
Human Breast Milk: The Key Role in the Maturation of Immune, Gastrointestinal and Central Nervous Systems: A Narrative Review

 , ,
, ,  ,
, 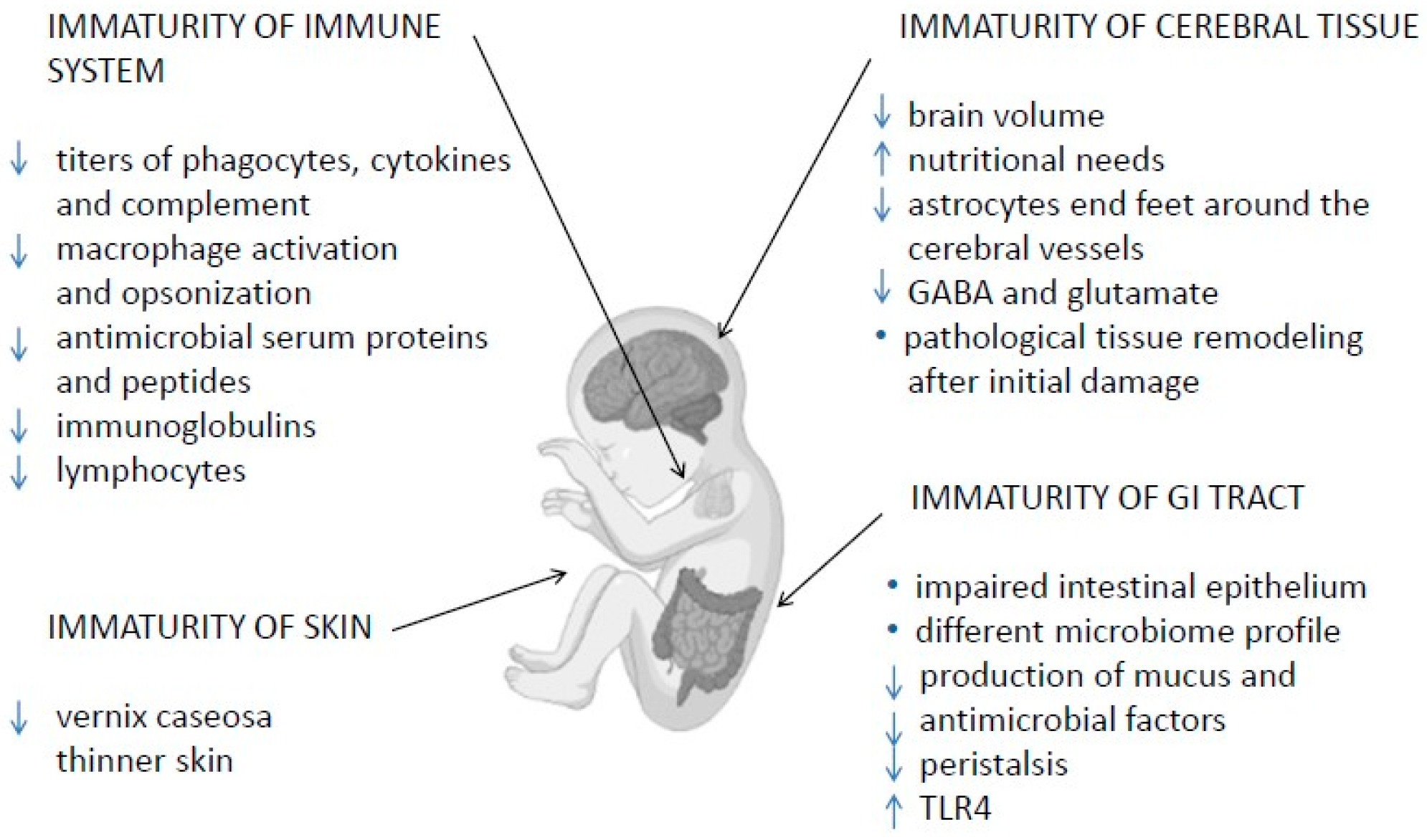{kind=link}
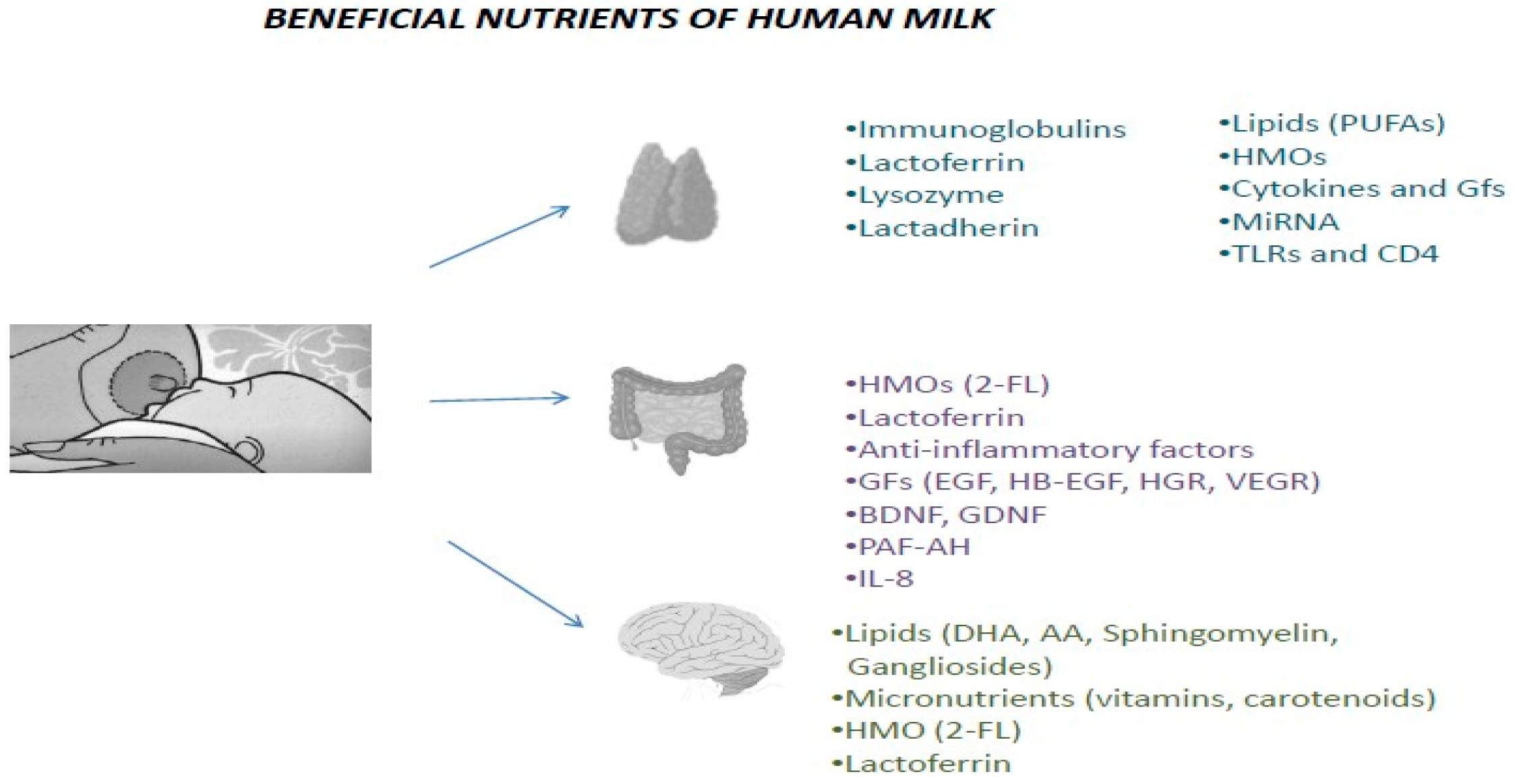{kind=link}
Abstract
:1. Introduction
2. Immune System Immaturity
2.1. Skin and Mucosal Barriers
2.2. Innate and Adaptive Immunity
2.3. Immaturity of Gastrointestinal Tract
2.4. Immaturity of Cerebral Tissue
3. The Importance of HBM for Very Preterm Neonates
3.1. Immunology of HBM
3.2. HBM and Gastrointestinal System
3.3. Significance of HBM for Cerebral Tissue
4. HBM Fortification
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| 2′ FL | 2′-fucosyllactose |
| BBB | Blood–brain barrier |
| BDNF | Brain-derived neurotrophic factor |
| CD14 | Cluster of differentiation 14 |
| CLASBIS | Central line-associated bloodstream infections |
| DCs | Dendritic cells |
| DHA | Docosahexaenoic acid |
| EPG | Epidermal growth factor |
| ELBW | Extremely low-birth-weight infants |
| FIRS | Fetal inflammatory response syndrome |
| GABA | Gamma-aminobutyric acid |
| G-CSF | Granulocyte colony-stimulating factor |
| GDNF | Glial cell-line derived neurotrophic factor |
| GFs | Growth factors |
| HBM | Human breast milk |
| HB-EGF | Heparin-binding epidermal growth factor-like growth factor |
| HGF | Hepatocyte growth factor |
| HMOs | Human breast milk oligosaccharides |
| IL-6 | Interleukin-6 |
| IL-7 | Interleukin-7 |
| IL-8 | Interleukin-8 |
| IL-10 | Interleukin-10 |
| INF-γ | Interferon gamma |
| LBW | Low-birth-weight infant |
| LCPUFAs | Long-chain polyunsaturated fatty acids |
| LISA | Less-invasive surfactant administration |
| LPS | Lipopolysaccharides |
| MHC | Major histocompatibility complex |
| n-CPAP | Nasal-continuous-positive-airway pressure (n-CPAP) |
| NEC | Necrotizing enterocolitis |
| PAF | Platelet-activating factor |
| PUFAs | Polyunsaturated fatty acids |
| TGF-β | Transformation growth factor beta |
| TLR | Toll-like receptors |
| TNF-a | Tumor necrosis factor-a |
| TNFR I and II | Tumor necrosis factor receptor I and II |
| VEGF | Vascular endothelial growth factor |
| VLBW | Very low-birth-weight infants |
| VPI | Very preterm infants |
| WHO | World Health Organization |
References
- Preterm Birth. Available online: https://www.who.int/news-room/fact-sheets/detail/preterm-birth (accessed on 22 August 2022).
- Baroutis, G.; Mousiolis, A.; Mesogitis, S.; Costalos, C.; Antsaklis, A. Preterm birth trends in Greece, 1980–2008: A rising concern. Acta Obstet. Gynecol. Scand. 2013, 92, 575–582. [Google Scholar] [CrossRef]
- Torchin, H.; Ancel, P.Y.; Jarreau, P.H.; Goffinet, F. Epidemiology of preterm birth: Prevalence, recent trends, short- and long-term outcomes. J. Gynecol. Obstet. Biol. Reprod. 2015, 44, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Barfield, W.D. Public Health Implications of Very Preterm Birth. Clin. Perinatol. 2018, 45, 565. [Google Scholar] [CrossRef]
- Thompson, T.; Reynolds, J. The results of intensive care therapy for neonates with respiratory distress syndrome: I. Neonatal mortality rates for neonates with RDS II. Long-term prognosis for survivors with RDS. J. Perinat. Med. 1977, 5, 149–171. [Google Scholar] [CrossRef] [PubMed]
- Iliodromiti, Z.; Christou, E.; Vrachnis, N.; Sokou, R.; Vrachnis, D.; Mihopoulou, G.; Boutsikou, T.; Iacovidou, N. Serum and Urinary N-Terminal Pro-brain Natriuretic Peptides as Biomarkers for Bronchopulmonary Dysplasia of Preterm Neonates. Front. Pediatr. 2020, 8, 588738. [Google Scholar] [CrossRef]
- Cavallin, F.; Doglioni, N.; Brombin, L.; Cavallin, F.; Doglioni, N.; Brombin, L.; Lolli, E.; Loddo, C.; Cavicchiolo, M.E.; Mardegan, V.; et al. Trends in respiratory management of transferred very preterm infants in the last two decades. Pediatr. Pulmonol. 2021, 56, 2604–2610. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.; Blennow, M.; Soll, R. Early surfactant administration with brief ventilation vs selective surfactant and continued mechanical ventilation for preterm infants with or at risk for respiratory distress syndrome. Cochrane Database Syst. Rev. 2007, 2007, CD003063. [Google Scholar] [CrossRef]
- Aldana-Aguirre, J.C.; Pinto, M.; Featherstone, R.M.; Kumar, M. Less invasive surfactant administration versus intubation for surfactant delivery in preterm infants with respiratory distress syndrome: A systematic review and meta-analysis. Arch. Dis. Child. Fetal Neonatal Ed. 2017, 102, F17–F23. [Google Scholar] [CrossRef]
- Miller, J.; Tonkin, E.; Damarell, R.A.; McPhee, A.J.; Suganuma, M.; Suganuma, H.; Middleton, P.F.; Makrides, M.; Collins, C.T. A Systematic Review and Meta-Analysis of Human Milk Feeding and Morbidity in Very Low Birth Weight Infants. Nutrients 2018, 10, 707. [Google Scholar] [CrossRef]
- Underwood, M.A. Human milk for the premature infant. Pediatr. Clin. North Am. 2013, 60, 189–207. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.L.; Kim, J.H. Human milk and necrotizing enterocolitis. Semin. Pediatr. Surg. 2018, 27, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Taïeb, A. Skin barrier in the neonate. Pediatr. Dermatol. 2018, 35 (Suppl. S1), s5–s9. [Google Scholar] [CrossRef] [PubMed]
- Kusari, A.; Han, A.M.; Virgen, C.A.; Matiz, C.; Rasmussen, M.; Friedlander, S.F.; Eichenfield, D.Z. Evidence-based skin care in preterm infants. Pediatr. Dermatol. 2019, 36, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Melville, J.M.; Moss, T.J.M. The immune consequences of preterm birth. Front. Neurosci. 2013, 7, 79. [Google Scholar] [CrossRef]
- Cuenca, A.G.; Wynn, J.L.; Moldawer, L.L.; Levy, O. Role of innate immunity in neonatal infection. Am. J. Perinatol. 2013, 30, 105–112. [Google Scholar] [CrossRef]
- Grases-Pintó, B.; Torres-Castro, P.; Abril-Gil, M.; Castell, M.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J.; Franch, À. A Preterm Rat Model for Immunonutritional Studies. Nutrients 2019, 11, 999. [Google Scholar] [CrossRef]
- Azizia, M.; Lloyd, J.; Allen, M.; Klein, N.; Peebles, D. Immune status in very preterm neonates. Pediatrics 2012, 129, e967–e974. [Google Scholar] [CrossRef]
- Segura-Cervantes, E.; Mancilla-Ramírez, J.; González-Canudas, J.; Alba, E.; Santillán-Ballesteros, R.; Morales-Barquet, D.; Sandoval-Plata, G.; Galindo-Sevilla, N. Inflammatory Response in Preterm and Very Preterm Newborns with Sepsis. Mediat. Inflamm. 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Walker, J.C.; Smolders, M.A.J.C.; Gemen, E.F.A.; Antonius, T.A.J.; Leuvenink, J.; De Vries, E. Development of Lymphocyte Subpopulations in Preterm Infants. Scand. J. Immunol. 2011, 73, 53–58. [Google Scholar] [CrossRef]
- Simister, N.E. Placental transport of immunoglobulin G. Vaccine 2003, 21, 3365–3369. [Google Scholar] [CrossRef]
- Clements, T.; Rice, T.F.; Vamvakas, G.; Barnett, S.; Barnes, M.; Donaldson, B.; Jones, C.E.; Kampmann, B.; Holder, B. Update on Transplacental Transfer of IgG Subclasses: Impact of Maternal and Fetal Factors. Front. Immunol. 2020, 11, 1920. [Google Scholar] [CrossRef]
- Denning, T.W.; Bhatia, A.M.; Kane, A.F.; Patel, R.M.; Denning, P.W. Pathogenesis of NEC: Role of the Innate and Adaptive Immune Response. Semin. Perinatol. 2017, 41, 15. [Google Scholar] [CrossRef] [PubMed]
- Bjarnason, I. Intestinal permeability. Gut 1994, 35 (Suppl. S1), S18. [Google Scholar] [CrossRef] [PubMed]
- Van Elburg, R.M.; Fetter, W.P.F.; Bunkers, C.M.; Heymans, H.S.A. Intestinal permeability in relation to birth weight and gestational and postnatal age. Arch. Dis. Child. Fetal Neonatal Ed. 2003, 88, F52. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.T.; Laker, M.F.; Nelson, R. Intestinal permeability in the newborn. Arch. Dis. Child. 1984, 59, 236. [Google Scholar] [CrossRef] [PubMed]
- McElroy, S.J.; Prince, L.S.; Weitkamp, J.H.; Reese, J.; Slaughter, J.C.; Polk, D.B. Tumor necrosis factor receptor 1-dependent depletion of mucus in immature small intestine: A potential role in neonatal necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G656. [Google Scholar] [CrossRef]
- Berseth, C.L. Gastrointestinal Motility in the Neonate. Clin. Perinatol. 1996, 23, 179–190. [Google Scholar] [CrossRef]
- Montagne, L.; Piel, C.; Lalles, J.P. Effect of diet on mucin kinetics and composition: Nutrition and health implications. Nutr. Rev. 2004, 62, 105–114. [Google Scholar] [CrossRef]
- Pammi, M.; Cope, J.; Tarr, P.I.; Warner, B.B.; Morrow, A.L.; Mai, V.; Gregory, K.E.; Kroll, J.S.; McMurtry, V.; Ferris, M.J.; et al. Intestinal dysbiosis in preterm infants preceding necrotizing enterocolitis: A systematic review and meta-analysis. Microbiome 2017, 5, 1–15. [Google Scholar] [CrossRef]
- Mai, V.; Young, C.M.; Ukhanova, M.; Wang, X.; Sun, Y.; Casella, G.; Theriaque, D.; Li, N.; Sharma, R.; Hudak, M.; et al. Fecal microbiota in premature infants prior to necrotizing enterocolitis. PLoS ONE 2011, 6, e20647. [Google Scholar] [CrossRef]
- Arboleya, S.; Ang, L.; Margolles, A.; Yiyuan, L.; Dongya, Z.; Liang, X.; Solís, G.; Fernández, N.; de Los Reyes-Gavilán, C.G.; Gueimonde, M. Deep 16S rRNA metagenomics and quantitative PCR analyses of the premature infant fecal microbiota. Anaerobe 2012, 18, 378–380. [Google Scholar] [CrossRef] [PubMed]
- Millar, M.; Wilks, M.; Costeloe, K. Probiotics for preterm infants? Arch. Dis. Child. Fetal Neonatal Ed. 2003, 88, F354–F358. [Google Scholar] [CrossRef] [PubMed]
- Reyman, M.; van Houten, M.A.; Watson, R.L.; Chu, M.L.J.N.; Arp, K.; de Waal, W.J.; Schiering, I.; Plötz, F.B.; Willems, R.J.L.; van Schaik, W.; et al. Effects of early-life antibiotics on the developing infant gut microbiome and resistome: A randomized trial. Nat. Commun. 2022, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.T.; Lauren Ruoss, J.; de la Cruz, D.; Li, N.; Bazacliu, C.; Patton, L.; McKinley, K.L.; Garrett, T.J.; Polin, R.A.; Triplett, E.W.; et al. Antibiotics and the developing intestinal microbiome, metabolome and inflammatory environment in a randomized trial of preterm infants. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Niño, D.F.; Sodhi, C.P.; Hackam, D.J. Necrotizing enterocolitis: New insights into pathogenesis and mechanisms. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 590. [Google Scholar] [CrossRef] [PubMed]
- Nosarti, C.; Nam, K.W.; Walshe, M.; Murray, R.M.; Cuddy, M.; Rifkin, L.; Allin, M.P. Preterm birth and structural brain alterations in early adulthood. NeuroImage Clin. 2014, 6, 180–191. [Google Scholar] [CrossRef]
- Ramel, S.E.; Georgieff, M.K. Preterm nutrition and the brain. World. Rev. Nutr. Diet. 2014, 110, 190–200. [Google Scholar] [CrossRef]
- El-Khoury, N.; Braun, A.; Hu, F.; Pandey, M.; Nedergaard, M.; Lagamma, E.F.; Ballabh, P. Astrocyte End-Feet in Germinal Matrix, Cerebral Cortex, and White Matter in Developing Infants. Pediatr. Res. 2006, 59, 673–679. [Google Scholar] [CrossRef]
- Malaeb, S.; Dammann, O. Fetal inflammatory response and brain injury in the preterm newborn. J. Child Neurol. 2009, 24, 1119–1126. [Google Scholar] [CrossRef]
- Raybaud, C.; Ahmad, T.; Rastegar, N.; Shroff, M.; Al Nassar, M. The premature brain: Developmental and lesional anatomy. Neuroradiology 2013, 55 (Suppl. S2), 23–40. [Google Scholar] [CrossRef]
- Basu, S.K.; Pradhan, S.; du Plessis, A.J.; Ben-Ari, Y.; Limperopoulos, C. GABA and glutamate in the preterm neonatal brain: In-vivo measurement by magnetic resonance spectroscopy. Neuroimage 2021, 238, 118215. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.K.; Pradhan, S.; Jacobs, M.B.; Said, M.; Kapse, K.; Murnick, J.; Whitehead, M.T.; Chang, T.; du Plessis, A.J.; Limperopoulos, C. Age and Sex Influences Gamma-aminobutyric Acid Concentrations in the Developing Brain of Very Premature Infants. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Scheinost, D.; Lacadie, C.; Benjamin, J.; Myers, E.; Qiu, M.; Schner, K.C.; Rothman, D.L.; Constable, R.T.; Ment, L.R. GABA, resting-state connectivity and the developing brain. Neonatology 2014, 106, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.J.; Hansen, N.; Fanaroff, A.A.; Wright, L.L.; Carlo, W.A.; Ehrenkranz, R.A.; Lemons, J.A.; Donovan, E.F.; Stark, A.R.; Tyson, J.E.; et al. Late-onset sepsis in very low birth weight neonates: The experience of the NICHD Neonatal Research Network. Pediatrics 2002, 110 Pt 1, 285–291. [Google Scholar] [CrossRef]
- Letouzey, M.; Foix-L’Hélias, L.; Torchin, H.; Mitha, A.; Morgan, A.S.; Zeitlin, J.; Kayem, G.; Maisonneuve, E.; Delorme, P.; Khoshnood, B.; et al. Cause of preterm birth and late-onset sepsis in very preterm infants: The EPIPAGE-2 cohort study. Pediatr. Res. 2021, 90, 584–592. [Google Scholar] [CrossRef]
- Collins, A.; Weitkamp, J.H.; Wynn, J.L. Why are preterm newborns at increased risk of infection? Arch. Dis. Child. Fetal Neonatal Ed. 2018, 103, F391–F394. [Google Scholar] [CrossRef]
- Steiner, L.; Diesner, S.C.; Voitl, P. Risk of infection in the first year of life in preterm children: An Austrian observational study. PLoS ONE 2019, 14, e0224766. [Google Scholar] [CrossRef]
- Yu, J.C.; Khodadadi, H.; Malik, A.; Davidson, B.; Salles, É.D.S.L.; Bhatia, J.; Hale, V.L.; Baban, B. Innate Immunity of Neonates and Infants. Front. Immunol. 2018, 9, 1759. [Google Scholar] [CrossRef]
- Anand, R.J.; Leaphart, C.L.; Mollen, K.P.; Hackam, D.J. The role of the intestinal barrier in the pathogenesis of necrotizing enterocolitis. Shock 2007, 27, 124–133. [Google Scholar] [CrossRef]
- Looi, K.; Evans, D.J.; Garratt, L.W.; Ang, S.; Hillas, J.K.; Kicic, A.; Simpson, S.J. Preterm birth: Born too soon for the developing airway epithelium? Paediatr. Respir. Rev. 2019, 31, 82–88. [Google Scholar] [CrossRef]
- Lewis, E.D.; Richard, C.; Larsen, B.M.; Field, C.J. The Importance of Human Milk for Immunity in Preterm Infants. Clin. Perinatol. 2017, 44, 23–47. [Google Scholar] [CrossRef] [PubMed]
- Berrington, J.E.; Barge, D.; Fenton, A.C.; Cant, A.J.; Spickett, G.P. Lymphocyte subsets in term and significantly preterm UK infants in the first year of life analysed by single platform flow cytometry. Clin. Exp. Immunol. 2005, 140, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Sim, K.; Shaw, A.G.; Randell, P.; Cox, M.J.; McClure, Z.E.; Li, M.S.; Haddad, M.; Langford, P.R.; Cookson, W.O.; Moffatt, M.F.; et al. Dysbiosis Anticipating Necrotizing Enterocolitis in Very Premature Infants. Clin. Infect. Dis. 2015, 60, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.K.; Shah, J.S. Necrotizing Enterocolitis in Very Low Birth Weight Infants: A Systemic Review. ISRN Gastroenterol. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bazacliu, C.; Neu, J. Necrotizing Enterocolitis: Long Term Complications. Curr. Pediatr. Rev. 2019, 15, 115–124. [Google Scholar] [CrossRef]
- Kamity, R.; Kapavarapu, P.K.; Chandel, A. Feeding Problems and Long-Term Outcomes in Preterm Infants—A Systematic Approach to Evaluation and Management. Children 2021, 8, 1158. [Google Scholar] [CrossRef]
- Thompson, A.M.; Bizzarro, M.J. Necrotizing enterocolitis in newborns: Pathogenesis, prevention and management. Drugs 2008, 68, 70–75. [Google Scholar] [CrossRef]
- Baranowski, J.R.; Claud, E.C. Necrotizing Enterocolitis and the Preterm Infant Microbiome. Adv. Exp. Med. Biol. 2019, 1125, 25–36. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- DiGiulio, D.B.; Gervasi, M.T.; Romero, R.; Vaisbuch, E.; Mazaki-Tovi, S.; Kusanovic, J.P.; Seok, K.S.; Gómez, R.; Mittal, P.; Gotsch, F.; et al. Microbial invasion of the amniotic cavity in pregnancies with small-for-gestational-age fetuses. J. Perinat. Med. 2010, 38, 495. [Google Scholar] [CrossRef] [Green Version]
- Ardissone, A.N.; de la Cruz, D.M.; Davis-Richardson, A.G.; Rechcigl, K.T.; Li, N.; Drew, J.C.; Murgas-Torrazza, R.; Sharma, R.; Hudak, M.L.; Triplett, E.W.; et al. Meconium Microbiome Analysis Identifies Bacteria Correlated with Premature Birth. PLoS ONE 2014, 9, e90784. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, H.J.; Wildeboer-Veloo, A.C.; Raangs, G.C.; Wagendorp, A.A.; Klijn, N.; Bindels, J.G.; Welling, G.W. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Sodhi, C.P.; Shi, X.H.; Richardson, W.M.; Grant, Z.S.; Shapiro, R.A.; Prindle, T., Jr.; Branca, M.; Russo, A.; Gribar, S.C.; Ma, C.; et al. Toll-like receptor-4 inhibits enterocyte proliferation via impaired beta-catenin signaling in necrotizing enterocolitis. Gastroenterology 2010, 138, 185–196. [Google Scholar] [CrossRef]
- Lu, P.; Sodhi, C.P.; Hackam, D.J. Toll-like Receptor Regulation of Intestinal Development and Inflammation in the Pathogenesis of Necrotizing Enterocolitis. Pathophysiology 2014, 21, 81. [Google Scholar] [CrossRef]
- Yazji, I.; Sodhi, C.P.; Lee, E.K.; Good, M.; Egan, C.E.; Afrazi, A.; Neal, M.D.; Jia, H.; Lin, J.; Ma, C.; et al. Endothelial TLR4 activation impairs intestinal microcirculatory perfusion in necrotizing enterocolitis via eNOS-NO-nitrite signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 9451–9456. [Google Scholar] [CrossRef]
- Bora, S.; Pritchard, V.E.; Moor, S.; Austin, N.C.; Woodward, L.J. Emotional and behavioural adjustment of children born very preterm at early school age. J. Paediatr. Child Health 2011, 47, 863–869. [Google Scholar] [CrossRef]
- Spittle, A.J.; Morgan, C.; Olsen, J.E.; Novak, I.; Cheong, J.L.Y. Early Diagnosis and Treatment of Cerebral Palsy in Children with a History of Preterm Birth. Clin. Perinatol. 2018, 45, 409–420. [Google Scholar] [CrossRef]
- Pritchard, V.E.; Clark, C.A.C.; Liberty, K.; Champion, P.R.; Wilson, K.; Woodward, L.J. Early school-based learning difficulties in children born very preterm. Early Hum. Dev. 2009, 85, 215–224. [Google Scholar] [CrossRef]
- Williams, J.; Lee, K.J.; Anderson, P.J. Prevalence of motor-skill impairment in preterm children who do not develop cerebral palsy: A systematic review. Dev. Med. Child Neurol. 2010, 52, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Woodward, L.J.; Moor, S.; Hood, K.M.; Champion, P.R.; Foster-Cohen, S.; Inder, T.E.; Austin, N.C. Very preterm children show impairments across multiple neurodevelopmental domains by age 4 years. Arch. Dis. Child. Fetal Neonatal Ed. 2009, 94, 339–344. [Google Scholar] [CrossRef]
- Skinner, A.M.; Narchi, H. Preterm nutrition and neurodevelopmental outcomes. World J. Methodol. 2021, 11, 278–293. [Google Scholar] [CrossRef] [PubMed]
- Malaeb, S.N.; Cohen, S.S.; Virgintino, D.; Stonestreet, B.S. Core ConceptsDevelopment of the Blood-Brain Barrier. Neoreviews 2012, 13, e241–e250. [Google Scholar] [CrossRef]
- Ballabh, P.; Braun, A.; Nedergaard, M. The blood–brain barrier: An overview: Structure, regulation, and clinical implications. Neurobiol. Dis. 2004, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Brunse, A.; Abbaspour, A.; Sangild, P.T. Brain Barrier Disruption and Region-Specific Neuronal Degeneration during Necrotizing Enterocolitis in Preterm Pigs. Dev. Neurosci. 2018, 40, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Wikström, S.; Ley, D.; Hansen-Pupp, I.; Rosén, I.; Hellström-Westas, L. Early amplitude-integrated EEG correlates with cord TNF-alpha and brain injury in very preterm infants. Acta Paediatr. 2008, 97, 915–919. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.R.; Yshii, L.M.; Viel, T.A.; Buck, H.S.; Mattson, M.P.; Scavone, C.; Kawamoto, E.M. Intermittent fasting attenuates lipopolysaccharide-induced neuroinflammation and memory impairment. J. Neuroinflammation 2014, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Konnikova, Y.; Zaman, M.M.; Makda, M.; D’Onofrio, D.; Freedman, S.D.; Martin, C.R. Late Enteral Feedings Are Associated with Intestinal Inflammation and Adverse Neonatal Outcomes. PLoS ONE 2015, 10, e0132924. [Google Scholar] [CrossRef] [PubMed]
- Kwok, T.C.; Dorling, J.; Gale, C. Early enteral feeding in preterm infants. Semin. Perinatol. 2019, 43, 151159. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.L.; Gibbins, S.; Kiss, A.; Bando, N.; Brennan-Donnan, J.; Ng, E.; Campbell, D.M.; Vaz, S.; Fusch, C.; Asztalos, E.; et al. Effect of Supplemental Donor Human Milk Compared With Preterm Formula on Neurodevelopment of Very Low-Birth-Weight Infants at 18 Months: A Randomized Clinical Trial. JAMA 2016, 316, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Chetta, K.E.; Schulz, E.V.; Wagner, C.L. Outcomes improved with human milk intake in preterm and full-term infants. Semin. Perinatol. 2021, 45, 151384. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, P.; Carneiro-Sampaio, M. Immunology of breast milk. Rev. Assoc. Med. Bras. 2016, 62, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Demers-Mathieu, V.; Huston, R.K.; Markell, A.M.; McCulley, E.A.; Martin, R.L.; Dallas, D.C. Antenatal Influenza A-Specific IgA, IgM, and IgG Antibodies in Mother’s Own Breast Milk and Donor Breast Milk, and Gastric Contents and Stools from Preterm Infants. Nutrients 2019, 11, 1567. [Google Scholar] [CrossRef]
- Demers-Mathieu, V.; Underwood, M.A.; Beverly, R.L.; Dallas, D.C. Survival of Immunoglobulins from Human Milk to Preterm Infant Gastric Samples at 1, 2, and 3 Hours Postprandial. Neonatology 2018, 114, 242. [Google Scholar] [CrossRef]
- Bryant, J.; Thistle, J. Anatomy, Colostrum. 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK513256/ (accessed on 8 January 2022).
- Hanson, L.Å.; Winberg, J. Breast milk and defence against infection in the newborn. Arch. Dis. Child. 1972, 47, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.; Petrova, A. Biologically active breast milk proteins in association with very preterm delivery and stage of lactation. J. Perinatol. 2011, 31, 58–62. [Google Scholar] [CrossRef]
- Ochoa, T.J.; Zegarra, J.; Cam, L.; Llanos, R.; Pezo, A.; Cruz, K.; Zea-Vera, A.; Cárcamo, C.; Campos, M.; Bellomo, S. Randomized Controlled Trial of Lactoferrin for Prevention of Sepsis in Peruvian Neonates <2500 Grams. Pediatr. Infect. Dis. J. 2015, 34, 571. [Google Scholar] [CrossRef]
- Pammi, M.; Suresh, G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2017, 2017, CD007137. [Google Scholar] [CrossRef]
- Razak, A.; Hussain, A. Lactoferrin Supplementation to Prevent Late-Onset Sepsis in Preterm Infants: A Meta-Analysis. Am. J. Perinatol. 2021, 38, 283–290. [Google Scholar] [CrossRef]
- Ellison, R.T.; Giehl, T.J. Killing of gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Investig. 1991, 88, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, C.E.; Kashyap, S.; Heird, W.C.; Thormar, H. Antiviral and antibacterial lipids in human milk and infant formula feeds. Arch. Dis. Child. 1990, 65, 861. [Google Scholar] [CrossRef] [PubMed]
- Schroten, H.; Hanisch, F.G.; Plogmann, R.; Hacker, J.; Uhlenbruck, G.; Nobis-Bosch, R.; Wahn, V. Inhibition of adhesion of S-fimbriated Escherichia coli to buccal epithelial cells by human milk fat globule membrane components: A novel aspect of the protective function of mucins in the nonimmunoglobulin fraction. Infect. Immun. 1992, 60, 2893–2899. [Google Scholar] [CrossRef] [PubMed]
- Salamone, M.; Di Nardo, V. Effects of human milk oligosaccharides (HMOs) on gastrointestinal health. Front. Biosci. 2020, 12, 183–198. [Google Scholar] [CrossRef]
- Triantis, V.; Bode, L.; van Neerven, J.R.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.Y.; Comstock, S.S.; Shunk, J.M.; Monaco, M.H.; Donovan, S.M. Natural killer cell populations and cytotoxic activity in pigs fed mother’s milk, formula, or formula supplemented with bovine lactoferrin. Pediatr. Res. 2013, 74, 402–407. [Google Scholar] [CrossRef]
- Garofalo, R. Cytokines in Human Milk. J. Pediatr. 2010, 156, S36–S40. [Google Scholar] [CrossRef]
- Aspinall, R.; Prentice, A.M.; Ngom, P.T. Interleukin 7 from Maternal Milk Crosses the Intestinal Barrier and Modulates T-Cell Development in Offspring. PLoS ONE 2011, 6, e20812. [Google Scholar] [CrossRef]
- Goldman, A.S.; Chheda, S. The Immune System in Human Milk: A Historic Perspective. Ann. Nutr. Metab. 2021, 77, 189–196. [Google Scholar] [CrossRef]
- Johnnidis, J.B.; Harris, M.H.; Wheeler, R.T.; Stehling-Sun, S.; Lam, M.H.; Kirak, O.; Brummelkamp, T.R.; Fleming, M.D.; Camargo, F.D. Regulation of progenitor cell proliferation and granulocyte function by microRNA-223. Nature 2008, 451, 1125–1129. [Google Scholar] [CrossRef]
- Ventura, A.; Young, A.G.; Winslow, M.M.; Lintault, L.; Meissner, A.; Erkeland, S.J.; Newman, J.; Bronson, R.T.; Crowley, D.; Stone, J.R.; et al. Targeted deletion reveals essential and overlapping functions of the miR-17 through 92 family of miRNA clusters. Cell 2018, 132, 875. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Rimer, J.M.; Good, M. The Role of Human Milk Oligosaccharides and Probiotics on the Neonatal Microbiome and Risk of Necrotizing Enterocolitis: A Narrative Review. Nutrients 2020, 12, 3052. [Google Scholar] [CrossRef] [PubMed]
- Young, L.; McGuire, W. Immunologic Properties of Human Milk and Clinical Implications in the Neonatal Population. Neoreviews 2020, 21, e809–e816. [Google Scholar] [CrossRef]
- York, D.J.; Smazal, A.L.; Robinson, D.T.; De Plaen, I.G. Human Milk Growth Factors and Their Role in NEC Prevention: A Narrative Review. Nutrients 2021, 13, 3751. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B. Milk Epidermal Growth Factor and Gut Protection. J. Pediatr. 2010, 156 (Suppl. S2), S31. [Google Scholar] [CrossRef]
- Radulescu, A.; Zhang, H.Y.; Chen, C.L.; Chen, Y.; Zhou, Y.; Yu, X.; Otabor, I.; Olson, J.K.; Besner, G.E. Heparin-Binding Egf-Like Growth Factor Promotes Intestinal Anastomotic Healing. J. Surg. Res. 2011, 171, 540. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Olson, J.; Yang, J.; Besner, G.E. Heparin-binding EGF-like growth factor promotes neuronal nitric oxide synthase expression and protects the enteric nervous system after necrotizing enterocolitis. Pediatr. Res. 2017, 82, 490–500. [Google Scholar] [CrossRef]
- Gila-Diaz, A.; Arribas, S.M.; Algara, A.; Martín-Cabrejas, M.A.; López de Pablo, Á.L.; Sáenz de Pipaón, M.; Ramiro-Cortijo, D. A Review of Bioactive Factors in Human Breastmilk: A Focus on Prematurity. Nutrients 2019, 11, 1307. [Google Scholar] [CrossRef]
- Li, R.; Xia, W.; Zhang, Z.; Wu, K. S100B Protein, Brain-Derived Neurotrophic Factor, and Glial Cell Line-Derived Neurotrophic Factor in Human Milk. PLoS ONE 2011, 6, e21663. [Google Scholar] [CrossRef]
- Furukawa, M.; Narahara, H.; Yasuda, K.; Johnston, J.M. Presence of platelet-activating factor-acetylhydrolase in milk. J. Lipid Res. 1993, 34, 1603–1609. [Google Scholar] [CrossRef]
- Maheshwari, A.; Lu, W.; Lacson, A.; Barleycorn, A.A.; Nolan, S.; Christensen, R.D.; Calhoun, D.A. Effects of interleukin-8 on the developing human intestine. Cytokine 2002, 20, 256–267. [Google Scholar] [CrossRef]
- González, H.F.; Visentin, S. Nutrients and neurodevelopment: Lipids. Update. Arch. Argent. Pediatr. 2016, 114, 472–476. [Google Scholar] [CrossRef]
- Martinat, M.; Rossitto, M.; Di Miceli, M.; Layé, S. Perinatal Dietary Polyunsaturated Fatty Acids in Brain Development, Role in Neurodevelopmental Disorders. Nutrients 2021, 13, 1185. [Google Scholar] [CrossRef]
- Agbaga, M.P.; Mandal, N.A.; Anderson, R.E. Retinal very long-chain PUFAs: New insights from studies on ELOVL4 protein. J. Lipid Res. 2010, 51, 1624. [Google Scholar] [CrossRef]
- Dalmeijer, G.W.; Wijga, A.H.; Gehring, U.; Renders, C.M.; Koppelman, G.H.; Smit, H.A.; van Rossem, L. Fatty acid composition in breastfeeding and school performance in children aged 12 years. Eur. J. Nutr. 2016, 55, 2199. [Google Scholar] [CrossRef]
- Jasani, B.; Simmer, K.; Patole, S.K.; Rao, S.C. Long chain polyunsaturated fatty acid supplementation in infants born at term. Cochrane Database Syst. Rev. 2017, 2017, CD000376. [Google Scholar] [CrossRef]
- Delgado-Noguera, M.F.; Calvache, J.A.; Bonfill Cosp, X.; Kotanidou, E.P.; Galli-Tsinopoulou, A. Supplementation with long chain polyunsaturated fatty acids (LCPUFA) to breastfeeding mothers for improving child growth and development. Cochrane Database Syst. Rev. 2015, 2015, CD007901. [Google Scholar] [CrossRef]
- Schneider, N.; Hauser, J.; Oliveira, M.; Cazaubon, E.; Mottaz, S.C.; O’Neill, B.V.; Steiner, P.; Deoni, S.C.L. Sphingomyelin in Brain and Cognitive Development: Preliminary Data. eNeuro 2019, 6, ENEURO.0421-18.2019. [Google Scholar] [CrossRef]
- Perea-Sanz, L.; Garcia-Llatas, G.; Lagarda, M.J. Gangliosides in human milk and infant formula: A review on analytical techniques and contents. Food Rev. Int. 2017, 34, 511–538. [Google Scholar] [CrossRef]
- Rahmann, H. Brain gangliosides and memory formation. Behav. Brain Res. 1995, 66, 105–116. [Google Scholar] [CrossRef]
- Rösner, H. Developmental expression and possible roles of gangliosides in brain development. Prog. Mol. Subcell. Biol. 2003, 32, 49–73. [Google Scholar] [CrossRef]
- Berger, P.K.; Plows, J.F.; Jones, R.B.; Alderete, T.L.; Yonemitsu, C.; Poulsen, M.; Ryoo, J.H.; Peterson, B.S.; Bode, L.; Goran, M.I. Human milk oligosaccharide 2’-fucosyllactose links feedings at 1 month to cognitive development at 24 months in infants of normal and overweight mothers. PLoS ONE 2020, 15, e0228323. [Google Scholar] [CrossRef]
- Boylan, L.M.; Hart, S.; Porter, K.B.; Driskell, J.A. Vitamin B-6 content of breast milk and neonatal behavioral functioning. J. Am. Diet. Assoc. 2002, 102, 1433–1438. [Google Scholar] [CrossRef]
- Zielinska, M.A.; Hamulka, J.; Grabowicz-Chadrzyńska, I.; Bryś, J.; Wesolowska, A. Association between Breastmilk LC PUFA, Carotenoids and Psychomotor Development of Exclusively Breastfed Infants. Int. J. Environ. Res. Public Health 2019, 16, 1144. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, Z.; Zhu, X.; Shi, Y.; Tian, D.; Zhao, F.; Liu, N.; Hüppi, P.S.; Troy, F.A., 2nd; Wang, B. Lactoferrin Promotes Early Neurodevelopment and Cognition in Postnatal Piglets by Upregulating the BDNF Signaling Pathway and Polysialylation. Mol. Neurobiol. 2015, 52, 256–269. [Google Scholar] [CrossRef]
- van de Looij, Y.; Ginet, V.; Chatagner, A.; Toulotte, A.; Somm, E.; Hüppi, P.S.; Sizonenko, S.V. Lactoferrin during lactation protects the immature hypoxic-ischemic rat brain. Ann. Clin. Transl. Neurol. 2014, 1, 955–967. [Google Scholar] [CrossRef]
- Patel, A.L.; Johnson, T.J.; Engstrom, J.L.; Fogg, L.F.; Jegier, B.J.; Bigger, H.R.; Meier, P.P. Impact of Early Human Milk on Sepsis and Health Care Costs in Very Low Birth Weight Infants. J. Perinatol. 2013, 33, 514. [Google Scholar] [CrossRef]
- Hurley, W.L.; Theil, P.K. Perspectives on Immunoglobulins in Colostrum and Milk. Nutrients 2011, 3, 442–474. [Google Scholar] [CrossRef]
- Sørensen, V.; Rasmussen, I.B.; Sunvold, V.; Michaelsen, T.E.; Sandlie, I. Structural requirements for incorporation of J chain into human IgM and IgA. Int. Immunol. 2000, 12, 19–27. [Google Scholar] [CrossRef]
- Thai, J.D.; Gregory, K.E. Bioactive Factors in Human Breast Milk Attenuate Intestinal Inflammation during Early Life. Nutrients 2020, 12, 581. [Google Scholar] [CrossRef] [Green Version]
- Newburg, D.S.; Peterson, J.A.; Ruiz-Palacios, G.M.; Matson, D.O.; Morrow, A.L.; Shults, J.; Guerrero, M.L.; Chaturvedi, P.; Newburg, S.O.; Scallan, C.D.; et al. Role of human-milk lactadherin in protectoin against symptomatic rotavirus infection. Lancet 1998, 351, 1160–1164. [Google Scholar] [CrossRef]
- Das, U.N. Arachidonic acid and other unsaturated fatty acids and some of their metabolites function as endogenous antimicrobial molecules: A review. J. Adv. Res. 2018, 11, 57. [Google Scholar] [CrossRef]
- Azagra-Boronat, I.; Massot-Cladera, M.; Mayneris-Perxachs, J.; Knipping, K.; Van’t Land, B.; Tims, S.; Stahl, B.; Garssen, J.; Franch, À.; Castell, M. Immunomodulatory and Prebiotic Effects of 2’-Fucosyllactose in Suckling Rats. Front. Immunol. 2019, 10, 1773. [Google Scholar] [CrossRef]
- M’Rabet, L.; Vos, A.P.; Boehm, G.; Garssen, J. Breast-Feeding and Its Role in Early Development of the Immune System in Infants: Consequences for Health Later in Life. J. Nutr. 2008, 138, 1782S–1790S. [Google Scholar] [CrossRef]
- Comstock, S.S.; Reznikov, E.A.; Contractor, N.; Donovan, S.M. Dietary bovine lactoferrin alters mucosal and systemic immune cell responses in neonatal piglets. J. Nutr. 2014, 144, 525–532. [Google Scholar] [CrossRef]
- Cacho, N.T.; Lawrence, R.M. Innate Immunity and Breast Milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef]
- Quitadamo, P.A.; Comegna, L.; Cristalli, P. Anti-Infective, Anti-Inflammatory, and Immunomodulatory Properties of Breast Milk Factors for the Protection of Infants in the Pandemic From COVID-19. Front. Public Health 2021, 8, 964. [Google Scholar] [CrossRef]
- Chowning, R.; Radmacher, P.; Lewis, S.; Serke, L.; Pettit, N.; Adamkin, D.H. A retrospective analysis of the effect of human milk on prevention of necrotizing enterocolitis and postnatal growth. J. Perinatol. 2015, 36, 221–224. [Google Scholar] [CrossRef]
- Herrmann, K.; Carroll, K. An exclusively human milk diet reduces necrotizing enterocolitis. Breastfeed. Med. 2014, 9, 184–190. [Google Scholar] [CrossRef]
- Thomson, P.; Medina, D.A.; Garrido, D. Human milk oligosaccharides and infant gut bifidobacteria: Molecular strategies for their utilization. Food Microbiol. 2018, 75, 37–46. [Google Scholar] [CrossRef]
- Arboleya, S.; Binetti, A.; Salazar, N.; Fernández, N.; Solís, G.; Hernández-Barranco, A.; Margolles, A.; de Los Reyes-Gavilán, C.G.; Gueimonde, M. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 2012, 79, 763–772. [Google Scholar] [CrossRef]
- Underwood, M.A.; Gaerlan, S.; De Leoz, M.L.; Dimapasoc, L.; Kalanetra, K.M.; Lemay, D.G.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Human Milk Oligosaccharides in Premature Infants: Absorption, Excretion and Influence on the Intestinal Microbiota. Pediatr. Res. 2015, 78, 670. [Google Scholar] [CrossRef]
- Lewis, Z.T.; Totten, S.M.; Smilowitz, J.T.; Popovic, M.; Parker, E.; Lemay, D.G.; Van Tassell, M.L.; Miller, M.J.; Jin, Y.S.; German, J.B.; et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 2015, 3, 13. [Google Scholar] [CrossRef]
- Singh, P.; Sanchez-Fernandez, L.L.; Ramiro-Cortijo, D.; Ochoa-Allemant, P.; Perides, G.; Liu, Y.; Medina-Morales, E.; Yakah, W.; Freedman, S.D.; Martin, C.R. Maltodextrin-induced intestinal injury in a neonatal mouse model. Dis. Model. Mech. 2020, 13, dmm044776. [Google Scholar] [CrossRef]
- Guglielmi, F.W.; Boggio-Bertinet, D.; Federico, A.; Forte, G.B.; Guglielmi, A.; Loguercio, C.; Mazzuoli, S.; Merli, M.; Palmo, A.; Panella, C.; et al. Total parenteral nutrition-related gastroenterological complications. Dig. Liver Dis. 2006, 38, 623–642. [Google Scholar] [CrossRef]
- Franco, S.; Goriacko, P.; Rosen, O.; Morgan-Joseph, T. Incidence of Complications Associated with Parenteral Nutrition in Preterm Infants. JPEN J. Parenter. Enter. Nutr. 2021, 45, 1204–1212. [Google Scholar] [CrossRef]
- Vohr, B.R.; Poindexter, B.B.; Dusick, A.M.; McKinley, L.T.; Higgins, R.D.; Langer, J.C.; Poole, W.K. Beneficial Effects of Breast Milk in the Neonatal Intensive Care Unit on the Developmental Outcome of Extremely Low Birth Weight Infants at 18 Months of Age. Pediatrics 2006, 118, e115–e123. [Google Scholar] [CrossRef]
- Vohr, B.R.; Poindexter, B.B.; Dusick, A.M.; McKinley, L.T.; Higgins, R.D.; Langer, J.C.; Poole, W.K. Persistent Beneficial Effects of Breast Milk Ingested in the Neonatal Intensive Care Unit on Outcomes of Extremely Low Birth Weight Infants at 30 Months of Age. Pediatrics 2007, 120, e953–e959. [Google Scholar] [CrossRef]
- Belfort, M.B.; Anderson, P.J.; Nowak, V.A.; Lee, K.J.; Molesworth, C.; Thompson, D.K.; Doyle, L.W.; Inder, T.E. Breast Milk Feeding, Brain Development, and Neurocognitive Outcomes: A 7-Year Longitudinal Study in Infants Born at Less Than 30 Weeks’ Gestation. J. Pediatr. 2016, 177, 133–139.e1. [Google Scholar] [CrossRef]
- Ong, M.L.; Belfort, M.B. Preterm infant nutrition and growth with a human milk diet. Semin. Perinatol. 2021, 45, 151383. [Google Scholar] [CrossRef]
- Hegar, B.; Wibowo, Y.; Basrowi, R.W.; Ranuh, R.G.; Sudarmo, S.M.; Munasir, Z.; Atthiyah, A.F.; Widodo, A.D.; Supriatmo Kadim, M. The Role of Two Human Milk Oligosaccharides, 2′-Fucosyllactose and Lacto-N-Neotetraose, in Infant Nutrition. Pediatr. Gastroenterol. Hepatol. Nutr. 2019, 22, 330. [Google Scholar] [CrossRef] [PubMed]
- Brown Belfort, M. The Science of Breastfeeding and Brain Development. Breastfeed. Med. 2017, 12, 459. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Shi, X.; Zhang, L.; Zhou, Y.; Cai, Y.; Jiang, W.; Zhou, Q. Evidence summary of human milk fortifier in preterm infants. Transl. Pediatr. 2021, 10, 3058. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, C.J.; Kang, N.M. Maternal Factors Affecting the Macronutrient Composition of Transitional Human Milk. Int. J. Environ. Res. Public Health 2022, 19, 3308. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimitroglou, M.; Iliodromiti, Z.; Christou, E.; Volaki, P.; Petropoulou, C.; Sokou, R.; Boutsikou, T.; Iacovidou, N. Human Breast Milk: The Key Role in the Maturation of Immune, Gastrointestinal and Central Nervous Systems: A Narrative Review. Diagnostics 2022, 12, 2208. https://doi.org/10.3390/diagnostics12092208
Dimitroglou M, Iliodromiti Z, Christou E, Volaki P, Petropoulou C, Sokou R, Boutsikou T, Iacovidou N. Human Breast Milk: The Key Role in the Maturation of Immune, Gastrointestinal and Central Nervous Systems: A Narrative Review. Diagnostics. 2022; 12(9):2208. https://doi.org/10.3390/diagnostics12092208
Chicago/Turabian StyleDimitroglou, Margarita, Zoi Iliodromiti, Evangelos Christou, Paraskevi Volaki, Chrysa Petropoulou, Rozeta Sokou, Theodora Boutsikou, and Nicoletta Iacovidou. 2022. "Human Breast Milk: The Key Role in the Maturation of Immune, Gastrointestinal and Central Nervous Systems: A Narrative Review" Diagnostics 12, no. 9: 2208. https://doi.org/10.3390/diagnostics12092208