Intra- and Inter-Modular Connectivity Alterations in the Brain Structural Network of Spinocerebellar Ataxia Type 3

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Image Acquisition and Cortical Feature-Based Structural Network
2.3. FD Analysis and Brain Structural Network
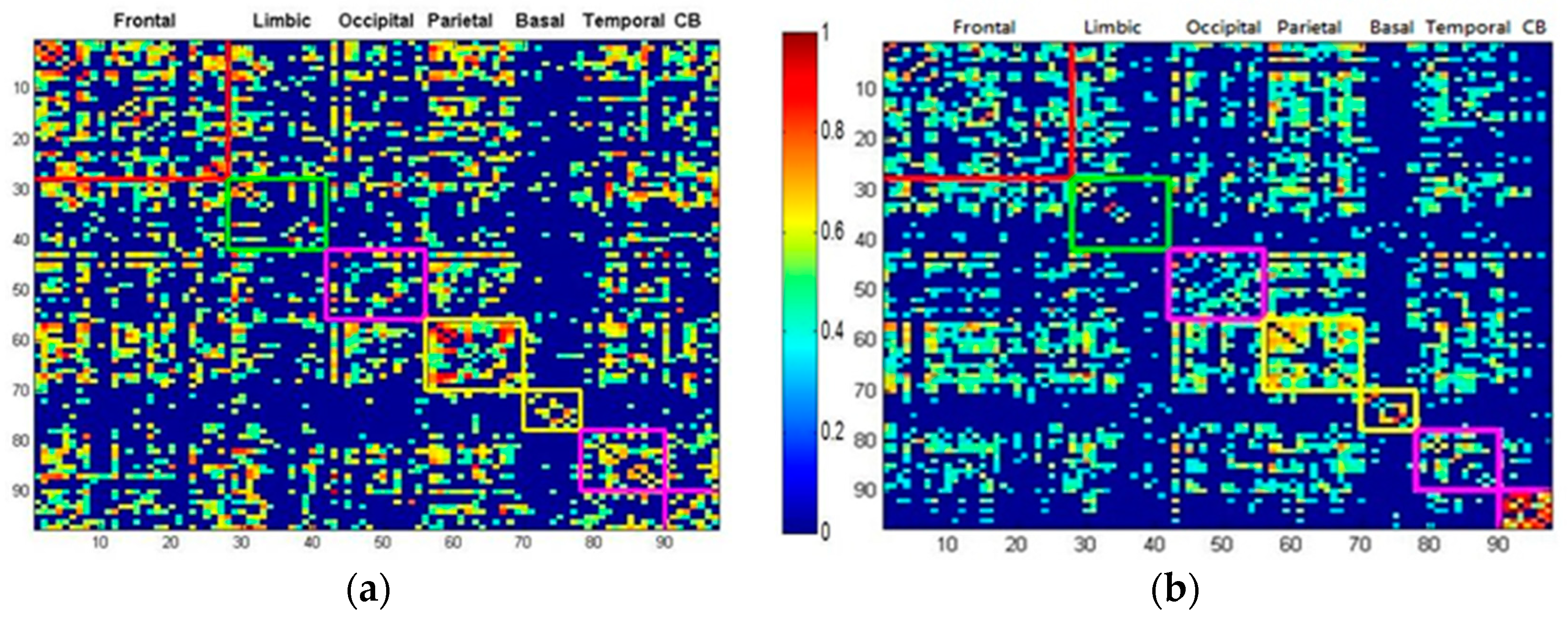
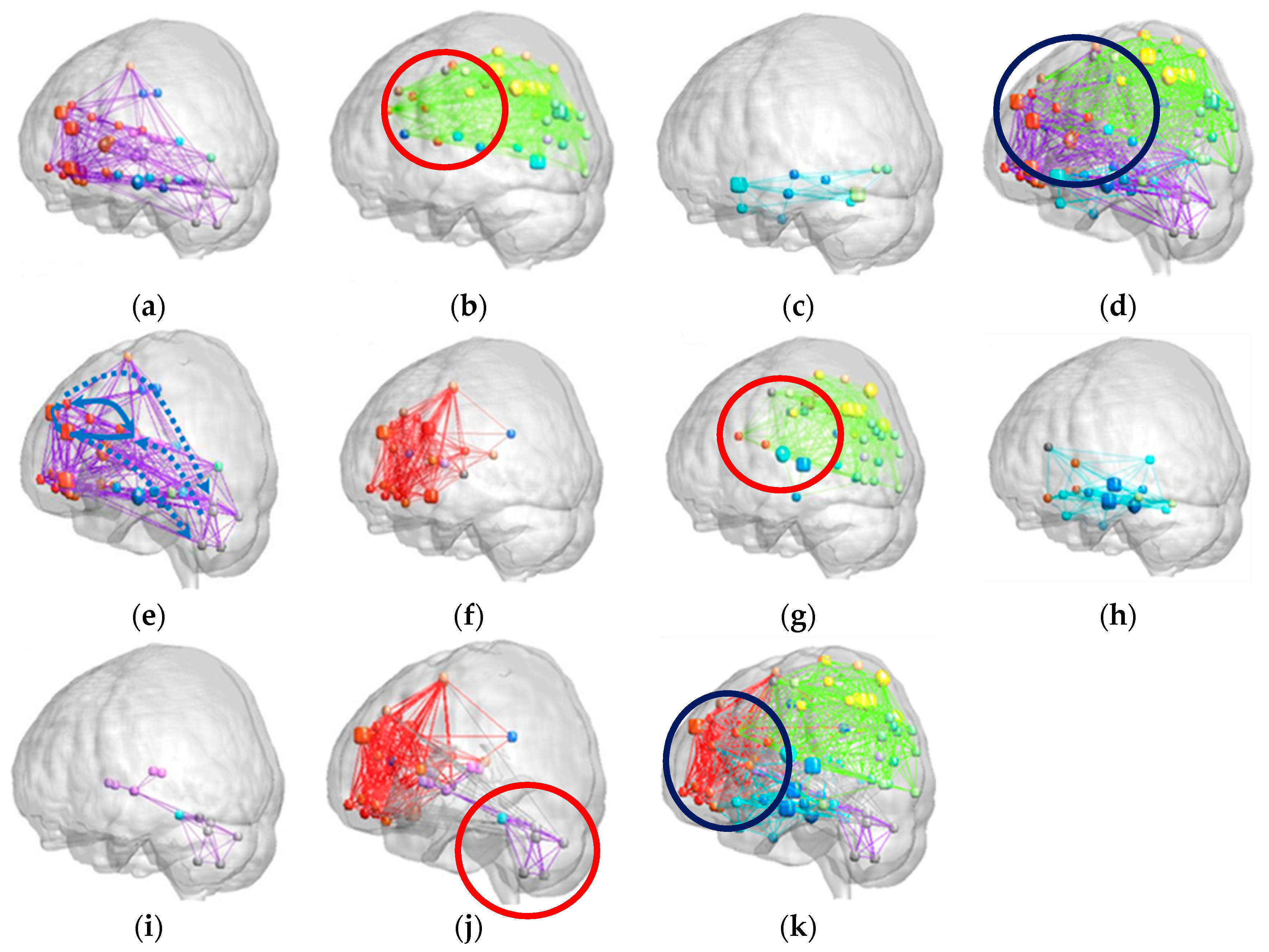
2.4. Modular Analysis
2.4.1. Intra-Modular Connectivity Analysis
2.4.2. Inter-Modular Connectivity Analysis
2.5. Statistical Analysis
3. Results
3.1. Patients with SCA3 Exhibited Significant Lateralized Atrophy with Extensive Involvement in the Left Supratentorial Regions
3.2. Patients with SCA3 Exhibited Lower Modularity Values and Less Dense Modular Networks
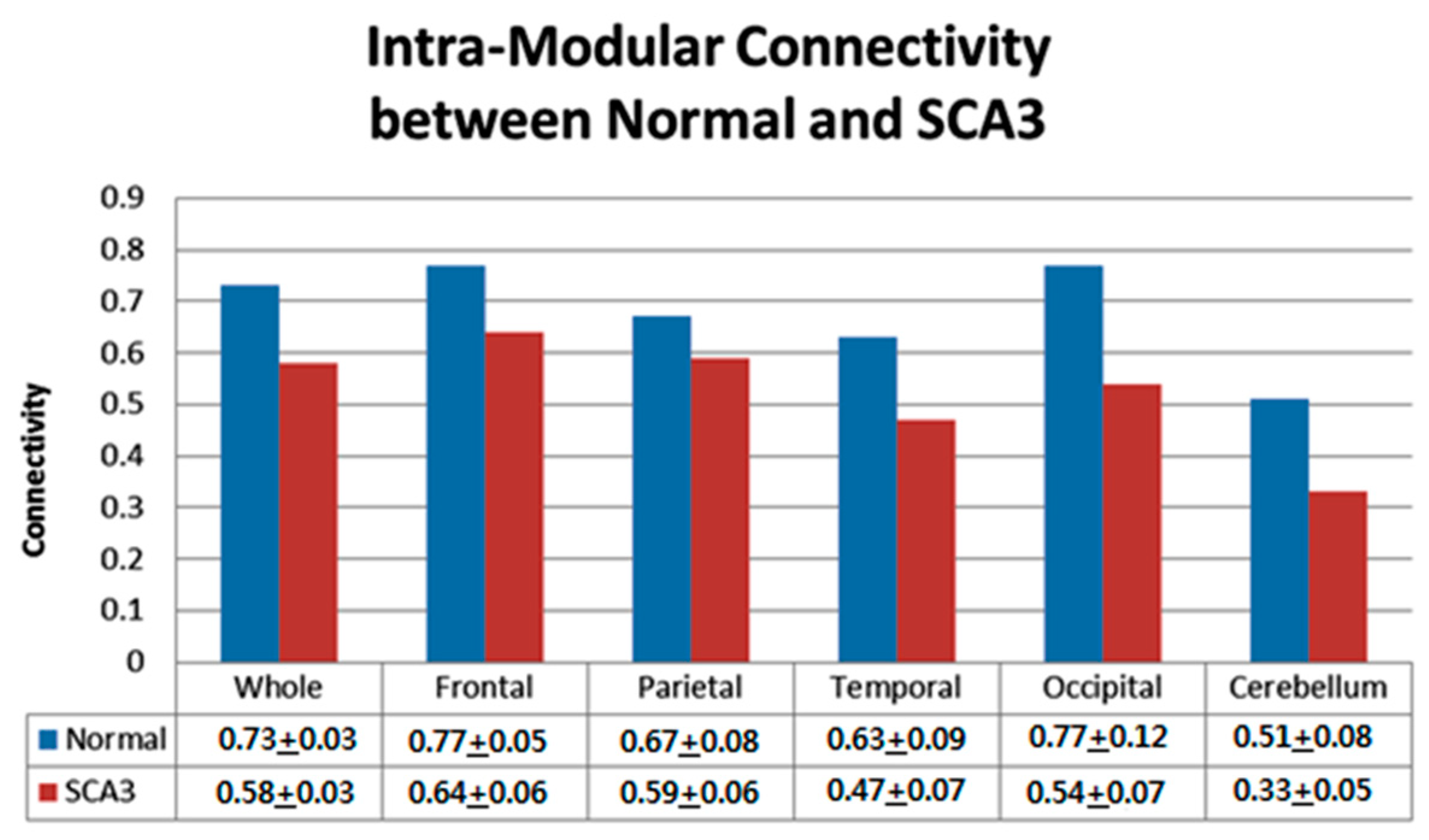
3.3. Significantly Decreased Intra-Modular Connectivity in Patients with SCA3
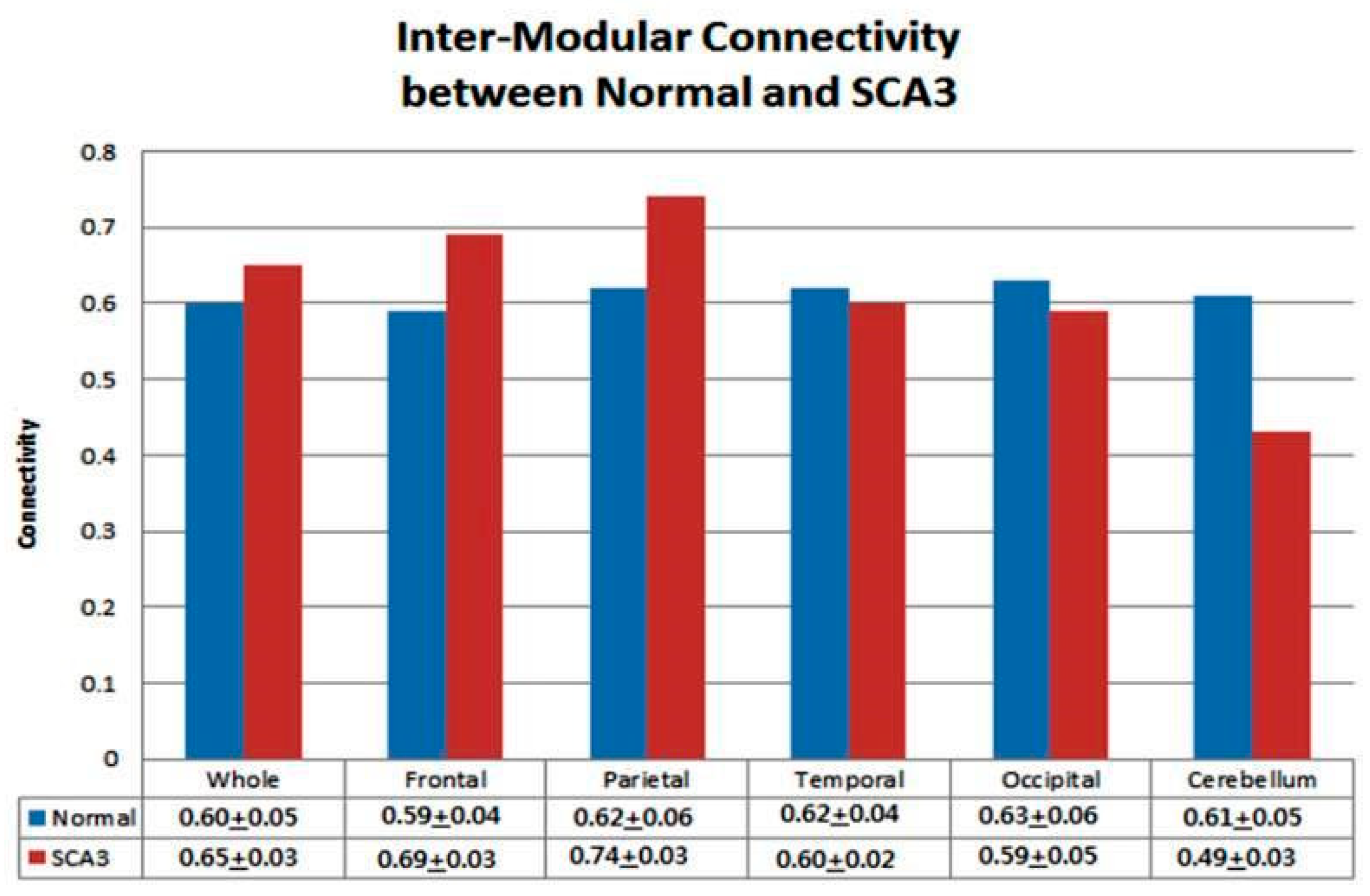
3.4. Significantly Increased Inter-Modular Connectivity in Patients with SCA3
4. Discussion
4.1. Patients with SCA3 Exhibited Cerebellar Cognitive Affective Syndrome (CCAS) and Mnemonic and Emotional Function Preservation
4.2. Increased Inter-Modular Connectivity in Supratentorial Regions in Patients with SCA3
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Riess, O.; Rub, U.; Pastore, A. SCA3: Neurological features, pathogenesis and animal models. Cerebellum 2008, 7, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Roeske, S.; Filla, I.; Heim, S.; Amunts, K.; Helmstaedter, C.; Wullner, U.; Wagner, M.; Klockgether, T.; Minnerop, M. Progressive cognitive dysfunction in spinocerebellar ataxia type 3. Mov. Disord. 2013, 8, 1435–1438. [Google Scholar] [CrossRef] [PubMed]
- Klinke, I.; Minnerop, M.; Schmitz-Hübsch, T.; Hendriks, M.; Klockgether, T.; Wüllner, U.; Helmstaedter, C. Neuropsychological features of patients with spinocerebellar ataxia (SCA) types 1, 2, 3, and 6. Cerebellum 2010, 9, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Fair, D.A.; Cohen, A.L.; Power, J.D.; Dosenbach, N.U.F.; Church, J.A.; Miezin, F.M.; Schlaggar, B.L.; Petersen, S.E. Functional Brain Networks Develop from a “Local to Distributed” Organization. PLoS Comput. Biol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Friston, K. Structural and functional brain networks: From connections to cognition. Science 2013, 342. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, S.; Barthelemy, M. Resolution limit in community detection. Proc. Natl. Acad. Sci. USA 2007, 104, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; He, Y.; Rosa-Neto, P.; Gong, G.; Evans, A.C. Age-related alterations in the modular organization of structural cortical network by using cortical thickness from MRI. NeuroImage 2011, 56, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D.; Pandya, D.N. Anatomic organization of the basilar pontine projections from prefrontal cortices in rhesus monkey. J. Neurosci. 1997, 17, 438–458. [Google Scholar] [CrossRef] [PubMed]
- Doyon, J.; Penhume, V.; Ungerleider, L.G. Distinct contribution of the cortico-striatal and cortico-cerebellar systems to motor skill learning. Neuropsychologia 2003, 41, 252–262. [Google Scholar] [CrossRef]
- Seeley, W.W.; Crawford, R.K.; Zhou, J.; Miller, B.L.; Greicius, M.D. Neurodegenerative diseases target large-scale human brain networks. Neuron 2009, 62, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Dagher, A.; Chen, Z. Impaired small-world efficiency in structural cortical networks in multiple sclerosis associated with white matter lesion load. Brain 2009, 132, 3366–3379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supekar, K.; Menon, V.; Rubin, D. Network analysis of intrinsic functional brain connectivity in Alzheimer’s disease. PLoS Comput. Biol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; He, Y.; Rosa-Neto, P.; Germann, J.; Evans, A.C. Revealing modular architecture of human brain structural networks by using cortical thickness from MRI. Cereb. Cortex 2008, 18, 2374–2381. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.G.; Wang, D.; Liu, K.; Weng, J.; Guan, Y.; Chan, K.C.C.; Chu, W.C.W.; Shi, L. Brain structure network analysis in patients with obstructive sleep apnea. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Sanabria-Diaz, G.; Melie-García, L.; Iturria-Medina, Y.; Alemán-Gómez, Y.; Hernández-González, G.; Valdés-Urrutia, L.; Galán, L.; Valdés-Sosa, P. Surface area and cortical thickness descriptors reveal different attributes of the structural human brain networks. NeuroImage 2010, 50, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Shyu, K.K.; Jao, C.W.; Wang, Z.Y.; Soong, B.W.; Wu, H.M.; Wang, P.S. Fractal dimension analysis for quantifying cerebellar morphological change of multiple system atrophy of the cerebellar type (MSA-C). NeuroImage 2010, 49, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.H.; Yoon, U.; Lee, K.J.; Shin, Y.W.; Lee, J.M.; Kim, I.Y.; Ha, K.S.; Kim, S.I.; Kwon, J.S. Fractal dimension of cerebral cortical surface in schizophrenia and obsessive–compulsive disorder. Neurosci. Lett. 2005, 384, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Sandu, A.L.; Rasmussen, I.A.; Lundervold, A.; Frank, K.F.; Neckelmann, G.; Hugdahl, K.; Specht, K. Fractal dimension analysis of MR images reveals grey matter structure irregularities in schizophrenia. Comput. Med. Imaging Graph. 2008, 32, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Kojadinovic, I.; Yan, J. Modeling multivariate distributions with continuous margins using the copula R package. J. Stat. Softw. 2010, 34, 1–20. [Google Scholar] [CrossRef]
- Newman, M.E. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [Green Version]
- Meunier, D.; Lambiotte, R.; Fornito, A.; Ersche, K.D.; Bullmore, E.T. Hierarchical modularity in human brain functional networks. Front. Neuroinform. 2009, 30. [Google Scholar] [CrossRef]
- Guimerà, R.; Amaral, L.A.N. Functional Cartography of Complex Metabolic Networks. Nature 2005, 433, 895–900. [Google Scholar] [CrossRef]
- Bullmore, E.T.; Suckling, J.; Overmeyer, S.; Rabe-Hesketh, S.; Taylor, E.; Brammer, M.J. Global, voxel, and cluster tests, by theory and permutation, for a difference between two groups of structural MR images of the brain. IEEE Trans. Med. Imaging 1999, 18, 32–42. [Google Scholar] [CrossRef]
- Hayhow, B.D.; Hassan, I.; Looi, J.C.; Gaillard, F.; Velakoulis, D.; Walterfang, M. The neuropsychiatry of hyperkinetic movement disorders: Insights from neuroimaging into the neural circuit bases of dysfunction. Tremor Other Hyperkinet. Mov. 2013, 3, tre-03-175-4242-1. [Google Scholar]
- Ramnani, N. The primate cortico-cerebellar system: Anatomy and function. Nat. Rev. Neurosci. 2006, 7, 511–522. [Google Scholar] [CrossRef]
- Krack, P.; Hariz, M.I.; Baunez, C.; Guridi, J.; Obeso, J.A. Deep brain stimulation: From neurology to psychiatry? Trends Neurosci. 2010, 33, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Nakayama, K.; Kosaka, S.; Yamada, E.; Shimada, H.; Miki, T.; Inoue, Y. Diffusion tensor imaging of the cortico-ponto-cerebellar pathway in patients with adult-onset ataxic neurodegenerative disease. Neuroradiology 2008, 50, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Burk, K.; Abele, M.; Fetter, M.; Dichgans, J.; Skalej, M.; Laccone, F.; Didierjean, O.; Brice, A.; Klockgether, T. Autosomal dominant cerebellar ataxia type I clinical features and MRI in families with SCA1, SCA2 and SCA3. Brain 1996, 119, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Braga-Neto, P.; Dutra, L.A.; Pedroso, J.L.; Felicio, A.C.; Alessi, H.; Santos-Galduroz, R.F.; Bertolucci, P.H.; Castiglioni, M.L.; Bressan, R.A.; de Garrido, G.E.; et al. Cognitive deficits in Machado-Joseph disease correlate with hypo-perfusion of visual system areas. Cerebellum 2012, 11, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, D.J.; Saleem, K.S.; Baker, C.I. A new neural framework for visuospatial processing. Nat. Rev. Neurosci. 2011, 12, 217–230. [Google Scholar] [CrossRef] [PubMed]
- D’Abreu, A.; Franca, M.C., Jr.; Yasuda, C.L.; Campos, B.A.; Lopes-Cendes, I.; Cendes, F. Neocortical atrophy in machado-joseph disease: A longitudinal neuroimaging study. J. Neuroimaging 2012, 22, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Lopes, T.M.; D′Abreu, A.; Junior, M.C.F.; Yasuda, C.L.; Betting, L.E.; Samara, A.B.; Castellano, G.; Somazz, J.C.; Balthazar, M.L.F.; Lopes-Cendes, I.; et al. Widespread neuronal damage and cognitive dysfunction in spinocerebellar ataxia type 3. J. Neurol. 2013, 260, 2370–2379. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Huang, S.R.; Jao, C.W.; Soong, B.W.; Lirng, J.F.; Wu, H.M.; Wang, P.S. Impaired Efficiency and Resilience of Structural Network in Spinocerebellar Ataxia Type 3. Front. Neurosci. 2018, 12, 935. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Group | P | |||
|---|---|---|---|---|---|
| Controls (N = 48) | SCA3 Patients (N = 48) | ||||
| Mean | SD | Mean | SD | ||
| Age (years) | 48.14 | 12.06 | 48.13 | 11.75 | 0.607 a |
| Duration (years) | -- | -- | 8.89 | 6.43 | -- |
| Clinical features | |||||
| SARA | -- | -- | 14 | 8.10 | -- |
| MMSE | -- | -- | 28.5 | 1.61 | -- |
| Cerebral atrophy/cerebellar atrophy (observed via visual inspection) | -- | -- | 9/39 | -- | -- |
| Gender | |||||
| Female | N = 24 | 50% | N = 21 | 43.75% | 0.544 a |
| Male | N = 24 | 50% | N = 27 | 56.25% | 0.544 a |
| Region(L/R) | Controls | SCA3 | Region (L/R) | Controls | SCA3 |
|---|---|---|---|---|---|
| Cerebellar Cortex | Parietal Lobe | ||||
| Entire | 2.56 ± 0.02 | 2.53 ± 0.04 | Post-central gyrus(L) | 2.17 ± 0.05 | 2.10 ± 0.06 |
| Anterior lobe (L) | 2.17 ± 0.04 | 2.11 ± 0.07 | Superior parietal gyrus(L) | 2.10 ± 0.06 | 2.03 ± 0.06 |
| Anterior lobe (R) | 2.15 ± 0.04 | 2.03 ± 0.08 | Superior parietal gyrus(R) | 2.08 ± 0.06 | 2.04 ± 0.07 |
| Posterior lobe(L) | 2.47 ± 0.03 | 2.45 ± 0.04 | Inferior parietal gyrus (L) | 2.19 ± 0.07 | 2.09 ± 0.08 |
| Posterior lobe(R) | 2.48 ± 0.03 | 2.44 ± 0.04 | Supramarginal gyrus (L) | 2.11 ± 0.05 | 2.04 ± 0.06 |
| Vermis | 2.15 ± 0.05 | 2.12 ± 0.04 | Angular gyrus (L) | 2.12 ± 0.07 | 2.00 ± 0.09 |
| Frontal Lobe | Precuneus (L) | 2.21 ± 0.03 | 2.17 ± 0.05 | ||
| Precentral gyrus(L) | 2.15 ± 0.07 | 2.07 ± 0.07 | Precuneus (R) | 2.14 ± 0.04 | 2.10 ± 0.04 |
| Superior frontal gyrus (L) | 2.08 ± 0.03 | 2.05 ± 0.05 | Occipital Lobe | ||
| Superior frontal gyrus (R) | 2.13 ± 0.04 | 2.10 ± 0.06 | Calcarine fissure and surrounding cortex (L) | 2.25 ± 0.04 | 2.22 ± 0.04 |
| Middle frontal gyrus (L) | 2.28 ± 0.04 | 2.25 ± 0.04 | Cuneus (L) | 2.13 ± 0.04 | 2.11 ± 0.04 |
| Orbitofrontal cortex(superior-medial) (L) | 2.11 ± 0.04 | 2.08 ± 0.05 | Lingual gyrus (L) | 2.20 ± 0.04 | 2.17 ± 0.05 |
| Orbitofrontal cortex(superior-medial) (R) | 2.14 ± 0.04 | 2.10 ± 0.05 | Superior occipital gyrus (L) | 1.95 ± 0.06 | 1.89 ± 0.08 |
| Inferior frontal gyrus (opercular) (R) | 2.10 ± 0.05 | 2.07 ± 0.05 | Middle occipital gyrus (L) | 2.19 ± 0.05 | 2.12 ± 0.07 |
| Inferior frontal gyrus (triangular) (L) | 2.27 ± 0.04 | 2.23 ± 0.05 | Temporal lobe | ||
| Supplementary motor area (L) | 2.19 ± 0.05 | 2.14 ± 0.05 | Superior temporal gyrus (L) | 2.18 ± 0.05 | 2.12 ± 0.05 |
| Superior frontal gyrus (medial) (L) | 2.17 ± 0.04 | 2.12 ± 0.06 | Middle temporal gyrus (L) | 2.34 ± 0.03 | 2.30 ± 0.05 |
| Superior frontal gyrus (medial) (R) | 2.09 ± 0.05 | 2.06 ± 0.07 | Limbic | ||
| Paracentral lobule (L) | 2.04 ± 0.07 | 1.99 ± 0.08 | Posterior cingulate gyrus (L) | 1.99 ± 0.05 | 1.95 ± 0.05 |
| Paracentral lobule (R) | 1.98 ± 0.07 | 1.92 ± 0.08 | Parahippocampal gyrus (R) | 2.16 ± 0.03 | 2.14 ± 0.03 |
| Subcortical Regions | Subcortical Regions | ||||
| Amygdala (R) | 1.94 ± 0.04 | 1.97 ± 0.04 | Caudate nucleus (R) | 2.09 ± 0.04 | 2.04 ± 0.05 |
| Caudate nucleus (L) | 2.08 ± 0.05 | 2.05 ± 0.05 | Lenticular nucleus, putamen (L) | 2.11 ± 0.08 | 2.07 ± 0.05 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jao, C.-W.; Soong, B.-W.; Wang, T.-Y.; Wu, H.-M.; Lu, C.-F.; Wang, P.-S.; Wu, Y.-T. Intra- and Inter-Modular Connectivity Alterations in the Brain Structural Network of Spinocerebellar Ataxia Type 3. Entropy 2019, 21, 317. https://doi.org/10.3390/e21030317
Jao C-W, Soong B-W, Wang T-Y, Wu H-M, Lu C-F, Wang P-S, Wu Y-T. Intra- and Inter-Modular Connectivity Alterations in the Brain Structural Network of Spinocerebellar Ataxia Type 3. Entropy. 2019; 21(3):317. https://doi.org/10.3390/e21030317
Chicago/Turabian StyleJao, Chi-Wen, Bing-Wen Soong, Tzu-Yun Wang, Hsiu-Mei Wu, Chia-Feng Lu, Po-Shan Wang, and Yu-Te Wu. 2019. "Intra- and Inter-Modular Connectivity Alterations in the Brain Structural Network of Spinocerebellar Ataxia Type 3" Entropy 21, no. 3: 317. https://doi.org/10.3390/e21030317