
KCTD15 Is Overexpressed in her2+ Positive Breast Cancer Patients and Its Silencing Attenuates Proliferation in SKBR3 CELL LINE

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Online Data and Statistical Analyses
2.3. RNA Extraction and qRT-PCR
- RPS18:fw 5′-CGATGGGCGGCGGAAAATA-3′;rev 5′-CTGCTTTCCTCAACACCACA-3′
- CyclinA:fw 5′-AAATGGGCAGTACAGGAGGA-3′;rev 5′-CCACAGTCAGGGAGTGCTTT-3′
- CyclinB:fw 5′-CATGGTGCACTTTCCTCCTT-3′;rev 5′ AGGTAATGTTGTAGAGTTGGTGTCC-3′
- CyclinE:fw 5′-GGCCAAAATCGACAGGAC-3′;rev 5′-GGGTCTGCACAGACTGCAT-3′
- CyclinD:fw 5′-GCTGTGCATCTACACCGACA-3′;rev 5′-TTGAGCTTGTTCACCAGGAG-3′
2.4. Western Blot Analysis
2.5. MTT Assay
2.6. Migration Assay
2.7. Flow Cytometry Experiments
2.8. Microscopy Analyses
2.9. Study Population
2.10. Immunohistochemistry Assay
3. Results
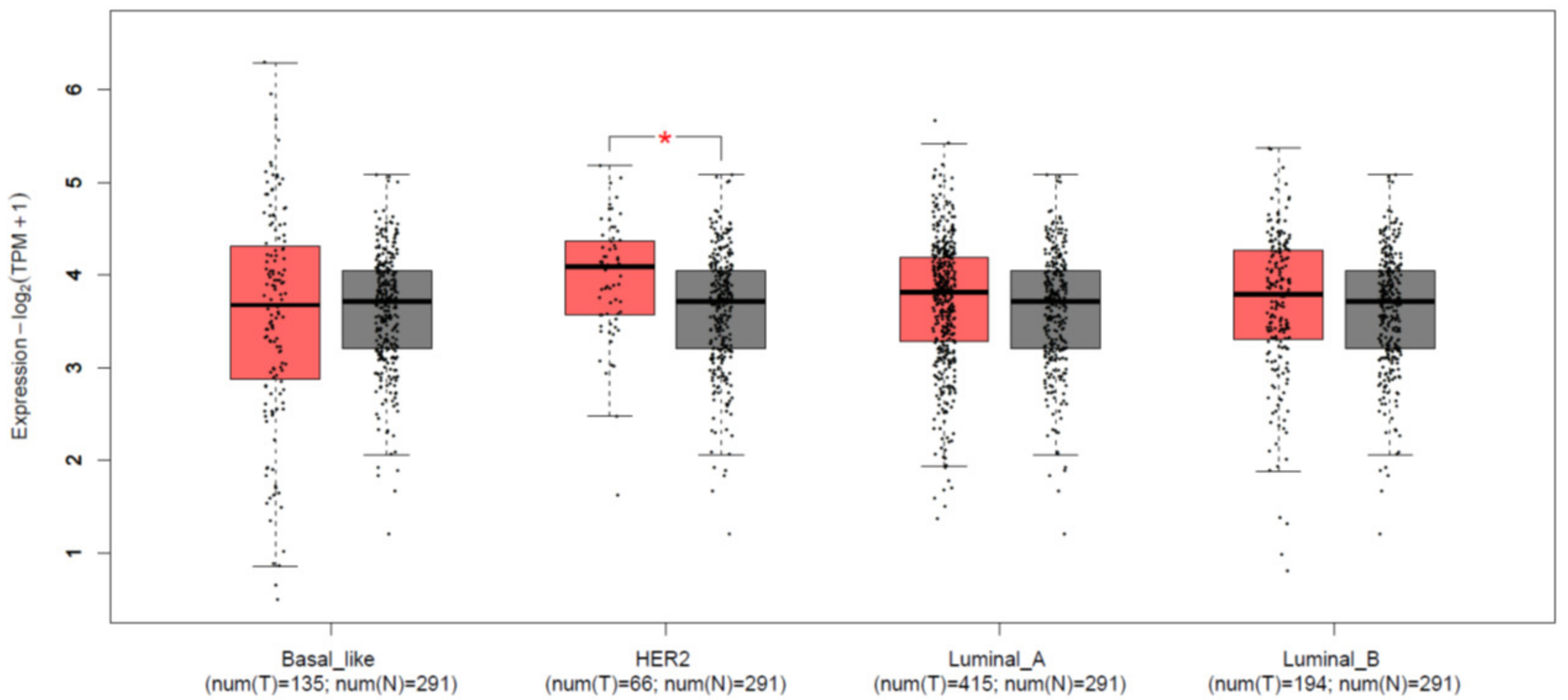
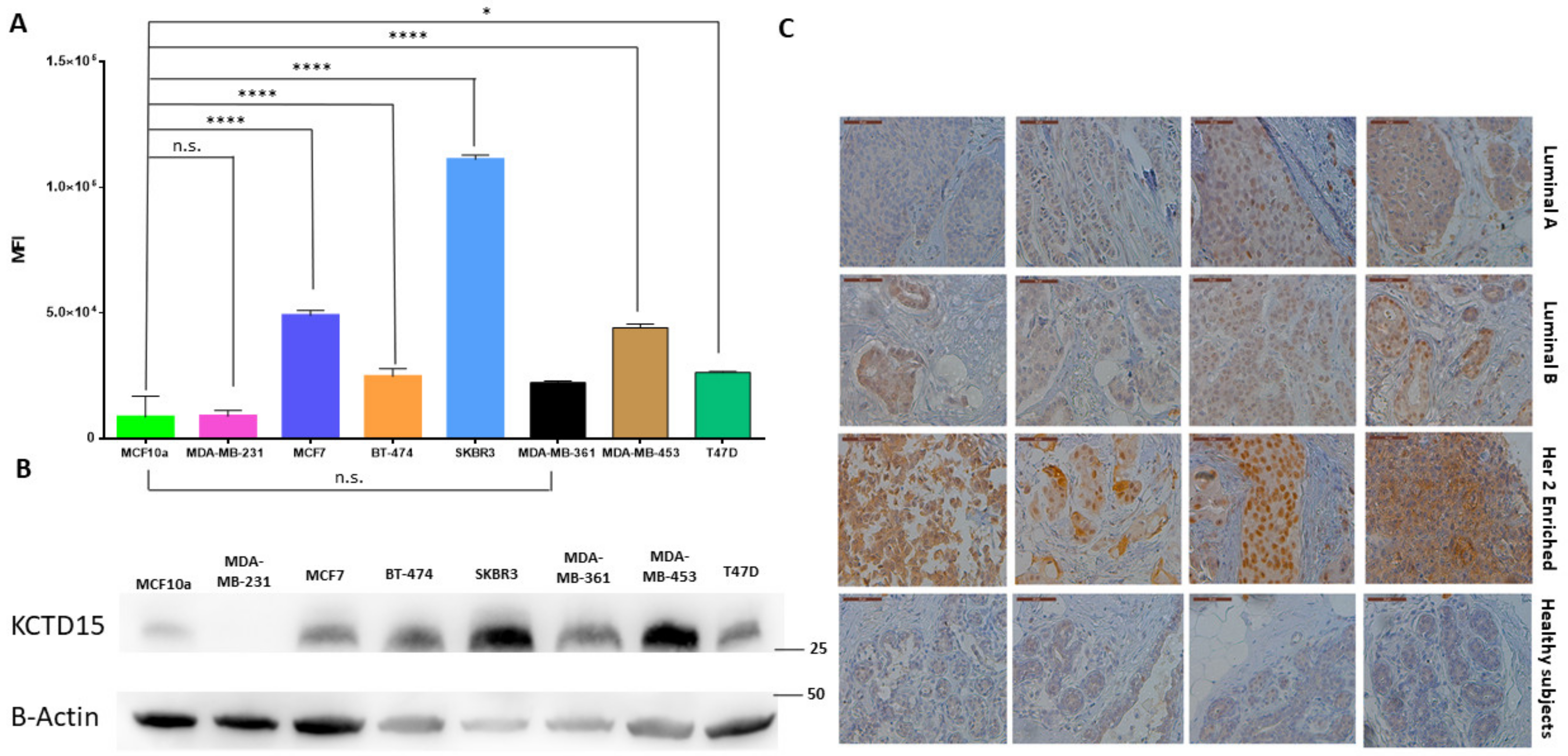
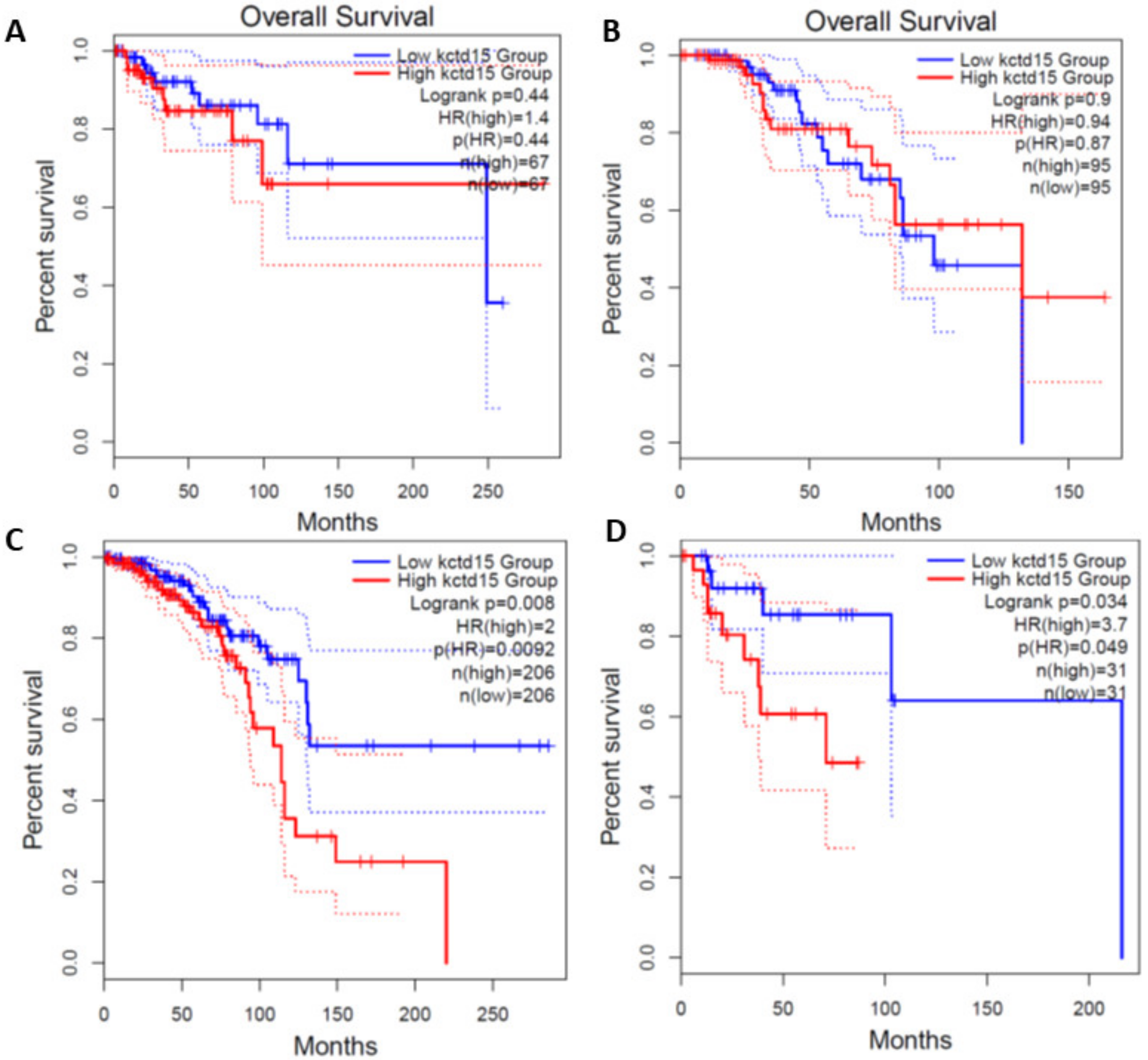
3.1. KCTD15 Is Over-Expressed in Breast Cancer HER2+
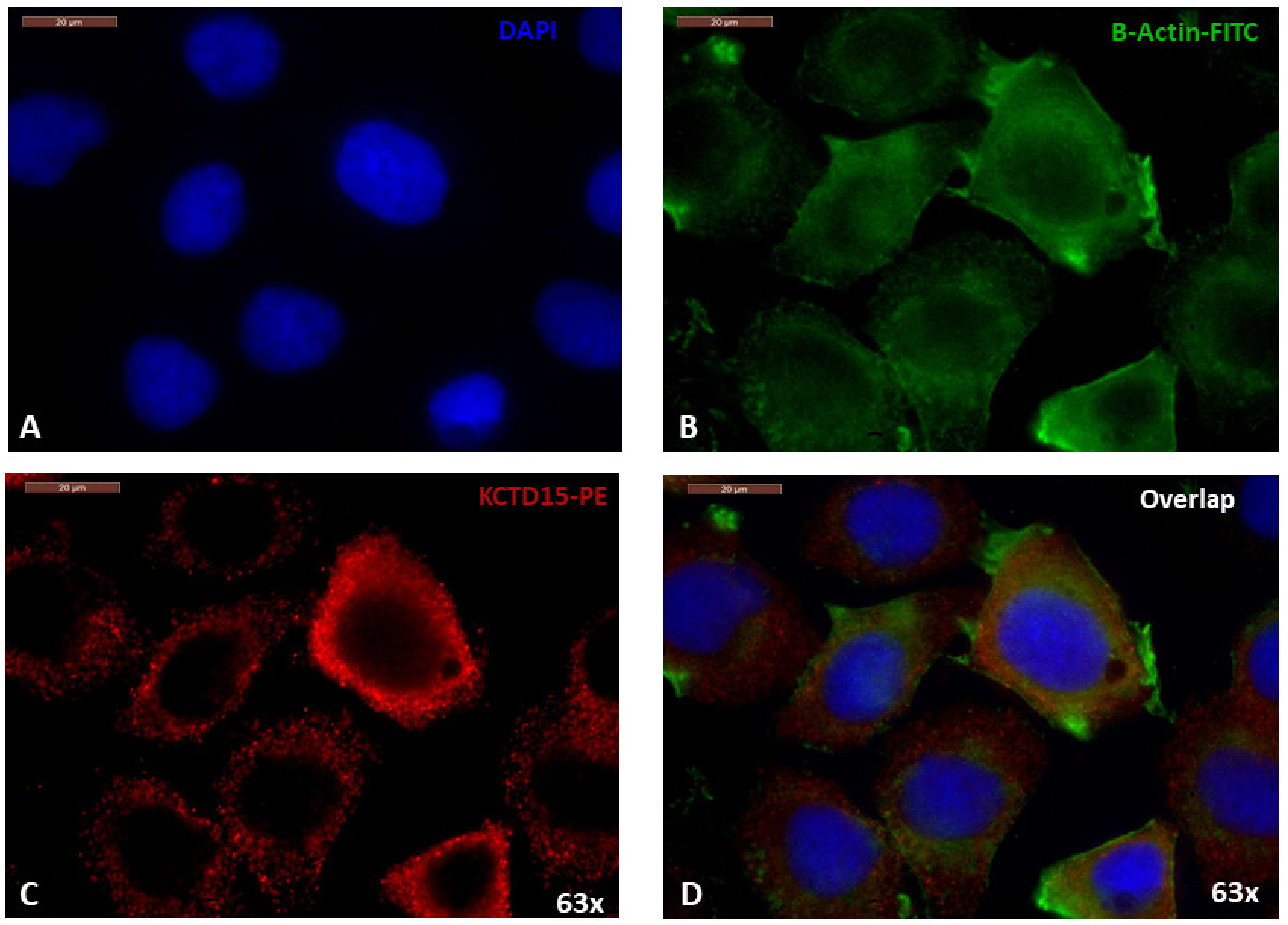
3.2. Cellular Localization of KCTD15 in SKBR3 Cell Line
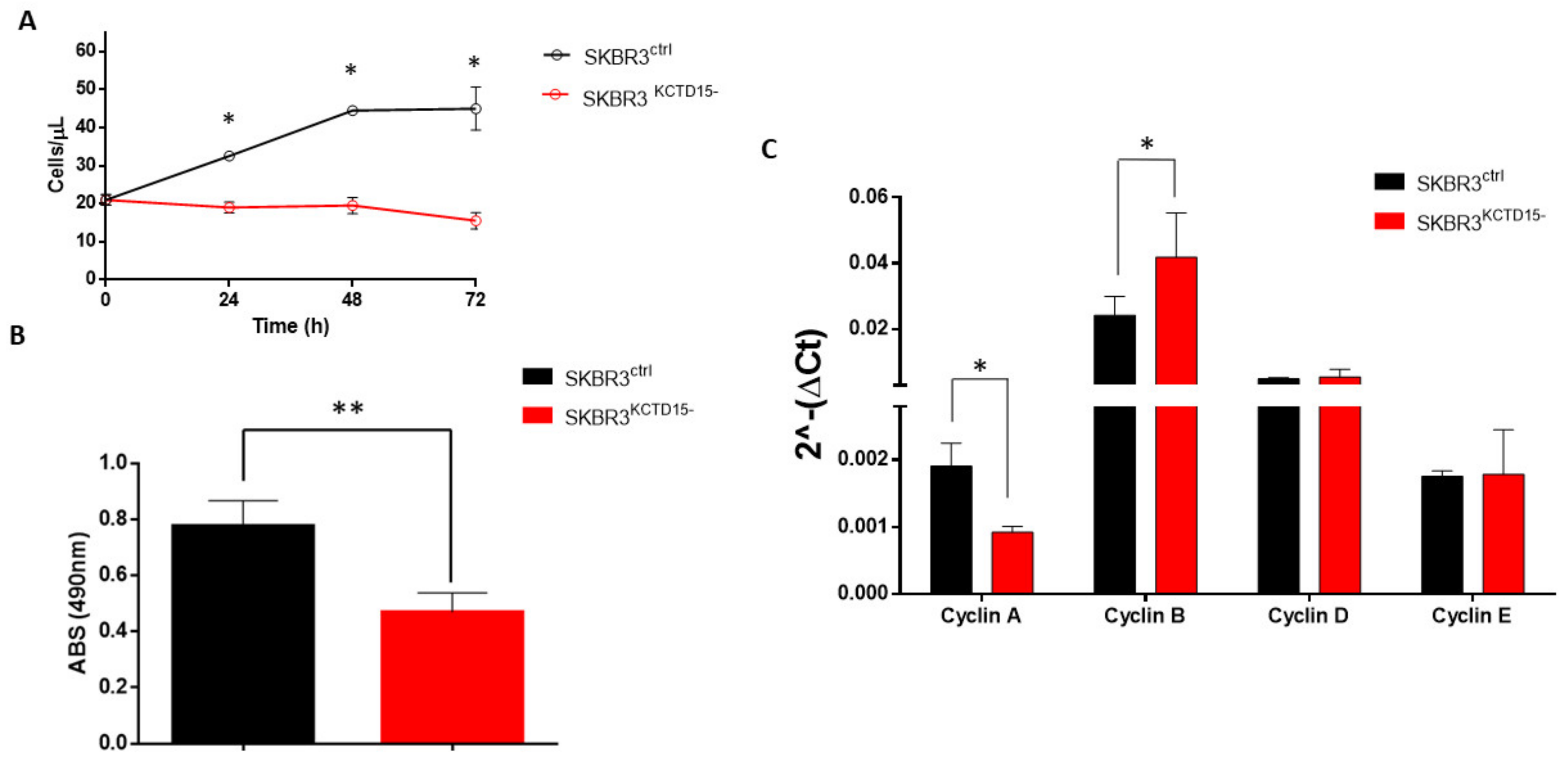
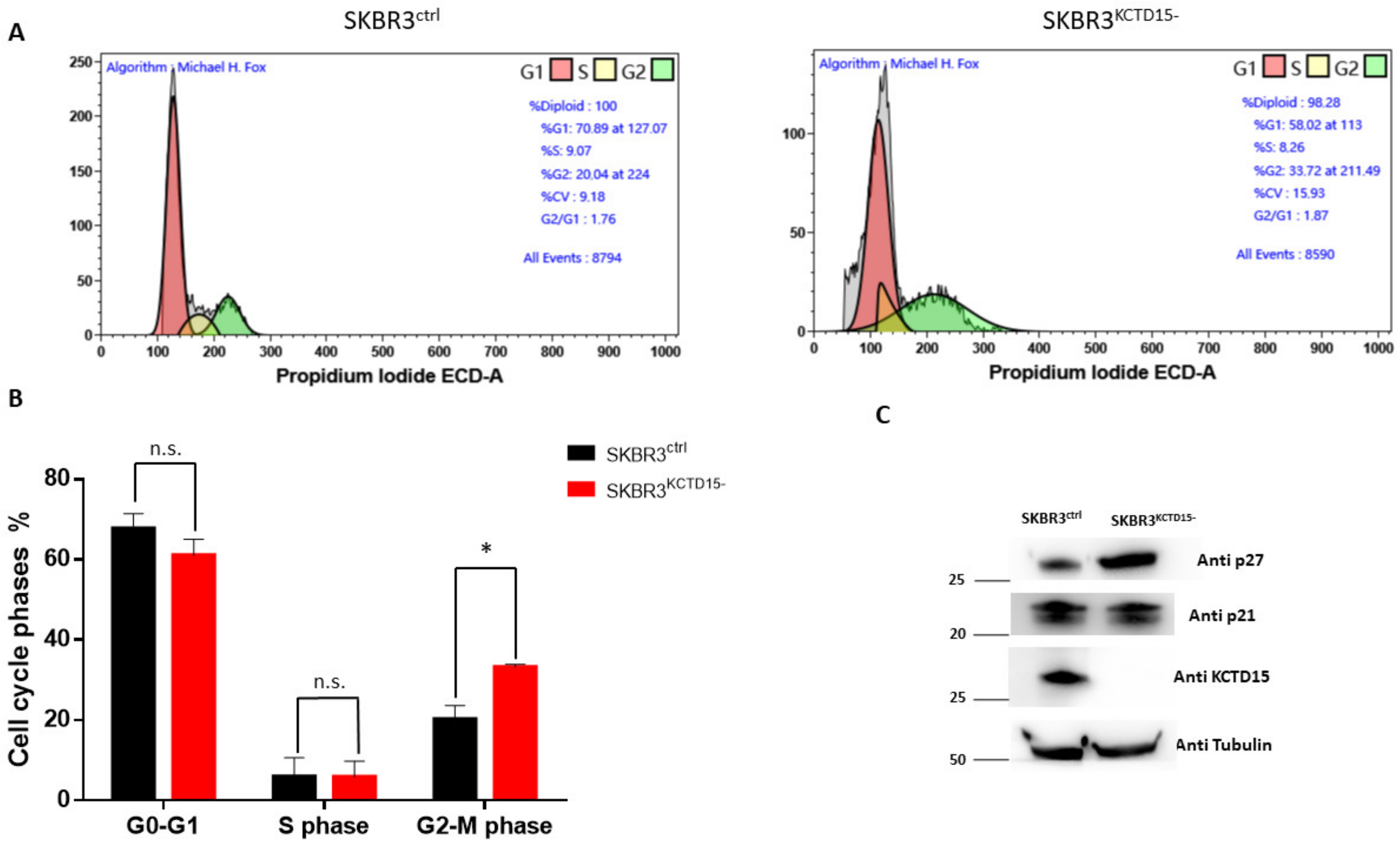
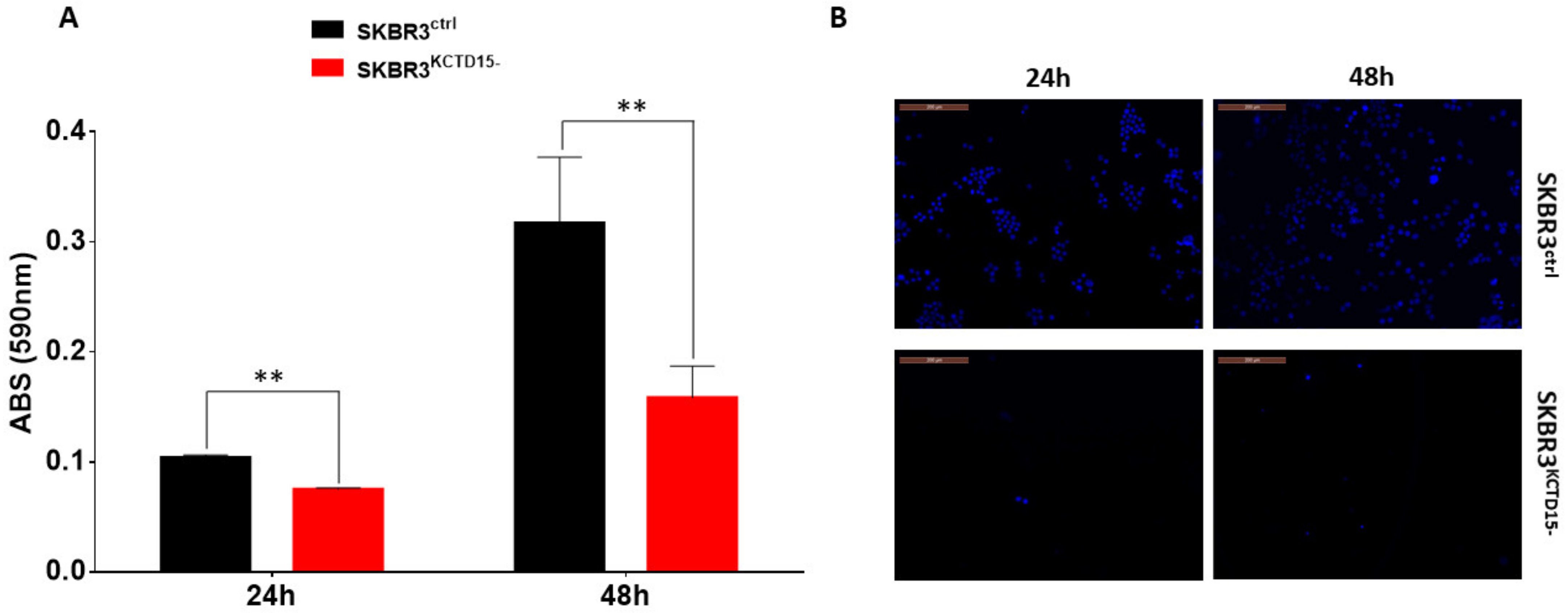
3.3. Silencing of KCTD15 Inhibits SKBR3 Proliferation and Migration, Determining a Cell Cycle Arrest
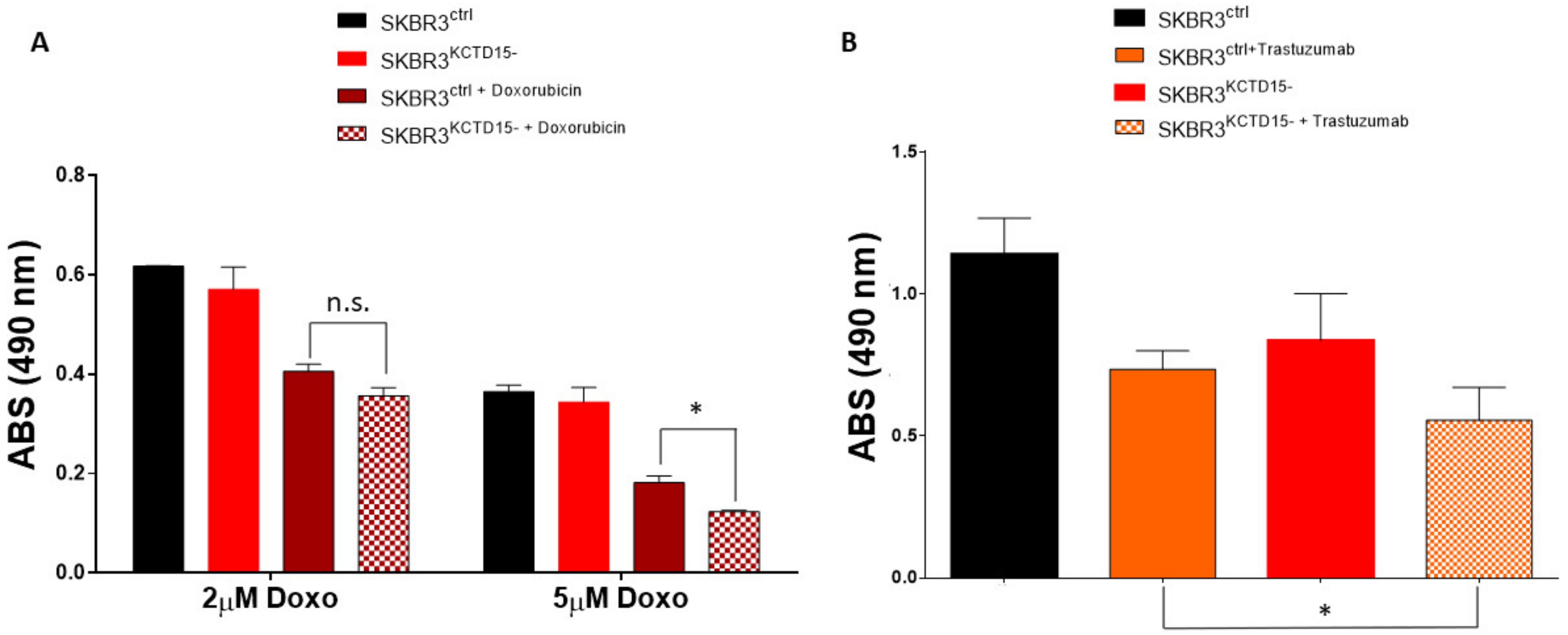
3.4. Silencing of KCTD15 Sensitizes SKBR3 Cells to Doxorubicin and Trastuzumab Treatments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BC | Breast Cancer |
| ER | estrogen receptor |
| PR | progesterone receptor |
| GTEx | Genotype-Tissue Expression |
| HER2 | human epidermal growth factor receptor 2 |
| KCTD | potassium channel tetramerization domain |
| IHC | Immunohistochemistry |
| MFI | Mean Fluorescence Intensity |
| FCM | Flow CytoMetry |
| OS | Overall Survival |
| FFPE | Formalin-Fixed and Paraffin-Embedded |
| TNBC | Triple-negative breast cancer |
| TCGA | Cancer Genome Atlas |
| RTKs | Tyrosine Kinase Receptors |
| EGFR | Epidermal Growth Factor |
| IGF-1R | Insulin-like Growth Factor 1 |
References
- Skoblov, M.; Marakhonov, A.; Marakasova, E.; Guskova, A.; Chandhoke, V.; Birerdinc, A.; Baranova, A. Protein partners of KCTD proteins provide insights about their functional roles in cell differentiation and vertebrate development. BioEssays News Rev. Mol. Cell. Dev. Biol. 2013, 35, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Teng, X.; Aouacheria, A.; Lionnard, L.; Metz, K.A.; Soane, L.; Kamiya, A.; Hardwick, J.M. KCTD: A new gene family involved in neurodevelopmental and neuropsychiatric disorders. CNS Neurosci. Ther. 2019, 25, 887–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canettieri, G.; Di Marcotullio, L.; Greco, A.; Coni, S.; Antonucci, L.; Infante, P.; Pietrosanti, L.; De Smaele, E.; Ferretti, E.; Miele, E.; et al. Histone deacetylase and Cullin3–RENKCTD11 ubiquitin ligase interplay regulates Hedgehog signalling through Gli acetylation. Nat. Cell Biol. 2010, 12, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Abreu, N.; Levitz, J.; Kruse, A.C. Structural basis for KCTD-mediated rapid desensitization of GABAB signalling. Nature 2019, 567, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Yang, J.; Xu, W.W.; Wang, Y.; Zheng, C.-C.; Li, B.; He, Q.-Y. KCTD12 promotes tumorigenesis by facilitating CDC25B/CDK1/Aurora A-dependent G2/M transition. Oncogene 2017, 36, 6177–6189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Bradfield, J.P.; Li, M.; Wang, K.; Zhang, H.; Kim, C.E.; Annaiah, K.; Glessner, J.; Thomas, K.; Garris, M.; et al. The Role of Obesity-associated Loci Identified in Genome-wide Association Studies in the Determination of Pediatric BMI. Obes. Silver Spring Md. 2009, 17, 2254–2257. [Google Scholar] [CrossRef] [PubMed]
- Smaldone, G.; Balasco, N.; Pirone, L.; Caruso, D.; Di Gaetano, S.; Pedone, E.M.; Vitagliano, L. Molecular basis of the scalp-ear-nipple syndrome unraveled by the characterization of disease-causing KCTD1 mutants. Sci. Rep. 2019, 9, 10519. [Google Scholar] [CrossRef]
- Spiombi, E.; Angrisani, A.; Fonte, S.; De Feudis, G.; Fabretti, F.; Cucchi, D.; Izzo, M.; Infante, P.; Miele, E.; Po, A.; et al. KCTD15 inhibits the Hedgehog pathway in Medulloblastoma cells by increasing protein levels of the oncosuppressor KCASH2. Oncogenesis 2019, 8, 64. [Google Scholar] [CrossRef]
- Ye, R.; Kuang, X.; Zeng, H.; Shao, N.; Lin, Y.; Wang, S. KCTD12 promotes G1/S transition of breast cancer cell through activating the AKT/FOXO1 signaling. J. Clin. Lab. Anal. 2020, 34, e23315. [Google Scholar] [CrossRef] [Green Version]
- Angrisani, A.; Di Fiore, A.; De Smaele, E.; Moretti, M. The emerging role of the KCTD proteins in cancer. Cell Commun. Signal. 2021, 19, 56. [Google Scholar] [CrossRef]
- Li, X.; Chen, C.; Wang, F.; Huang, W.; Liang, Z.; Xiao, Y.; Wei, K.; Wan, Z.; Hu, X.; Xiang, S.; et al. KCTD1 Suppresses Canonical Wnt Signaling Pathway by Enhancing β-catenin Degradation. PLoS ONE 2014, 9, e94343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, S.; Dawid, I.B. Kctd15 inhibits neural crest formation by attenuating Wnt/β-catenin signaling output. Dev. Camb. Engl. 2010, 137, 3013–3018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamero-Villarroel, C.; González, L.M.; Rodríguez-López, R.; Albuquerque, D.; Carrillo, J.A.; García-Herráiz, A.; Flores, I.; Gervasini, G. Influence ofTFAP2BandKCTD15genetic variability on personality dimensions in anorexia and bulimia nervosa. Brain Behav. 2017, 7, e00784. [Google Scholar] [CrossRef] [Green Version]
- Pirone, L.; Smaldone, G.; Spinelli, R.; Barberisi, M.; Beguinot, F.; Vitagliano, L.; Miele, C.; Di Gaetano, S.; Raciti, G.A.; Pedone, E. KCTD1: A novel modulator of adipogenesis through the interaction with the transcription factor AP2α. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2019, 1864, 158514. [Google Scholar] [CrossRef] [PubMed]
- Smaldone, G.; Pirone, L.; Capolupo, A.; Vitagliano, L.; Monti, M.C.; Di Gaetano, S.; Pedone, E. The essential player in adipogenesis GRP78 is a novel KCTD15 interactor. Int. J. Biol. Macromol. 2018, 115, 469–475. [Google Scholar] [CrossRef]
- Marneros, A.G.; Beck, A.E.; Turner, E.H.; McMillin, M.J.; Edwards, M.J.; Field, M.; Sobreira, N.L.D.M.; Perez, A.B.A.; Fortes, J.A.; Lampe, A.K.; et al. Mutations in KCTD1 Cause Scalp-Ear-Nipple Syndrome. Am. J. Hum. Genet. 2013, 92, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Smaldone, G.; Coppola, L.; Incoronato, M.; Parasole, R.; Ripaldi, M.; Vitagliano, L.; Mirabelli, P.; Salvatore, M. KCTD15 Protein Expression in Peripheral Blood and Acute Myeloid Leukemia. Diagnostics 2020, 10, 371. [Google Scholar] [CrossRef]
- Smaldone, G.; Beneduce, G.; Incoronato, M.; Pane, K.; Franzese, M.; Coppola, L.; Cordella, A.; Parasole, R.; Ripaldi, M.; Nassa, G.; et al. KCTD15 is overexpressed in human childhood B-cell acute lymphoid leukemia. Sci. Rep. 2019, 9, 20108. [Google Scholar] [CrossRef] [Green Version]
- Smaldone, G.; Coppola, L.; Pane, K.; Franzese, M.; Beneduce, G.; Parasole, R.; Menna, G.; Vitagliano, L.; Salvatore, M.; Mirabelli, P. KCTD15 deregulation is associated with alterations of the NF-κB signaling in both pathological and physiological model systems. Sci. Rep. 2021, 11, 18237. [Google Scholar] [CrossRef]
- Carey, L.A.; Perou, C.M.; Livasy, C.A.; Dressler, L.G.; Cowan, D.; Conway, K.; Karaca, G.; Troester, M.A.; Tse, C.K.; Edmiston, S.; et al. Race, Breast Cancer Subtypes, and Survival in the Carolina Breast Cancer Study. JAMA J. Am. Med Assoc. 2006, 295, 2492–2502. [Google Scholar] [CrossRef] [Green Version]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [Green Version]
- Greenwalt, I.; Zaza, N.; Das, S.; Li, B.D. Precision Medicine and Targeted Therapies in Breast Cancer. Surg. Oncol. Clin. N. Am. 2020, 29, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2009, 123, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [Green Version]
- Canonici, A.; Gijsen, M.; Mullooly, M.; Bennett, R.; Bouguern, N.; Pedersen, K.; O’Brien, A.N.; Roxanis, I.; Li, J.-L.; Bridge, E.; et al. Neratinib overcomes trastuzumab resistance in HER2 amplified breast cancer. Oncotarget 2013, 4, 1592–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, M.H. A model for the computer analysis of synchronous DNA distributions obtained by flow cytometry. Cytometry 1980, 1, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Untch, M.; Gerber, B.; Harbeck, N.; Jackisch, C.; Marschner, N.; Möbus, V.; Von Minckwitz, G.; Loibl, S.; Beckmann, M.W.; Blohmer, J.-U.; et al. 13th st. Gallen international breast cancer conference 2013: Primary therapy of early breast cancer evidence, controversies, consensus—Opinion of a german team of experts (zurich 2013). Breast Care Basel Switz. 2013, 8, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Mirabelli, P.; Incoronato, M.; Coppola, L.; Infante, T.; Parente, C.A.; Nicolai, E.; Soricelli, A.; Salvatore, M. SDN Biobank: Bioresource of Human Samples Associated with Functional and/or Morphological Bioimaging Results for the Study of Oncological, Cardiological, Neurological, and Metabolic Diseases. Open J. Bioresour. 2017, 4, 2. [Google Scholar] [CrossRef]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.-P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Yu, Y.; Wang, Z.; Wang, H.; Bieerkehazhi, S.; Zhao, Y.; Suzuk, L.; Zhang, H. Second-generation proteasome inhibitor carfilzomib enhances doxorubicin-induced cytotoxicity and apoptosis in breast cancer cells. Oncotarget 2016, 7, 73697–73710. [Google Scholar] [CrossRef] [Green Version]
- Vranic, S.; Gatalica, Z.; Wang, Z.-Y. Update on the molecular profile of the MDA-MB-453 cell line as a model for apocrine breast carcinoma studies. Oncol. Lett. 2011, 2, 1131–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbastabar, M.; Kheyrollah, M.; Azizian, K.; Bagherlou, N.; Tehrani, S.S.; Maniati, M.; Karimian, A. Multiple functions of p27 in cell cycle, apoptosis, epigenetic modification and transcriptional regulation for the control of cell growth: A double-edged sword protein. DNA Repair 2018, 69, 63–72. [Google Scholar] [CrossRef]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.; Braybrooke, J.; Gray, R.; Hills, R.; Liu, Z.; Peto, R.; Davies, L.; Dodwell, D.; McGale, P.; Pan, H.; et al. Trastuzumab for early-stage, HER2-positive breast cancer: A meta-analysis of 13 864 women in seven randomised trials. Lancet Oncol. 2021, 22, 1139–1150. [Google Scholar] [CrossRef]
- Eroles, P.; Bosch, A.; Pérez-Fidalgo, J.A.; Lluch, A. Molecular biology in breast cancer: Intrinsic subtypes and signaling pathways. Cancer Treat. Rev. 2012, 38, 698–707. [Google Scholar] [CrossRef]
- Osborne, C.K.; Schiff, R. Mechanisms of Endocrine Resistance in Breast Cancer. Annu. Rev. Med. 2011, 62, 233–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baselga, J. Targeting the Phosphoinositide-3 (PI3) Kinase Pathway in Breast Cancer. Oncologist 2011, 16, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, E.; Hisamatsu, Y.; Tanaka, K.; Yamashita, N.; Saeki, H.; Oki, E.; Kitao, H.; Maehara, Y. Molecular mechanisms regulating the hormone sensitivity of breast cancer. Cancer Sci. 2014, 105, 1377–1383. [Google Scholar] [CrossRef]
- Mitri, Z.; Constantine, T.; O’Regan, R. The HER2 Receptor in Breast Cancer: Pathophysiology, Clinical Use, and New Advances in Therapy. Chemother. Res. Pract. 2012, 2012, 743193. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P. Biology and Management of Patients with Triple-Negative Breast Cancer. Oncologist 2016, 21, 1050–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkhofer, E.C.; Cogswell, P.; Baldwin, A.S. Her2 activates NF-κB and induces invasion through the canonical pathway involving IKKα. Oncogene 2009, 29, 1238–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Liu, J.; Jian, J.; Li, J.; Wan, Y.; Huang, C. IKK-β/NF-κB p65 mediates p27Kip1 protein degradation in arsenite response. Biochem. Biophys. Res. Commun. 2014, 447, 563–568. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, J.; Tang, X.; Liu, Y.; Yu, X.; Wang, Z.; Liu, W. Anthocyanins inhibit trastuzumab-resistant breast cancer in vitro and in vivo. Mol. Med. Rep. 2016, 13, 4007–4013. [Google Scholar] [CrossRef] [Green Version]
- Sajadimajd, S.; Yazdanparast, R. Differential behaviors of trastuzumab-sensitive and -resistant SKBR3 cells treated with menadione reveal the involvement of Notch1/Akt/FOXO1 signaling elements. Mol. Cell. Biochem. 2015, 408, 89–102. [Google Scholar] [CrossRef] [PubMed]
- de Paola, I.; Pirone, L.; Palmieri, M.; Balasco, N.; Esposito, L.; Russo, L.; Mazzà, D.; Di Marcotullio, L.; Di Gaetano, S.; Malgieri, G.; et al. Cullin3—BTB Interface: A Novel Target for Stapled Peptides. PLoS ONE 2015, 10, e0121149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Xiang, Y.; Sun, G. The KCTD family of proteins: Structure, function, disease relevance. Cell Biosci. 2013, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Balasco, N.; Pirone, L.; Smaldone, G.; Di Gaetano, S.; Esposito, L.; Pedone, E.M.; Vitagliano, L. Molecular recognition of Cullin3 by KCTDs: Insights from experimental and computational investigations. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2014, 1844, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Maekawa, M.; Kawai, K.; Nakayama, J.; Araki, N.; Semba, K.; Taguchi, T.; Kamei, Y.; Takada, Y.; Higashiyama, S. Cullin-3/KCTD10 E3 complex is essential for Rac1 activation through RhoB degradation in human epidermal growth factor receptor 2-positive breast cancer cells. Cancer Sci. 2018, 110, 650–661. [Google Scholar] [CrossRef]
- Grinchuk, O.V.; Motakis, E.; Kuznetsov, V.A. Complex sense-antisense architecture of TNFAIP1/POLDIP2 on 17q11.2 represents a novel transcriptional structural-functional gene module involved in breast cancer progression. BMC Genom. 2010, 11, S9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canales, J.; Cruz, P.; Díaz, N.; Riquelme, D.; Leiva-Salcedo, E.; Cerda, O. K+ Channel Tetramerization Domain 5 (KCTD5) Protein Regulates Cell Migration, Focal Adhesion Dynamics and Spreading through Modulation of Ca2+ Signaling and Rac1 Activity. Cells 2020, 9, 2273. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Control Sample (n = 4) | |
|---|---|
| 34–55 | (42.5) |
| Breast Cancer Sample (n = 30) | |
| 39–87 | (65) |
| Sex | |
| Woman | 34 |
| Man | 0 |
| Histologic Types | |
| Invasive Ductal Carcinoma | 30 |
| Hyperplasia | 2 |
| Gynecomastia | 2 |
| Subtype | |
| Luminal A | 10 |
| Luminal B | 10 |
| HER2 + | 10 |
| Ki67 | |
| Low (0–29%) | 20 |
| High (30–100%) | 14 |
| Grade | |
| G1 | 1 |
| G2 | 18 |
| G3 | 11 |
| Tumor size | |
| (0.1–2 cm) | 20 |
| (2–5 cm) | 10 |
| >5cm | 0 |
| Lymph node Status (N) | |
| Involved | 10 |
| Uninvolved | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppola, L.; Baselice, S.; Messina, F.; Giannatiempo, R.; Farina, A.; Vitagliano, L.; Smaldone, G.; Salvatore, M. KCTD15 Is Overexpressed in her2+ Positive Breast Cancer Patients and Its Silencing Attenuates Proliferation in SKBR3 CELL LINE. Diagnostics 2022, 12, 591. https://doi.org/10.3390/diagnostics12030591
Coppola L, Baselice S, Messina F, Giannatiempo R, Farina A, Vitagliano L, Smaldone G, Salvatore M. KCTD15 Is Overexpressed in her2+ Positive Breast Cancer Patients and Its Silencing Attenuates Proliferation in SKBR3 CELL LINE. Diagnostics. 2022; 12(3):591. https://doi.org/10.3390/diagnostics12030591
Chicago/Turabian StyleCoppola, Luigi, Simona Baselice, Francesco Messina, Rosa Giannatiempo, Amalia Farina, Luigi Vitagliano, Giovanni Smaldone, and Marco Salvatore. 2022. "KCTD15 Is Overexpressed in her2+ Positive Breast Cancer Patients and Its Silencing Attenuates Proliferation in SKBR3 CELL LINE" Diagnostics 12, no. 3: 591. https://doi.org/10.3390/diagnostics12030591