaks of fungal stem and branch cankers and bacterial cankers have been observed, while a range of stem boring and leaf chewing herbivores can cause considerable damage [28,30]. Leaf weevils [30] and leaf beetles [31] are among the most serious defoliators of P. tremula × tremuloides.
Many species of the leaf beetle family Chrysomelidae feed on leaves of deciduous trees and species in the genus Phratora are specialised at feeding on the tree family Salicaceae, with both adults and larvae feeding on poplar and willow leaves [31]. Phratora laticollis Suffrian (Coleoptera: Chrysomelidae) is a common European leaf beetle that feeds mainly on poplars in the Nordic countries. Adults overwinter in leaf litter or under the bark of dead wood; in June adults feed on leaves, mate and lay eggs on leaves that are also consumed by developing larvae and then by the second-generation adults [32]. In this study, P. laticollis adults were utilised to investigate the differential induction of VOCs by male and female beetles and the differences in orientation of beetles in response to gender specific HIPVs. The following two questions were addressed: (1) Do P. tremula × tremuloides saplings differ in the volatile profiles they emit in response to herbivore-feeding by female and male leaf beetles in gender specific and mixed aggregates? (2) Do male and female P. laticollis beetles differ in their orientation to VOCs induced by female and male leaf beetles in gender-specific and mixed aggregates? The hypotheses for the study were (1) that the VOCs in the headspace of plants infested with beetles of the two genders individually and mixed would vary subtly, and (2) that foraging adult beetles would be able to detect differences in VOC blends and use them to fine-tune their orientation choices.
2. Materials and Methods
2.1. Plants and Insects
The plants used in this study were hybrid aspen (Populus tremula L. × Populus tremuloides Michx., known also as Populus × wettsteinii L.Hämet-Ahti) clone 55, which originated from Anjalankoski (60°45′N, 26°59′E) and was originally produced by crossing P. tremula from Southern Finland with P. tremuloides from Canada. Seedlings were micropropagated in the laboratory [7] and grown individually in pots containing a mixture of peat and sand (3:1) in the greenhouse. At 1–2 days before experimental manipulations, seedlings (8–10 weeks old and 90–120 cm tall) were transferred to the laboratory and grown under room temperature and natural light through the laboratory window (approximately 18–20 h/day). Supplementary lighting (approximately 300 μmol m−2 s−1) was provided with fluorescent lamps in a 12h L/12h D cycle. We selected plants without visible signs of disease and herbivory for all experiments.
Phratora laticollis larvae and egg masses were collected in 2015 from aspen saplings at the University of Eastern Finland Ruohoniemi field site (62°13′N, 27°13′E, 80 m a.s.l.). P. laticollis populations at the Ruohoniemi field site were originally derived from populations collected in 2011–2014 from aspen plantations at the experimental site of the Finnish Forest Research Institute in Suonenjoki, Finland (62°39′N, 27°03′E, 120 m a.s.l.). Larvae and egg masses were reared on hybrid aspen seedlings in the laboratory under natural light and room temperature, and their offspring were used in the experiments.
2.2. Odor Sources for Dual-Choice Olfactory Bioassays
To assess whether P. laticollis adults can use plant volatiles as host location cues, we carried out Y-tube olfactometer bioassays to assess their olfactory responses to VOCs from plants subjected to different treatments. The olfactory responses to the following odor pairs were investigated: (a) Undamaged leaves versus charcoal filtered air, (b) undamaged leaves versus leaves damaged by male adults, (c) undamaged leaves versus leaves damaged by female adults, and (d) undamaged leaves versus leaves damaged by a mixture of female and male adults. Each damaged plant was prepared by enclosing the top 11–13 leaves in a mesh bag together with 24 adults (i.e., 12 males and 12 females for those plants damaged by mixed genders). Control plants were bagged in the same way, but without insects inside. Feeding lasted for approximately 48 h on all but two plants (one plant damaged by females and one plant damaged by mixed genders), where feeding lasted for about 72 h. Over this feeding period, feeding caused, on average, approximately 13.5% (range 5–20%) of leaf area loss on the enclosed leaf material based on visual estimation. There was no significant difference in the leaf area consumption among experiments (Table S1). After feeding treatment, plants were used for olfactory bioassays (see below). To avoid any effects of insect-derived odors, mesh bags, insects and feces were carefully removed from leaves before the start of bioassays. In total, five plants were investigated for undamaged leaves versus charcoal filtered air; four, five and six plant pairs were investigated for undamaged versus leaves damaged by females, males and mixed genders, respectively. Each plant pair was assessed for both male and female adults, with between six and 22 insects tested for each gender depending on the availability of the insects. In two plant pairs from undamaged versus male-damaged leaves, bioassays were conducted only for males due to the unavailability of females. The experimental period lasted for approximately three months from 9 June–29 August 2015, and the treatments with different insect genders were conducted sequentially one after another (See Table S2 for the timelines of the bioassays).
2.3. Dual-Choice Olfactory Bioassays
Dual choice bioassays were carried out in glass Y-shaped olfactometers (12-cm base arm length, 8-cm side arm length, 1.6-cm interior diameter) using P. laticollis adults starved for 12–24 h. Each insect was used for only one trial, and insects were not reused between replicates.
VOCs from the trees were directed into the olfactometer as follows. A pre-cleaned (120 °C for 2 h) polyethylene terephthalate (PET) bag (25 cm wide × 55 cm deep; Rainbow, Helsinki, Finland) was placed over the top 11 leaves. A hole was cut in the top corner of the bag, a Teflon tube was inserted, and compressed air, purified by an activated charcoal filter and MnO2 ozone scrubber was pumped into the bag until it inflated. A hole was then cut in the remaining top corner, and a second Teflon tube was inserted and connected to one arm of the olfactometer. The second plant was connected to the other arm of the olfactometer in the same way (Figure 1). Air was channelled through both bags and into the olfactometer at a flow rate of approximately 500 mL min−1. Teflon tubing was used to connect the different parts of the system. Plants were illuminated by artificial growth lights throughout the olfactometry assays.
The olfactometer was placed inside a green plastic box, which was illuminated from above and covered with aluminum foil during the bioassays to exclude visual cues and reduce visual bias. At the start of each trial, a beetle was introduced into the olfactometer by placing a plastic tube in it with a mesh-covered end and inserting the tube into the base of the olfactometer. The time was started when the beetle left the tube and entered the base arm. Beetles were recorded as having made a choice after walking at least two-thirds of the way down one side arm and remaining there for at least 30 s. Beetles that did not make a choice within 5 min were recorded as “no choice”. In addition, the time spent by insects in the odor fields were recorded during the 5-min period. To account for any potential effect of associative learning and the traces left by the adults on the choices made, each insect was tested only once and a clean olfactometer was used for each test. To compensate for any unforeseen asymmetry inherent to the experimental setup, we alternated odor sources after every five to 10 trials. After each trial, olfactometers were thoroughly cleaned with 70% ethanol and oven-dried at 120 °C for a minimum 20 min before reuse.
2.4. VOC Collection and Analysis
We performed volatile collections for all odor comparisons mentioned above, except for the odor pairs of plants versus filtered air. VOCs were collected immediately after olfactory bioassays, i.e., 4–6 h after feeding termination, using a push–pull headspace sampling system. The incoming air was maintained as it was during the olfactory bioassay, with a flow rate of about 500 mL min−1. The headspace was pulled out with a vacuum pump at 200 mL min−1 through a stainless steel adsorbent cartridge filled with Tenax TA and Carbopack B (150 mg of each, mesh 60/80; Markes International, Llantrisant, RCT, UK). The adsorbent cartridges were loosely tied to the PET bags to allow excess air to exit. Volatiles were collected for 30 min.
Volatile samples were analyzed by gas chromatography–mass spectrometry (GC–MS; Agilent GC 7890A, MSD 5975C, New York, USA). Trapped compounds were thermally desorbed (TD100, Markes International, Llantrisant, RCT, UK) at 250 °C for 10 min, cryofocused at 10 °C and injected onto an HP-5 capillary column (50 m × 0.2 mm; film thickness 0.5 μm) with helium as a carrier gas at a flow of 1.2 mL min−1. The column temperature was held at 40 °C for 1 min, then ramped at 5 °C min−1 to 210 °C, and ramped again at 20 °C min−1 to 250 °C. Chromatograms were analyzed using the software PARADISe (version 3.9) [33]. Individual VOCs were tentatively identified by comparing mass spectra to those in the NIST 2014 mass spectral library, verified by chromatography with authentic standards where available and quantified based on external standards. For those compounds that had no authentic standards available, quantification was estimated based on α-pinene. Emission rates are expressed in ng gram dry weight per hour (ng gDW−1 h−1).
2.5. Statistical Analysis
All statistical analyses were performed in R version 3.6.3 (R Core Developmental Team 2018). All dual-choice bioassays were analyzed with binomial tests to determine whether insect preferences differed significantly from a 50:50 distribution (p = q = 0.5, two-tailed, α = 0.05). During the bioassays, the difference in the time spent between the two odor fields were assessed with a paired-sample T-test when the data were normally distributed or a paired-sample Wilcoxon test when this was not the case. Normal distribution of the data was assessed with the Shapiro–Wilk test. Likewise, differences in volatile emissions between control and damaged plants were analyzed separately for single and mixed genders using either a paired-sample T-test or Wilcoxon test depending on whether the normality assumption was met or violated. In addition, gender-specific differences in volatile emissions were tested using Kruskal–Wallis tests, followed by pairwise Wilcoxon–Mann–Whitney tests using Benjamini and Hochberg post-hoc correction.
To characterize differences in volatile blends among treatments, we used a Random Forest (RF) between-group classification algorithm as described previously [34]. RF analysis is similar to a principal component analysis, but is more suitable for nonparametric datasets that comprise many more variables than samples [34]. We performed RF analysis on the emission rates of individual compounds of the entire blend. For each analysis, 100,000 bootstrap samples were drawn with 10 variables (i.e., volatiles) randomly selected at each node. The number of variables selected is based on the square root of all variables. Differences among treatments were then visualized using multidimensional scaling (MDS) plots. The importance of each compound for the distinction is expressed as the mean decrease in accuracy (MDA) and the odds of the compound being improperly classified is expressed as the out-of-bag (OOB) error rate. We conducted the RF analysis using the package randomForest in R.
3. Results
3.1. Role of Plant Volatiles in Host Orientation Behaviour of P. laticollis Adults
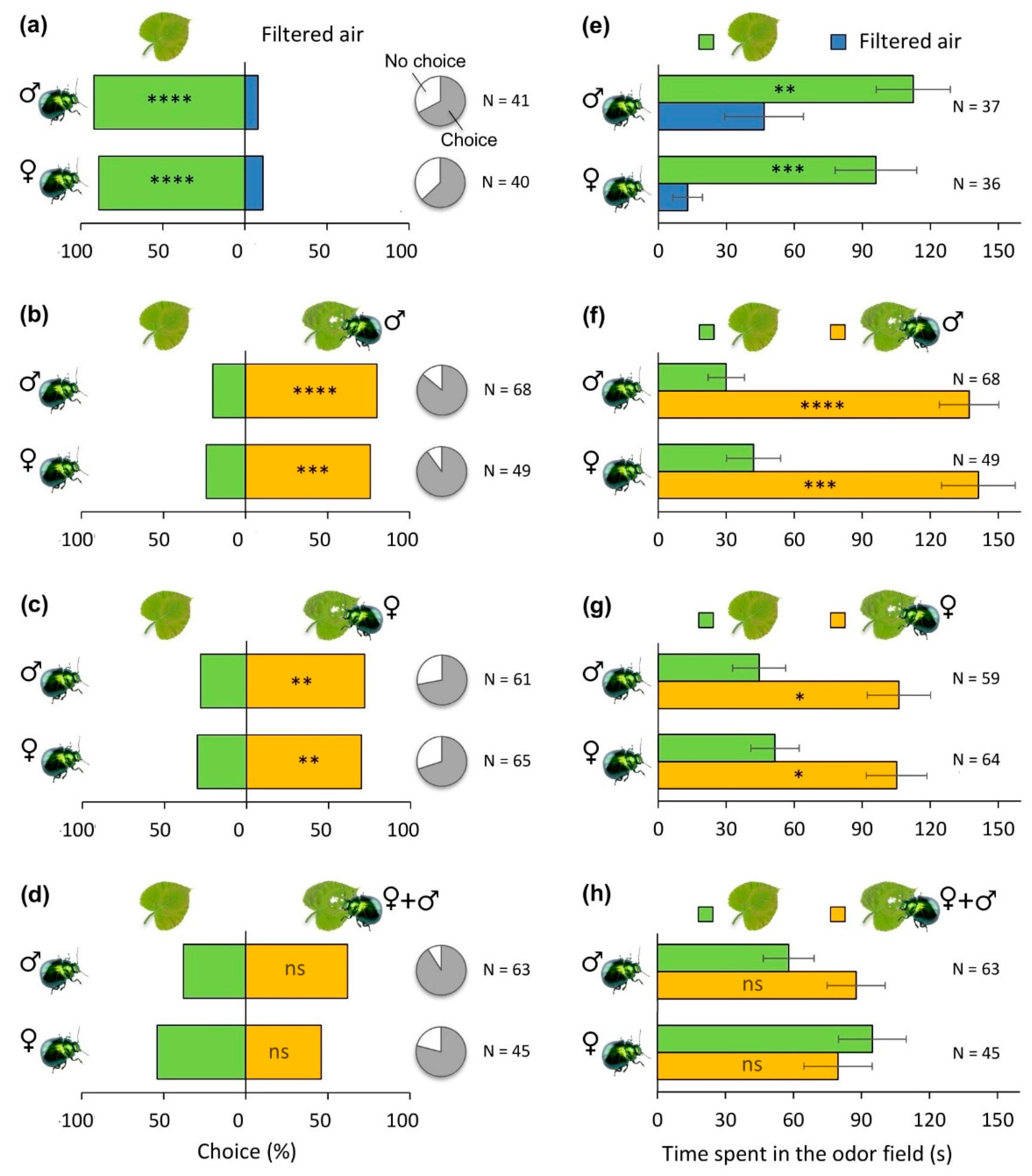
In the Y-tube bioassays, both female and male P. laticollis adults strongly preferred volatiles from undamaged plants to charcoal filtered air (Figure 2a; p < 0.0001) and spent more time in the odor field corresponding to the undamaged plants (Figure 2e; p < 0.001). When offered a choice between volatiles emanating from undamaged and P. laticollis male-infested plants, both male and female adults showed a strong preference for the male-induced odor (Figure 2b; p < 0.001) and spent more time in this odor field (Figure 2f; p < 0.0001). When offered a choice between volatiles emanating from undamaged and P. laticollis female damaged plants, both male and female adults still preferred volatiles released from damaged plants (Figure 2c; p < 0.01) and spent more time in the induced odor field (Figure 2g; p < 0.01). Interestingly, when volatiles released from plants damaged by a mixture of P. laticollis females and males were tested against volatiles released from undamaged plants, both male and female adults did not discriminate between the two odors, though there was a tendency for male attraction toward volatiles from damaged plants (Figure 2d; p = 0.118). In both cases, the amount of time spent in control versus induced odor fields was very similar (Figure 2h; p > 0.15).
3.2. Effects of P. laticollis Herbivory on Host Volatile Emission
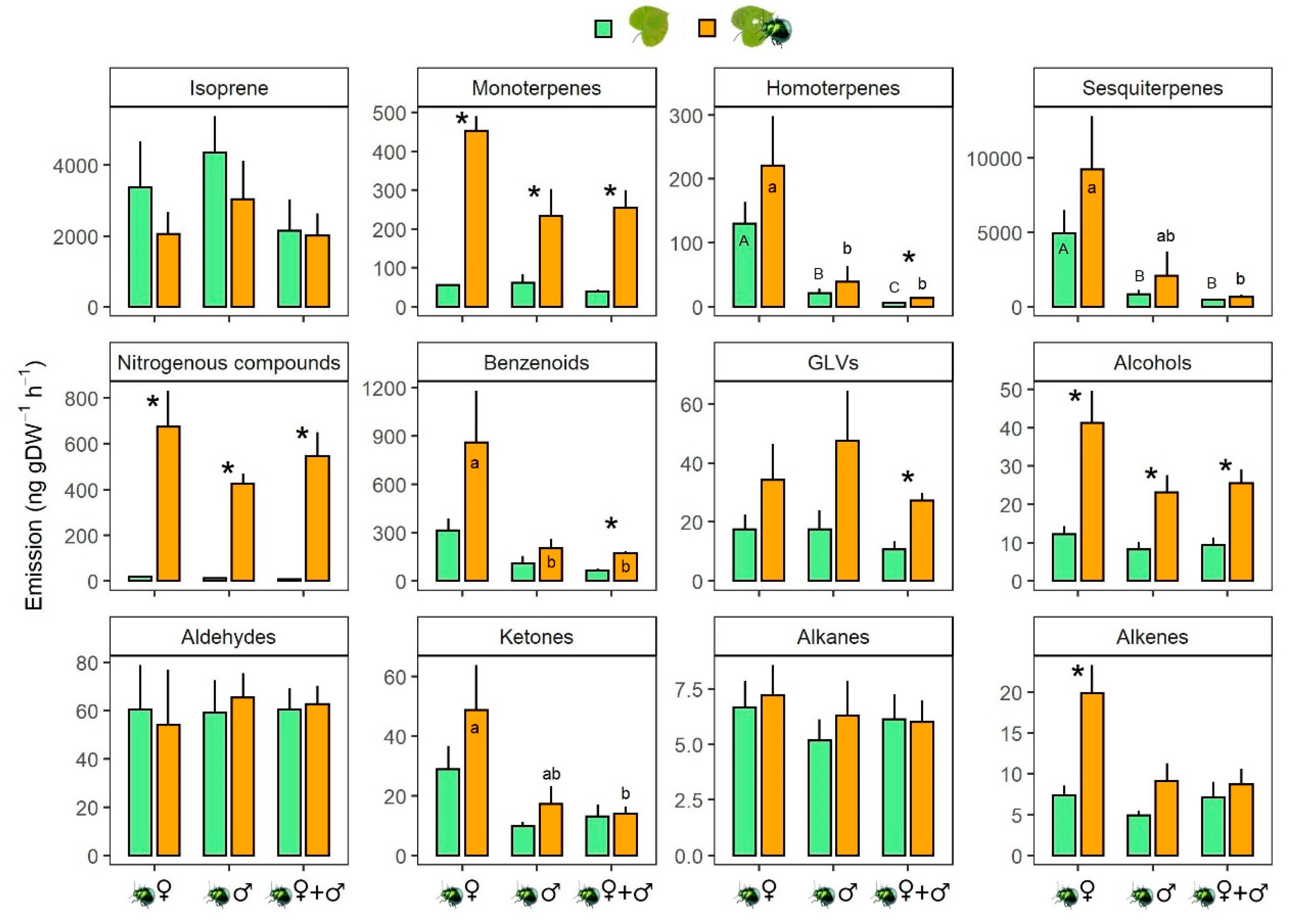
We detected 108 VOCs using GC–MS analysis of 30 headspace collections from 15 undamaged and 15 damaged hybrid aspen seedlings (Table S3). Terpenoids, including isoprene, 27 monoterpenes, three homoterpenes and 17 sesquiterpenes, dominated the composition of the VOC blends, followed by 17 nitrogen-containing volatiles and 17 benzenoids. The remaining volatile groups include five alcohols, five alkanes, five ketones, four aldehydes, four green leaf volatile (GLVs) and three alkenes. Feeding by P. laticollis adults for approximately 48 h prior to 4–6 h olfactory bioassays significantly increased the emission rates of monoterpenes, nitrogenous compounds and alcohols, independently of the sexual identity of the attacking herbivores (Figure 3). Specifically, when averaged across the sexual identity, emissions of monoterpenes, nitrogenous compounds and alcohols from damaged plants were two, five and 44 times higher, respectively, than emissions from undamaged plants. Feeding also enhanced emissions of benzenoids, homoterpenes, GLVs and alkenes, but inconsistently, depending on the sexual identity of the attacking herbivores (Figure 3).
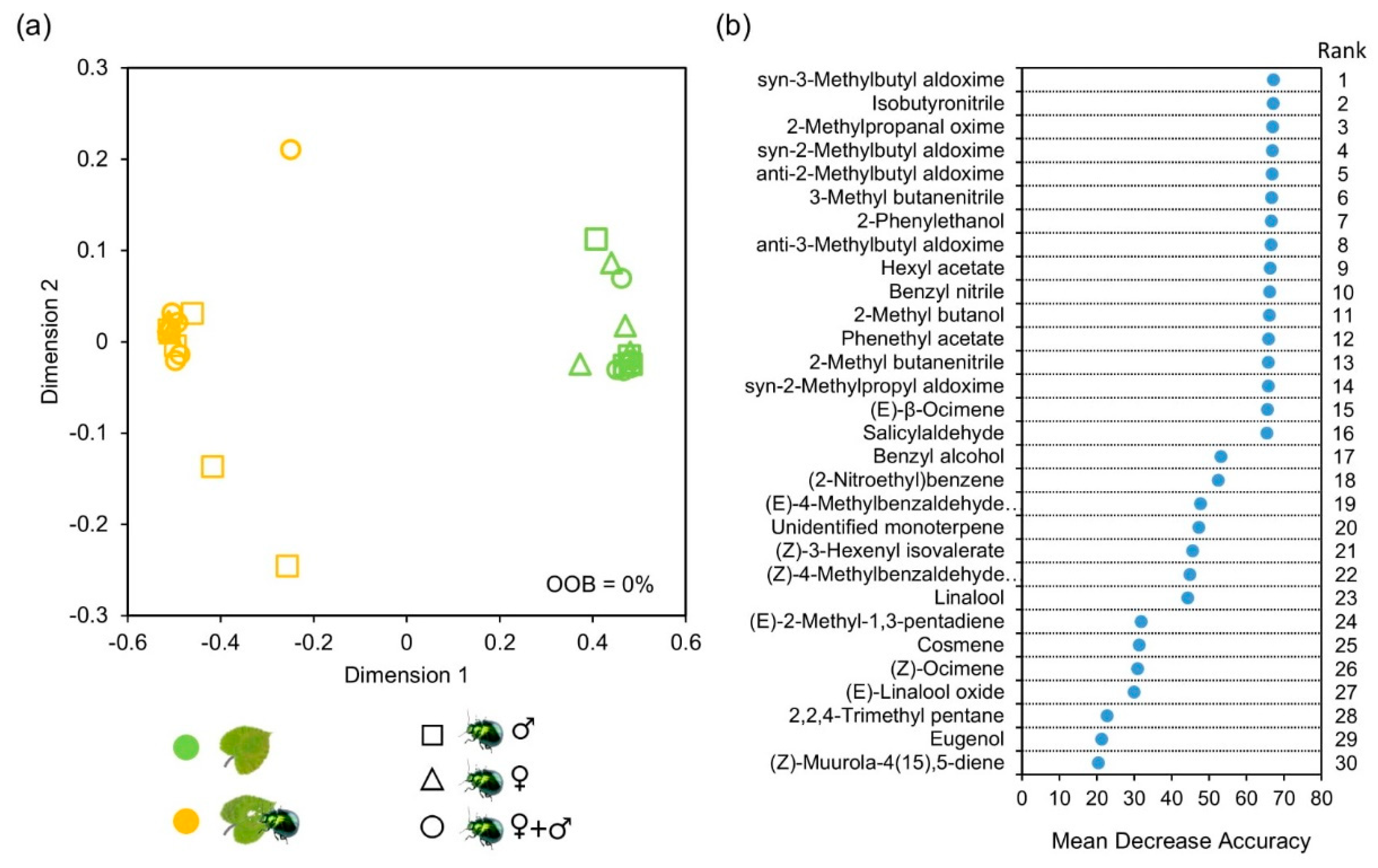
Apart from accelerating VOC emission rates, herbivore feeding significantly altered VOC composition. Multidimensional scaling (MDS) following Random Forest (RF) analysis of the proportional data revealed that overall volatile emission profiles were clearly distinguished between undamaged and damaged plants (Figure 4a). The RF classification model comparing undamaged and damaged plants had an out-of-bag error of 0% for classification by herbivory during 100,000 replicated tree constructions and revealed 16 compounds that were ranked as most important in discriminating between the two groups (Figure 4b). Ten out of these 16 VOCs were nitrogen-containing compounds, such as syn-3-methylbutyl aldoxime, 2-methyl butanenitrile, isobutyronitrile and benzyl nitrile (Figure 4b). The remaining six compounds included three benzenoids (phenethyl acetate, 2-phenylethanol and salicylaldehyde), one monoterpene ((E)-β-ocimene), one GLV ((Z)-3-hexenyl isovalerate) and one alcohol (2-methyl butanol). Each of these 16 compounds was emitted in greater amounts from damaged plants and absent or just present in trace amounts from undamaged plants (Table S3). Volatile blends released by damaged plants could not be clearly separated among sexual identities of the attacking herbivores, as revealed by the high out-of-error rate at 53.3%, even though plants damaged by females tended to stand alone against plants damaged by males or a mixture of males and females based on the MDS plot (Figure S1).
4. Discussion
The results of this study indicate that both females and males of Phratora laticollis leaf beetles prefer the volatiles emitted by undamaged aspen plants when compared to filtered air. When comparing emission from aspen plants damaged by conspecific leaf beetle adults of either gender, both females and males preferred damaged-plant volatiles over the volatiles emitted by undamaged plants. However, if the damage was induced by aggregations of both genders of the conspecific beetles, there was no attraction of either gender in preference to volatiles from undamaged plants. Our results clearly showed that foliage areas already accommodating both sexes feeding are not evaluated by beetles as better feeding and mating sites than undamaged foliage areas.
Leaf beetles differ from many other insect herbivore groups because adult beetles do similar leaf damage to host plants as larvae. This avoidance of mixed gender-induced VOC emissions might be an adaptation to avoid the overcrowding of the host plant before the larvae start feeding. Typical behaviour for leaf beetle populations is to move constantly to undamaged foliage and leave the occupied foliage after oviposition to secure food for the larvae [35]. Therefore, single gender-specific VOC emissions induced in host plants could be important signals by which leaf beetles evaluate the suitability of the host for mating, and VOC emissions induced by both genders simultaneously could signal risk of overcrowding. The gender-specific induction of host plant VOCs and the significance for insect behaviour is, thus far, poorly understood [19,20,36]. The omnivorous plant bugs predate on herbivorous insects, but also suck the fluids of the host plant where they lay their eggs [20,36]. Feeding by Nezara viridula Reuter (Heteroptera: Miridae) females induced the emission of the sesquiterpene (E)-β-farnesene on a sesame host plant but not on an eggplant [20]. Feeding the Macrolophus caliginosus Wagner (Hemiptera: Miridae) on beans also induced the emission of (E)-β-farnesene, suggesting that these sesquiterpenes could be signalling compounds that attract males and promote mating [20,36]. However, no evidence of how plant VOCs induced by mixed-gender feeding could affect behaviour of conspecifics has been published. Our study is possibly the first to give evidence of gender-related induced VOC signals of crowding.
We removed the insects and their feces from the damaged plants before the olfactometric bioassays and we tested the response of each insect in a single clean Y-tube; therefore, the involvement of insect pheromones or secretions in orientation behaviours in our tests is unlikely. This means that the orientating beetles should have based their decision on the qualitative changes in the emissions of the herbivore-damaged plants when compared to emissions from undamaged plants. Our analysis of VOC emission rates showed that leaf feeding by adult beetles significantly increased the total emissions of monoterpenes, alcohols and nitrogen compounds in all feeding treatments when compared to undamaged plants. This observation agrees with an earlier report [37] demonstrating that Populus × euroamericana showed an increased proportion of monoterpenes in total VOC emission from leaf beetle-damaged young leaves. Monoterpene emissions were also found to be attractive to newly emerged leaf beetle adults in this Populus species.
The increases of emission rates of monoterpenes, alcohols and nitrogen compounds did not differ between single-gender damaged and mixed-gender damaged plants, hence they may not have any value in explaining the differences between these treatments. However, the herbivore-induced GLV and benzenoid and homoterpene emissions in the mixed-gender damage treatment were significantly increased compared to undamaged plants, while in the single-gender damage treatments the increasing trend in induced GLV and homoterpene emissions was not significant. Phratora species are sensitive to changes in GLV composition in damaged plant emissions [38]. In the mixed-gender treatment, the overall homoterpene and GLV emission rates were also at a lower level in control plants, which might have partly affected the significance of the damage-induced increase. Therefore, the lack of a significant difference in beetle orientation towards undamaged and damaged plants in the mixed-gender treatment may indicate that the emission of homoterpenes and GLVs could have some explanatory value for leaf beetle behaviour, leading to a lower attractiveness of plants having feeding damage by a mixed-gender population. Emissions of GLVs and the homoterpene DMNT from tree foliage are typically related to increased feeding pressure on the plant foliage and the density of feeding insects on defoliated plants [39,40,41]. It can be speculated that, in mixed populations, the feeding activity of females was probably increased because a higher percentage of females had already mated. Greater emissions of GLVs and homoterpenes from beetle-damaged than undamaged foliage could indicate a higher risk of within-species competition, higher leaf consumption rate due to competition [39], increased pressure by parasitoids [42,43] and predators [44,45] attracted by GLV and homoterpene emissions, and eventually overcrowding and food shortage in damaged foliage [46,47]. In terms of willows, it has been shown [38] that Phratora spp. adults have also capacity to learn to use the induced GLV compound composition to differentiate damaged and intact willow species.
An analysis of the individual monoterpenes emitted in response to leaf-beetle damage (Table S3) showed that only the non-oxygenated monoterpene (E)-β-ocimene, known to be a plant stress-indicator [48], was consistently increased in all three feeding treatments. This suggests that the feeding treatments produced a generally recognised stress response in damaged plants. Furthermore, (E)-DMNT was only emitted in significantly higher amounts than control plants in the mixed-gender treatment. Emissions of the sesquiterpene α-farnesene are often indicative of insect damage in foliage [17], but, in this study, the undamaged plants also had relatively high emission rates of this compound and no treatment effects were found. GLV emissions normally respond to insect feeding on tree foliage within seconds of the herbivore mouth parts starting to crush the cell membranes of the leaves [49]. Emission rates of the GLV (Z)-3-hexenyl isovalerate were consistently increased in all feeding treatments, although total GLV emissions were only emitted at significantly increased rates in the mixed-gender treatment.
Random Forest analysis revealed that a group of nitrogen-containing compounds, such as syn- and anti-2-methylbutyl aldoxime, syn- and anti-3-methylbutyl aldoxime, and benzyl nitrile, are the most characteristic volatiles of plants damaged by leaf beetles. These compounds were released in higher amounts from damaged plants, irrespective of the gender identity of the attacking beetles, while their emission from undamaged plants were at trace levels. Similarly, earlier studies on black poplar (Populus nigra) have shown that the same aldoximes and nitriles as reported in our study are induced upon attack by caterpillars of the gypsy moth (Lymantria dispar) and poplar hawkmoth (Laothoe populi), and are the most important compounds distinguishing control and damaged plants [12,13,14]. Together, these studies suggest that induction of aldoximes and nitriles upon herbivore attack might be widespread in Populus species and be used as non-destructive diagnostic markers in Populus pest management. While the ecological roles aldoximes and nitriles play in mediating trophic interactions have been given much less experimental attention than other classes of plant volatiles including GLVs and terpenoids, some studies have demonstrated that they may serve as essential cues attracting herbivore enemies [14,50]. In the present study, nitrogen-containing compounds seem unlikely to mediate leaf beetle aggregation on host plants, as indicated by the observed lack of preference between control plants and plants damaged by mixed genders. However, these compounds may be exploited by natural enemies of leaf beetles, a possibility that deserves future investigation.
5. Conclusions
Our behavioural bioassays revealed that leaf beetle feeding can modify the VOC emissions of aspen foliage and that these emissions are attractive to conspecifics, but that certain VOC compositions may be less attractive and not different to the attractiveness of undamaged plants. These findings provide new insights into the role of the insect gender in the release of herbivore-induced VOCs from hybrid aspens. The gender of the feeding beetles may especially affect the VOC composition and the aggregation rates of the defoliating leaf beetles. Further behavioural tests with a combination of electroantennogram analyses [47] are needed to detect insect antennal responses to specific volatile compounds and their significance in the interaction between hybrid aspen plants and their herbivores. Our results may also give valuable basic information for the development of sustainable pest control methods in intensive short-rotation hybrid aspen production on formerly arable land [27]. The novel hybrid aspen ecosystems established in agricultural environments and separated from natural forests resemble more low-diversity agricultural monocultures and the aspen trees could be much more vulnerable to leaf beetle outbreaks than in more diverse mixed forest ecosystems.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/11/6/638/s1, Figure S1. Multidimensional scaling plot of Random Forest (RF) classification analysis of volatile profiles released from aspen seedlings damaged by different genders. Table S1: Leaf area consumption. Table S2: Time lines of olfactory bioassays. Table S3: Volatile compound emissions of hybrid aspen seedlings upon attack by P. laticollis adults of different genders.
Author Contributions
Conceptualization, T.L. and J.D.B.; methodology, T.L., K.G.-G. and J.D.B.; investigation, K.G.-G.; T.L.; data analysis K.G.-G.; T.L.; resources, J.D.B. and J.K.H.; writing—review and editing, T.L., J.D.B. and J.K.H.; funding acquisition, T.L., J.D.B. and J.K.H.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Academy of Finland decision numbers 309425, 256050, 251898 and 283122, UEF International Top-Level Research Area BORFOR, and Marie Curie project: European Union’s Horizon 2020 research and innovation programme under the Marie Sklodowska-Curie (grant agreement 751684).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aljbory, Z.; Chen, M. Indirect plant defense against insect herbivores: A review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Xiao, L.; Carrillo, J.; Siemann, E.; Ding, J. Herbivore-specific induction of indirect and direct defensive responses in leaves and roots. Aob. Plants 2019, 11, plz003. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Huber, D.; Bohlmann, J. Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa × deltoides): cDNA cloning, functional characterization, and patterns of gene expression of (-)-germacrene D synthase, PtdTPS1. Plant J. 2004, 37, 603–616. [Google Scholar] [CrossRef]
- Staudt, M.; Lhoutellier, L. Volatile organic compound emission from holm oak infested by gypsy moth larvae: Evidence for distinct responses in damaged and undamaged leaves. Tree Physiol. 2007, 27, 1433–1440. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Baldwin, I.T. Herbivory-induced signalling in plants: Perception and action. Plant Cell Environ. 2009, 32, 1161–1174. [Google Scholar] [CrossRef]
- Li, T.; Holopainen, J.K.; Kokko, H.; Tervahauta, A.I.; Blande, J.D. Herbivore-induced aspen volatiles temporally regulate two different indirect defences in neighbouring plants. Funct. Ecol. 2012, 26, 1176–1185. [Google Scholar] [CrossRef]
- Frost, C.J.; Appel, M.; Carlson, J.E.; De Moraes, C.M.; Mescher, M.C.; Schultz, J.C. Within-plant signalling via volatiles overcomes vascular constraints on systemic signalling and primes responses against herbivores. Ecol. Lett. 2007, 10, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Kost, C. Priming of indirect defences. Ecol. Lett. 2006, 9, 813–817. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Schultz, J.C. Rapid changes in tree leaf chemistry induced by damage: Evidence for communication between plants. Science 1983, 221, 277–279. [Google Scholar] [CrossRef]
- Lackner, S.; Lackus, N.D.; Paetz, C.; Koellner, T.G.; Unsicker, S.B. Aboveground phytochemical responses to belowground herbivory in poplar trees and the consequence for leaf herbivore preference. Plant Cell Environ. 2019, 42, 3293–3307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, A.C.; Boeckler, G.A.; Koellner, T.G.; Gershenzon, J.; Unsicker, S.B. The timing of herbivore-induced volatile emission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cues for herbivore enemies. BMC Plant Biol. 2014, 14, 304. [Google Scholar] [CrossRef] [Green Version]
- McCormick, A.C.; Irmisch, S.; Boeckler, G.A.; Gershenzon, J.; Koellner, T.G.; Unsicker, S.B. Herbivore-induced volatile emission from old-growth black poplar trees under field conditions. Sci. Rep. 2019, 9, 7714. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.C.; Irmisch, S.; Reinecke, A.; Boeckler, G.A.; Veit, D.; Reichelt, M.; Hansson, B.S.; Gershenzon, J.; Koellner, T.G.; Unsicker, S.B. Herbivore-induced volatile emission in black poplar: Regulation and role in attracting herbivore enemies. Plant Cell Environ. 2014, 37, 1909–1923. [Google Scholar] [CrossRef]
- Cope, O.L.; Lindroth, R.L. Clonal Saplings of Trembling Aspen Do Not Coordinate Defense Induction. J. Chem. Ecol. 2018, 44, 1045–1050. [Google Scholar] [CrossRef]
- Fabisch, T.; Gershenzon, J.; Unsicker, S.B. Specificity of Herbivore Defense Responses in a Woody Plant, Black Poplar (Populus nigra). J. Chem. Ecol. 2019, 45, 162–177. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Blande, J.D. Volatile-Mediated within-Plant Signaling in Hybrid Aspen: Required for Systemic Responses. J. Chem. Ecol. 2017, 43, 327–338. [Google Scholar] [CrossRef]
- Frost, C.J.; Mescher, M.C.; Dervinis, C.; Davis, J.M.; Carlson, J.E.; De Moraes, C.M. Priming defense genes and metabolites in hybrid poplar by the green leaf volatile cis-3-hexenyl acetate. New Phytol. 2008, 180, 722–733. [Google Scholar] [CrossRef]
- Williams, L.; Rodriguez-Saona, C.; Pare, P.W.; Crafts-Brandner, S.J. The piercing-sucking herbivores Lygus hesperus and Nezara viridula induce volatile emissions in plants. Arch. Insect Biochem. Physiol. 2005, 58, 84–96. [Google Scholar] [CrossRef]
- Rim, H.; Uefune, M.; Ozawa, R.; Takabayashi, J. An omnivorous arthropod, Nesidiocoris tenuis, induces gender-specific plant volatiles to which conspecific males and females respond differently. Arthropod Plant Interact 2018, 12, 495–503. [Google Scholar] [CrossRef]
- Gols, R. Direct and indirect chemical defences against insects in a multitrophic framework. Plant Cell Environ. 2014, 37, 1741–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aartsma, Y.; Bianchi, F.J.; van der Werf, W.; Poelman, E.H.; Dicke, M. Herbivore-induced plant volatiles and tritrophic interactions across spatial scales. New Phytol. 2017, 216, 1054–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blande, J.D. Plant communication with herbivores. Adv. Bot. Res. 2017, 82, 281–304. [Google Scholar] [CrossRef]
- Dicke, M. Behavioural and community ecology of plants that cry for help. Plant Cell Environ. 2009, 32, 654–665. [Google Scholar] [CrossRef]
- Vet, L.; Wackers, F.L.; Dicke, M. How to hunt for hiding hosts-the reliability-detectability problem in foraging parasitoids. Neth. J. Zool. 1991, 41, 202–213. [Google Scholar]
- Xu, H.; Desurmont, G.; Degen, T.; Zhou, G.; Laplanche, D.; Henryk, L.; Turlings, T.C.J. Combined use of herbivore-induced plant volatiles and sex pheromones for mate location in braconid parasitoids. Plant Cell Environ. 2017, 40, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Fahlvik, N.; Rytter, L.; Stener, L. Production of hybrid aspen on agricultural land during one rotation in southern Sweden. J. For. Res. 2019, 1–9. [Google Scholar] [CrossRef]
- Tullus, A.; Rytter, L.; Tullus, T.; Weih, M.; Tullus, H. Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scand. J. For. Res. 2012, 27, 10–29. [Google Scholar] [CrossRef]
- Hytonen, J.; Beuker, E.; Vihera-Aarnio, A. Clonal variation in basic density, moisture content and heating value of wood, bark and branches in hybrid aspen. Silva Fenn. 2018, 52, 9938. [Google Scholar] [CrossRef] [Green Version]
- Freiwald, V.; Haikio, E.; Julkunen-Tiitto, R.; Holopainen, J.K.; Oksanen, E. Elevated ozone modifies the feeding behaviour of the common leaf weevil on hybrid aspen through shifts in developmental, chemical, and structural properties of leaves. Entomol. Exp. Appl. 2008, 128, 66–72. [Google Scholar] [CrossRef]
- Kopf, A.; Rank, N.E.; Roininen, H.; Julkunen-Tiitto, R.; Pasteels, J.M.; Tahvanainen, J. The evolution of host-plant use and sequestration in the leaf beetle genus Phratora (Coleoptera: Chrysomelidae). Evolution 1998, 52, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Saalas, U. Suomen Metsähyönteiset [Finnish Forest Insects]; WSOY: Helsinki, Finland, 1949; p. 719. [Google Scholar]
- Johnsen, L.G.; Skou, P.B.; Khakimov, B.; Bro, R. Gas chromatography—Mass spectrometry data processing made easy. J. Chromatogr. A 2017, 1503, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, Y.; Borges, R.M. Reducing the babel in plant volatile communication: Using the forest to see the trees. Plant Biol. 2010, 12, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.R.; Zalucki, M.P.; Madden, J.L.; Patel, V.S.; Paterson, S.C. Local dispersion of the Eucalyptus leaf beetle Chrysophtharta bimaculata (Coleoptera: Chrysomelidae), and implications for forest protection. J. Appl. Ecol. 1997, 34, 807–816. [Google Scholar] [CrossRef]
- Moayeri, H.R.S.; Ashouri, A.; Brodsgaard, H.F.; Enkegaard, A. Males of the predatory mirid bug Macrolophus caliginosus exploit plant volatiles induced by conspecifics as a sexual synomone. Entomol. Exp. Appl. 2007, 123, 49–55. [Google Scholar] [CrossRef]
- Brilli, F.; Ciccioli, P.; Frattoni, M.; Prestininzi, M.; Spanedda, A.F.; Loreto, F. Constitutive and herbivore-induced monoterpenes emitted by Populus × euroamericana leaves are key volatiles that orient Chrysomela populi beetles. Plant Cell Environ. 2009, 32, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Austel, N.; Bjorkman, C.; Hilker, M.; Meiners, T. Phenotypic plasticity in host plant preference of the willow leaf beetle Phratora vulgatissima: The impact of experience made by adults. Agric. For. Entomol. 2014, 16, 417–425. [Google Scholar] [CrossRef]
- Copolovici, L.; Kaennaste, A.; Remmel, T.; Vislap, V.; Niinemets, U. Volatile emissions from Alnus glutionosa induced by herbivory are quantitatively related to the extent of damage. J. Chem. Ecol. 2011, 37, 18–28. [Google Scholar] [CrossRef]
- Miresmailli, S.; Gries, R.; Gries, G.; Zamar, R.H.; Isman, M.B. Population density and feeding duration of cabbage looper larvae on tomato plants alter the levels of plant volatile emissions. Pest Manag. Sci. 2012, 68, 101–107. [Google Scholar] [CrossRef]
- Yli-Pirilä, P.; Copolovici, L.; Kannaste, A.; Noe, S.; Blande, J.D.; Mikkonen, S.; Klemola, T.; Pulkkinen, J.; Virtanen, A.; Laaksonen, A.; et al. Herbivory by an Outbreaking Moth Increases Emissions of Biogenic Volatiles and Leads to Enhanced Secondary Organic Aerosol Formation Capacity. Environ. Sci. Technol. 2016, 50, 11501–11510. [Google Scholar] [CrossRef] [Green Version]
- Kos, M.; Houshyani, B.; Overeem, A.; Bouwmeester, H.J.; Weldegergis, B.T.; van Loon, J.J.; Dicke, M.; Vet, L.E.M. Genetic engineering of plant volatile terpenoids: Effects on a herbivore, a predator and a parasitoid. Pest Manag. Sci. 2013, 69, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Ikeda, A.; Shiojiri, K.; Ozawa, R.; Shiki, K.; Nagai-Kunihiro, N.; Fujita, K.; Sugimoto, K.; Yamato, K.T.; Dohra, H.; et al. Identification of a hexenal reductase that modulates the composition of green leaf volatiles. Plant Physiol. 2018, 178, 552–564. [Google Scholar] [CrossRef] [Green Version]
- Mäntylä, E.; Alessio, G.A.; Blande, J.D.; Heijari, J.; Holopainen, J.K.; Laaksonen, T.; Piirtola, P.; Klemola, T. From plants to birds: Higher avian predation rates in trees responding to insect herbivory. PLoS ONE 2008, 3, e2832. [Google Scholar]
- Allmann, S.; Baldwin, I.T. Insects betray themselves in nature to predators by rapid isomerization of green leaf volatiles. Science 2010, 329, 1075–1078. [Google Scholar] [CrossRef] [PubMed]
- Loughrin, J.H.; Potter, D.A.; Hamilton-Kemp, T.R. Volative Compounds Induced by Herbivory Act as Aggregation Kairomones for the Japanese-Beetle (Popillia japonica Newman). J. Chem. Ecol. 1995, 21, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, G.; Cai, X.; Jin, S.; Gao, Y.; Chen, Z. The Tea Weevil, Myllocerinus aurolineatus, is Attracted to Volatiles Induced by Conspecifics. J. Chem. Ecol. 2010, 36, 388–395. [Google Scholar] [CrossRef]
- Copolovici, L.; Kaennaste, A.; Remmel, T.; Niinemets, U. Volatile organic compound emissions from Alnus glutinosa under interacting drought and herbivory stresses. Environ. Exp. Bot. 2014, 100, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Maja, M.M.; Kasurinen, A.; Yli-Pirila, P.; Joutsensaari, J.; Klemola, T.; Holopainen, T.; Holopainen, J.K. Contrasting responses of silver birch VOC emissions to short- and long-term herbivory. Tree Physiol. 2014, 34, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Kugimiya, S.; Shimoda, T.; Tabata, J.; Takabayashi, J. Present or Past Herbivory: A screening of volatiles released from Brassica rapa under caterpillar attacks as attractants for the solitary parasitoid, Cotesia vestalis. J. Chem. Ecol. 2010, 36, 620–628. [Google Scholar] [CrossRef]
Figure 1.
Olfactory bioassay illustration. (a) The entire experimental setup. (b) A closer look at the dual choice olfactory bioassays.
Figure 1.
Olfactory bioassay illustration. (a) The entire experimental setup. (b) A closer look at the dual choice olfactory bioassays.

Figure 2.
Attraction of P. laticollis adults to volatiles released from undamaged and damaged hybrid aspen seedlings. (a–d) Proportion of insects choosing the two odors, where the pie charts illustrate the relative proportion of insects that made or did not make a choice. (e–h) Average residence time (mean ± SEM) spent by insects in the two odor fields. Seedlings were subjected to feeding by leaf beetles for approximately 48 h, followed by dual-choice olfactory assays in Y-shaped olfactometers. The following odor pairs were tested for both female (♀) and male (♂) adults: (a,e) undamaged leaves versus charcoal filtered air, (b,f) undamaged leaves versus leaves damaged by male adults, (c,g) undamaged leaves versus leaves damaged by female adults, and (d,h) undamaged leaves versus leaves damaged by a mixture of female and male adults (♀+♂). Significant differences in proportion data and residence time were assessed by binomial tests and Wilcoxon matched-pair signed-rank tests, respectively. Asterisks indicate significance level: * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns, not significant.
Figure 2.
Attraction of P. laticollis adults to volatiles released from undamaged and damaged hybrid aspen seedlings. (a–d) Proportion of insects choosing the two odors, where the pie charts illustrate the relative proportion of insects that made or did not make a choice. (e–h) Average residence time (mean ± SEM) spent by insects in the two odor fields. Seedlings were subjected to feeding by leaf beetles for approximately 48 h, followed by dual-choice olfactory assays in Y-shaped olfactometers. The following odor pairs were tested for both female (♀) and male (♂) adults: (a,e) undamaged leaves versus charcoal filtered air, (b,f) undamaged leaves versus leaves damaged by male adults, (c,g) undamaged leaves versus leaves damaged by female adults, and (d,h) undamaged leaves versus leaves damaged by a mixture of female and male adults (♀+♂). Significant differences in proportion data and residence time were assessed by binomial tests and Wilcoxon matched-pair signed-rank tests, respectively. Asterisks indicate significance level: * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns, not significant.

Figure 3.
Emission of different groups of volatiles from hybrid aspen in response to leaf beetle attack. Seedlings were infested (orange bars) for approximately 48 h by female adults of leaf beetle (♀), or male adults (♂), or a mixture of females and males (♀+♂), with their respective control left uninfested (green bars). Volatiles were collected following insect olfactory bioassays. Asterisks indicate significant differences between control and damaged plants within each gender (p values < 0.05) after a paired T-test or Wilcoxon matched-pair signed-rank tests. Different upper- and lower-case letters indicate significant differences among genders within control and damage treatments, respectively, based on pairwise Wilcoxon–Mann–Whitney tests using Benjamini and Hochberg post-hoc correction following Kruskal–Wallis tests. Data represent mean ± SEM. n = 4 (♀), 5 (♂) and 6 (♀+♂). GLVs: green leaf volatiles. See Table S3 for breakdown of individual compounds within each compound class.
Figure 3.
Emission of different groups of volatiles from hybrid aspen in response to leaf beetle attack. Seedlings were infested (orange bars) for approximately 48 h by female adults of leaf beetle (♀), or male adults (♂), or a mixture of females and males (♀+♂), with their respective control left uninfested (green bars). Volatiles were collected following insect olfactory bioassays. Asterisks indicate significant differences between control and damaged plants within each gender (p values < 0.05) after a paired T-test or Wilcoxon matched-pair signed-rank tests. Different upper- and lower-case letters indicate significant differences among genders within control and damage treatments, respectively, based on pairwise Wilcoxon–Mann–Whitney tests using Benjamini and Hochberg post-hoc correction following Kruskal–Wallis tests. Data represent mean ± SEM. n = 4 (♀), 5 (♂) and 6 (♀+♂). GLVs: green leaf volatiles. See Table S3 for breakdown of individual compounds within each compound class.

Figure 4.
Random Forest (RF) classification analysis of volatile profiles from undamaged and damaged hybrid aspen plants. Seedlings were infested (orange symbols) for approximately 42 h by female adults of leaf beetle (♀), or male adults (♂), or a mixture of females and males (♀+♂), with their respective control left uninfested (green symbols). Volatiles were collected following insect olfactory bioassays. (a) Multidimensional scaling plot of RF model classifying volatile bouquets released from undamaged and damaged plants, independent of gender identity of the attacking leaf beetles. During 100,000 replicated tree constructions samples were always assigned correctly, with the out-of-bag (OOB) error being 0%. (b) Importance ranking of volatile compounds contributing to separation between undamaged and damaged plants based on mean decrease in accuracy (MDA) following RF analysis.
Figure 4.
Random Forest (RF) classification analysis of volatile profiles from undamaged and damaged hybrid aspen plants. Seedlings were infested (orange symbols) for approximately 42 h by female adults of leaf beetle (♀), or male adults (♂), or a mixture of females and males (♀+♂), with their respective control left uninfested (green symbols). Volatiles were collected following insect olfactory bioassays. (a) Multidimensional scaling plot of RF model classifying volatile bouquets released from undamaged and damaged plants, independent of gender identity of the attacking leaf beetles. During 100,000 replicated tree constructions samples were always assigned correctly, with the out-of-bag (OOB) error being 0%. (b) Importance ranking of volatile compounds contributing to separation between undamaged and damaged plants based on mean decrease in accuracy (MDA) following RF analysis.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, T.; Grauer-Gray, K.; Holopainen, J.K.; Blande, J.D. Herbivore Gender Effects on Volatile Induction in Aspen and on Olfactory Responses in Leaf Beetles. Forests 2020, 11, 638. https://doi.org/10.3390/f11060638
AMA Style
Li T, Grauer-Gray K, Holopainen JK, Blande JD. Herbivore Gender Effects on Volatile Induction in Aspen and on Olfactory Responses in Leaf Beetles. Forests. 2020; 11(6):638. https://doi.org/10.3390/f11060638
Chicago/Turabian StyleLi, Tao, Kristen Grauer-Gray, Jarmo K. Holopainen, and James D. Blande. 2020. "Herbivore Gender Effects on Volatile Induction in Aspen and on Olfactory Responses in Leaf Beetles" Forests 11, no. 6: 638. https://doi.org/10.3390/f11060638
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.