

miRNAs as a Potential Biomarker in the COVID-19 Infection and Complications Course, Severity, and Outcome
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. miRNAs Role in COVID-19 Disease
3.2. miRNAs as a Modulator of Cell Protein ACE2
3.3. miRNAs as a Modulator of Cell Protein TMPRSS2
3.4. miRNAs against Cellular Mediator NRP1
3.5. miRNAs Role in Obesity
3.6. miRNAs Role in Pregnancy
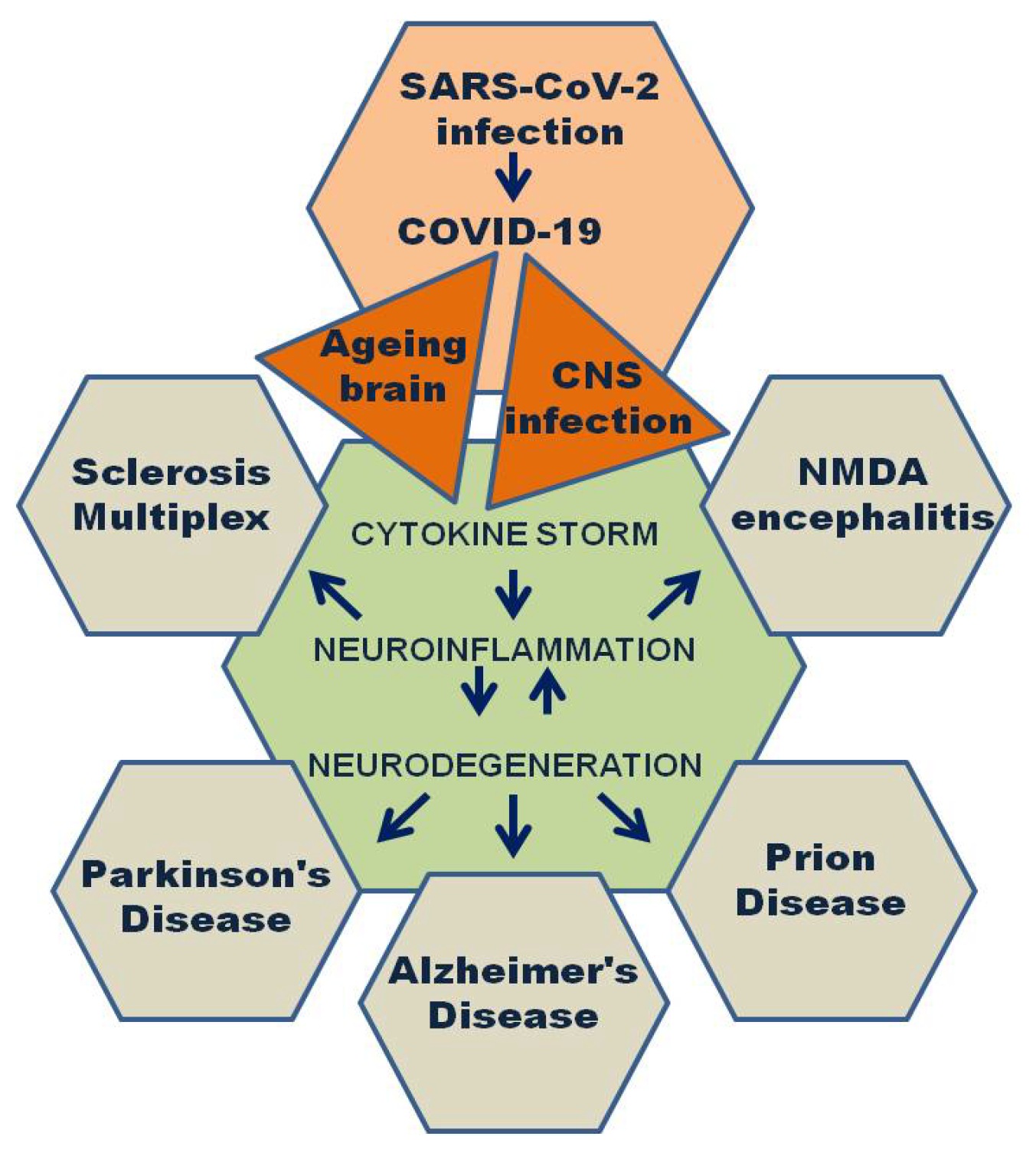
3.7. miRNAs Role in Neurological Diseases
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Bastard, P.; COVID Human Genetic Effort; Cobat, A.; Casanova, J.L. Human genetic and immunological deter-minants of critical COVID-19 pneumonia. Nature 2022, 603, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, A.; Ridolfo, A.L.; Pezzati, L.; Oreni, L.; Carrozzo, G.; Beltrami, M.; Poloni, A.; Caloni, B.; Lazzarin, S.; Colombo, M.; et al. Mortality rates among COVID-19 patients hospitalised during the first three waves of the epidemic in Milan, Italy: A prospective observational study. PLoS ONE 2022, 17, e0263548. [Google Scholar] [CrossRef]
- Ozonoff, A.; Schaenman, J.; Jayavelu, N.D.; Milliren, C.E.; Calfee, C.S.; Cairns, C.B.; Kraft, M.; Baden, L.R.; Shaw, A.C.; Krammer, F.; et al. Phenotypes of disease severity in a cohort of hospitalized COVID-19 patients: Results from the IMPACC study. Ebiomedicine 2022, 83, 104208. [Google Scholar] [CrossRef] [PubMed]
- Zsichla, L.; Müller, V. Risk Factors of Severe COVID-19: A Review of Host, Viral and Environmental Factors. Viruses 2023, 15, 175. [Google Scholar] [CrossRef]
- Niemi, M.E.K.; Daly, M.J.; Ganna, A. The human genetic epidemiology of COVID-19. Nat. Rev. Genet. 2022, 23, 533–546. [Google Scholar] [CrossRef]
- van der Made, C.I.; Netea, M.G.; van der Veerdonk, F.L.; Hoischen, A. Clinical implications of host genetic variation and susceptibility to severe or critical COVID-19. Genome Med. 2022, 14, 96. [Google Scholar] [CrossRef]
- Quintana-Murci, L. Human Immunology through the Lens of Evolutionary Genetics. Cell 2019, 177, 184–199. [Google Scholar] [CrossRef] [Green Version]
- Shelton, J.F.; Shastri, A.J.; Ye, C.; Weldon, C.H.; Filshtein-Sonmez, T.; Coker, D.; Symons, A.; Esparza-Gordillo, J.; 23andMe COVID-19 Team; Aslibekyan, S. Trans-ancestry analysis reveals genetic and nongenetic associations with COVID-19 suscep-tibility and severity. Nat. Genet. 2021, 53, 801–808. [Google Scholar] [CrossRef]
- Severe Covid-19 GWAS Group; Ellinghaus, D.; Degenhardt, F.; Bujanda, L.; Buti, M.; Albillos, A.; Invernizzi, P.; Fernández, J.; Prati, D.; Baselli, G.; et al. Genomewide Association Study of Severe Covid-19 with Respiratory Failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar]
- Tangye, S.G.; Bucciol, G.; Meyts, I. Mechanisms underlying host defense and disease pathology in response to severe acute respiratory syndrome (SARS)-CoV2 infection: Insights from inborn errors of immunity. Curr. Opin. Allergy Clin. Immunol. 2021, 21, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Niknam, Z.; Jafari, A.; Golchin, A.; Danesh Pouya, F.; Nemati, M.; Rezaei-Tavirani, M.; Rasmi, Y. Potential therapeutic options for COVID-19: An update on current evidence. Eur. J. Med. Res. 2022, 27, 6. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.E.; Martin, M.M.; Feldman, D.S.; Terry, A.V., Jr.; Nuovo, G.J.; Elton, T.S. Experimental validation of miRNA targets. Methods 2008, 44, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoletti, A.D.S.; Visacri, M.B.; da Ronda, C.R.d.S.C.; Vasconcelos, P.E.D.N.S.; Quintanilha, J.C.F.; de Souza, R.N.; Ventura, D.D.S.; Eguti, A.; Silva, L.F.d.S.; Junior, M.W.P.; et al. Differentially expressed plasmatic microRNAs in Brazilian patients with Coronavirus disease 2019 (COVID-19): Preliminary results. Mol. Biol. Rep. 2022, 49, 6931–6943. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Cullen, B.R. Viruses, microRNAs, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.Y.; Chen, Y.H.; Liu, P.J.; Hu, W.C.; Lu, K.C.; Tsai, K.W. The emerging role of miRNAs in the pathogenesis of COVID-19: Protective effects of nutraceutical polyphenolic compounds against SARS-CoV-2 infection. Int. J. Med. Sci. 2022, 19, 1340–1356. [Google Scholar] [CrossRef]
- Paul, S.; Vázquez, L.A.B.; Reyes-Pérez, P.R.; Estrada-Meza, C.; Alburquerque, R.A.A.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Chakraborty, S.; Srivastava, A. The role of microRNAs in solving COVID-19 puzzle from infection to therapeutics: A mini-review. Virus Res. 2021, 308, 198631. [Google Scholar] [CrossRef]
- Giannella, A.; Riccetti, S.; Sinigaglia, A.; Piubelli, C.; Razzaboni, E.; Di Battista, P.; Agostini, M.; Molin, E.D.; Manganelli, R.; Gobbi, F.; et al. Circulating microRNA signatures associated with disease severity and outcome in COVID-19 patients. Front. Immunol. 2022, 13, 4409. [Google Scholar] [CrossRef]
- Fernández-Pato, A.; Virseda-Berdices, A.; Resino, S.; Ryan, P.; Martínez-González, O.; Pérez-García, F.; Martin-Vicente, M.; Valle-Millares, D.; Brochado-Kith, O.; Blancas, R.; et al. Plasma miRNA profile at COVID-19 onset predicts severity status and mortality. Emerg. Microbes Infect. 2022, 11, 676–688. [Google Scholar] [CrossRef]
- Marchi, R.; Sugita, B.; Centa, A.; Fonseca, A.S.; Bortoletto, S.; Fiorentin, K.; Ferreira, S.; Cavalli, L.R. The role of microRNAs in modulating SARS-CoV-2 infection in human cells: A systematic review. Infect. Genet. Evol. 2021, 91, 104832. [Google Scholar] [CrossRef]
- Parray, A.; Mir, F.A.; Doudin, A.; Iskandarani, A.; Danjuma, I.M.M.; Kuni, R.A.T.; Abdelmajid, A.; Abdelhafez, I.; Arif, R.; Mulhim, M.; et al. SnoRNAs and miRNAs Networks Underlying COVID-19 Disease Severity. Vaccines 2021, 9, 1056. [Google Scholar] [CrossRef] [PubMed]
- Saçar Demirci, M.D.; Adan, A. Computational analysis of microRNA-mediated interactions in SARS-CoV-2 infection. PeerJ 2020, 8, e9369. [Google Scholar] [CrossRef] [PubMed]
- Khokhar, M.; Tomo, S.; Purohit, P. MicroRNAs based regulation of cytokine regulating immune expressed genes and their transcription factors in COVID-19. Meta Gene 2021, 31, 100990. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.S.; Rocha, L.P.C.; Moreira, P.R. Genetic and epigenetic control of ACE2 expression and its possible role in COVID-19. Cell Biochem. Funct. 2021, 39, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, X.; Li, L.; Li, J. Differential microRNA expression in the peripheral blood from human patients with COVID-19. J. Clin. Lab. Anal. 2020, 34, e23590. [Google Scholar] [CrossRef]
- Donyavi, T.; Bokharaei-Salim, F.; Baghi, H.B.; Khanaliha, K.; Janat-Makan, M.A.; Karimi, B.; Nahand, J.S.; Mirzaei, H.; Khatami, A.; Garshasbi, S.; et al. Acute and post-acute phase of COVID-19: Analyzing expression patterns of miRNA-29a-3p, 146a-3p, 155–5p, and let-7b-3p in PBMC. Int. Immunopharmacol. 2021, 97, 107641. [Google Scholar] [CrossRef]
- Calderon-Dominguez, M.; Trejo-Gutierrez, E.; González-Rovira, A.; Beltrán-Camacho, L.; Rojas-Torres, M.; Eslava-Alcón, S.; Sanchez-Morillo, D.; Calderon-Dominguez, J.; Martinez-Nicolás, M.P.; Gonzalez-Beitia, E.; et al. Serum microRNAs target-ing ACE2 and RAB14 genes distinguish asymptomatic from critical COVID-19 patients. Mol. Ther. Nucleic Acids. 2022, 29, 76–87. [Google Scholar] [CrossRef]
- Garg, A.; Seeliger, B.; Derda, A.A.; Xiao, K.; Gietz, A.; Scherf, K.; Sonnenschein, K.; Pink, I.; Hoeper, M.M.; Welte, T.; et al. Circulating cardiovascular microRNAs in critically ill COVID-19 patients. Eur. J. Heart Fail. 2021, 23, 468–475. [Google Scholar] [CrossRef]
- Gaytán-Pacheco, N.; Ibáñez-Salazar, A.; Herrera-Van Oostdam, A.S.; Oropeza-Valdez, J.J.; Magaña-Aquino, M.; Adrián López, J.; Monárrez-Espino, J.; López-Hernández, Y. miR-146a, miR-221, and miR-155 are Involved in Inflammatory Immune Re-sponse in Severe COVID-19 Patients. Diagnostics 2022, 13, 133. [Google Scholar] [CrossRef]
- McDonald, J.T.; Enguita, F.J.; Taylor, D.; Griffin, R.J.; Priebe, W.; Emmett, M.R.; Sajadi, M.M.; Harris, A.D.; Clement, J.; Dybas, J.M.; et al. Role of miR-2392 in driving SARS-CoV-2 infection. Cell Rep. 2021, 37, 109839. [Google Scholar] [CrossRef]
- Benhamou, R.I.; Choudhary, S.; Lekah, E.; Tong, Y.; Disney, M.D. Bioinformatic Searching for Optimal RNA Targets of Dimeric Compounds Informs Design of a MicroRNA-27a Inhibitor. ACS Chem. Biol. 2021, 17, 5–10. [Google Scholar] [CrossRef]
- Bugnon, L.A.; Raad, J.; Merino, G.A.; Yones, C.; Ariel, F.; Milone, D.H.; Stegmayer, G. Deep Learning for the discovery of new pre-miRNAs: Helping the fight against COVID-19. Mach. Learn. Appl. 2021, 6, 100150. [Google Scholar] [CrossRef]
- Lodhi, N.; Singh, R.; Rajput, S.P.; Saquib, Q. SARS-CoV-2: Understanding the Transcriptional Regulation of ACE2 and TMPRSS2 and the Role of Single Nucleotide Polymorphism (SNP) at Codon 72 of p53 in the Innate Immune Response against Virus Infection. Int. J. Mol. Sci. 2021, 22, 8660. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.O.; Jakhar, K.; Nema, V.; Krishnaraj, A.; Choudhari, R. Effect of miRNAs, Proinflammatory Cytokines and ACE2 in COVID-19 Pathophysiology. Coronaviruses 2021, 2, e020721192021. [Google Scholar] [CrossRef]
- Ahmed, J.Q.; Maulud, S.Q.; Dhawan, M.; Choudhary, O.P.; Jalal, P.J.; Ali, R.K.; Tayib, G.A.; Hasan, D.A. MicroRNAs in the development of potential therapeutic targets against COVID-19: A narrative review. J. Infect. Public Health 2022, 15, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Choudhari, R.; Nema, V.; Khan, A.A. ACE2 and TMPRSS2 polymorphisms in various diseases with special reference to its impact on COVID-19 disease. Microb. Pathog. 2020, 150, 104621. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Chatterjee, S.; Xiao, K.; Riedel, I.; Wang, Y.; Foo, R.; Bär, C.; Thum, T. MicroRNAs targeting the SARS-CoV-2 entry receptor ACE2 in cardiomyocytes. J. Mol. Cell. Cardiol. 2020, 148, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Nersisyan, S.; Shkurnikov, M.; Turchinovich, A.; Knyazev, E.; Tonevitsky, A. Integrative analysis of miRNA and mRNA sequencing data reveals potential regulatory mechanisms of ACE2 and TMPRSS2. PLoS ONE 2020, 15, e0235987. [Google Scholar] [CrossRef]
- Eyileten, C.; Wicik, Z.; Simões, S.N.; Martins, D.C., Jr.; Klos, K.; Wlodarczyk, W.; Assinger, A.; Soldacki, D.; Chcialowski, A.; Siller-Matula, J.M.; et al. Thrombosis-related circulating miR-16-5p is associated with disease severity in patients hospitalised for COVID-19. RNA Biol. 2022, 19, 963–979. [Google Scholar] [CrossRef]
- Senapati, S.; Banerjee, P.; Bhagavatula, S.; Kushwaha, P.P.; Kumar, S. Contributions of human ACE2 and TMPRSS2 in deter-mining host-pathogen interaction of COVID-19. J. Genet. 2021, 100, 12. [Google Scholar] [CrossRef]
- Matarese, A.; Gambardella, J.; Sardu, C.; Santulli, G. miR-98 Regulates TMPRSS2 Expression in Human Endothelial Cells: Key Implications for COVID-19. Biomedicines 2020, 8, 462. [Google Scholar] [CrossRef]
- Katopodis, P.; Randeva, H.S.; Spandidos, D.A.; Saravi, S.; Kyrou, I.; Karteris, E. Host cell entry mediators implicated in the cellular tropism of SARS CoV 2, the pathophysiology of COVID 19 and the identification of microRNAs that can modulate the expression of these mediators (Review). Int. J. Mol. Med. 2022, 49, 20. [Google Scholar] [CrossRef]
- Naidoo, N.; Moodley, J.; Khaliq, O.P.; Naicker, T. Neuropilin-1 in the pathogenesis of preeclampsia, HIV-1, and SARS-CoV-2 infection: A review. Virus Res. 2022, 319, 198880. [Google Scholar] [CrossRef] [PubMed]
- Mone, P.; Gambardella, J.; Wang, X.; Jankauskas, S.S.; Matarese, A.; Santulli, G. miR-24 targets SARS-CoV-2 co-factor Neuro-pilin-1 in human brain microvascular endothelial cells: Insights for COVID-19 neurological manifestations. Res. Sq. 2021, rs.3.rs-192099. [Google Scholar] [CrossRef]
- Bellae Papannarao, J.; Schwenke, D.O.; Manning, P.; Katare, R. Upregulated miR-200c is associated with downregulation of the functional receptor for severe acute respiratory syndrome coronavirus 2 ACE2 in individuals with obesity. Int. J. Obes. (Lond.) 2022, 46, 238–241. [Google Scholar] [CrossRef]
- Saccon, T.D.; Mousovich-Neto, F.; Ludwig, R.G.; Carregari, V.C.; Dos Anjos Souza, A.B.; Dos Passos, A.S.C.; Martini, M.C.; Barbosa, P.P.; de Souza, G.F.; Muraro, S.P.; et al. SARS-CoV-2 infects adipose tissue in a fat depot- and viral lineage-dependent manner. Nat. Commun. 2022, 13, 5722. [Google Scholar] [CrossRef]
- Sudhakar, M.; Winfred, S.B.; Meiyazhagan, G.; Venkatachalam, D.P. Mechanisms contributing to adverse outcomes of COVID-19 in obesity. Mol. Cell. Biochem. 2022, 477, 1155–1193. [Google Scholar] [CrossRef] [PubMed]
- Elemam, N.M.; Hasswan, H.; Aljaibeji, H.; Sharif-Askari, N.S.; Halwani, R.; Taneera, J.; Sulaiman, N. Profiling Levels of Serum microRNAs and Soluble ACE2 in COVID-19 Patients. Life 2022, 12, 575. [Google Scholar] [CrossRef]
- Tseng, Y.-H. Adipose tissue in communication: Within and without. Nat. Rev. Endocrinol. 2023, 19, 70–71. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, H.; Li, X.; Zhao, Y. Current understanding of the role of microRNAs from adipose-derived extracellular vesicles in obesity. Biochem. Soc. Trans. 2021, 50, 447–457. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Zhang, Z.; Zhang, X.; Zhu, Y.; Zhang, C.; Bi, Y. Extracellular vesicles mediate the communication of adipose tissue with brain and promote cognitive impairment associated with insulin resistance. Cell Metab. 2022, 34, 1264–1279.e8. [Google Scholar] [CrossRef] [PubMed]
- Sierra, A.C.; Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez, J.A. Diet, Gut Microbiota, and Obesity: Links with Host Genetics and Epigenetics and Potential Applications. Adv. Nutr. 2019, 10, S17–S30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, C.R.; Wang, G.; Lee, K.Y. Altered adipose tissue and adipocyte function in the pathogenesis of metabolic syndrome. J. Clin. Investig. 2019, 129, 3990–4000. [Google Scholar] [CrossRef] [PubMed]
- Vettori, A.; Pompucci, G.; Paolini, B.; Del Ciondolo, I.; Bressan, S.; Dundar, M.; Kenanoğlu, S.; Unfer, V.; Bertelli, M.; Geneob Project. Genetic background, nutrition and obesity: A review. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1751–1761. [Google Scholar]
- Roganović, J. Downregulation of microRNA-146a in diabetes, obesity and hypertension may contribute to severe COVID-19. Med. Hypotheses 2020, 146, 110448. [Google Scholar] [CrossRef]
- Mormile, R. Obesity and Severe COVID-19 in the Young: Is Downregulation of miR-126 a Piece of the SARS-COV2 Patho-genicity Puzzle? Arch. Med. Res. 2022, 53, 228–230. [Google Scholar] [CrossRef]
- Dashraath, P.; Wong, J.L.J.; Lim, M.X.K.; Lim, L.M.; Li, S.; Biswas, A.; Choolani, M.; Mattar, C.; Su, L.L. Coronavirus disease 2019 (COVID-19) pandemic and pregnancy. Am. J. Obstet. Gynecol. 2020, 222, 521–531. [Google Scholar] [CrossRef]
- Royal College of Obstetricians and Gynaecologists. Coronavirus (COVID-19) Infection in Pregnancy: Information for Healthcare Professionals Version 13; Royal College of Obstetricians and Gynaecologists: London, UK, 2021. [Google Scholar]
- Panda, M.; Kalita, E.; Singh, S.; Kumar, K.; Rao, A.; Prajapati, V.K. MiRNA-SARS-CoV-2 dialogue and prospective an-ti-COVID-19 therapies. Life Sci. 2022, 305, 120761. [Google Scholar] [CrossRef]
- Mourad, M.; Jacob, T.; Sadovsky, E.; Bejerano, S.; Simone, G.S.; Bagalkot, T.R.; Zucker, J.; Yin, M.T.; Chang, J.Y.; Liu, L.; et al. Placental response to maternal SARS-CoV-2 infection. Sci. Rep. 2021, 11, 14390. [Google Scholar] [CrossRef]
- Gonlepa, M.K.; Rufai, O.H.; Ofuonye, C.G.; Sebaka, L. Coronavirus-linked pregnancy complications: A comparative study. Egypt. J. Med. Hum. Genet. 2022, 23, 13. [Google Scholar] [CrossRef]
- Pierce-Williams, R.A.; Burd, J.; Felder, L.; Khoury, R.; Bernstein, P.S.; Avila, K.; Penfield, C.A.; Roman, A.S.; DeBolt, C.A.; Stone, J.L.; et al. Clinical course of severe and critical coronavirus disease 2019 in hospitalized pregnancies: A United States cohort study. Am. J. Obstet. Gynecol. MFM 2020, 2, 100134. [Google Scholar] [CrossRef] [PubMed]
- Metz, T.D.; Clifton, R.G.; Hughes, B.L.; Sandoval, G.; Saade, G.R.; Grobman, W.A.; Manuck, T.A.; Miodovnik, M.; Sowles, A.; Clark, K.; et al. Disease Severity and Perinatal Outcomes of Pregnant Patients With Coronavirus Disease 2019 (COVID-19). Obstet. Gynecol. 2021, 137, 571–580. [Google Scholar] [CrossRef]
- Tan, J.S.; Liu, N.N.; Guo, T.T.; Hu, S.; Hua, L. Genetic predisposition to COVID-19 may increase the risk of hypertension dis-orders in pregnancy: A two-sample Mendelian randomization study. Pregnancy Hypertens. 2021, 26, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Gayen Nee’ Betal, S.; Urday, P.; Al-Kouatly, H.B.; Solarin, K.; Chan, J.S.Y.; Addya, S.; Boelig, R.C.; Aghai, Z.H. COVID-19 Infection During Pregnancy Induces Differential Gene Expression in Human Cord Blood Cells From Term Neonates. Front. Pediatr. 2022, 10, 834771. [Google Scholar] [CrossRef]
- Saulle, I.; Garziano, M.; Fenizia, C.; Cappelletti, G.; Parisi, F.; Clerici, M.; Cetin, I.; Savasi, V.; Biasin, M. MiRNA Profiling in Plasma and Placenta of SARS-CoV-2-Infected Pregnant Women. Cells 2021, 10, 1788. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, J.; Wang, H.; Shi, J.; Wu, K.; Liu, S.; Liu, Y.; Wu, J. HCV-Induced miR-21 Contributes to Evasion of Host Immune System by Targeting MyD88 and IRAK1. PLOS Pathog. 2013, 9, e1003248. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, A.; Vega-Guedes, B.; Brito-Casillas, Y.; Wägner, A.M. Diabetes in Pregnancy and MicroRNAs: Promises and Limitations in Their Clinical Application. Non-Coding RNA 2018, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Cirkovic, A.; Stanisavljevic, D.; Milin-Lazovic, J.; Rajovic, N.; Pavlovic, V.; Milicevic, O.; Savic, M.; Peric, J.K.; Aleksic, N.; Milic, N.; et al. Preeclamptic Women Have Disrupted Placental microRNA Expression at the Time of Preeclampsia Diagnosis: Meta-Analysis. Front. Bioeng. Biotechnol. 2021, 9, 782845. [Google Scholar] [CrossRef]
- Inchley, C.S.; Sonerud, T.; O Fjærli, H.; Nakstad, B. Nasal mucosal microRNA expression in children with respiratory syncytial virus infection. BMC Infect. Dis. 2015, 15, 150. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, C.; Dong, X.; Gou, W. MicroRNA-155 inhibits migration of trophoblast cells and contributes to the pathogenesis of severe preeclampsia by regulating endothelial nitric oxide synthase. Mol. Med. Rep. 2014, 10, 550–554. [Google Scholar] [CrossRef] [Green Version]
- Abel, T.; Moodley, J.; Naicker, T. The Involvement of MicroRNAs in SARS-CoV-2 Infection Comorbid with HIV-Associated Preeclampsia. Curr. Hypertens. Rep. 2021, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Li, Y.; Xu, Y. Decreased placental miR-126 expression and vascular endothelial growth factor levels in patients with pre-eclampsia. J. Int. Med. Res. 2014, 42, 1243–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardar, R.; Satish, D.; Birla, S.; Gupta, D. Integrative analyses of SARS-CoV-2 genomes from different geographical locations reveal unique features potentially consequential to host-virus interaction, pathogenesis and clues for novel therapies. Heliyon 2020, 6, e04658. [Google Scholar] [CrossRef] [PubMed]
- Abbas-Hanif, A.; Rezai, H.; Ahmed, S.F.; Ahmed, A. The impact of COVID-19 on pregnancy and therapeutic drug development. Br. J. Pharmacol. 2021, 179, 2108–2120. [Google Scholar] [CrossRef]
- Lin, Y.-P.; Hsieh, Y.-S.; Cheng, M.-H.; Shen, C.-F.; Shen, C.-J.; Cheng, C.-M. Using MicroRNA Arrays as a Tool to Evaluate COVID-19 Vaccine Efficacy. Vaccines 2022, 10, 1681. [Google Scholar] [CrossRef]
- Prosperini, L.; Tortorella, C.; Haggiag, S.; Ruggieri, S.; Galgani, S.; Gasperini, C. Determinants of COVID-19-related lethality in multiple sclerosis: A meta-regression of observational studies. J. Neurol. 2022, 269, 2275–2285. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Chen, C.; Dong, X.-P. Impact of COVID-19 Pandemic on Patients With Neurodegenerative Diseases. Front. Aging Neurosci. 2021, 13, 664965. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yang, H.; Hou, C.; Chen, W.; Zhang, H.; Ying, Z.; Hu, Y.; Sun, Y.; Qu, Y.; Feychting, M.; et al. COVID-19 related outcomes among individuals with neurodegenerative diseases: A cohort analysis in the UK biobank. BMC Neurol. 2022, 22, 15. [Google Scholar] [CrossRef]
- De Marchi, F.; Gallo, C.; Sarnelli, M.F.; De Marchi, I.; Saraceno, M.; Cantello, R.; Mazzini, L. Accelerated Early Progression of Amyotrophic Lateral Sclerosis over the COVID-19 Pandemic. Brain Sci. 2021, 11, 1291. [Google Scholar] [CrossRef]
- Musson, L.S.; Collins, A.; Opie-Martin, S.; Bredin, A.; Hobson, E.V.; Barkhouse, E.; Coulson, M.C.; Stavroulakis, T.; Gould, R.L.; Al-Chalabi, A.; et al. Impact of the covid-19 pandemic on amyotrophic lateral sclerosis care in the UK. Amyotroph. Lateral Scler. Front. Degener. 2022, 24, 91–99. [Google Scholar] [CrossRef]
- Danics, K.; Forrest, S.L.; Kapas, I.; Erber, I.; Schmid, S.; Törő, K.; Majtenyi, K.; Kovacs, G.G. Neurodegenerative proteinopathies associated with neuroinfections. J. Neural Transm. 2021, 128, 1551–1566. [Google Scholar] [CrossRef]
- Vigasova, D.; Nemergut, M.; Liskova, B.; Damborsky, J. Multi-pathogen infections and Alzheimer’s disease. Microb. Cell Factories 2021, 20, 25. [Google Scholar] [CrossRef] [PubMed]
- Piekut, T.; Hurła, M.; Banaszek, N.; Szejn, P.; Dorszewska, J.; Kozubski, W.; Prendecki, M. Infectious agents and Alzheimer’s disease. J. Integr. Neurosci. 2022, 21, 73. [Google Scholar] [CrossRef]
- Burwinkel, M.; Riemer, C.; Schwarz, A.; Schultz, J.; Neidhold, S.; Bamme, T.; Baier, M. Role of cytokines and chemokines in prion infections of the central nervous system. Int. J. Dev. Neurosci. 2004, 22, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Ciolac, D.; Racila, R.; Duarte, C.; Vasilieva, M.; Manea, D.; Gorincioi, N.; Condrea, A.; Crivorucica, I.; Zota, E.; Efremova, D.; et al. Clinical and Radiological Deterioration in a Case of Creutzfeldt–Jakob Disease following SARS-CoV-2 Infection: Hints to Accelerated Age-Dependent Neurodegeneration. Biomedicines 2021, 9, 1730. [Google Scholar] [CrossRef] [PubMed]
- Choe, K.; Park, H.Y.; Ikram, M.; Lee, H.J.; Park, T.J.; Ullah, R.; Kim, M.O. Systematic Review of the Common Pathophysiological Mechanisms in COVID-19 and Neurodegeneration: The Role of Bioactive Compounds and Natural Antioxidants. Cells 2022, 11, 1298. [Google Scholar] [CrossRef]
- Lingor, P.; Demleitner, A.F.; Wolff, A.W.; Feneberg, E. SARS-CoV-2 and neurodegenerative diseases: What we know and what we don’t. J. Neural. Transm (Vienna) 2022, 129, 1155–1167. [Google Scholar] [CrossRef]
- Baazaoui, N.; Iqbal, K. COVID-19 and Neurodegenerative Diseases: Prion-Like Spread and Long-Term Consequences. J. Alzheimer’s Dis. 2022, 88, 399–416. [Google Scholar] [CrossRef]
- Rossi, E.; Mutti, L.; Morrione, A.; Giordano, A. Neuro–Immune Interactions in Severe COVID-19 Infection. Pathogens 2022, 11, 1256. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, W.; Lukiw, W. Ubiquity of the SARS-CoV-2 receptor ACE2 and upregulation in limbic regions of Alzheimer’s disease brain. Folia Neuropathol. 2021, 59, 232–238. [Google Scholar] [CrossRef]
- Piras, M.; Cau, F.; Manchia, M.; Paribello, P.; Saba, L.; Suri, J.S.; Gerosa, C. Strong ACE-2 expression in the choroidal vessels: Do high choroid plexuses serve as a gateway for SARS-CoV-2 infection on the human brain? Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 3025–3029. [Google Scholar] [PubMed]
- Shahzad, S.; Willcox, M. The Possible Role of Prion-Like Viral Protein Domains on the Emergence of Novel Viruses as SARS-CoV-2. J. Mol. Evol. 2022, 90, 227–230. [Google Scholar] [CrossRef]
- Young, M.J.; O’Hare, M.; Matiello, M.; Schmahmann, J.D. Creutzfeldt-Jakob disease in a man with COVID-19: SARS-CoV-2-accelerated neurodegeneration? Brain Behav. Immun. 2020, 89, 601–603. [Google Scholar] [CrossRef]
- Kuvandık, A.; Özcan, E.; Karaduman, S.; Sungurtekin, H. Creutzfeldt-Jakob Disease After the Coronavirus Disease-2019 Vaccination. Turk. J. Intensiv. Care 2022, 20, 61–64. [Google Scholar] [CrossRef]
- Bernardini, A.; Gigli, G.L.; Janes, F.; Pellitteri, G.; Ciardi, C.; Fabris, M.; Valente, M. Creutzfeldt-Jakob disease after COVID-19: Infection-induced prion protein misfolding? A case report. Prion 2022, 16, 78–83. [Google Scholar] [CrossRef]
- Olivo, S.; Furlanis, G.; Stella, A.B.; Fabris, M.; Milanic, R.; Zanusso, G.; Manganotti, P. Rapidly evolving Creutzfeldt–Jakob disease in COVID-19: From early status epilepticus to fatal outcome. Acta Neurol. Belg. 2022, 1–4. [Google Scholar] [CrossRef]
- Lee, H.; Jeon, J.H.; Choi, H.M.; Koh, S.-H.M.; Lee, K.-Y.M.; Lee, Y.J.M.; Kwon, H.S.M. Anti-N-methyl-D-aspartate receptor encephalitis after coronavirus disease 2019: A case report and literature review. Medicine 2022, 101, e30464. [Google Scholar] [CrossRef] [PubMed]
- Tayyebi, G.; Malakouti, S.K.; Shariati, B.; Kamalzadeh, L. COVID-19-associated encephalitis or Creutzfeldt–Jakob disease: A case report. Neurodegener. Dis. Manag. 2022, 12, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Haroun, R.A.; Osman, W.H.; Amin, R.E.; Hassan, A.K.; Abo-Shanab, W.S.; Eessa, A.M. Circulating plasma miR-155 is a potential biomarker for the detection of SARS-CoV-2 infection. Pathology 2022, 54, 104–110. [Google Scholar] [CrossRef]
- Song, J.; Lee, J.E. miR-155 is involved in Alzheimer’s disease by regulating T lymphocyte function. Front. Aging Neurosci. 2015, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Tili, E.; Mezache, L.; Michaille, J.J.; Amann, V.; Williams, J.; Vandiver, P.; Quinonez, M.; Fadda, P.; Mikhail, A.; Nuovo, G. microRNA 155 up regulation in the CNS is strongly correlated to Down’s syndrome dementia. Ann. Diagn. Pathol. 2018, 34, 103–109. [Google Scholar] [CrossRef]
- Maciak, K.; Dziedzic, A.; Miller, E.; Saluk-Bijak, J. miR-155 as an Important Regulator of Multiple Sclerosis Pathogenesis: A Review. Int. J. Mol. Sci. 2021, 22, 4332. [Google Scholar] [CrossRef]
- Zingale, V.D.; Gugliandolo, A.; Mazzon, E. MiR-155: An Important Regulator of Neuroinflammation. Int. J. Mol. Sci. 2021, 23, 90. [Google Scholar] [CrossRef] [PubMed]
- Kenny, A.; McArdle, H.; Calero, M.; Rabano, A.; Madden, S.F.; Adamson, K.; Forster, R.; Spain, E.; Prehn, J.H.; Henshall, D.C.; et al. Elevated Plasma microRNA-206 Levels Predict Cognitive Decline and Progression to Dementia from Mild Cognitive Impairment. Biomolecules 2019, 9, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Liu, Y.; Huang, J.; Gao, L.; Wu, Z.; Wang, L.; Fan, L. Let 7b 5p promotes cell apoptosis in Parkinson’s disease by targeting HMGA2. Mol. Med. Rep. 2021, 24, 820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, X.; Zhao, S.; Gong, Z.; Liu, P.; Guan, W.; He, X.; Wang, T.; Peng, T.; Teng, J.; et al. The Expression and Significance of the Plasma Let-7 Family in Anti-N-methyl-d-aspartate Receptor Encephalitis. J. Mol. Neurosci. 2015, 56, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. COVID−19, Anti-NMDA Receptor Encephalitis and MicroRNA. Front. Immunol. 2022, 13, 1184. [Google Scholar] [CrossRef]
- Bai, X.; Bian, Z. MicroRNA-21 Is a Versatile Regulator and Potential Treatment Target in Central Nervous System Disorders. Front. Mol. Neurosci. 2022, 15, 5. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.-H.; Hua, P.; Chen, Y.; Li, L.-T.; Yu, C.-Y.; Yan, L.; Zhang, H.; He, Y.; Zheng, H.; Chen, H.; et al. Identification of microRNAs for the early diagnosis of Parkinson’s disease and multiple system atrophy. J. Integr. Neurosci. 2020, 19, 429–436. [Google Scholar] [CrossRef]
- Keikha, R.; Hashemi-Shahri, S.M.; Jebali, A. The relative expression of miR-31, miR-29, miR-126, and miR-17 and their mRNA targets in the serum of COVID-19 patients with different grades during hospitalization. Eur. J. Med. Res. 2021, 26, 75. [Google Scholar] [CrossRef]
- Farr, R.J.; Rootes, C.L.; Rowntree, L.C.; Nguyen, T.H.O.; Hensen, L.; Kedzierski, L.; Cheng, A.C.; Kedzierska, K.; Au, G.G.; Marsh, G.A.; et al. Altered microRNA expression in COVID-19 patients enables identification of SARS-CoV-2 infection. PLoS Pathog. 2021, 17, e1009759. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, C.-F.; Wang, A.-H.; Lin, Q.-F. MiR-16 regulates cell death in Alzheimer’s disease by targeting amyloid precursor protein. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4020–4027. [Google Scholar]
- de Gonzalo-Calvo, D.; Benítez, I.D.; Pinilla, L.; Carratalá, A.; Moncusí-Moix, A.; Gort-Paniello, C.; Molinero, M.; González, J.; Torres, G.; Bernal, M.; et al. Circulating microRNA profiles predict the severity of COVID-19 in hospitalized patients. Transl. Res. 2021, 236, 147–159. [Google Scholar] [CrossRef]
- Burak, K.; Lamoureux, L.; Boese, A.; Majer, A.; Saba, R.; Niu, Y.; Frost, K.; Booth, S.A. MicroRNA-16 targets mRNA involved in neurite extension and branching in hippocampal neurons during presymptomatic prion disease. Neurobiol. Dis. 2018, 112, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Galván-Román, J.M.; Lancho-Sánchez, Á.; Luquero-Bueno, S.; Vega-Piris, L.; Curbelo, J.; Manzaneque-Pradales, M.; Gómez, M.; De La Fuente, H.; Ortega-Gómez, M.; Aspa, J. Usefulness of circulating microRNAs miR-146a and miR-16-5p as prognostic biomarkers in community-acquired pneumonia. PLoS ONE 2020, 15, e0240926. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Reddy, P.H. Are circulating microRNAs peripheral biomarkers for Alzheimer’s disease? Biochim. Biophys. Acta 2016, 1862, 1617–1627. [Google Scholar] [CrossRef]
- Swarbrick, S.; Wragg, N.; Ghosh, S.; Stolzing, A. Systematic Review of miRNA as Biomarkers in Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 6156–6167. [Google Scholar] [CrossRef] [Green Version]
- Lukiw, W.J. microRNA-146a Signaling in Alzheimer’s Disease (AD) and Prion Disease (PrD) Front. Neurol. 2020, 11, 462. [Google Scholar]
- Pogue, A.I.; Lukiw, W.J. microRNA-146a-5p, Neurotropic Viral Infection and Prion Disease (PrD). Int. J. Mol. Sci. 2021, 22, 9198. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zou, T.; Zhang, M.; Fan, W.; Zhang, T.; Jiang, Y.; Cai, Y.; Chen, F.; Chen, X.; Sun, Y.; et al. MicroRNA-146a switches microglial phenotypes to resist the pathological processes and cognitive degradation of Alzheimer’s disease. Theranostics 2021, 11, 4103–4121. [Google Scholar] [CrossRef]
- Poenaru, S.; Abdallah, S.J.; Corrales-Medina, V.; Cowan, J. COVID-19 and post-infectious myalgic encephalomyelitis/chronic fatigue syndrome: A narrative review. Ther. Adv. Infect. Dis. 2021, 8, 20499361211009385. [Google Scholar] [CrossRef]
- Rivera, M.C.; Mastronardi, C.; Silva-Aldana, C.T.; Arcos-Burgos, M.; Lidbury, B.A. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: A Comprehensive Review. Diagnostics 2019, 9, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Rawaf, H.A.; Alghadir, A.H.; Gabr, S.A. MicroRNAs as Biomarkers of Pain Intensity in Patients With Chronic Fatigue Syndrome. Pain Pract. 2019, 19, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Nepotchatykh, E.; Elremaly, W.; Caraus, I.; Godbout, C.; Leveau, C.; Chalder, L.; Beaudin, C.; Kanamaru, E.; Kosovskaia, R.; Lauzon, S.; et al. Profile of circulating microRNAs in myalgic encephalomyelitis and their relation to symptom severity, and disease pathophysiology. Sci. Rep. 2020, 10, 19620. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
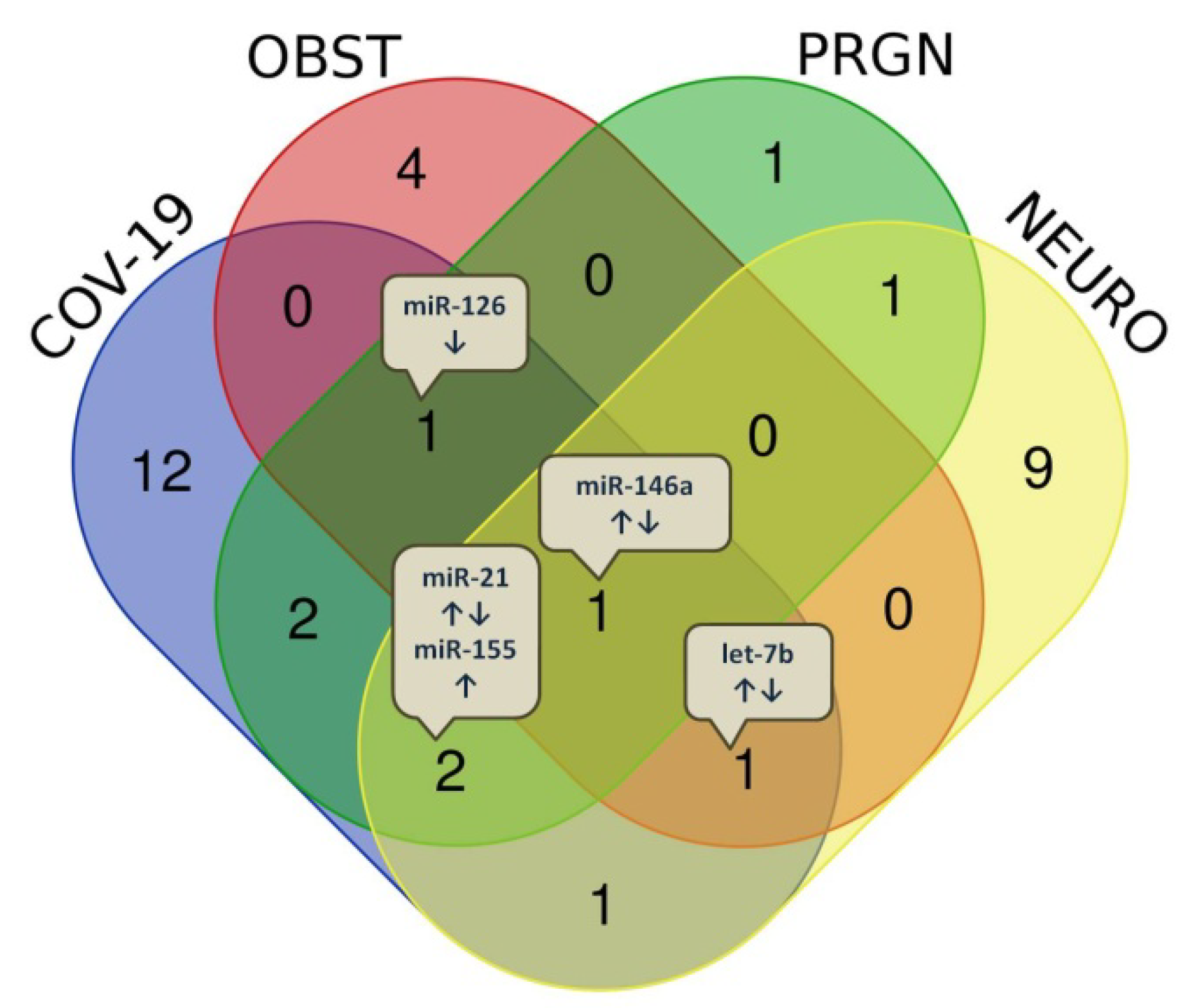
| COVID-19 | COVID-19 and Obesity | COVID-19 and Pregnancy | COVID-19 and Neurological Disorders |
|---|---|---|---|
| miR-16 ↑ miR-6501 ↑ miR-618 ↑ miR-627 ↑ miR-183 ↓ miR-144 ↓ miR-29a ↑ miR-146a ↑↓ miR-155 ↑ let-7b ↑ miR-98 ↑ miR-423 ↑ miR-1246 ↑ miR-32 ↑ miR-21 ↑ miR-208a ↑ miR-499 ↑ miR-126 ↓ miR-221 ↑ miR-2392 ↑ | miR-421 ↑ miR-3909 ↑ miR-4677 ↑ miR-200c ↑ let-7b ↑ miR-146a ↓ miR-126 ↓ | miR-29a ↑ miR-29c ↑ miR-21 ↑ miR-98 ↑ miR-150 ↑ miR-155 ↑ miR-146a ↑ miR-126 ↓ | miR-155 ↑ let-7b ↑↓ miR-107 ↑ miR-29b ↑ let-7a ↓ let-7f ↓ miR-26b ↑ miR-21 ↓ miR-31 ↓ miR-16 ↑ miR-146a ↑↓ miR-558 ↓ miR-150 ↓ miR-124 ↓ miR-143 ↓ |
| Diseases Overlap | miRNAs, n | miRNAs, Name |
|---|---|---|
| COVID-19 Obesity Pregnancy Neurol. diseases | 1 | miR-146a ↑↓ |
| COVID-19 Obesity Pregnancy | 1 | miR-126 ↓ |
| COVID-19 Obesity Neurol. diseases | 1 | let-7b ↑↓ |
| COVID-19 Pregnancy Neurol. diseases | 2 | miR-21 ↑↓ miR-155 ↑ |
| COVID-19 Pregnancy | 2 | miR-29a ↑ miR-98 ↑ |
| COVID-19 Neurol. diseases | 1 | miR-16 ↑ |
| Pregnancy Neurol. diseases | 1 | miR-150 ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankovic, M.; Nikolic, D.; Novakovic, I.; Petrovic, B.; Lackovic, M.; Santric-Milicevic, M. miRNAs as a Potential Biomarker in the COVID-19 Infection and Complications Course, Severity, and Outcome. Diagnostics 2023, 13, 1091. https://doi.org/10.3390/diagnostics13061091
Jankovic M, Nikolic D, Novakovic I, Petrovic B, Lackovic M, Santric-Milicevic M. miRNAs as a Potential Biomarker in the COVID-19 Infection and Complications Course, Severity, and Outcome. Diagnostics. 2023; 13(6):1091. https://doi.org/10.3390/diagnostics13061091
Chicago/Turabian StyleJankovic, Milena, Dejan Nikolic, Ivana Novakovic, Bojana Petrovic, Milan Lackovic, and Milena Santric-Milicevic. 2023. "miRNAs as a Potential Biomarker in the COVID-19 Infection and Complications Course, Severity, and Outcome" Diagnostics 13, no. 6: 1091. https://doi.org/10.3390/diagnostics13061091