
CRISPR-Cas Systems-Based Bacterial Detection: A Scoping Review
by
 , , and
, , and
Kasturi Selvam
1, 
Mohamad Ahmad Najib
1 ,
,
Muhammad Fazli Khalid
1,
Mehmet Ozsoz
1,2 and
Ismail Aziah
1,* 1
Institute for Research in Molecular Medicine (INFORMM), Health Campus, Universiti Sains Malaysia, Kubang Kerian 16150, Kelantan, Malaysia
2
Department of Biomedical Engineering, Near East University, Nicosia 99138, Turkey
*
Author to whom correspondence should be addressed.
Diagnostics 2022, 12(6), 1335; https://doi.org/10.3390/diagnostics12061335
Submission received: 15 March 2022
/
Revised: 22 May 2022
/
Accepted: 24 May 2022
/
Published: 27 May 2022
(This article belongs to the Section Diagnostic Microbiology and Infectious Disease)
Abstract
:Recently, CRISPR-Cas system-based assays for bacterial detection have been developed. The aim of this scoping review is to map existing evidence on the utilization of CRISPR-Cas systems in the development of bacterial detection assays. A literature search was conducted using three databases (PubMed, Scopus, and Cochrane Library) and manual searches through the references of identified full texts based on a PROSPERO-registered protocol (CRD42021289140). Studies on bacterial detection using CRISPR-Cas systems that were published before October 2021 were retrieved. The Critical Appraisal Skills Programme (CASP) qualitative checklist was used to assess the risk of bias for all the included studies. Of the 420 studies identified throughout the search, 46 studies that met the inclusion criteria were included in the final analysis. Bacteria from 17 genera were identified utilising CRISPR-Cas systems. Most of the bacteria came from genera such as Staphylococcus, Escherichia, Salmonella, Listeria, Mycobacterium and Streptococcus. Cas12a (64%) is the most often used Cas enzyme in bacterial detection, followed by Cas13a (13%), and Cas9 (11%). To improve the signal of detection, 83% of the research exploited Cas enzymes’ trans-cleavage capabilities to cut tagged reporter probes non-specifically. Most studies used the extraction procedure, whereas only 17% did not. In terms of amplification methods, isothermal reactions were employed in 66% of the studies, followed by PCR (23%). Fluorescence detection (67%) was discovered to be the most commonly used method, while lateral flow biosensors (13%), electrochemical biosensors (11%), and others (9%) were found to be less commonly used. Most of the studies (39) used specific bacterial nucleic acid sequences as a target, while seven used non-nucleic acid targets, including aptamers and antibodies particular to the bacteria under investigation. The turnaround time of the 46 studies was 30 min to 4 h. The limit of detection (LoD) was evaluated in three types of concentration, which include copies per mL, CFU per mL and molarity. Most of the studies used spiked samples (78%) rather than clinical samples (22%) to determine LoD. This review identified the gap in clinical accuracy evaluation of the CRISPR-Cas system in bacterial detection. More research is needed to assess the diagnostic sensitivity and specificity of amplification-free CRISPR-Cas systems in bacterial detection for nucleic acid-based tests.
1. Introduction
Bacterial infection occurs when bacteria enter the body, multiply, and cause a reaction in the body. Many patients with suspected bacterial infections are given empiric antimicrobial medicine instead of proper treatment, which leads to an increase in antimicrobial resistance [1]. The ESKAPE bacteria (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumanii, Pseudomonas aeruginosa, and Enterobacteriaceae) are the microorganisms that are primarily involved in the resistance process, emphasising their ability to “escape” from common antibacterial treatments [2]. Antibiotic resistance pathogens have emerged as a result of a lack of rapid diagnostic tests with high sensitivity and specificity.
The majority of clinical microbiology laboratories still use the culture method for the detection of most bacteria from clinical samples; however, this process takes days to weeks to complete, relies on phenotypic biochemical characterization and requires skilled laboratory staff [3,4]. Apart from that, antibody detection methods have been used to detect circulating antibodies that are specific to respective bacteria; however, during acute infection, these results may be negative because the patients have not yet generated antibody response, and cross-reactions with unrelated IgM can occur [5]. Indeed, over the last two decades, there has been a surge in the development of diagnostic tests based on amplification and detection of specific bacterial nucleotide sequences. The majority of nucleic acid amplification methods use polymerase chain reaction (PCR) and can detect a pathogen of interest with high sensitivity and specificity; however, the requirement for expensive instruments (thermocycler) and reagents prevents such diagnostic tests from being used in areas with limited resources, such as on the battlefield or in developing countries [6]. As a result, the test cannot meet the ASSURED criteria (Affordable, Sensitive, Specific, User-friendly, Robust and rapid, Equipment-free, Deliverable) for developing point-of-care (POC) diagnostic tests [7].
Recently, enzymes from clustered, regularly interspaced, short palindromic repeats (CRISPR)- CRISPR associated protein (Cas) systems have been adapted for the specific, rapid, sensitive, and portable sensing of nucleic acids. The CRISPR–Cas system is composed of RNA-guided endonucleases, and it is an adaptive immune system that protects its hosts from bacteriophage predation and parasitism by other mobile genetic elements (MGEs) [8]. CRISPR-Cas system has been hailed as a versatile and reliable method for genome editing since its discovery. The CRISPR-Cas system has a diverse set of Cas proteins and genomic loci architecture, which has piqued researchers’ attention in a variety of biotech disciplines, including infectious disease detection [9]. Various CRISPR–Cas system-based methods have been developed for bacterial detection. However, there are limitations in terms of the number of comprehensive reports on successful bacterial detection using CRISPR–Cas technology. Therefore, this review aims to highlight the advances made in using the CRISPR-Cas systems to detect bacterial diseases.
2. Methods
The present scoping review utilized the updated Preferred Reporting Items for Systematic review and Meta-Analyses extension for Scoping Reviews (PRISMA-ScR) guidelines [10,11]. The PRISMA-ScR aims to provide guidance on the reporting of scoping reviews. This review has been registered in the International Prospective Register of Systematic Reviews (PROSPERO), with registration number CRD42021289140.
2.1. Search Strategy
The literature search was conducted in October 2021 through three databases (PubMed, Scopus, and Cochrane Library) using lists of keywords referring to the Medical Subject Headings (MeSH) thesaurus. These keywords were combined using the Boolean operators as follows: [“CRISPR”] AND [“bacteria”] AND [“detection”]. An additional search was conducted by manually screening the references of the retrieved literature.
2.2. Selection of Studies
Articles were excluded if (i) the studies did not involve the development and evaluation of CRISPR-based detection; (ii) the studies were published in languages other than English or Malay; (iii) the studies detected pathogens other than bacteria. The retrieved literature was downloaded into the Endnote reference manager and duplicates were identified and removed. The references were distributed to two authors (K.S. and M.A.N.), who independently reviewed all the articles for title and abstract screening. A satisfactory agreement for the screening process was assessed between the authors. Discrepancies between the authors were solved through a discussion with a third author (M.F.K.). Two authors (K.S. and M.A.N.) performed full-text screening and summarized the findings.
2.3. Questions for the Quality Assessment of Retrieved Studies
The studies included from PubMed, Scopus and Cochrane Library databases were analysed according to their quality standards by seven questions defined by the Critical Appraisal Skills Programme (CASP) qualitative checklist. For each retrieved article, the questions were answered by two authors who filled in an Excel table with the answers “no”, “yes” or “unclear”. Discrepancies between these authors were solved through a discussion with third authors. The questions are listed as follows: 1. Was there a clear statement of the aims of the research? 2. Was the research design appropriate to address the aims of the research? 3. Was the execution of the index test described in sufficient detail to permit replication of the test? 4. Did the study provide a clear definition of what was considered to be a positive result? 5. If necessary, have ethical issues been taken into consideration? 6. Was the data analysis sufficiently rigorous? 7. Is there a clear statement of findings?
2.4. Data Extraction
The following data were extracted for descriptive analysis and comparison in terms of percentage: type of bacteria, type of CRISPR-Cas enzymes, the occurrence of trans-cutting, extraction utilization, amplification methods, detection techniques, type of targets, assay time, and limit of detection (LoD).
3. Results
3.1. Search Results
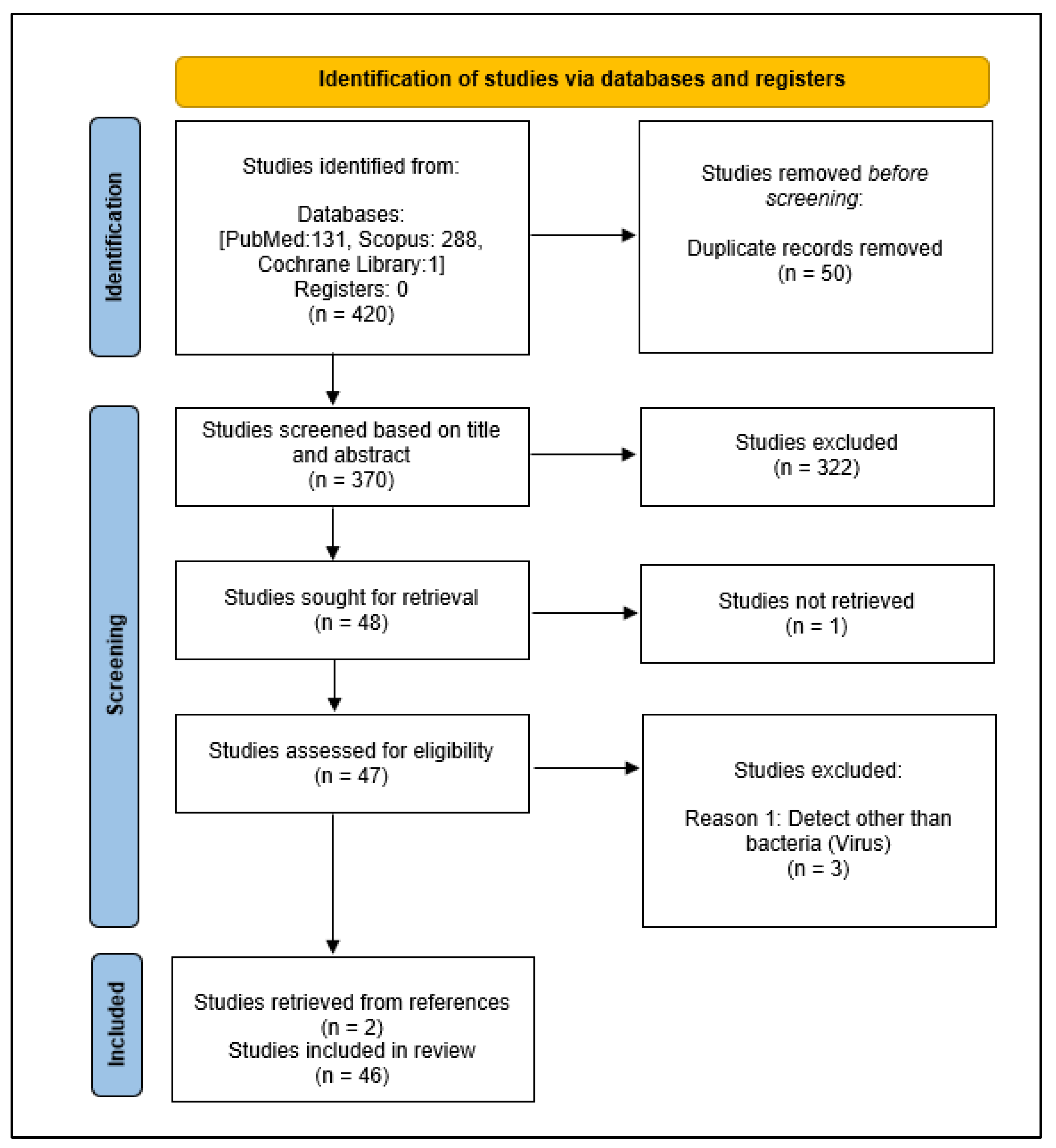
A total of 420 studies were identified from the three databases and 50 duplicates were removed. After screening the titles and abstracts, 322 studies that are not relevant were excluded. One article was not retrieved. Based on the study criteria, three studies were excluded during full-text screening. A total of two new studies were identified through manual searches of the lists of references. The remaining 46 studies were included in the final review (Figure 1). The characteristics of the 46 studies were summarized in Table 1.
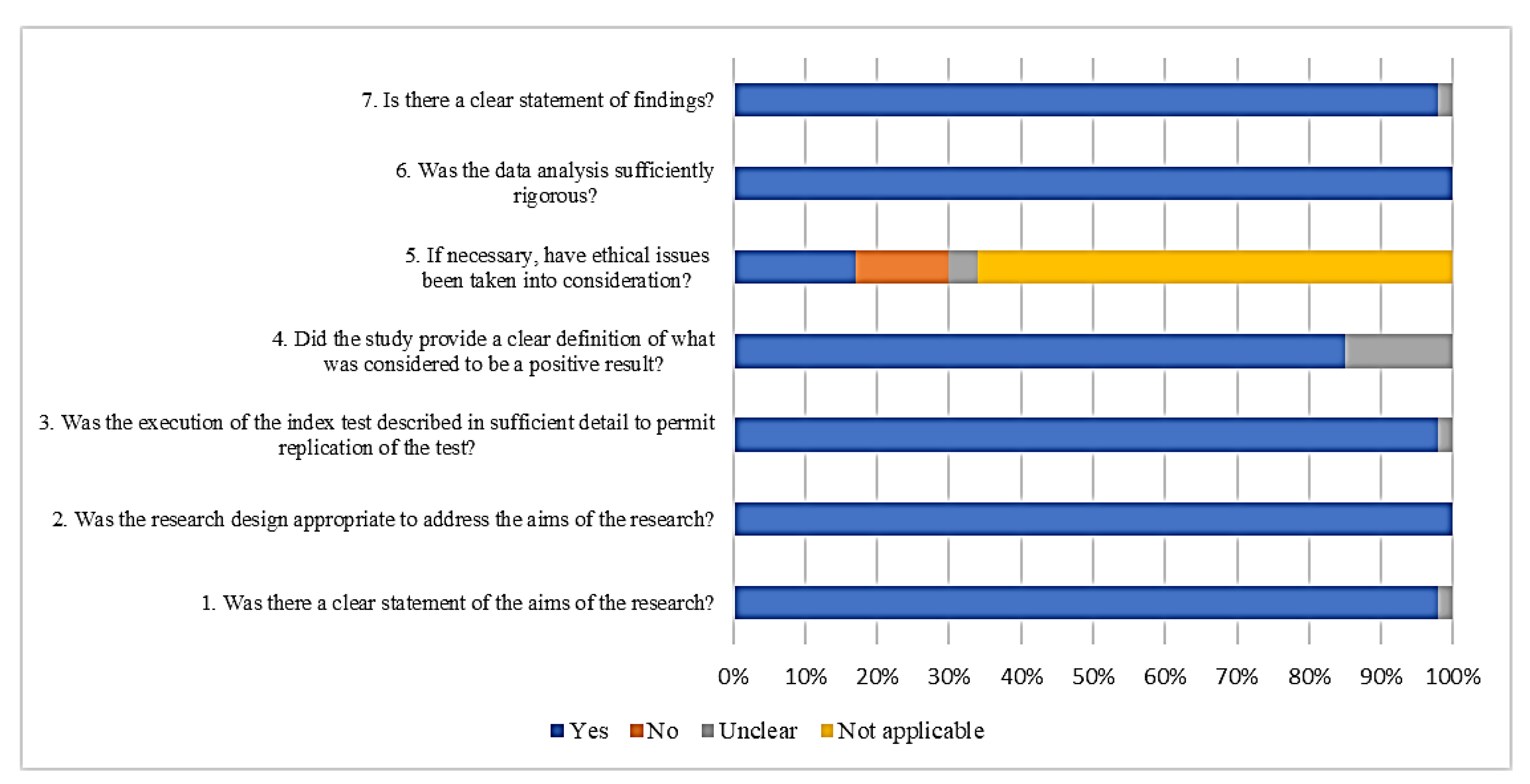
A summary of the CASP Qualitative Checklist assessment is presented in Figure 2. The overall results of the quality assessment showed a low risk of bias in all 46 studies. Regarding the answers for each quality question, around 2% of the studies did not clearly state the aims of the research, but all the studies showed an appropriate experimental design (100%). Most of the studies (98%) explained the index test in detail. When we analysed the definition of a positive result for those studies testing diagnostic approaches, 15% of the studies failed to report a cut off value. Thirteen percent of the studies did not consider ethical issues when biological samples were used. All studies (100%) reported sufficiently rigorous analyses of their data. Most of the studies (98%) displayed their findings clearly.
3.2. Types of Bacteria
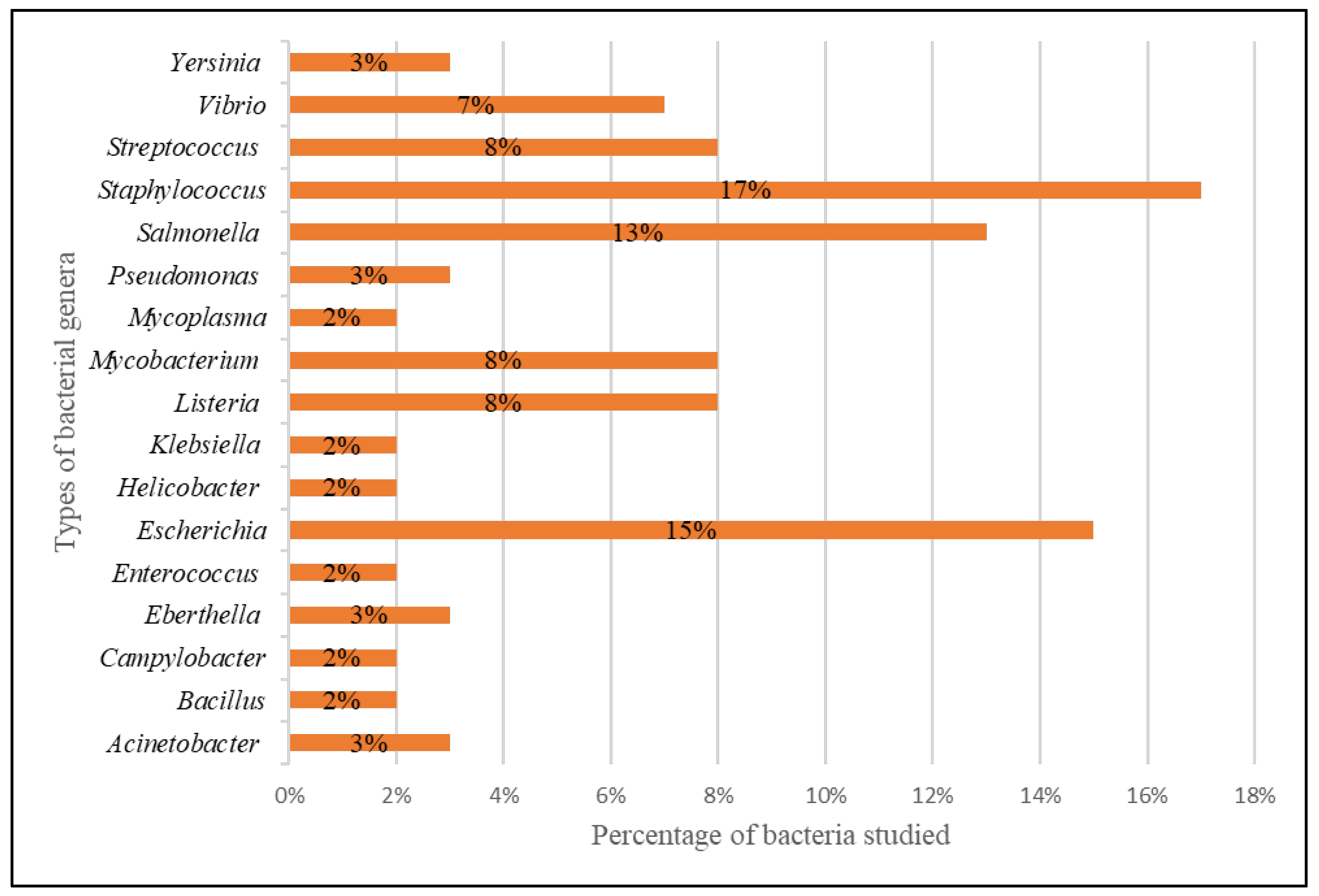
All 46 studies reported the development of diagnostics based on the CRISPR-Cas systems for the detection of harmful bacteria. The diagnostic platform has detected bacteria from 17 different genera, as shown in Figure 3. Most of the bacteria discovered in the retrieved studies came from the genera Staphylococcus (17%), Escherichia (15%), Salmonella (13%), Listeria (8%), Mycobacterium (8%) and Streptococcus (8%).
3.3. Detection Techniques
Table 2 showed a subgroup analysis of CRISPR-Cas-based-bacterial detection. Subgroup analysis based on CRISPR-Cas enzymes showed that Cas12a (64%) is the most commonly employed Cas enzyme in bacterial detection, followed by Cas13a (13%), and Cas9 (11%). Among 46 studies, there are several orthologs of Cas12a (LbCas12a, EnGen® LbaCas12a, LbaCas12a and FnCas12a), Cas13 (LbuCas13a, LwaCas13a and LwCas13a) and Cas9 (dCas9). Cas12b, Cas14a, and dCas9 were utilised in an equal number of studies (4% each). The number of studies that employed trans-cleavage activity of CRISPR-Cas enzymes was higher (83%) and the remaining 17% of the studies used signalling molecules such as Raman reporter (methylene blue) and SYBR Green 1 because these studies predominantly used dCas9 which lacks trans-cleavage activity.
The extraction step is crucial to isolate bacterial genetic materials before amplification and CRISPR-Cas systems-based detection. Eighty-three percent of research employed an extraction process, while only 17% did not. Rather than isolating their genetic components, most of this research used aptamers, which are specific for selected bacteria that were amplified subsequent steps before being detected using the CRISPR-Cas systems. Subgroup analysis regarding the amplification method showed that 66% of the studies used isothermal reactions such as RCA, RPA, LAMP, SDA, EXPAR and RAA followed by PCR (23%) and amplification-free (11%). In terms of detection methods, the majority of research employed fluorescence (67%) based detection rather than lateral flow biosensor (13%), electrochemical biosensor (11%), and others (9%) such as surface-enhanced Raman scattering (SERS), gel electrophoresis, and colorimetric assay.
3.4. Types of Biomarkers
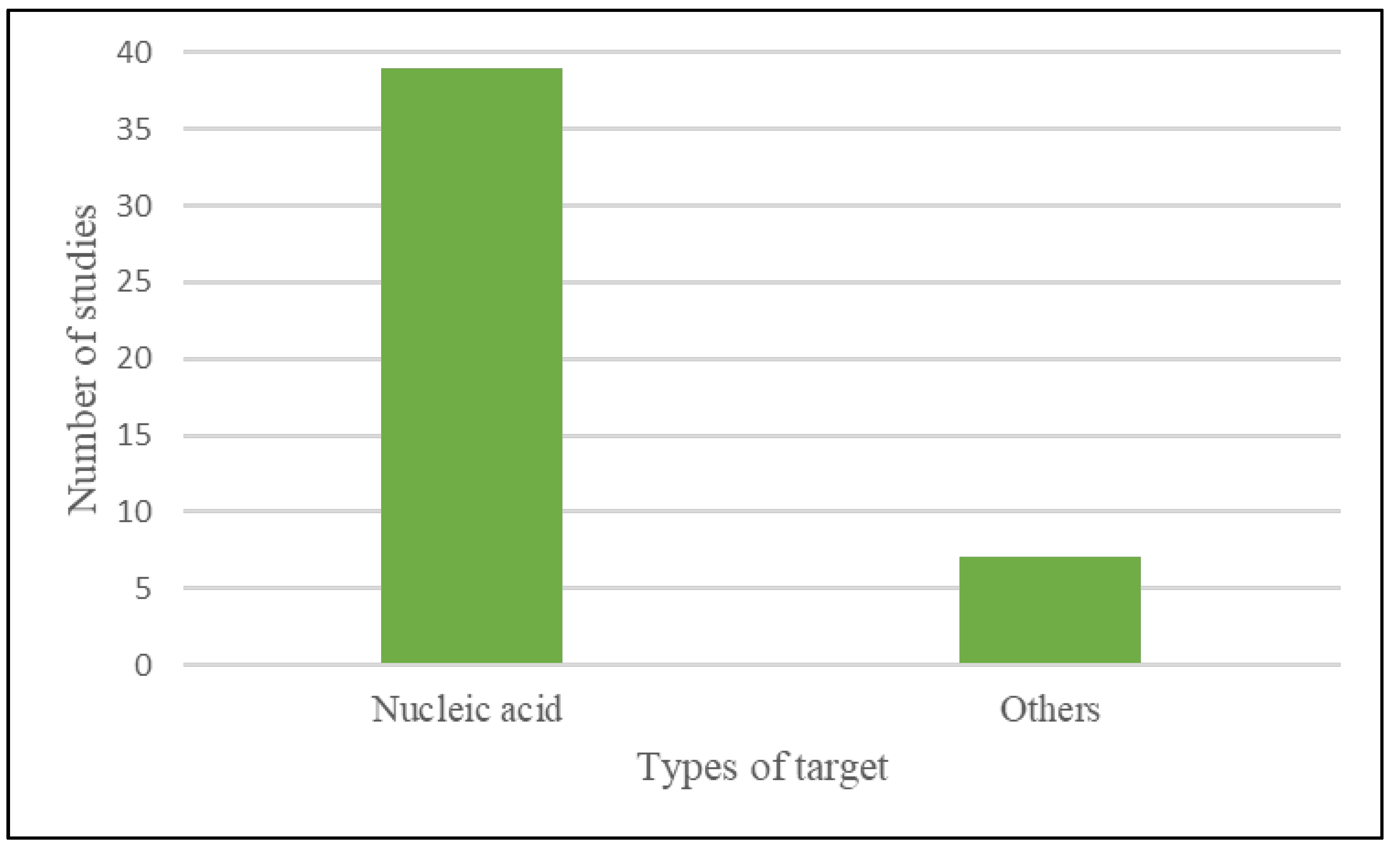
The majority of research (39) employed nucleic acid as a target, while seven used non-nucleic acid such as aptamers and antibodies that are specific for the bacteria being detected as shown in Figure 4.
3.5. Assessment of Study Outcomes
Of the 46 studies, the turnaround times were 30 min to 4 h. The CRISPR-mediated DNA-FISH for the detection of Methicillin-resistant Staphylococcus aureus (MRSA) showed the fastest turnaround times of 30 min [44]. The rapid turnaround time was attributed to the amplification free method utilizing the dCas9 enzyme. Three studies did not report the turnaround times of the assays [30,45,56]. With regards to the analytical sensitivity of the CRISPR-Cas assays, the limit of detection (LoD) was evaluated in three types of concentration which include copies per mL, CFU per mL and molarity. For studies reporting the LoD in copies per mL, detection of Mycobacterium tuberculosis H37Rv utilizing EnGen LbaCas12a reported the lowest LoD of 1 copy/uL [40]. Meanwhile, most of the studies reported the LoD in CFU/mL, of which seven studies showed LoD of 1 CFU/mL. Five studies reported the LoD in molarity [12,17,18,28,38,39]. Of these, CAS-EXPAR for the detection of Listeria monocytogenes showed the lowest LoD up to the attomolar level [38]. Three studies did not report the LoD of the assays [42,48,55]. To determine the LoD of the target, most studies used spiked samples (78%) compared to clinical samples (22%).
4. Discussion
The CRISPR-Cas system has been used in various applications such as gene editing, identification of genotypes and SNPs, detection of antibiotics resistance and virulence genes, and diagnosis of infectious diseases [58,59,60]. Diagnostic techniques based on the CRISPR-Cas systems have recently attracted the attention of researchers due to their excellent accuracy. Three key aspects of the CRISPR-Cas systems contribute to high sensitivity and specificity in the diagnosis of disease, including the detection of bacteria. First, CRISPR– Cas systems identify specific amplicon sequences, distinguish them from amplification byproducts, and cut the sequences (cis-cleavage), a single-turnover method that improves specificity. Second, multiple turnover trans-cleavage activity of Cas12 and Cas13 causes nucleic acid signalling reporters to be cleaved several times, resulting in amplified readout signals for detection and hence improved sensitivity. Third, CRISPR–Cas systems make it easier to generate a variety of readout signals, which broadens their usefulness [61,62].
Bacteria from the genera Staphylococcus (17%), Escherichia (15%), Salmonella (13%), Listeria (8%), Mycobacterium (8%) and Streptococcus (8%) were mostly recognised by the CRISPR-Cas systems. Apart from that, the CRISPR-Cas systems were also utilised to detect bacteria from the genera (i) Vibrio (7%), (ii) Yersinia (3%), (iii) Pseudomonas (3%), (iv) Eberthella (3%), (v) Acinetobacter (3%), (vi) Bacillus (2%), (vii) Campylobacter (2%), (viii) Enterococcus (2%), (ix) Helicobacter (2%), (x) Klebsiella (2%), and (xi) Mycoplasma (2%). The majority of these bacteria are microbes responsible for the most common foodborne infections [63]. These pathogens are also antibiotic resistance bacteria and are on the World Health Organization (WHO)’s priority list for new antibiotic research and development [64,65]. A report by the United States Center for Disease Control and Prevention (CDC) provides an overview of the annual morbidity and mortality of antibiotic-resistant infections in the United States, estimating their number at approximately 2.8 million and the number of deaths associated with these infections at 35,000 [66].
The Cas enzyme is an endonuclease that may be programmed to detect DNA and RNA. CRISPR-Cas systems are divided into two classes (Class 1 and Class 2). Class 1 employs a multi-subunit crRNA-Cas protein, whereas Class 2 employs a single multidomain crRNA-Cas protein [67]. Class 2 is only found in bacteria, and accounts for less than 5% of all known systems. Each class has at least three types as well as several subtypes [68]. In Class 1, there are three types: I, III, and IV. Class 2 enzymes include the II (Cas9), V (Cas12), and VI (Cas13) enzyme classes, as well as subtypes such as V-A (Cas12a or Cpf1), V-B (Cas12b or C2c1), V-C (Cas12c or C2c3), V-F (Cas12f), VI-A (Cas13a or C2c2), VI-B (Cas13b or C2c4), VI-C (Cas13c or C2c7) and VI-D (Cas13d), which have evolved in separate evolutionary paths [69,70].
Of the 46 studies, 30 (64%) employed Cas12a enzymes to identify bacteria, while six (13%) studies used Cas13a, and five (11%) studies used Cas9. Cas12a has two major benefits over other Cas enzymes, namely, it does not require an additional reverse-transcription of amplicons step to detect bacterial DNA as compared to Cas13a [71], and it has trans-cleavage activity to non-specifically cut reporter probes to improve sensitivity, whereas Cas9 can only serve as a nickase to cut dsDNA (cis-cleavage) with no indication of trans-cleavage [62]. According to the Table 1, there are several Cas12a and Cas13a orthologs compared to Cas9, such as LbCas12a, EnGen® LbaCas12a, LbaCas12a (Lachnspiraceace bacterium ND2006), FnCas12a (Francisella novicida U112), LbuCas13a (Leptotrichia buccalis), and LwaCas13a and LwCas13 (Leptotrichia wadei). Cas enzyme orthologs are important because they can recognise other protospacer adjacent motifs (PAM) sequences in addition to typical PAM sequences, allowing for a wider targeting range while maintaining target specificity [72].
Cas12b, Cac12f, and dCas9 were utilised in a similar number of studies (4% each). Recently, researchers have shown interest in a new ortholog Cas12b (AapCas12b) because this enzyme (from Alicyclobacillus acidiphilus) can tolerate high temperature (60 °C) of isothermal reaction (e.g., LAMP) compared to Cas12a, which operates at a lower temperature (e.g., 25–40 °C), and so is incompatible with high-temperature conditions and leads to one-pot assays [73,74]. As well as AapCas12b, BrCas12b from (thermophile bacterium Brevibacillus sp. SYSU G02855) is also capable of binding and cleaving target DNA at high temperatures, making it a good candidate for diagnostic development [75]. Apart from Cas12b, Cas14a (Cas12f) is becoming more widely utilised in the diagnostic field because it demands full complementarity in the seed region of sgRNA, a trait that is important for obtaining single nucleotide specificity [76]. Cas12f, a type V effector protein, was previously known as Cas14. Cas14 is similar to Cas12 in that it can also target dsDNA and is dependent on T-rich PAM, hence it has been classed into the Cas12 family (Karvelis et al., 2020). Cas9 from Streptococcus pyogenes (SpCas9) is one of the simplest systems, drawing a lot of attention for its gene-editing capabilities [77]. However, in recent years, it has become a good bio-recognition element after being modified in a deactivated form (dCas9) resulting in an “antibody-like” mechanism. Researchers created a dCas9 by introducing two-point mutations, H840A and D10A, into the HNH and RuvC nuclease domains (dCas9). DNA cleavage activity is absent in dCas9, but DNA binding activity is unaffected [78].
Furthermore, Class 2 Cas enzyme selection is based on the enzymes’ properties. Cas9 (type II) has two nuclease domains, HNH and RuvC, which each cleaves one strand of double-stranded DNA (dsDNA) [79]. Cas12 (type V) has only one RuvC domain that cleaves dsDNA and single-stranded DNA (ssDNA) in the presence of cation ions such as magnesium and calcium ions (Mg2+ and Ca2+) [80]. Cas13 (type VI) has two predicted higher eukaryotic and prokaryotic nucleotide (HEPN) domains to cut single-stranded RNA (ssRNA) [81]. Cas9 requires both tracrRNA and crRNA, whereas Cas12 (except for Cas12b and Cas12c) and Cas13 utilise crRNA only. This is because Cas12 and Cas13 can cleave crRNA arrays to produce their crRNAs (self-processing) [81,82].
Cas9 recognise a specific PAM sequence (5′ NGG 3′) (N represents any nucleotide) in a non-target DNA strand, distant 10–12 nucleotides apart from the PAM sequence [79]. Cas12 identifies a 5′-T-rich PAM at the distal end. The PAM sequence required for Cas12 to bind dsDNA induces the catalytic activation of a crRNA-complementary dsDNA, but not of a crRNA-complementary ssDNA [83]. Cas13a identifies a 3′ end non-G protospacer flanking site (PFS) while 5′ end non-C PFS for Cas13b [84,85]. Moreover, Cas9 generate a blunt dsDNA break while Cas12 and Cas13 generate sticky and near U and A break respectively [80,86]. In addition, Cas9 does not exhibit trans-cleavage activity, but Cas12 and Cas13 do, allowing for powerful signal amplification [87,88]. These discoveries result in the development of the DETECTR (DNA endonuclease targeted CRISPR trans reporter) and SHERLOCK (Specific High Sensitivity Enzymatic Reporter UnLOCKing) diagnostic platforms for Cas12 and Cas13 respectively [89,90]. Trans-cleavage activity by Cas12a and Cas13 were used in 83% of studies, while the remaining 17% of studies mostly used Cas9, which lacks trans-cleavage activity and hence used other signalling molecules.
Extracting bacterial genetic materials from samples necessitates determining the appropriate nucleic acid sequence (DNA or RNA) [91]. It is an important preanalytical stage in the development of any successful molecular diagnostic procedure, ensuring a reliable result. The extraction method was first used in 83% of studies before continuing to amplify and before CRISPR-Cas system-based detection. Extraction entails lysing the cells, purifying the nucleic acid to remove extraneous cell components, inhibiting compounds, degrading enzymes, and recovering the necessary nucleic acid [92,93]. The commercially available kits are also used for DNA extraction.
To improve sensitivity, the CRISPR-Cas system needs to be combined with a target nucleic acid amplification step performed by either PCR or isothermal technologies. This allows for the enrichment of rare and low-abundance nucleic acid targets and for the depletion of unwanted abundant nucleic acids. PCR is used in terms of targeted amplification of desired sequences, but it has several disadvantages, including the need for large equipment and trained personnel to operate it [94]. Isothermal amplification technologies address these constraints. CRISPR–Cas9 systems are effective for creating isothermal exponential amplification strategies because they can unwind dsDNA to ssDNA at a moderate temperature (37 °C) [61]. Isothermal methods were used in 66% of the investigations, while PCR methods were used in 23% of the studies. Among isothermal methods, RPA has been widely used in CRISPR-Cas system based bacterial detection followed by LAMP and SDA. RPA is notable for its ease of use, high sensitivity, selectivity, compatibility with multiplexing, exceptionally rapid amplification (20–60 min), and ability to operate at a low (37–42 °C) and constant temperature without the requirement for an initial denaturation phase or numerous primers [59,95]. The amplification technique, on the other hand, is not only time consuming but also poses a risk of aerosol contamination. Several research groups have worked hard to produce amplification-free CRISPR-Cas systems and investigate their use in pathogen identification. Only five (11%) of the 46 research studies found used an amplification-free CRISPR-Cas system-based bacterial detection.
Fluorescence detection (67%), lateral flow biosensor (13%), and electrochemical biosensor (11%) are three key detection approaches employed in CRISPR-Cas systems-based bacteria detection. Fluorescence-based sensing has several advantages, including background-free sensing, which dramatically improves the signal-to-noise ratio when compared to other optical approaches, but which necessitates the use of instruments (fluorescence reader, either portable or not, and a real-time PCR machine) [96]. The main benefit of LFAs is that their results are simple to interpret and can be read with the naked eye without the use of expensive instruments. LFAs, on the other hand, are inefficient and inaccurate, making these traditional paper-based platforms unsuitable for the higher quantitative analyses required in clinical applications [97]. Biosensors have several advantages over traditional analytical techniques, including excellent selectivity and sensitivity, the potential for miniaturization and portability, low cost, real-time detection, small sample volumes, and quick reaction [98].
5. Conclusions
This review detailed and analysed several key points, including the origin of CRISPR-Cas systems, the types and properties of CRISPR-Cas enzymes, extraction methods, amplification techniques, detection methods, assay time and LoD. CRISPR-Cas systems were first used to modify genes and are now being used to diagnose infectious diseases. Cas12, Cas13, and Cas9 are three types of CRISPR-Cas enzymes employed in bacterial detection. Nucleic acid extraction prior to isothermal amplification, and fluorescence detection steps after CRISPR-Cas were used in most identified studies in this review. The assay takes about 4 h and obtains various LoD using spiked samples. This review reveals that only 20% of the studies reported on the clinical accuracy of the CRISPR-Cas system in bacterial detection. As a result, we recommend that the development and evaluation of the CRISPR-Cas system should be conducted using clinical samples. It is suggested that more studies will focus on the development of nucleic acid amplification-free CRISPR-Cas systems for bacterial detection toward the fulfilment of the ASSURED criteria.
Author Contributions
Conceptualization, I.A. and M.O.; validation, I.A. and M.O.; writing—original draft preparation, K.S. and M.A.N.; writing—review and editing, I.A., M.O., M.A.N., M.F.K. and K.S.; visualization, I.A. and M.A.N.; supervision, I.A. and M.O.; project administration, I.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Higher Institution Centre of Excellence (HICoE), Ministry of Higher Education, Malaysia (311/CIPPM/4401005).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kootallur, B.N.; Thangavelu, C.P.; Mani, M. Bacterial identification in the diagnostic laboratory: How much is enough? Indian J. Med. Microbiol. 2011, 29, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Figdor, D.; Gulabivala, K. Survival against the odds: Microbiology of root canals associated with post-treatment disease. Endod. Top. 2008, 18, 62–77. [Google Scholar] [CrossRef]
- Bursle, E.; Robson, J. Non-culture methods for detecting infection. Aust. Prescr. 2016, 39, 171–175. [Google Scholar] [CrossRef]
- Caliendo, A.M.; Gilbert, D.N.; Ginocchio, C.C.; Hanson, K.E.; May, L.; Quinn, T.C.; Tenover, F.C.; Alland, D.; Blaschke, A.J.; Bonomo, R.A.; et al. Better tests, better care: Improved diagnostics for infectious diseases. Clin. Infect. Dis. 2013, 57 (Suppl. 3), S139–S170. [Google Scholar] [CrossRef] [Green Version]
- Kosack, C.S.; Page, A.-L.; Klatser, P.R. A guide to aid the selection of diagnostic tests. Bull. World Health Organ. 2017, 95, 639–645. [Google Scholar] [CrossRef]
- Leon, L.M.; Mendoza, S.D.; Bondy-Denomy, J. How bacteria control the CRISPR-Cas arsenal. Curr. Opin. Microbiol. 2018, 42, 87–95. [Google Scholar] [CrossRef]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and classification of the CRISPR-Cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Munn, Z.; Peters, M.D.J.; Stern, C.; Tufanaru, C.; McArthur, A.; Aromataris, E. Systematic review or scoping review? Guidance for authors when choosing between a systematic or scoping review approach. BMC Med. Res. Methodol. 2018, 18, 143. [Google Scholar] [CrossRef]
- Bonini, A.; Poma, N.; Vivaldi, F.; Biagini, D.; Bottai, D.; Tavanti, A.; Di Francesco, F. A label-free impedance biosensing assay based on CRISPR/Cas12a collateral activity for bacterial DNA detection. J. Pharm. Biomed. Anal. 2021, 204, 114268. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, Y.; Zhang, L.; Liu, S.; Zhang, M.; Wang, J.; Ning, B.; Peng, Y.; He, J.; Hu, Y.; et al. CRISPR-Cas9 Triggered Two-Step Isothermal Amplification Method for E. coli O157:H7 Detection Based on a Metal–Organic Framework Platform. Anal. Chem. 2020, 92, 3032–3041. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.-W.; Zhou, X.; Xu, T.; Yang, M.; Chen, Y.; He, G.-Q.; Pan, N.; Cai, Y.; Li, Y.; Wang, X.; et al. CRISPR-based rapid and ultra-sensitive diagnostic test for Mycobacterium tuberculosis. Emerg. Microbes Infect. 2019, 8, 1361–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Zhou, X.; Shan, Y.; Yue, H.; Huang, R.; Hu, J.; Xing, D. Sensitive detection of a bacterial pathogen using allosteric probe-initiated catalysis and CRISPR-Cas13a amplification reaction. Nat. Commun. 2020, 11, 267. [Google Scholar] [CrossRef] [Green Version]
- Sam, I.K.; Chen, Y.-Y.; Ma, J.; Li, S.-Y.; Ying, R.-Y.; Li, L.-X.; Ji, P.; Wang, S.-J.; Xu, J.; Bao, Y.-J.; et al. TB-QUICK: CRISPR-Cas12b-assisted rapid and sensitive detection of Mycobacterium tuberculosis. J. Infect. 2021, 83, 54–60. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, X.; Cai, Z.; Dong, X.; Chen, G.; Li, Z.; Qiu, L.; He, L.; Liang, B.; Liu, X.; et al. An Isothermal Method for Sensitive Detection of Mycobacterium tuberculosis Complex Using Clustered Regularly Interspaced Short Palindromic Repeats/Cas12a Cis and Trans Cleavage. J. Mol. Diagn. 2020, 22, 1020–1029. [Google Scholar] [CrossRef]
- Kim, H.; Lee, S.; Seo, H.W.; Kang, B.; Moon, J.; Lee, K.G.; Yong, D.; Kang, H.; Jung, J.; Lim, E.-K.; et al. Clustered Regularly Interspaced Short Palindromic Repeats-Mediated Surface-Enhanced Raman Scattering Assay for Multidrug-Resistant Bacteria. ACS Nano 2020, 14, 17241–17253. [Google Scholar] [CrossRef]
- Qiu, E.; Jin, S.; Xiao, Z.; Chen, Q.; Wang, Q.; Liu, H.; Xie, C.; Chen, C.; Li, Z.; Han, S. CRISPR-based detection of Helicobacter pylori in stool samples. Helicobacter 2021, 26, e12828. [Google Scholar] [CrossRef]
- Wei, J. Accurate and sensitive analysis of Staphylococcus aureus through CRISPR-Cas12a based recycling signal amplification cascades for early diagnosis of skin and soft tissue infections. J. Microbiol. Methods 2021, 183, 106167. [Google Scholar] [CrossRef]
- Jiang, L.; Zeng, W.; Wu, W.; Deng, Y.; He, F.; Liang, W.; Huang, M.; Huang, H.; Li, Y.; Wang, X.; et al. Development and Clinical Evaluation of a CRISPR-Based Diagnostic for Rapid Group B Streptococcus Screening. Emerg. Infect. Dis. 2021, 27, 2379–2388. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Wang, R.; Qiao, Z.; Yang, W. Single-digit Salmonella detection with the naked eye using bio-barcode immunoassay coupled with recombinase polymerase amplification and a CRISPR-Cas12a system. Analyst 2021, 146, 5271–5279. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ke, Y.; Liu, W.; Sun, Y.; Ding, X. A One-Pot Toolbox Based on Cas12a/crRNA Enables Rapid Foodborne Pathogen Detection at Attomolar Level. ACS Sens. 2020, 5, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, Y.; Shi, Y.; Wang, L.; Zhang, M.; Wu, J.; Chen, H. Carrying out pseudo dual nucleic acid detection from sample to visual result in a polypropylene bag with CRISPR/Cas12a. Biosens. Bioelectron. 2021, 178, 113001. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Zeng, H.; Liu, X.; Jiang, W.; Wang, Y.; Ouyang, W.; Tang, X. RPA-Cas12a-FS: A frontline nucleic acid rapid detection system for food safety based on CRISPR-Cas12a combined with recombinase polymerase amplification. Food Chem. 2021, 334, 127608. [Google Scholar] [CrossRef]
- Wang, T.; Liu, Y.; Sun, H.-H.; Yin, B.-C.; Ye, B.-C. An RNA-Guided Cas9 Nickase-Based Method for Universal Isothermal DNA Amplification. Angew. Chem. Int. Ed. 2019, 58, 5382–5386. [Google Scholar] [CrossRef]
- Mukama, O.; Wu, J.; Li, Z.; Liang, Q.; Yi, Z.; Lu, X.; Liu, Y.; Liu, Y.; Hussain, M.; Makafe, G.G.; et al. An ultrasensitive and specific point-of-care CRISPR/Cas12 based lateral flow biosensor for the rapid detection of nucleic acids. Biosens. Bioelectron. 2020, 159, 112143. [Google Scholar] [CrossRef]
- Chen, G.; Lyu, Y.; Wang, D.; Zhu, L.; Cao, S.; Pan, C.; Feng, E.; Zhang, W.; Liu, X.; Cui, Y.; et al. Obtaining Specific Sequence Tags for Yersinia pestis and Visually Detecting Them Using the CRISPR-Cas12a System. Pathogens 2021, 10, 562. [Google Scholar] [CrossRef]
- Song, F.; Wei, Y.; Wang, P.; Ge, X.; Li, C.; Wang, A.; Yang, Z.; Wan, Y.; Li, J. Combining tag-specific primer extension and magneto-DNA system for Cas14a-based universal bacterial diagnostic platform. Biosens. Bioelectron. 2021, 185, 113262. [Google Scholar] [CrossRef]
- Chen, Z.; Ma, L.; Bu, S.; Zhang, W.; Chen, J.; Li, Z.; Hao, Z.; Wan, J. CRISPR/Cas12a and immuno-RCA based electrochemical biosensor for detecting pathogenic bacteria. J. Electroanal. Chem. 2021, 901, 115755. [Google Scholar] [CrossRef]
- Bu, S.; Liu, X.; Wang, Z.; Wei, H.; Yu, S.; Li, Z.; Hao, Z.; Liu, W.; Wan, J. Ultrasensitive detection of pathogenic bacteria by CRISPR/Cas12a coupling with a primer exchange reaction. Sens. Actuators B Chem. 2021, 347, 130630. [Google Scholar] [CrossRef]
- Yin, L.; Duan, N.; Chen, S.; Yao, Y.; Liu, J.; Ma, L. Ultrasensitive pathogenic bacteria detection by a smartphone-read G-quadruplex-based CRISPR-Cas12a bioassay. Sens. Actuators B Chem. 2021, 347, 130586. [Google Scholar] [CrossRef]
- Huang, Y.; Gu, D.; Xue, H.; Yu, J.; Tang, Y.; Huang, J.; Zhang, Y.; Jiao, X. Rapid and Accurate Campylobacter jejuni Detection With CRISPR-Cas12b Based on Newly Identified Campylobacter jejuni-Specific and -Conserved Genomic Signatures. Front. Microbiol. 2021, 12, 802. [Google Scholar] [CrossRef]
- Sheng, A.; Wang, P.; Yang, J.; Tang, L.; Chen, F.; Zhang, J. MXene Coupled with CRISPR-Cas12a for Analysis of Endotoxin and Bacteria. Anal. Chem. 2021, 93, 4676–4681. [Google Scholar] [CrossRef]
- Wang, L.; Shen, X.; Wang, T.; Chen, P.; Qi, N.; Yin, B.-C.; Ye, B.-C. A lateral flow strip combined with Cas9 nickase-triggered amplification reaction for dual food-borne pathogen detection. Biosens. Bioelectron. 2020, 165, 112364. [Google Scholar] [CrossRef]
- Peng, L.; Zhou, J.; Yin, L.; Man, S.; Ma, L. Integration of logic gates to CRISPR/Cas12a system for rapid and sensitive detection of pathogenic bacterial genes. Anal. Chim. Acta 2020, 1125, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xiong, E.; Tian, T.; Cheng, M.; Lin, W.; Wang, H.; Zhang, G.; Sun, J.; Zhou, X. Clustered Regularly Interspaced Short Palindromic Repeats/Cas9-Mediated Lateral Flow Nucleic Acid Assay. ACS Nano 2020, 14, 2497–2508. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Zhou, X.; Wang, H.; Xing, D. Clustered Regularly Interspaced Short Palindromic Repeats/Cas9 Triggered Isothermal Amplification for Site-Specific Nucleic Acid Detection. Anal. Chem. 2018, 90, 2193–2200. [Google Scholar] [CrossRef]
- Suea-Ngam, A.; Howes, P.D.; deMello, A.J. An amplification-free ultra-sensitive electrochemical CRISPR/Cas biosensor for drug-resistant bacteria detection. Chem. Sci. 2021, 12, 12733–12743. [Google Scholar] [CrossRef]
- Li, H.; Cui, X.; Sun, L.; Deng, X.; Liu, S.; Zou, X.; Li, B.; Wang, C.; Wang, Y.; Liu, Y.; et al. High concentration of Cas12a effector tolerates more mismatches on ssDNA. FASEB J. 2021, 35, e21153. [Google Scholar] [CrossRef]
- Gao, S.; Liu, J.; Li, Z.; Ma, Y.; Wang, J. Sensitive detection of foodborne pathogens based on CRISPR-Cas13a. J. Food Sci. 2021, 86, 2615–2625. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Zhang, S.; Liang, Z.; Li, G.; Fang, M.; Liu, Y.; Zhang, J.; Ou, M.; He, X.; Zhang, T.; et al. Identification of Mycobacterium abscessus species and subspecies using the Cas12a/sgRNA-based nucleic acid detection platform. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 551–558. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.; Zhang, L.; Yang, Y.; Yang, S.; Yang, L.; Chen, H.; Liu, C.; Li, J.; Xie, G. Integration of multiplex PCR and CRISPR-Cas allows highly specific detection of multidrug-resistant Acinetobacter Baumannii. Sens. Actuators B Chem. 2021, 334, 129600. [Google Scholar] [CrossRef]
- Guk, K.; Keem, J.O.; Hwang, S.G.; Kim, H.; Kang, T.; Lim, E.-K.; Jung, J. A facile, rapid and sensitive detection of MRSA using a CRISPR-mediated DNA FISH method, antibody-like dCas9/sgRNA complex. Biosens. Bioelectron. 2017, 95, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhou, W.; Lin, X.; Khan, M.R.; Deng, S.; Zhou, M.; He, G.; Wu, C.; Deng, R.; He, Q. Light-up RNA aptamer signaling-CRISPR-Cas13a-based mix-and-read assays for profiling viable pathogenic bacteria. Biosens. Bioelectron. 2021, 176, 112906. [Google Scholar] [CrossRef]
- Zhou, J.; Yin, L.; Dong, Y.; Peng, L.; Liu, G.; Man, S.; Ma, L. CRISPR-Cas13a based bacterial detection platform: Sensing pathogen Staphylococcus aureus in food samples. Anal. Chim. Acta 2020, 1127, 225–233. [Google Scholar] [CrossRef]
- Ma, L.; Peng, L.; Yin, L.; Liu, G.; Man, S. CRISPR-Cas12a-Powered Dual-Mode Biosensor for Ultrasensitive and Cross-Validating Detection of Pathogenic Bacteria. ACS Sens. 2021, 6, 2920–2927. [Google Scholar] [CrossRef]
- Xu, L.; Dai, Q.; Shi, Z.; Liu, X.; Gao, L.; Wang, Z.; Zhu, X.; Li, Z. Accurate MRSA identification through dual-functional aptamer and CRISPR-Cas12a assisted rolling circle amplification. J. Microbiol. Methods 2020, 173, 105917. [Google Scholar] [CrossRef]
- Schultzhaus, Z.; Wang, Z.; Stenger, D. Systematic analysis, identification, and use of CRISPR/Cas13a–associated crRNAs for sensitive and specific detection of the lcrV gene of Yersinia pestis. Diagn. Microbiol. Infect. Dis. 2021, 99, 115275. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, C.; Shi, Y.; Wu, J.; Wu, J.; Chen, H. Selective endpoint visualized detection of Vibrio parahaemolyticus with CRISPR/Cas12a assisted PCR using thermal cycler for on-site application. Talanta 2020, 214, 120818. [Google Scholar] [CrossRef]
- You, Y.; Zhang, P.; Wu, G.; Tan, Y.; Zhao, Y.; Cao, S.; Song, Y.; Yang, R.; Du, Z. Highly Specific and Sensitive Detection of Yersinia pestis by Portable Cas12a-UPTLFA Platform. Front. Microbiol. 2021, 12, 1702. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ye, Q.; Chen, M.; Zhou, B.; Zhang, J.; Pang, R.; Xue, L.; Wang, J.; Zeng, H.; Wu, S.; et al. An ultrasensitive CRISPR/Cas12a based electrochemical biosensor for Listeria monocytogenes detection. Biosens. Bioelectron. 2021, 179, 113073. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ye, Q.; Chen, M.; Xiang, X.; Zhang, J.; Pang, R.; Xue, L.; Wang, J.; Gu, Q.; Lei, T.; et al. Cas12aFDet: A CRISPR/Cas12a-based fluorescence platform for sensitive and specific detection of Listeria monocytogenes serotype 4c. Anal. Chim. Acta 2021, 1151, 338248. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, Y.; Yang, Q.; Peng, C.; Wang, X.; Zhang, M.; Qian, S.; Xu, J.; Wu, J. A reversible valve-assisted chip coupling with integrated sample treatment and CRISPR/Cas12a for visual detection of Vibrio parahaemolyticus. Biosens. Bioelectron. 2021, 188, 113352. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, R.; Wang, D.; Wu, J.; Li, J.; Wang, J.; Liu, H.; Wang, Y. Cas12aVDet: A CRISPR/Cas12a-Based Platform for Rapid and Visual Nucleic Acid Detection. Anal. Chem. 2019, 91, 12156–12161. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Meng, T.; Tan, X.; Wei, Y.; Tao, Z.; Yang, Z.; Song, F.; Wang, P.; Wan, Y. Cas14a1-mediated nucleic acid detectifon platform for pathogens. Biosens. Bioelectron. 2021, 189, 113350. [Google Scholar] [CrossRef]
- Xiao, X.; Lin, Z.; Huang, X.; Lu, J.; Zhou, Y.; Zheng, L.; Lou, Y. Rapid and Sensitive Detection of Vibrio vulnificus Using CRISPR/Cas12a Combined With a Recombinase-Aided Amplification Assay. Front. Microbiol. 2021, 12, 3251. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Battalapalli, D.; Hakeem, M.J.; Selamneni, V.; Zhang, P.; Draz, M.S.; Ruan, Z. Engineered CRISPR-Cas systems for the detection and control of antibiotic-resistant infections. J. Nanobiotechnol. 2021, 19, 401. [Google Scholar] [CrossRef]
- Kaminski, M.M.; Abudayyeh, O.O.; Gootenberg, J.S.; Zhang, F.; Collins, J.J. CRISPR-based diagnostics. Nat. Biomed. Eng. 2021, 5, 643–656. [Google Scholar] [CrossRef]
- Hu, Y.; Li, W. Development and Application of CRISPR-Cas Based Tools. Front. Cell Dev. Biol. 2022, 10, 834646. [Google Scholar] [CrossRef]
- Feng, W.; Newbigging, A.M.; Tao, J.; Cao, Y.; Peng, H.; Le, C.; Wu, J.; Pang, B.; Li, J.; Tyrrell, D.L.; et al. CRISPR technology incorporating amplification strategies: Molecular assays for nucleic acids, proteins, and small molecules. Chem. Sci. 2021, 12, 4683–4698. [Google Scholar] [CrossRef]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529–563. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [Green Version]
- CDC. Antibiotic Resistance Threats in the United States; CDC: Atlanta, GA, USA, 2019. [Google Scholar]
- Nidhi, S.; Anand, U.; Oleksak, P.; Tripathi, P.; Lal, J.A.; Thomas, G.; Kuca, K.; Tripathi, V. Novel CRISPR-Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives. Int. J. Mol. Sci. 2021, 22, 3327. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR–Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shmakov, S.; Abudayyeh, O.O.; Makarova, K.S.; Wolf, Y.I.; Gootenberg, J.S.; Semenova, E.; Minakhin, L.; Joung, J.; Konermann, S.; Severinov, K.; et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. Mol. Cell 2015, 60, 385–397. [Google Scholar] [CrossRef] [Green Version]
- Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; et al. Diversity and evolution of class 2 CRISPR-Cas systems. Nat. Rev. Microbiol. 2017, 15, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Kellner, M.J.; Koob, J.G.; Gootenberg, J.S.; Abudayyeh, O.O.; Zhang, F. SHERLOCK: Nucleic acid detection with CRISPR nucleases. Nat. Protoc. 2019, 14, 2986–3012. [Google Scholar] [CrossRef]
- Kleinstiver, B.P.; Sousa, A.A.; Walton, R.T.; Tak, Y.E.; Hsu, J.Y.; Clement, K.; Welch, M.M.; Horng, J.E.; Malagon-Lopez, J.; Scarfò, I.; et al. Engineered CRISPR-Cas12a variants with increased activities and improved targeting ranges for gene, epigenetic and base editing. Nat. Biotechnol. 2019, 37, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, S.; Wu, N.; Wu, J.; Wang, G.; Zhao, G.; Wang, J. HOLMESv2: A CRISPR-Cas12b-Assisted Platform for Nucleic Acid Detection and DNA Methylation Quantitation. ACS Synth. Biol. 2019, 8, 2228–2237. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-mediated isothermal amplification (LAMP): Principle, features, and future prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Liu, R.-R.; Xian, W.-D.; Xiong, M.; Xiao, M.; Li, W.-J. A novel thermal Cas12b from a hot spring bacterium with high target mismatch tolerance and robust DNA cleavage efficiency. Int. J. Biol. Macromol. 2020, 147, 376–384. [Google Scholar] [CrossRef]
- Harrington, L.B.; Burstein, D.; Chen, J.S.; Paez-Espino, D.; Ma, E.; Witte, I.P.; Cofsky, J.C.; Kyrpides, N.C.; Banfield, J.F.; Doudna, J.A. Programmed DNA destruction by miniature CRISPR-Cas14 enzymes. Science 2018, 362, 839–842. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna Jennifer, A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [Green Version]
- Garneau, J.E.; Dupuis, M.-È.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Fremaux, C.; Horvath, P.; Magadán, A.H.; Moineau, S. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [Green Version]
- Pickar-Oliver, A.; Gersbach, C.A. The next generation of CRISPR–Cas technologies and applications. Nat. Rev. Mol. Cell Biol. 2019, 20, 490–507. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Y.; Cheng, Q.-X.; Liu, J.-K.; Nie, X.-Q.; Zhao, G.-P.; Wang, J. CRISPR-Cas12a has both cis- and trans-cleavage activities on single-stranded DNA. Cell Res. 2018, 28, 491–493. [Google Scholar] [CrossRef] [PubMed]
- Anantharaman, V.; Makarova, K.S.; Burroughs, A.M.; Koonin, E.V.; Aravind, L. Comprehensive analysis of the HEPN superfamily: Identification of novel roles in intra-genomic conflicts, defense, pathogenesis and RNA processing. Biol. Direct 2013, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Li, X.; Wang, J.; Wang, M.; Chen, P.; Yin, M.; Li, J.; Sheng, G.; Wang, Y. Two Distant Catalytic Sites Are Responsible for C2c2 RNase Activities. Cell 2017, 168, 121–134.e12. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Gao, P.; Rajashankar, K.R.; Patel, D.J. PAM-Dependent Target DNA Recognition and Cleavage by C2c1 CRISPR-Cas Endonuclease. Cell 2016, 167, 1814–1828.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- East-Seletsky, A.; O’Connell, M.R.; Knight, S.C.; Burstein, D.; Cate, J.H.D.; Tjian, R.; Doudna, J.A. Two distinct RNase activities of CRISPR-C2c2 enable guide-RNA processing and RNA detection. Nature 2016, 538, 270–273. [Google Scholar] [CrossRef]
- Sashital, D.G. Pathogen detection in the CRISPR-Cas era. Genome Med. 2018, 10, 32. [Google Scholar] [CrossRef]
- Hatoum-Aslan, A. CRISPR Methods for Nucleic Acid Detection Herald the Future of Molecular Diagnostics. Clin. Chem. 2018, 64, 1681–1683. [Google Scholar] [CrossRef] [Green Version]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Rampazzo, R.d.C.P.; Costa, A.D.T.; Krieger, M.A. Current Nucleic Acid Extraction Methods and Their Implications to Point-of-Care Diagnostics. BioMed Res. Int. 2017, 2017, 9306564. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.H. Nucleic Acid Extraction Techniques. In Advanced Techniques in Diagnostic Microbiology; Tang, Y.W., Stratton, C., Eds.; Springer: Boston, MA, USA, 2013. [Google Scholar] [CrossRef]
- Gupta, N. DNA Extraction and Polymerase Chain Reaction. J. Cytol. 2019, 36, 116–117. [Google Scholar] [CrossRef] [PubMed]
- Scheler, O.; Glynn, B.; Kurg, A. Nucleic acid detection technologies and marker molecules in bacterial diagnostics. Expert Rev. Mol. Diagn. 2014, 14, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trends Anal. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, J.E.; Berendsen, J.T.W.; Steenbergen, R.D.M.; Wolthuis, R.M.F.; Eijkel, J.C.T.; Segerink, L.I. Point-of-care CRISPR/Cas nucleic acid detection: Recent advances, challenges and opportunities. Biosens. Bioelectron. 2020, 166, 112445. [Google Scholar] [CrossRef] [PubMed]
- Kasetsirikul, S.; Shiddiky, M.J.A.; Nguyen, N.-T. Challenges and perspectives in the development of paper-based lateral flow assays. Microfluid. Nano Fluid. 2020, 24, 17. [Google Scholar] [CrossRef]
- Nambiar, S.; Yeow, J.T.W. Conductive polymer-based sensors for biomedical applications. Biosens. Bioelectron. 2011, 26, 1825–1832. [Google Scholar] [CrossRef]
Figure 1.
PRISMA-ScR flow diagram showing the process of selecting studies.

Figure 2.
Quality assessment of the retrieved studies.

Figure 3.
Bacteria from various genera were detected via CRISPR-Cas systems.

Figure 4.
Types of targets used in CRISPR-Cas system-based bacterial detection.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the included studies.
| No | CRISPR-Cas Enzymes | Name of Methods | Types of Bacteria | Trans Cutting | Reporter Probes | Amplification Methods | Extraction | Samples (Types & n) | Detection Methods | Requirements of Instruments | Targets | Assay Time (min) | LOD/ Detection Range | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | LbaCas12a | NR | Escherichia coli & Staphylococcus aureus | Yes | ssDNA | PCR | Yes | Reference laboratory strains & panel of clinical isolates | EB | Thermocycler & impedance analyser | mdh & nuc | 90 | 3 nM | [12] |
| 2 | Cas9 Cis (dsDNA) | NR | Escherichia coli O157:H7 | No | ssDNA with a metal-organic framework (MOF) [UiO66] | SDA & RCA | Yes | Spiked spring water, skim milk & orange juice | F | Thermocycler & fluorescence spectrophotometer | hlyA | 120 | 40 CFU/mL | [13] |
| 3 | Cas12a | CRISPR-MTB | Mycobacterium tuberculosis H37Ra strain | Yes | ssDNA | RPA | Yes | Pulmonary samples: sputum, & BALF and extrapulmonary samples: cerebrospinal fluid, pleural fluid, ascites, pus, pericardial effusion, urine & synovial fluid (n = 179) | F | qPCR machine | IS6110 | 90 | 50 CFU/mL | [14] |
| 4 | LbuCas13a | APC-Cas | Salmonella enteritidis | Yes | ssRNA | SDA & reverse transcription | No | Spiked milk and drinking water | F | Real-time PCR machine | Aptamer SE-3 against live Salmonella Enteritidis | 140 | 1 CFU | [15] |
| 5 | AacCas12b | TB-QUICK | Mycobacterium tuberculosis H37Ra strain | Yes | ssDNA | LAMP | Yes | Sputum, BALF & EDTA anticoagulant plasma (n = 147) | F | Real-Time PCR machine | IS6110 | 120 | 1.3 copy/μL | [16] |
| 6 | LbCas12a | NR | Mycobacterium tuberculosis | Yes | ssDNA | RPA | Yes | Sputum (n = 193) | F | Real-Time PCR machine | IS1081 | 240 | 4.48 fmol/L | [17] |
| 7 | dCas9 | NR | Staphylococcus aureus, Acinetobacter baumannii & Klebsiella pneumoniae | No | Raman reporter: MB | No | Yes | Mice lung, spleen & liver tissues | SERS | Raman spectrometer | spa, pgi & uge | 60 | 14.1 fM, 9.7 fM & 8.1 fM | [18] |
| 8 | Cas12a | CRISPR-HP | Helicobacter pylori | Yes | ssDNA | RPA | Yes | Stool (n = 41) | LFB | No | Genomic DNA | 60 | 5 copies/µL | [19] |
| 9 | EnGen® LbaCas12a | NR | Methicillin-resistant Staphylococcus aureus (MRSA) | Yes | ssDNA | SDA | No | Reference laboratory strains | F | Fluorescence spectrophotometer | Aptamer against PBP2a | ~180 | 106 to 102 CFU/ mL (detection range) | [20] |
| 10 | Cas13a | CRISPR-GBS | Streptococcus agalactiae | Yes | ssRNA | RPA | Yes | Vaginal–rectal swabs (n = 412) | F | qPCR machine | atoB | 90 | 50 CFU/mL | [21] |
| 11 | EnGen® LbaCas12a | BCA–RPA–Cas12a | Salmonella typhimurium | Yes | ssDNA | RPA | No | Spiked milk | F | Blue light | Antibodies specific against Salmonella Typhimurium | 60 | 1 CFU/mL | [22] |
| 12 | LbCas12a | OCTOPUS | Escherichia coli O157:H7 & Streptococcus aureus | Yes | ssDNA | RPA | Yes | Spiked milk | F | Portable fluorescence reader | rfbE & nuc | 50 | 1 CFU/mL | [23] |
| 13 | Cas12a | NR | Salmonella typhimurium | Yes | ssDNA | LAMP | Yes | Inactivated bacterial culture | F | Portable UV lamp & milk warmer | Genomic DNA | 60 | 800 CFU/mL | [24] |
| 14 | LbCas12a | RPA-Cas12a-FS | Escherichia coli, Listeria monocytogenes, Staphylococcus aureus & Vibrio parahaemolyticus | Yes | ssDNA | RPA | Yes | Pork, duck meat, and beef | F | Handheld fluorometer | Genomic DNA | 45 | 10 copies (Escherichia coli, Listeria monocytogenes, Staphylococcus aureus) & 100 copies (Vibrio parahaemolyticus) | [25] |
| 15 | Cas9 Cis (dsDNA) | Cas9nAR | Salmonella typhimurium | No | SYBR Green I | Cas9nAR | Yes | Bacteria isolates | F | Fluorescence reader | invA | 60 | 1 copy/10µL | [26] |
| 16 | LbaCas12a | CIA | Pseudomonas aeruginosa | Yes | ssDNA | LAMP | Yes | Spiked human serum, milk & clinical sputum | LFB | No | Acetyltransferase | 50 | 1 CFU/mL | [27] |
| 17 | LbCas12a | NR | Yersinia pestis | Yes | ssDNA | RPA | Yes | Reference laboratory strains & clinical isolates | F & LFB | Real-time PCR machine | Chromosomal DNA (4 tag sites) | ~50 (LFB)-150 (F) | 103 fg/µL (YP-1, YP-2 & YP-3)-106 fg/µL (YP-4) [F] | [28] |
| 18 | Cas12f(a) | Cas-TSPE | Escherichia coli, Eberthella typhi, Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus pyogenes & Enterococcus faecalis | Yes | ssDNA | PCR | Yes | Spiked blood & urine | F | Fluorescence reader | Variable regions (V3) of 16S rRNA | ~210 | 1 CFU/mL (Streptococcus pyogenes) | [29] |
| 19 | Cas12a | NR | Escherichia coli O157:H7 | Yes | ssDNA | RCA | No | Spiked skimmed milk powder | EB | CHI660E electrochemical workstation & CE | Aptamer | NR | 10 CFU/mL | [30] |
| 20 | LbaCas12a | NR | Escherichia coli O157:H7 | Yes | ssDNA | Primer exchange reaction & SDA | No | Spiked milk | EB | CHI660E electrochemical workstation & CE | Aptamer | ~180 | 19 CFU/mL | [31] |
| 21 | LbCas12a | NR | Salmonella spp. | No. Catalyze TMB-H2O2 reaction (Blue to yellow) | G-quadruplex hemin (DNAzyme) | RPA | Yes | Spiked beer & juice | Colorimetric & quantitative analysis | Smartphone readout with Color Picker APP | invA | 180 | 1 CFU/mL | [32] |
| 22 | AacCas12b | NR | Campylobacter jejuni | Yes | ssDNA | PCR | Yes | Spiked chicken (n = 55) | F | Thermal Cycler & blue light | flhA | 40 | 10 CFU/g | [33] |
| 23 | EnGen® LbaCas12a | NR | Gram-negative bacteria: Escherichia coli (LPS) | Yes | ssDNA | No | No (purchased LPS) | Spiked purified water, milk, grapefruit juice & green tea | F (inhibitory effect) | Fluorescence spectrometer | Aptamer | ~140 | 23 CFU/mL | [34] |
| 24 | Cas9 Cis (dsDNA) nickase | NR | Salmonella typhimurium & Escherichia coli | No | ssDNA (fluorescence tagged Primers) | Cas9nAR | Yes | Spiked milk | LFB | Portable test strip reader | invA & UidA | 180 | 100 CFU/mL | [35] |
| 25 | LbCas12a | NR | Staphylococcus aureus | Yes | ssDNA | PCR | Yes | Spiked milk | Elementary OR AND INHIBIT logic gates | Microplate reader | femA | 120 | 103 CFU/mL | [36] |
| 26 | Cas9 | CASLFA | Listeria monocytogenes | No | AuNP-DNA Probe | PCR | Yes | Reference laboratory strains | LFB | Thermocycler | hlyA | 60 | 150 copies | [37] |
| 27 | Cas9 Cis (ssDNA) | CAS-EXPAR | Listeria monocytogenes | No | SYBR Green I | EXPAR | Yes | Bacterial cells | F | Real-time PCR | hly | 60 | 0.82 amol | [38] |
| 28 | Cas12a | E-Si-CRISPR | Methicillin-resistant Staphylococcus aureus (MRSA) | Yes | ssDNA | No | Yes | Spiked human serum | EB | PGSTAT204 AutoLab, SPGE & impedance analyser | mecA | 90 | 3.5 fM | [39] |
| 29 | EnGen® LbaCas12a | NR | Mycobacterium tuberculosis H37Rv | Yes | ssDNA | RPA | Yes | BALB, hydrothorax, and homogenate of needle biopsy (n = 69) | Gel electrophoresis | No | IS6110 | 40 | 1 copy/uL | [40] |
| 30 | LwCas13a | PCF | Salmonella spp. | Yes | ssRNA | PCR & reverse transcription | Yes | Reference laboratory strains & bacterial isolates | F | Thermocycler & fluorescence reader | invA | 120 | 10 CFU/mL | [41] |
| 31 | FnCas12a | NR | Mycobacterium abscessus species and subspecies | Yes | ssDNA | PCR | Yes | Clinical isolates | F | Thermocycler & Fluorescence reader | rpoB & erm (41) | ~240 | NR | [42] |
| 32 | LbaCas12a | NR | Multidrug-resistant Acinetobacter baumannii (MDRAB) | Yes | ssDNA | PCR | Yes | Reference laboratory strains | F | Thermocycler & fluorescence spectrophotometer | glt A & β-lactamase genes | 120 | 50 CFU/mL | [43] |
| 33 | dCas9 | CRISPR-mediated DNA-FISH | Methicillin-resistant Staphylococcus aureus (MRSA) | No | SYBR Green I | No | Yes | Bacterial cells | F | Fluorescence spectroscopy | mecA | 30 | 10 CFU/mL | [44] |
| 34 | Cas13a | NR | Bacillus cereus | Yes | Light-up RNA aptamer (Broccoli) | No | Yes | Spiked milk & rice | F | Fluorescence microplate reader | 16s rRNA | NR | 9.83 CFU | [45] |
| 35 | LwCas13a | CCB-Detection | Staphylococcus aureus | Yes | ssRNA | PCR & reverse transcription | Yes | Spiked milk, juice, beer & water | F | Microplate reader | nuc | 240 | 1 CFU/mL | [46] |
| 36 | LbCas12a | NR | Salmonella spp. | Yes | ssDNA | PCR | Yes | Spiked milk | F | Portable colorimeter & portable NIR irradiator | invA | 90 | 1 CFU/mL | [47] |
| 37 | Cas12a | NR | Methicillin resistant Staphylococcus aureus (MRSA) | Yes | ssDNA | RCA | No | Spiked serum | F | Fluorescence spectroscopy | Aptamers against protein A & PBP2a | ~75 | NR | [48] |
| 38 | LwaCas13a | SHERLOCK | Yersinia pestis | Yes | ssRNA | RPA | Yes | Reference laboratory strains & bacterial isolates | F | Microplate reader | lcrV | 150 | 420 copies/mL | [49] |
| 39 | Cas12a | NR | Vibrio parahaemolyticus | Yes | ssDNA | PCR | Yes | Spiked shrimp | F | Homemade UV light & mini thermal cycler | tlh | ~100 | 102 copies/μL | [50] |
| 40 | LbCas12a | Cas12a-UPTLFA | Yersinia pestis | Yes | ssDNA | RPA | Yes | Spiked blood | LFB | UPT biosensor | pla | 80 | 1 CFU/µL | [51] |
| 41 | EnGen® LbaCas12a | RAA-based E-CRISPR | Listeria monocytogenes | Yes | ssDNA with methylene blue | RAA | Yes | Flammulina velutipes | EB | CHI 660E electrochemical workstation & gold electrodes | LMOSLCC2755_0090 | 120 | 26 CFU/mL | [52] |
| 42 | EnGen® LbaCas12a | Cas12aFDet | Listeria monocytogenes serotype 4c | Yes | ssDNA | RAA | Yes | Spiked fresh grass carp | F | Fluorescence reader | LMOSLCC2755_0090 | 60 | 135 CFU/mL | [53] |
| 43 | Cas12a | NR | Vibrio parahaemolyticus | Yes | ssDNA | LAMP | Yes | Spiked shrimp | F | Portable cartridge | tlh | 50 | 30 copies/reaction | [54] |
| 44 | EnGen® LbaCas12a | Cas12aVDet | Mycoplasma | Yes | ssDNA | RPA | No | Cell culture supernatant | F | Blue light | 16s rRNA | 30 | NR | [55] |
| 45 | Cas12f(a1) | CMP | Streptococcus pyogenes & Eberthella typhi | Yes | ssDNA | Reverse-transcription & APCR | Yes | Spiked milk | F | Fluorescence plate reader | 16S rRNA gene V3 hypervariable region | NR | 103 & 104 CFU/mL | [56] |
| 46 | Cas12a | RAA-CRISPR/Cas12a | Vibrio vulnificus | Yes | ssDNA | RAA | Yes | Spiked human blood & stool | F | UV torch | vvhA | 40 | 2 copies/reaction | [57] |
Table 2.
Summary of subgroup analysis of CRISPR-Cas system-based-bacterial detection.
| Subgroup | Number of Studies in Percentage |
|---|---|
| All studies | 46 (100%) |
| Extraction of nucleic acids | |
| Yes | 83% |
| No | 17% |
| Amplification of nucleic acids | |
| Isothermal reaction | 66% |
| Polymerase chain reaction | 23% |
| No | 11% |
| CRISPR-Cas enzymes | |
| Cas12a | 64% |
| Cas12b | 4% |
| Cas13a | 13% |
| Cas12f | 4% |
| Cas9 | 11% |
| dCas9 | 4% |
| Trans-cleavage activity | |
| Yes | 83% |
| No | 17% |
| Detection methods | |
| Fluorescence | 67% |
| Lateral flow biosensor | 13% |
| Electrochemical biosensor | 11% |
| Others | 9% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Selvam, K.; Ahmad Najib, M.; Khalid, M.F.; Ozsoz, M.; Aziah, I. CRISPR-Cas Systems-Based Bacterial Detection: A Scoping Review. Diagnostics 2022, 12, 1335. https://doi.org/10.3390/diagnostics12061335
AMA Style
Selvam K, Ahmad Najib M, Khalid MF, Ozsoz M, Aziah I. CRISPR-Cas Systems-Based Bacterial Detection: A Scoping Review. Diagnostics. 2022; 12(6):1335. https://doi.org/10.3390/diagnostics12061335
Chicago/Turabian StyleSelvam, Kasturi, Mohamad Ahmad Najib, Muhammad Fazli Khalid, Mehmet Ozsoz, and Ismail Aziah. 2022. "CRISPR-Cas Systems-Based Bacterial Detection: A Scoping Review" Diagnostics 12, no. 6: 1335. https://doi.org/10.3390/diagnostics12061335
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.