Monthly Radial Growth Model of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.), and the Relationships between Radial Increment and Climate Factors

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Plot Selection
2.3. Tree Increment Data
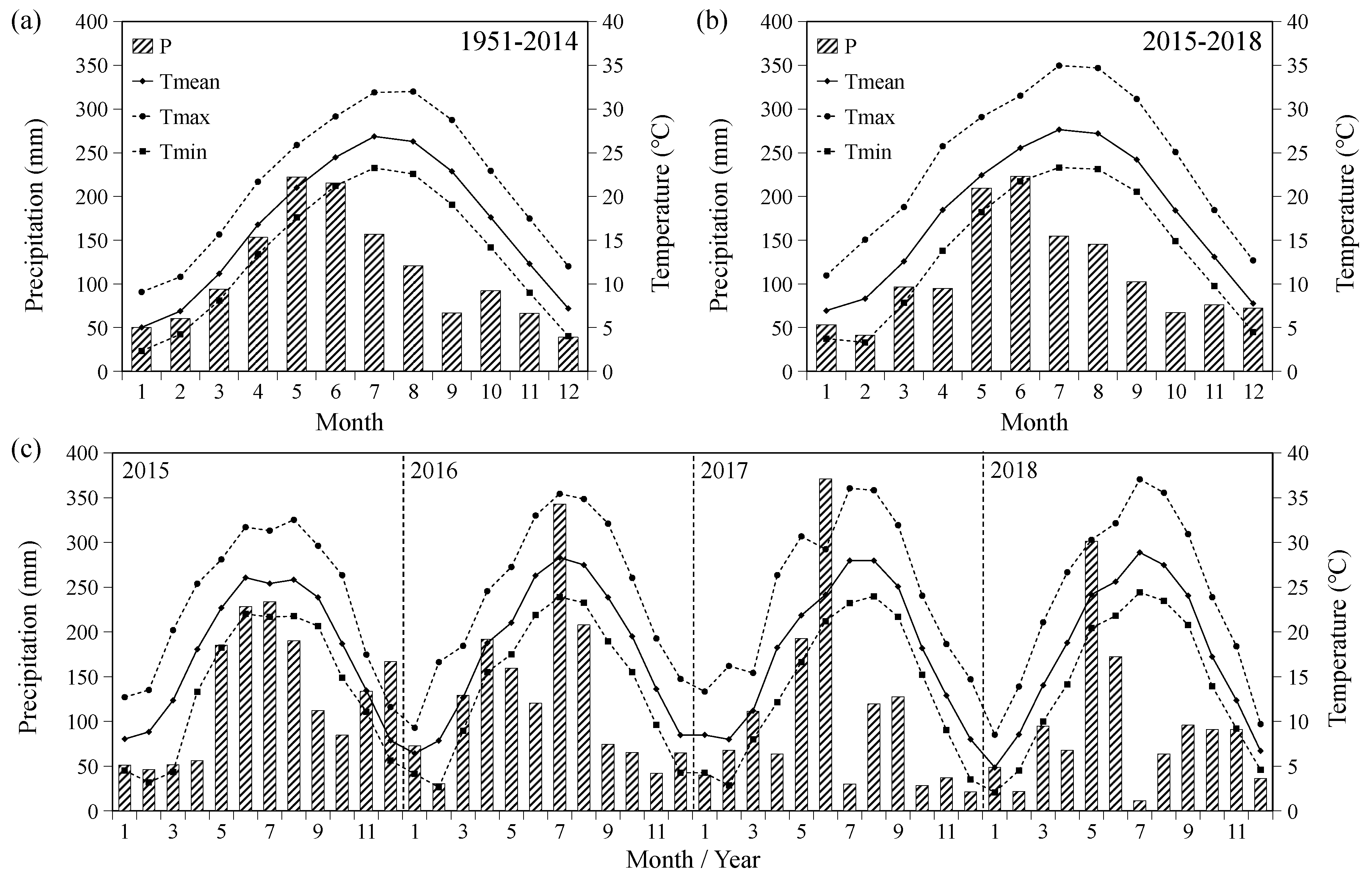
2.4. Climate Data
2.5. Data Analysis
2.5.1. Sample Grouping
2.5.2. Growth Pattern and Rate
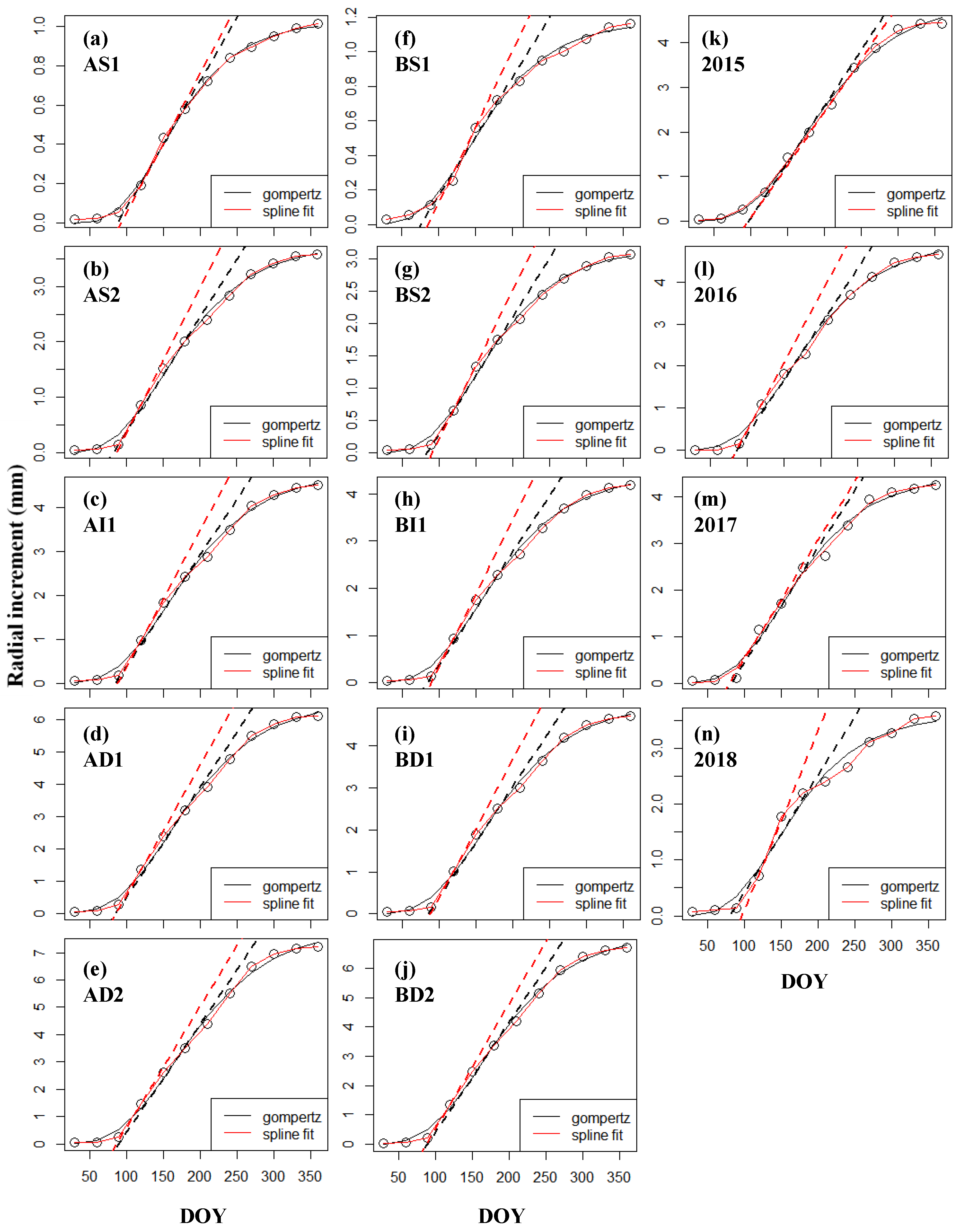
2.5.3. Growth Curves Modelling
2.5.4. Relationship between Climate and Radial Growth
3. Results
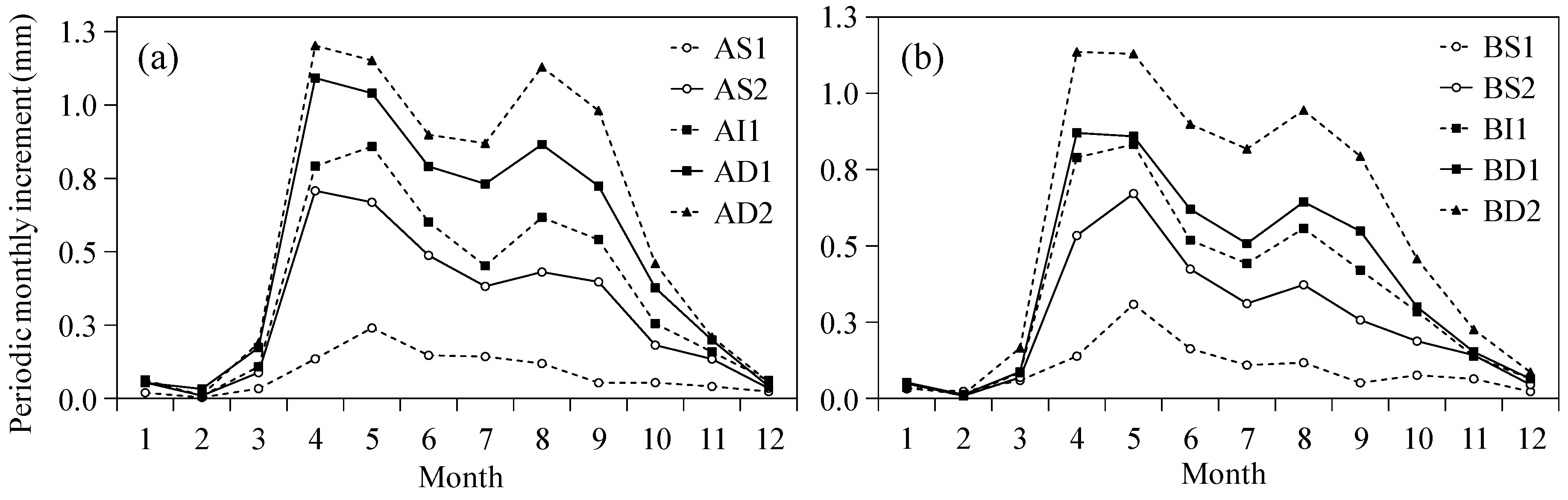
3.1. Monthly Radial Growth
3.2. Growth Curves’ Modelling
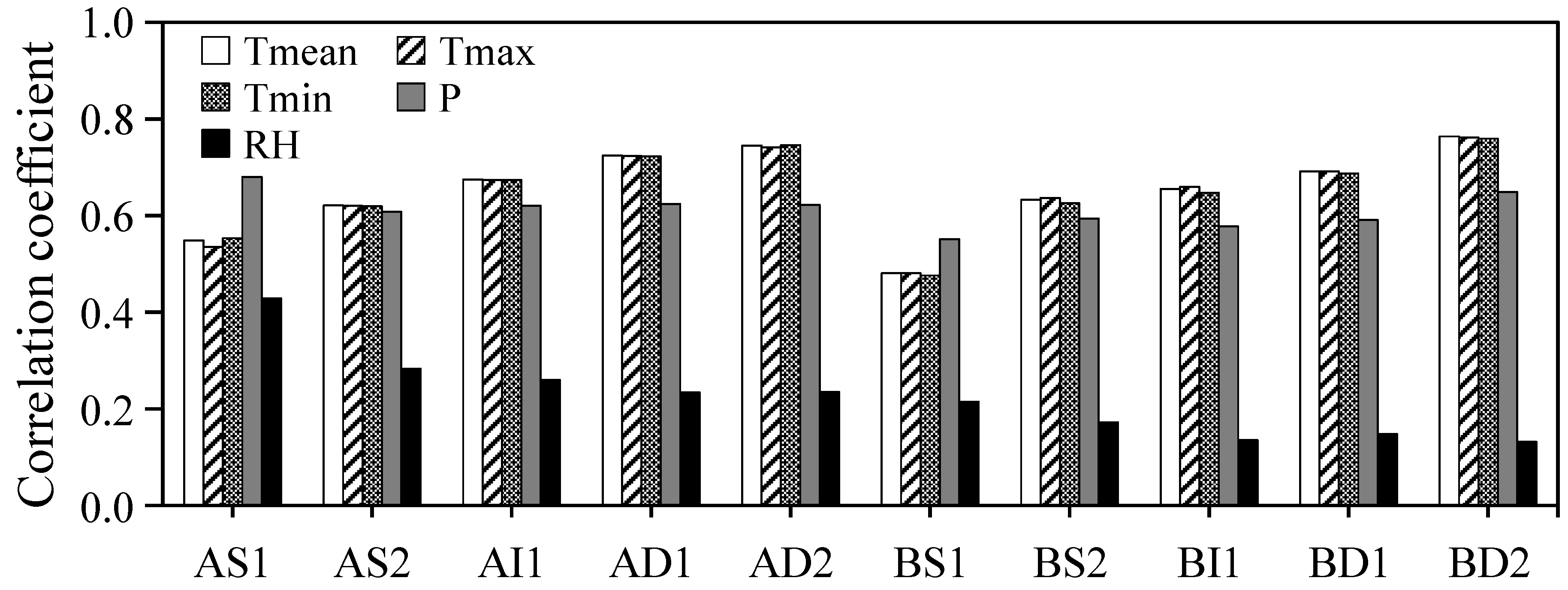
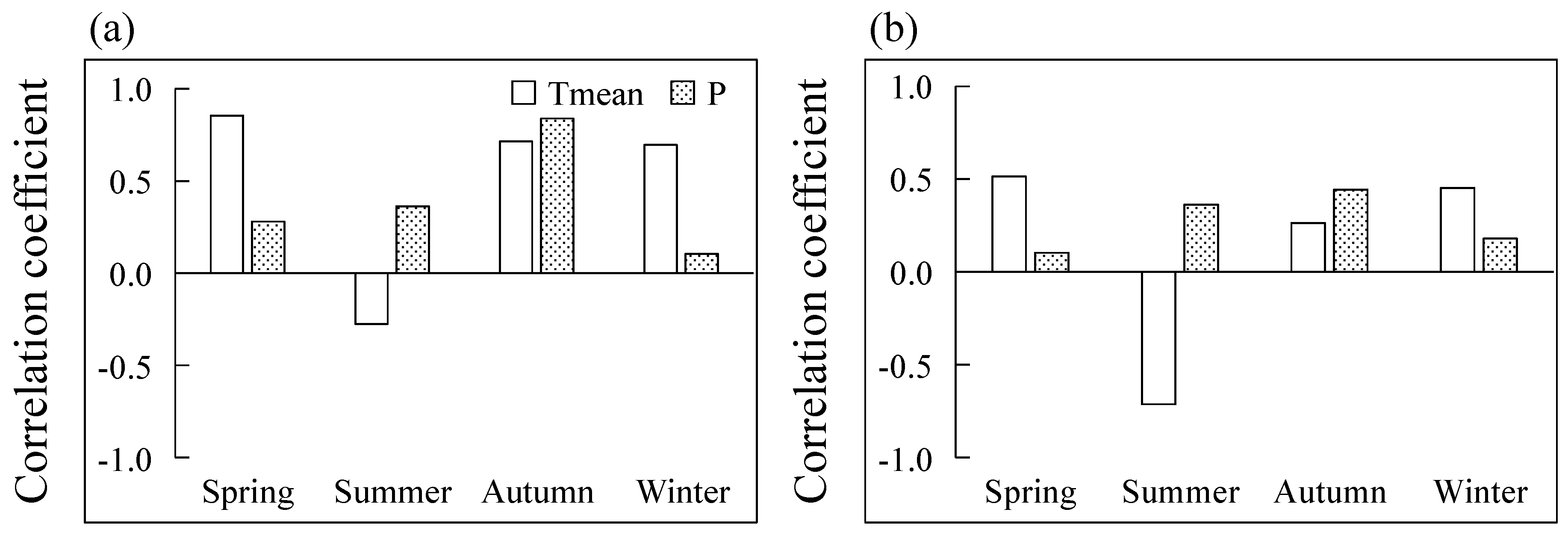
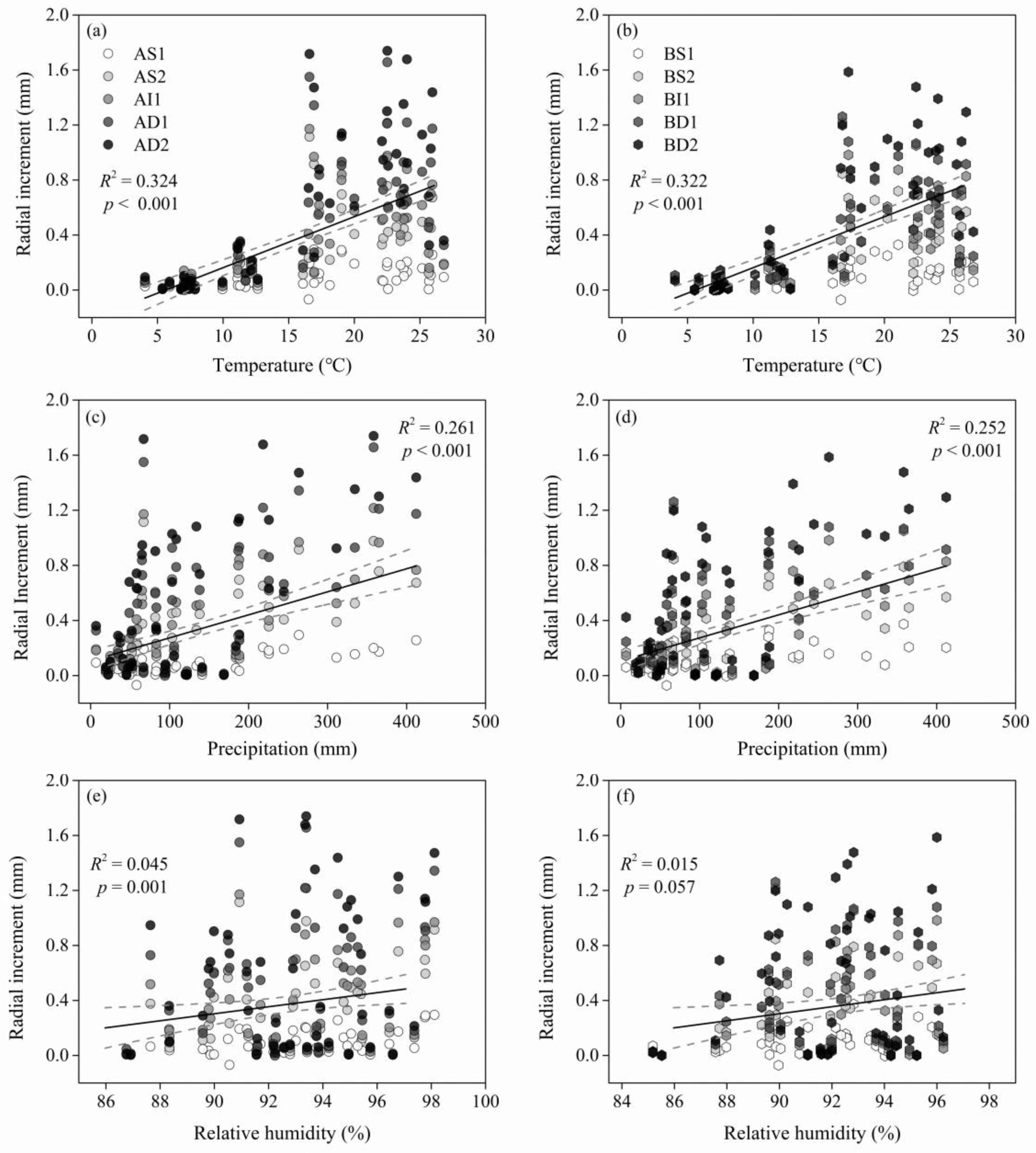
3.3. Radial Increment Responses of Climate Factors
4. Discussion
4.1. Influence of Trees’ Social Status on Radial Growth
4.2. Effects of Climate Factors on the Double-Peaks Pattern
4.3. Model Curves for Radial Growth
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thuiller, W.; Lavergne, S.; Roquet, C.; Boulangeat, I.; Lafourcade, B.; Araujo, M.B. Consequences of climate change on the tree of life in europe. Nature 2011, 470, 531–534. [Google Scholar] [CrossRef]
- Prentice, I.C.; Heimann, M.; Sitch, S. The carbon balance of the terrestrial biosphere: Ecosystem models and atmospheric observations. Ecol. Appl. 2000, 10, 1553–1573. [Google Scholar] [CrossRef]
- Szeicz, J.M. Growth trends and climatic sensitivity of trees in the north Patagonian rain forest of Chile. Can. J. For. Res. 1997, 27, 1003–1014. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef]
- Rathgeber, C.B.; Rossi, S.; Bontemps, J.D. Cambial activity related to tree size in a mature silver-fir plantation. Ann. Bot. 2011, 108, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, V.; Mcmahon, S.M.; Detto, M.; Lutz, J.A.; Davies, S.J.; Changyang, C.H.; Kristina, J.A.T. Tree circumference dynamics in four forests characterized using automated dendrometer bands. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Oberhuber, W.; Kofler, W. Topographic influences on radial growth of scots pine (Pinus sylvestris L.) at small spatial scales. Plant Ecol. 2000, 146, 229–238. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Saranpää, P. Seasonal changes in stem radius and production of new tracheids in Norway spruce. Tree Physiol. 2003, 23, 959. [Google Scholar] [CrossRef]
- Frank, D.; Esper, J. Characterization and climate response patterns of a high-elevation, multi-species tree-ring network in the European Alps. Dendrochronologia 2005, 22, 107–121. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Griesbauer, H.P.; Green, D.S. Growth responses of three coexisting conifer species to climate across wide geographic and climate ranges in Yukon and British Columbia. For. Ecol. Manag. 2010, 259, 514–523. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Formentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature (London) 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Gou, X.H.; Pederson, N.; Zhang, F.; Niu, H.; Zhao, S.; Wang, F. Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiol. 2018. [Google Scholar] [CrossRef]
- Tsai, C.C.; Hung, L.F.; Chung, J.D.; Chen, S.J.; Chien, C.T.; Kao, W.Y.; Kuo-Huang, L.L. Radial growth of Cinnamomum kanehirae, hayata displays a larger temperature sensitivity in dominant than codominant trees. Ann. For. Sci. 2018, 75, 52. [Google Scholar] [CrossRef]
- Biondi, F.; Hartsough, P. Using automated point dendrometers to analyze tropical treeline stem growth at Nevado de Colima, Mexico. Sensors 2010, 10, 5827–5844. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Morin, H. Application of the Gompertz equation for the study of xylem cell development. Dendrochronologia 2004, 21, 33–39. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T. Assessment of cambial activity and xylogenesis by microsampling tree species: An example at the alpine timberline. IAWA J. 2006, 27, 383–394. [Google Scholar] [CrossRef]
- Michelot, A.; Simard, S.; Rathgeber, C.; Dufrêne, E.; Damesin, C. Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol. 2012, 32, 1033. [Google Scholar] [CrossRef]
- Oswald, S.A.; Nisbet, I.C.T.; Chiaradia, A.; Arnold, J.M. FlexParamCurve: R package for flexible fitting of non-linear parametric curves. Methods Ecol. Evol. 2012, 3, 1073–1077. [Google Scholar] [CrossRef]
- Duan, A.; Zhang, J.; Zhang, X.; He, C. Relationship between modelling accuracy and inflection point attributes of several equations while modelling stand diameter distributions. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Linares, J.C.; Camarero, J.J.; Carreira, J.A. Plastic responses of Abies pinsapo xylogenesis to drought and competition. Tree Physiol. 2009, 29, 1525–1536. [Google Scholar] [CrossRef]
- Güney, A.; Küppers, M.; Rathgeber, C.; Sahin, M.; Zimmermann, R. Intra-annual stem growth dynamics of lebanon cedar along climatic gradients. Trees 2017, 19, 442–450. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Borghetti, M.M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 2006, 170, 301–310. [Google Scholar] [CrossRef]
- Li, X.; Liang, E.; Gričar, J.; Prislan, P.; Rossi, S.; Čufar, K. Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan plateau. Tree Physiol. 2013, 33, 48. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Urbinati, C.; Carrer, M. Daily weather response of balsam fir (Abies balsamea, (L.) mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees 2003, 17, 477–484. [Google Scholar] [CrossRef]
- Thibeaultmartel, M.; Krause, C.H.; Rossi, S. Cambial activity and intra-annual xylem formation in roots and stems of Abies balsamea and Picea mariana. Ann. Bot. 2008, 102, 667–674. [Google Scholar] [CrossRef]
- Ni, J. Impacts of climate change on Chinese ecosystems: Key vulnerable regions and potential thresholds. Reg. Environ. Chang. 2011, 11, 49–64. [Google Scholar] [CrossRef]
- Lucas, C.; Puchi, P.; Profumo, L.; Ferreira, A.; Muñoz, A. Effect of climate on tree growth in the pampa biome of southeastern south america: First tree-ring chronologies in uruguay. Dendrochronologia 2018. [Google Scholar] [CrossRef]
- Wen, H.F.; Deng, X.W.; Zhang, Y.F.; Wei, X.C.; Wang, G.J.; Zhou, B.; Xiang, W.H.; Zhu, N.H. Cunninghamia lanceolata variant with red-heart wood: A mini-review. Dendrobiology 2018, 79, 156–167. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, J.; Zeng, S.; Wu, D.; Jacobs, D.F.; Sloan, J.L. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata, plantations in south China. J. Soils Sediments 2016, 17, 1–9. [Google Scholar] [CrossRef]
- Li, Y.F.; Deng, X.W.; Zhang, Y.F.; Huang, Y.Q.; Wang, C.Y.; Xiang, W.H.; Xiao, F.M.; Wei, X.C. Chemical Characteristics of Heartwood and Sapwood of Red Heart Chinese Fir (Cunninghamia lanceolata). For. Prod. J. 2019, 69, 103–109. [Google Scholar] [CrossRef]
- Lu, Y.; Coops, N.C.; Wang, T.; Wang, G. A process-based approach to estimate Chinese fir (Cunninghamia lanceolata) distribution and productivity in southern China under climate change. Forests 2015, 6, 360–379. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Deng, X.W.; Huang, Y.Q.; Li, Y.F.; Xiang, W.H.; Yan, W.D. Quantification of individual tree competition index taking Chinese-fir plantations in subtropical low hilly area as an example. Pol. J. Ecol. 2019, 67, 1–16. [Google Scholar] [CrossRef]
- Song, J. A brief discussion on the relationship between the growth of Chinese fir and the environmental conditions. Sci. Technol. West China 2008, 7, 54–55. [Google Scholar]
- Tian, D.L.; Shen, Y.; Kang, W.X.; Deng, X.W. Characteristics of nutrient cycling in first and second rotations of Chinese fir plantations. Acta Ecol. Sin. 2011, 31, 5025–5032. [Google Scholar]
- Vitas, A. Seasonal Growth Variations of Pine, Spruce, and Birch Recorded by Band Dendrometers in NE Lithuania. Balt. For. 2011, 17, 197–204. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R. Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Kahm, M.; Hasenbrink, G.; Lichtenbergfrate, H.; Ludwig, J.; Kschischo, M. Grofit: Fitting biological growth curves. J. Stat. Softw. 2010, 33, 1–21. [Google Scholar] [CrossRef]
- Lin, S.Z.; Huang, S.G.; Hong, W. Interspecific competition of Chinese fir and its mixed species in Chinese fir and broad-leaved mixed forest. Sci. Silva Sin. 2004, 40, 160–164. [Google Scholar]
- Grotta, A.T.; Gartner, B.L.; Radosevich, S.R.; Huso, M. Influence of red alder competition on cambial phenology and latewood formation in Douglas-fir. IAWA J. 2005, 26, 309–324. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H. Intra-annual tracheid production in balsam fir stems and the effect of meteorological variables. Trees 2005, 19, 402–408. [Google Scholar] [CrossRef]
- Rossi, S.; Girard, M.J.; Morin, H. Lengthening of the duration of xylogenesis engenders disproportionate increases in xylem production. Glob. Chang. Biol. 2014, 20, 2261. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Deslauriers, A.; Griçar, J.; Seo, J.W.; Rathgeber, C.B.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Dong, M.Y.; Jiang, Y.; Yang, H.C.; Wang, M.C.; Zhang, W.T.; Guo, Y.Y. Dynamics of stem radial growth of Picea meyeri during the growing season at the treeline of Luya Mountain, China. Chin. J. Plant Ecol. 2012, 36, 956–964. [Google Scholar] [CrossRef]
- Kellomäki, S.; Wang, K.Y. Growth and resource use of birch seedlings under elevated carbon dioxide and temperature. Ann. Bot. 2001, 87, 669–682. [Google Scholar] [CrossRef]
- Strand, M.; Lundmark, T.; Söderbergh, I.; Mellander, P.E. Impacts of seasonal air and soil temperatures on photosynthesis in scots pine trees. Tree Physiol. 2002, 22, 839. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Nie, Y.Q.; Wen, F. Seasonal Dynamics of Stem Radial Increment of Pinus taiwanensis Hayata and Its Response to Environmental Factors in the Lushan Mountains, Southeastern China. Forests 2018, 9, 387. [Google Scholar] [CrossRef]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Seasonal and daily cycles of stem radial variation of Pinus pinaster in a drought-prone environment. Agric. For. Meteorol. 2013, 180, 173–181. [Google Scholar] [CrossRef]
- Oladi, R.; Elzami, E.; Pourtahmasi, K.; Bräuning, A. Weather factors controlling growth of Oriental beech are on the turn over the growing season. Eur. J. For. Res. 2017, 136, 345–356. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, B.Q.; Dong, M.Y.; Huang, Y.M.; Wang, M.C.; Wang, B. Response of daily stem radial growth of Platycladus orientalis to environmental factors in a semi-arid area of North China. Trees 2015, 29, 87–96. [Google Scholar] [CrossRef]
- Zweifel, R.; Zimmermann, L.; Zeugin, F.; Newbery, D.M. Intra-annual radial growth and water relations of trees: Implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [CrossRef]
- Urrutia-Jalabert, R.; Rossi, S.; Deslauriers, A.; Malhi, Y.; Lara, A. Environmental correlates of stem radius change in the endangered Fitzroya cupressoides forests of southern Chile. Agric. For. Meteorol. 2015, 200, 209–221. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Groups | Number | Age (years) | Slope (°) | Slope Aspect | Average DBH | Max DBH | Min DBH | Variance |
|---|---|---|---|---|---|---|---|---|---|
| A | AS1 | 22 | 22 | 25 | Northeast | 15.32 | 18.93 | 12.50 | 4.96 |
| A | AS2 | 20 | 22 | 25 | Northeast | 18.10 | 23.23 | 14.26 | 4.72 |
| A | AI1 | 17 | 22 | 25 | Northeast | 21.73 | 23.98 | 19.21 | 1.61 |
| A | AD1 | 14 | 22 | 25 | Northeast | 23.56 | 25.87 | 20.05 | 1.84 |
| A | AD2 | 9 | 22 | 25 | Northeast | 25.63 | 26.72 | 24.26 | 0.74 |
| B | BS1 | 23 | 30 | 27 | Northeast | 17.12 | 19.85 | 13.55 | 2.37 |
| B | BS2 | 20 | 30 | 27 | Northeast | 20.63 | 22.64 | 17.22 | 2.72 |
| B | BI1 | 16 | 30 | 27 | Northeast | 24.11 | 26.99 | 19.72 | 2.73 |
| B | BD1 | 12 | 30 | 27 | Northeast | 26.85 | 33.40 | 25.40 | 4.63 |
| B | BD2 | 9 | 30 | 27 | Northeast | 29.39 | 36.49 | 26.50 | 8.69 |
| Items | A | μ | λ | ||||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | ||
| Year | 2015 | 4.901 | 0.140 | 0.025 | 0.001 | 97.627 | 4.180 |
| 2016 | 4.959 | 0.146 | 0.026 | 0.002 | 87.183 | 4.957 | |
| 2017 | 4.470 | 0.168 | 0.025 | 0.002 | 82.967 | 6.833 | |
| 2018 | 3.592 | 0.161 | 0.021 | 0.002 | 81.016 | 8.784 | |
| Group | AS1 | 1.027 | 0.015 | 0.006 | 0.000 | 88.612 | 2.964 |
| AS2 | 3.743 | 0.095 | 0.021 | 0.001 | 82.675 | 4.627 | |
| AI1 | 4.791 | 0.134 | 0.025 | 0.001 | 85.232 | 4.763 | |
| AD1 | 6.572 | 0.174 | 0.034 | 0.002 | 86.121 | 4.400 | |
| AD2 | 7.893 | 0.255 | 0.040 | 0.002 | 90.275 | 4.953 | |
| BS1 | 1.175 | 0.030 | 0.007 | 0.000 | 76.824 | 5.201 | |
| BS2 | 3.161 | 0.076 | 0.018 | 0.001 | 84.360 | 4.521 | |
| BI1 | 4.391 | 0.125 | 0.024 | 0.001 | 84.708 | 5.021 | |
| BS1 | 5.009 | 0.143 | 0.027 | 0.002 | 86.255 | 4.818 | |
| BS2 | 7.194 | 0.186 | 0.038 | 0.002 | 89.684 | 4.162 | |
| Factors Site A | A | μ | λ | |||
| F | p | F | p | F | p | |
| Year | 9.472 | 0.002 ** | 1.339 | 0.308 | 19.989 | <0.001 *** |
| Group | 416.193 | <0.001 *** | 201.847 | <0.001 *** | 0.009 | 0.926 |
| Year × Group | 2.908 | 0.078 | 1.030 | 0.414 | 0.774 | 0.531 |
| Factors Site B | A | μ | λ | |||
| F | p | F | p | F | p | |
| Year | 7.298 | 0.005 ** | 4.241 | 0.029 * | 6.371 | 0.008 ** |
| Group | 351.113 | <0.001 *** | 234.306 | <0.001 *** | 9.730 | 0.009 ** |
| Year × Group | 4.452 | 0.025 * | 2.316 | 0.128 | 2.402 | 0.119 |
| Intercept | Tmean | P | Tmax | Tmin | RH | |
|---|---|---|---|---|---|---|
| AS1 | −1.0648 * | −0.0263 | 0.0003 ** | 0.0303 | - | 0.0101 * |
| AS2 | −3.0882 * | −0.1251 * | 0.0010 * | 0.1416 * | - | 0.0274 |
| AI1 | −3.0692 | −0.1256 | 0.0012 * | 0.1480 * | - | 0.0265 |
| AD1 | −3.5633 | −0.1499 | 0.0015 ** | 0.1832 * | - | 0.0296 |
| AD2 | −0.4898 *** | - | 0.0020 *** | 0.0430 *** | - | - |
| BS1 | −1.1378 | −0.0478 * | 0.0003 * | 0.0506 * | - | 0.0107 |
| BS2 | −3.0093 * | −0.1305 * | 0.0008 * | 0.1422 ** | - | 0.0274 * |
| BI1 | −3.5720 * | −0.1618 * | 0.0010 * | 0.1796 ** | - | 0.0318 |
| BD1 | −3.6035 | −0.1371 | 0.0011 * | 0.1601 * | - | 0.0324 |
| BD2 | −3.5852 | −0.1415 | 0.0017 ** | 0.1762 * | - | 0.0304 |
| Items | Site A | Site B | ||
|---|---|---|---|---|
| F | p | F | p | |
| Temperature | 186.92 | <0.001 *** | 182.44 | <0.001 *** |
| Group | 27.38 | <0.001 *** | 26.22 | <0.001 *** |
| Temperature × Group | 12.41 | <0.001 *** | 12.24 | <0.001 *** |
| Precipitation | 120.72 | <0.001 *** | 113.99 | <0.001 *** |
| Group | 21.99 | <0.001 *** | 20.98 | <0.001 *** |
| Precipitation × Group | 5.99 | <0.001 *** | 6.27 | <0.001 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Deng, X.; Zhao, Z.; Xiang, W.; Yan, W.; Ouyang, S.; Lei, P. Monthly Radial Growth Model of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.), and the Relationships between Radial Increment and Climate Factors. Forests 2019, 10, 757. https://doi.org/10.3390/f10090757
Huang Y, Deng X, Zhao Z, Xiang W, Yan W, Ouyang S, Lei P. Monthly Radial Growth Model of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.), and the Relationships between Radial Increment and Climate Factors. Forests. 2019; 10(9):757. https://doi.org/10.3390/f10090757
Chicago/Turabian StyleHuang, Yaqi, Xiangwen Deng, Zhonghui Zhao, Wenhua Xiang, Wende Yan, Shuai Ouyang, and Pifeng Lei. 2019. "Monthly Radial Growth Model of Chinese Fir (Cunninghamia lanceolata (Lamb.) Hook.), and the Relationships between Radial Increment and Climate Factors" Forests 10, no. 9: 757. https://doi.org/10.3390/f10090757