Plasma Exosomal Brain-Derived Neurotrophic Factor Correlated with the Postural Instability and Gait Disturbance–Related Motor Symptoms in Patients with Parkinson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Clinical Assessments
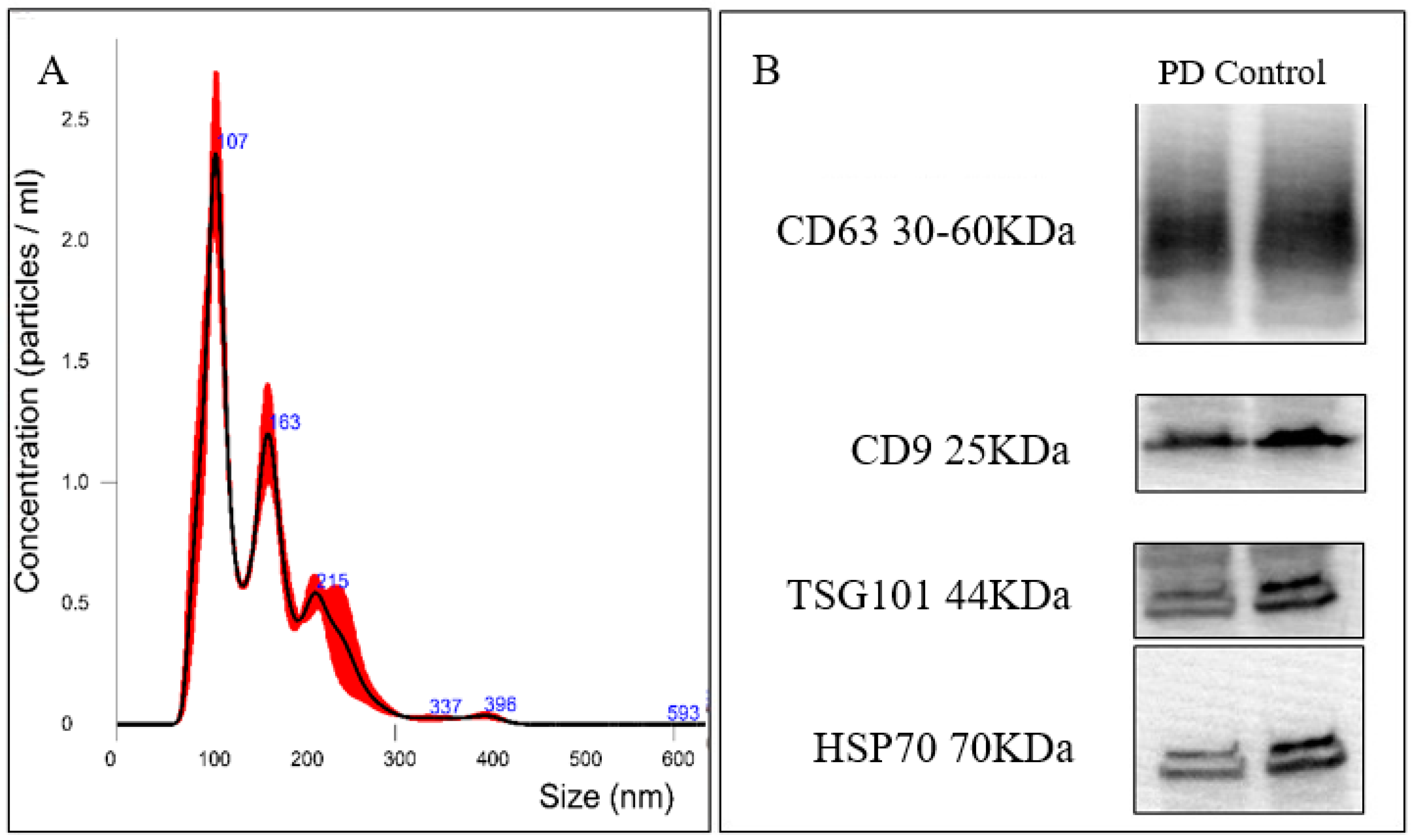
2.3. Plasma Exosomes Separation and Validation
2.4. Assessment of BDNF
2.5. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- De Lau, L.M.; Breteler, M.M. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D. Neurotrophic Factors: An Overview. In Neurotrophic Factors: Methods and Protocols; Skaper, S.D., Ed.; Springer: New York, NY, USA, 2018; pp. 1–17. [Google Scholar]
- Howells, D.W.; Porritt, M.J.; Wong, J.Y.F.; Batchelor, P.E.; Kalnins, R.; Hughes, A.J.; Donnan, G.A. Reduced BDNF mRNA Expression in the Parkinson’s Disease Substantia Nigra. Exp. Neurol. 2000, 166, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Parain, K.; Murer, M.G.; Yan, Q.; Faucheux, B.; Agid, Y.; Hirsch, E.; Raisman-Vozari, R. Reduced expression of brain-derived neurotrophic factor protein in Parkinson’s disease substantia nigra. Neuroreport 1999, 10, 557–561. [Google Scholar] [CrossRef]
- Palasz, E.; Wysocka, A.; Gasiorowska, A.; Chalimoniuk, M.; Niewiadomski, W.; Niewiadomska, G. BDNF as a Promising Therapeutic Agent in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 1170. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, P.; Brassard, P.; Adser, H.; Pedersen, M.V.; Leick, L.; Hart, E.; Secher, N.H.; Pedersen, B.K.; Pilegaard, H. Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp. Physiol. 2009, 94, 1062–1069. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, H.; Wang, C.; Ming, F.; Shi, X.; Yang, M. Serum level of brain-derived neurotrophic factor in Parkinson’s disease: A meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 88, 168–174. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Kang, Y.S.; Buciak, J.L. Transport of human recombinant brain-derived neurotrophic factor (BDNF) through the rat blood-brain barrier in vivo using vector-mediated peptide drug delivery. Pharm. Res. 1994, 11, 738–746. [Google Scholar] [CrossRef]
- Jan, A.T.; Malik, M.A.; Rahman, S.; Yeo, H.R.; Lee, E.J.; Abdullah, T.S.; Choi, I. Perspective Insights of Exosomes in Neurodegenerative Diseases: A Critical Appraisal. Front. Aging Neurosci. 2017, 9, 317. [Google Scholar] [CrossRef] [Green Version]
- Porro, C.; Trotta, T.; Panaro, M.A. Microvesicles in the brain: Biomarker, messenger or mediator? J. Neuroimmunol. 2015, 288, 70–78. [Google Scholar] [CrossRef]
- D’Anca, M.; Fenoglio, C.; Serpente, M.; Arosio, B.; Cesari, M.; Scarpini, E.A.; Galimberti, D. Exosome Determinants of Physiological Aging and Age-Related Neurodegenerative Diseases. Front. Aging Neurosci. 2019, 11, 232. [Google Scholar] [CrossRef] [Green Version]
- Kapogiannis, D.; Boxer, A.; Schwartz, J.B.; Abner, E.L.; Biragyn, A.; Masharani, U.; Frassetto, L.; Petersen, R.C.; Miller, B.L.; Goetzl, E.J. Dysfunctionally phosphorylated type 1 insulin receptor substrate in neural-derived blood exosomes of preclinical Alzheimer’s disease. FASEB J. 2015, 29, 589–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullins, R.J.; Mustapic, M.; Goetzl, E.J.; Kapogiannis, D. Exosomal biomarkers of brain insulin resistance associated with regional atrophy in Alzheimer’s disease. Hum. Brain Mapp. 2017, 38, 1933–1940. [Google Scholar] [CrossRef] [Green Version]
- Irwin, D.J.; Hurtig, H.I. The Contribution of Tau, Amyloid-Beta and Alpha-Synuclein Pathology to Dementia in Lewy Body Disorders. J. Alzheimer’s Dis. Parkinsonism 2018, 8, 444. [Google Scholar] [CrossRef]
- Hughes, A.J.; Daniel, S.E.; Kilford, L.; Lees, A.J. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: A clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry 1992, 55, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Kotagal, V. Is PIGD a legitimate motor subtype in Parkinson disease? Ann. Clin. Transl. Neurol. 2016, 3, 473–477. [Google Scholar] [CrossRef]
- Lee, J.W.; Song, Y.S.; Kim, H.; Ku, B.D.; Lee, W.W. Alteration of Tremor Dominant and Postural Instability Gait Difficulty Subtypes During the Progression of Parkinson’s Disease: Analysis of the PPMI Cohort. Front. Neurol. 2019, 10, 471. [Google Scholar] [CrossRef] [Green Version]
- Radka, S.F.; Holst, P.A.; Fritsche, M.; Altar, C.A. Presence of brain-derived neurotrophic factor in brain and human and rat but not mouse serum detected by a sensitive and specific immunoassay. Brain Res. 1996, 709, 122–301. [Google Scholar] [CrossRef]
- Klein, A.B.; Williamson, R.; Santini, M.A.; Clemmensen, C.; Ettrup, A.; Rios, M.; Knudsen, G.M.; Aznar, S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int. J. Neuropsychopharmacol. 2011, 14, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, W.; Banks, W.A.; Fasold, M.B.; Bluth, J.; Kastin, A.J. Transport of brain-derived neurotrophic factor across the blood-brain barrier. Neuropharmacology 1998, 37, 1553–1561. [Google Scholar] [CrossRef]
- Zhang, Y.; Pardridge, W.M. Neuroprotection in transient focal brain ischemia after delayed intravenous administration of brain-derived neurotrophic factor conjugated to a blood-brain barrier drug targeting system. Stroke 2001, 32, 1378–1384. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Pardridge, W.M. Neuroprotection with noninvasive neurotrophin delivery to the brain. Proc. Natl. Acad. Sci. USA 1999, 96, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.K.S.; Ho, C.S.H.; Tam, W.W.S.; Kua, E.H.; Ho, R.C.-M. Decreased Serum Brain-Derived Neurotrophic Factor (BDNF) Levels in Patients with Alzheimer’s Disease (AD): A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- Arancibia, S.; Silhol, M.; Moulière, F.; Meffre, J.; Höllinger, I.; Maurice, T.; Tapia-Arancibia, L. Protective effect of BDNF against beta-amyloid induced neurotoxicity in vitro and in vivo in rats. Neurobiol. Dis. 2008, 31, 316–326. [Google Scholar] [CrossRef]
- Sakane, T.; Pardridge, W.M. Carboxyl-directed pegylation of brain-derived neurotrophic factor markedly reduces systemic clearance with minimal loss of biologic activity. Pharm. Res. 1997, 14, 1085–1091. [Google Scholar] [CrossRef]
- Agliardi, C.; Clerici, M. Blood extracellular vesicles (EVs) of central nervous system origin: A window into the brain. Neural Regen. Res. 2020, 15, 55–56. [Google Scholar] [CrossRef]
- Saint-Pol, J.; Gosselet, F.; Duban-Deweer, S.; Pottiez, G.; Karamanos, Y. Targeting and Crossing the Blood-Brain Barrier with Extracellular Vesicles. Cells 2020, 9, 851. [Google Scholar] [CrossRef] [Green Version]
- Suire, C.N.; Eitan, E.; Shaffer, N.C.; Tian, Q.; Studenski, S.; Mattson, M.P.; Kapogiannis, D. Walking speed decline in older adults is associated with elevated pro-BDNF in plasma extracellular vesicles. Exp. Gerontol. 2017, 98, 209–216. [Google Scholar] [CrossRef]
- Okazawa, H.; Murata, M.; Watanabe, M.; Kamei, M.; Kanazawa, I. Dopaminergic stimulation up-regulates the in vivo expression of brain-derived neurotrophic factor (BDNF) in the striatum. FEBS Lett. 1992, 313, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg-Katz, K.; Herman, T.; Jacob, Y.; Giladi, N.; Hendler, T.; Hausdorff, J.M. Gray matter atrophy distinguishes between Parkinson disease motor subtypes. Neurology 2013, 80, 1476–1484. [Google Scholar] [CrossRef] [Green Version]
- Pötter-Nerger, M.; Volkmann, J. Deep brain stimulation for gait and postural symptoms in Parkinson’s disease. Mov. Disord. 2013, 28, 1609–1615. [Google Scholar] [CrossRef]
- Bohnen, N.I.; Albin, R.L. White matter lesions in Parkinson disease. Nat. Rev. Neurol. 2011, 7, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Chiu, W.T.; Chan, L.; Wu, D.; Ko, T.H.; Chen, D.Y.; Hong, C.T. Cerebral Microbleeds are Associated with Postural Instability and Gait Disturbance Subtype in People with Parkinson’s Disease. Eur. Neurol. 2018, 80, 335–340. [Google Scholar] [CrossRef]
- Olson, M.; Lockhart, T.E.; Lieberman, A. Motor Learning Deficits in Parkinson’s Disease (PD) and Their Effect on Training Response in Gait and Balance: A Narrative Review. Front. Neurol. 2019, 10, 62. [Google Scholar] [CrossRef] [Green Version]
- Carretón, O.; Giralt, A.; Torres-Peraza, J.F.; Brito, V.; Lucas, J.J.; Ginés, S.; Canals, J.M.; Alberch, J. Age-dependent decline of motor neocortex but not hippocampal performance in heterozygous BDNF mice correlates with a decrease of cortical PSD-95 but an increase of hippocampal TrkB levels. Exp. Neurol. 2012, 237, 335–345. [Google Scholar] [CrossRef]
- Ploughman, M.; Windle, V.; MacLellan, C.L.; White, N.; Doré, J.J.; Corbett, D. Brain-derived neurotrophic factor contributes to recovery of skilled reaching after focal ischemia in rats. Stroke 2009, 40, 1490–1495. [Google Scholar] [CrossRef] [Green Version]
- Helm, E.E.; Tyrell, C.M.; Pohlig, R.T.; Brady, L.D.; Reisman, D.S. The presence of a single-nucleotide polymorphism in the BDNF gene affects the rate of locomotor adaptation after stroke. Exp. Brain Res. 2016, 234, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Bechara, R.G.; Kelly, Á.M. Exercise improves object recognition memory and induces BDNF expression and cell proliferation in cognitively enriched rats. Behav. Brain Res. 2013, 245, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.; Luciano, M.S.; Carlson, K.; Luo, P.; Marks, W.J., Jr.; Larson, P.S.; Starr, P.A.; Follett, K.A.; Weaver, F.M.; Stern, M.B.; et al. Differential effects of deep brain stimulation target on motor subtypes in Parkinson’s disease. Ann. Neurol. 2015, 77, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; You, Y.; Joseph, C.; Mirzaei, M.; Klistorner, A.; Graham, S.L.; Gupta, V. BDNF Polymorphism: A Review of Its Diagnostic and Clinical Relevance in Neurodegenerative Disorders. Aging Dis. 2018, 9, 523–536. [Google Scholar] [CrossRef] [Green Version]
- Foltran, R.B.; Diaz, S.L. BDNF isoforms: A round trip ticket between neurogenesis and serotonin? J. Neurochem. 2016, 138, 204–221. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Controls | PD | p Value | |
|---|---|---|---|
| Number of patients | 42 | 114 | - |
| Age (years) | 67.02 ± 7.00 | 69.67 ± 8.44 | 0.12 |
| Female patients | 13 | 52 | 0.14 |
| Disease duration (years) | - | 2.70 ± 2.45 | - |
| MMSE | 28.50 ± 1.21 | 24.88 ± 5.03 | <0.001 |
| MoCA | 24.05 ± 2.93 | 20.41 ± 6.08 | 0.001 |
| UPDRS-I | 2.44 ± 1.98 | - | |
| UPDRS-II | 8.06 ± 5.80 | - | |
| UPDRS-III | 22.89 ± 10.00 | - |
| Plasma Exosomal BDNF at Q1 | Plasma Exosomal BDNF at Q2 | Plasma Exosomal BDNF at Q3 | Plasma Exosomal BDNF at Q4 | p for Trend | |
|---|---|---|---|---|---|
| UPDRS I | 2.96 ± 2.70 | 2.17 ± 1.51 | 2.24 ± 1.70 | 2.43 ± 1.89 | 0.447 |
| UPDRS II | 10.59 ± 6.78 | 7.45 ± 5.00 | 8.03 ± 6.19 | 6.29 ± 4.46 | 0.042 |
| UPDRS III | 26.04 ± 11.14 | 21.24 ± 9.73 | 21.52 ± 10.15 | 23.00 ± 8.71 | 0.263 |
| B | SE. | p Value | |
|---|---|---|---|
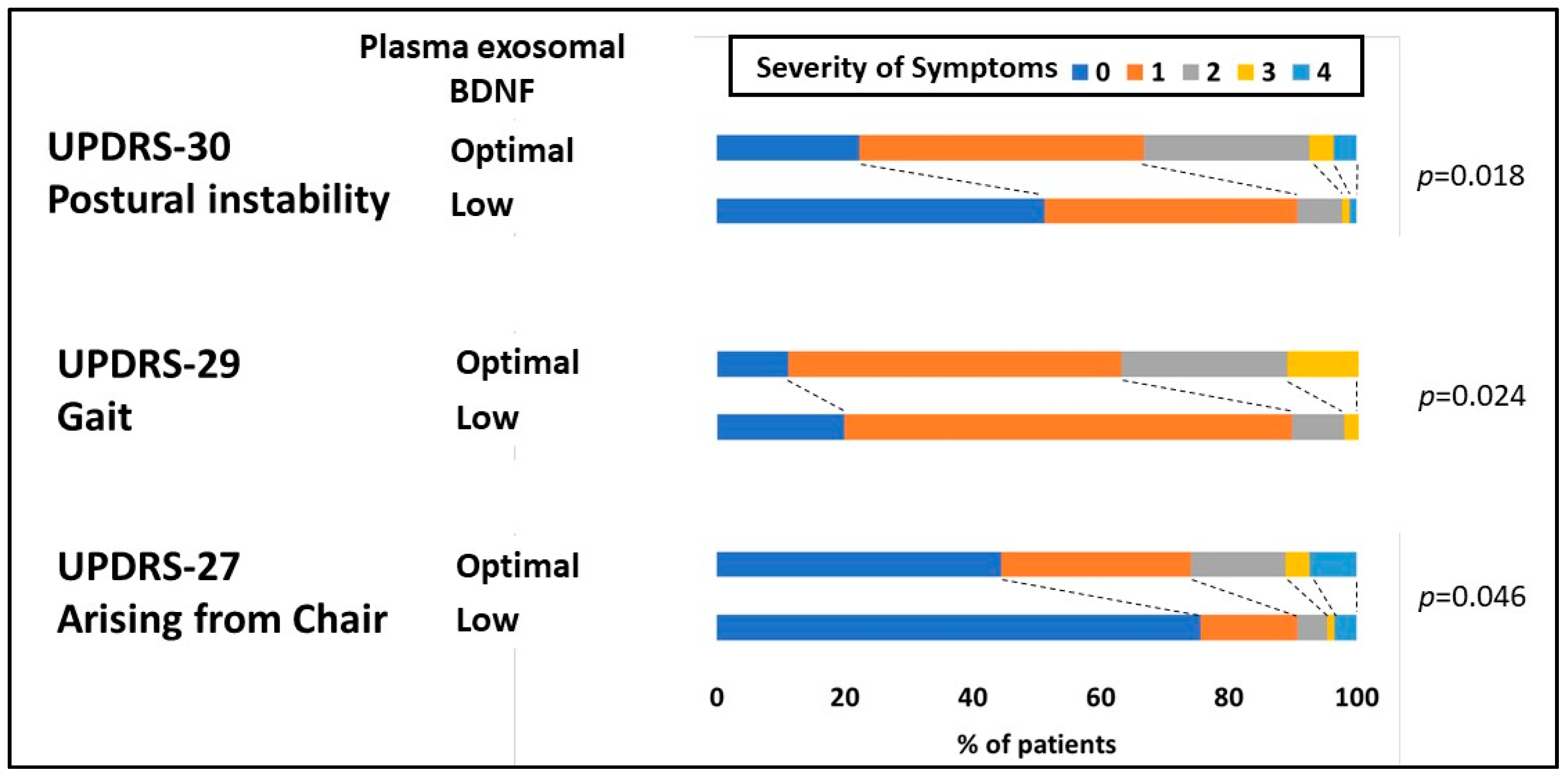
| Arising from the chair | 0.460 | 0.217 | 0.036 |
| Gait | 0.364 | 0.146 | 0.014 |
| Postural stability | 0.546 | 0.180 | 0.003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, C.C.; Huang, P.H.; Chan, L.; Chen, J.-H.; Chien, L.-N.; Hong, C.T. Plasma Exosomal Brain-Derived Neurotrophic Factor Correlated with the Postural Instability and Gait Disturbance–Related Motor Symptoms in Patients with Parkinson’s Disease. Diagnostics 2020, 10, 684. https://doi.org/10.3390/diagnostics10090684
Chung CC, Huang PH, Chan L, Chen J-H, Chien L-N, Hong CT. Plasma Exosomal Brain-Derived Neurotrophic Factor Correlated with the Postural Instability and Gait Disturbance–Related Motor Symptoms in Patients with Parkinson’s Disease. Diagnostics. 2020; 10(9):684. https://doi.org/10.3390/diagnostics10090684
Chicago/Turabian StyleChung, Chen Chih, Pai Hao Huang, Lung Chan, Jia-Hung Chen, Li-Nien Chien, and Chien Tai Hong. 2020. "Plasma Exosomal Brain-Derived Neurotrophic Factor Correlated with the Postural Instability and Gait Disturbance–Related Motor Symptoms in Patients with Parkinson’s Disease" Diagnostics 10, no. 9: 684. https://doi.org/10.3390/diagnostics10090684