
Trends in Stream Biodiversity Research since the River Continuum Concept


Abstract
:1. Introduction
- (1)
- What are the major questions that investigators asked with regard to diversity in lotic systems? We considered a number of aspects that defined research questions including the focal organism(s), the measured predictors of biodiversity, whether studies were explicitly motivated by investigating anthropogenic impacts on diversity, the way in which diversity was measured, and whether questions explicitly dealt with the relationship between biodiversity and evolution. We also considered the major underlying theories or themes motivating the research.
- (2)
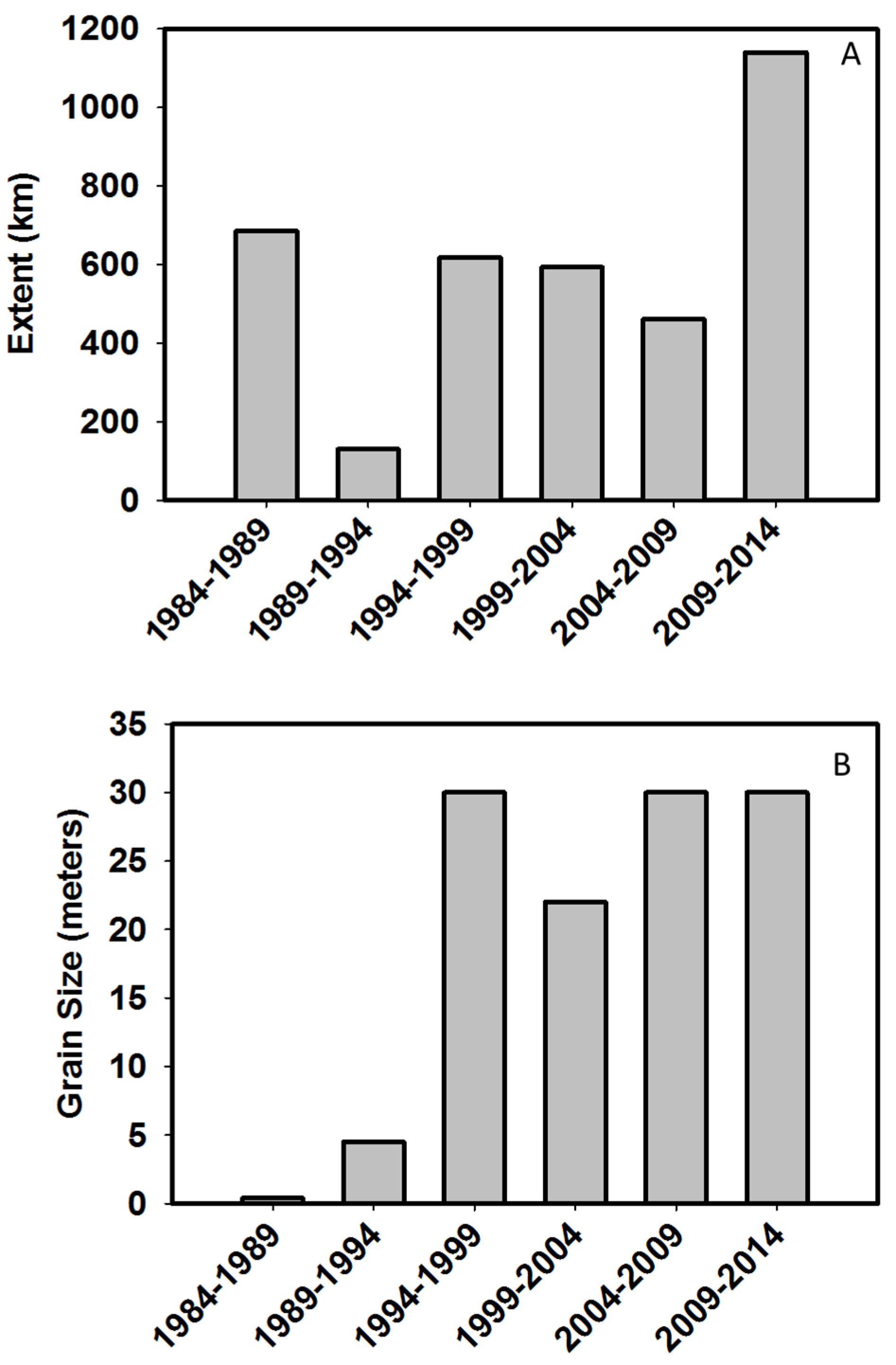
- At what spatial scales are researchers investigating biodiversity? We estimated both grain size and extent of studies, as well as whether investigations were explicitly designed as multi-scale studies. Additionally, we examined keywords that are indicative of multi-scale research.
- (3)
- What proportion of studies had an experimental component? We were particularly interested in how many studies approached the question of biodiversity using experiments to investigate mechanisms.
- (4)
- What factors influence biodiversity in lotic systems according to published studies? Can our current compilation of research on stream biodiversity suggest which commonly considered factors most strongly influence biodiversity patterns?
- (5)
- How have approaches for studying biodiversity in stream systems changed through time? For all of the previous questions, we were also interested in temporal shifts in approaches, questions, or results that signify adaptive responses of stream ecologists to emerging theory and available analytical and observational tools.
2. Experimental Section
Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field | Search Term |
|---|---|
| Title | Stream * OR river * |
| Topic | biodiversity OR diversity OR richness |
| Year | 1981–2014 |
| Journal | Proceedings of the National Academy of Sciences |
| Nature | |
| Science | |
| Proceedings of the Royal Society B | |
| PeerJ | |
| PLOS One | |
| PLOS Biology | |
| Ecology Letters | |
| Trends in Ecology and Evoloution (TREE) | |
| Annual Review of Ecology, Evolution, and Systematics | |
| Ecology | |
| Ecological Applications | |
| Ecological Monographs | |
| Frontiers in Ecology and Environment | |
| Ecosphere | |
| Oikos | |
| Oecologia | |
| The American Naturalist | |
| Journal of Ecology | |
| Journal of Animal Ecology | |
| Ecography | |
| Conservation Ecology | |
| Conservation Biology | |
| Ecological Entomology | |
| Freshwater Science | |
| Freshwater Biology | |
| Limnology and Oceanography | |
| Canadian Journal of Fisheries and Aquatic Science | |
| J. of Aquatic Sci. | |
| Archiv für Hydrobiologie | |
| Hydrobiologia |
| Field Type | Field | Field Value | Field Description |
|---|---|---|---|
| Study descriptors | Author | Text (e.g., “Allan”) | Author surname(s) |
| Year | Numeric (e.g., 1990) | Publication year | |
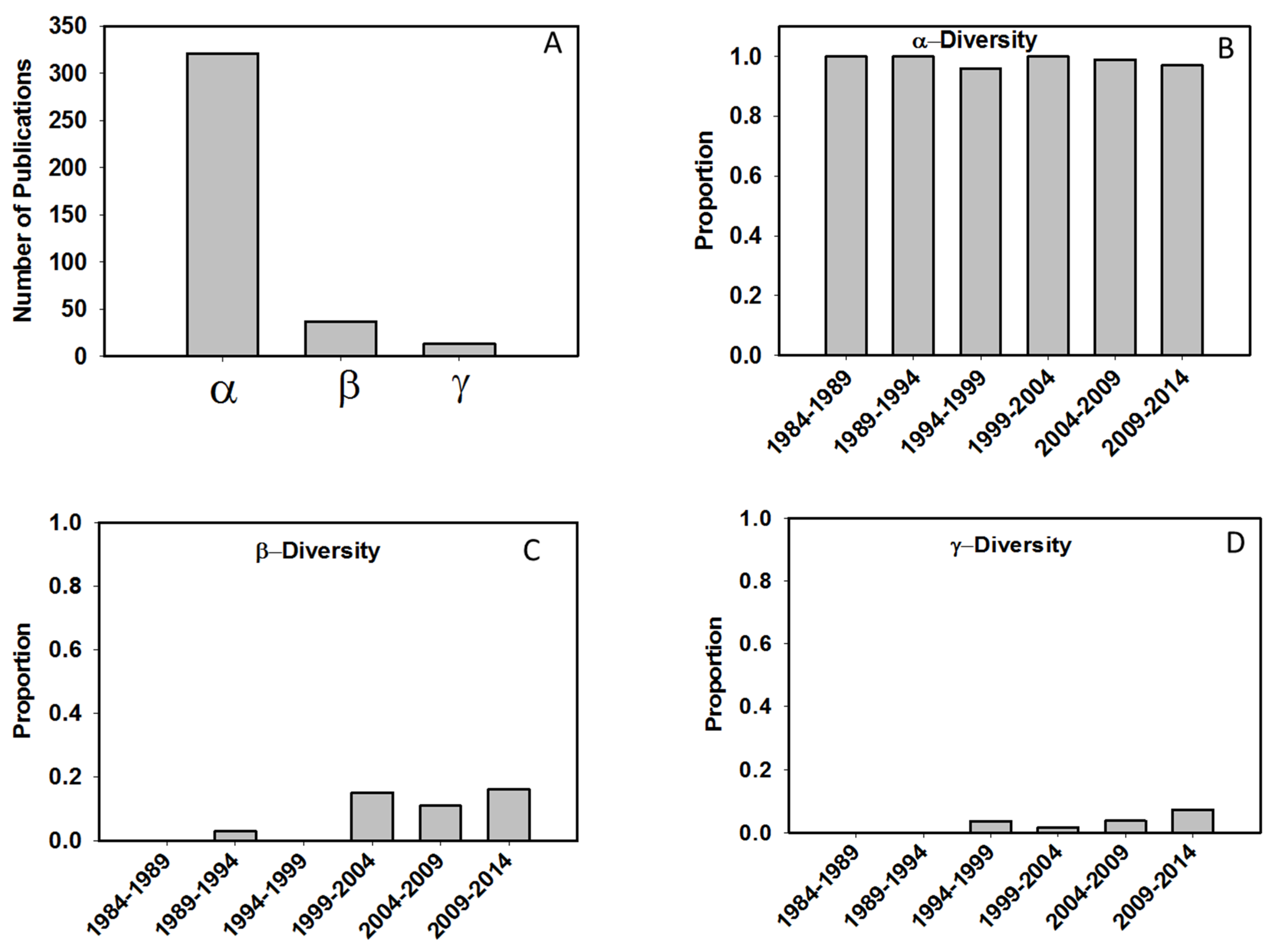
| Response | Text (e.g., alpha, beta, gamma) | Metrics used to quantify stream or river biodiversity and used as response variables in the study | |
| Grain Size (m) | >0 | Approximate size of the unit of inference | |
| Extent (km) | >0 | Approximate study extent (i.e., furthest distance separating two observations) | |
| Survey? | Binary | Was the study a survey? | |
| Experiment? | Binary | Was the study an experiment? | |
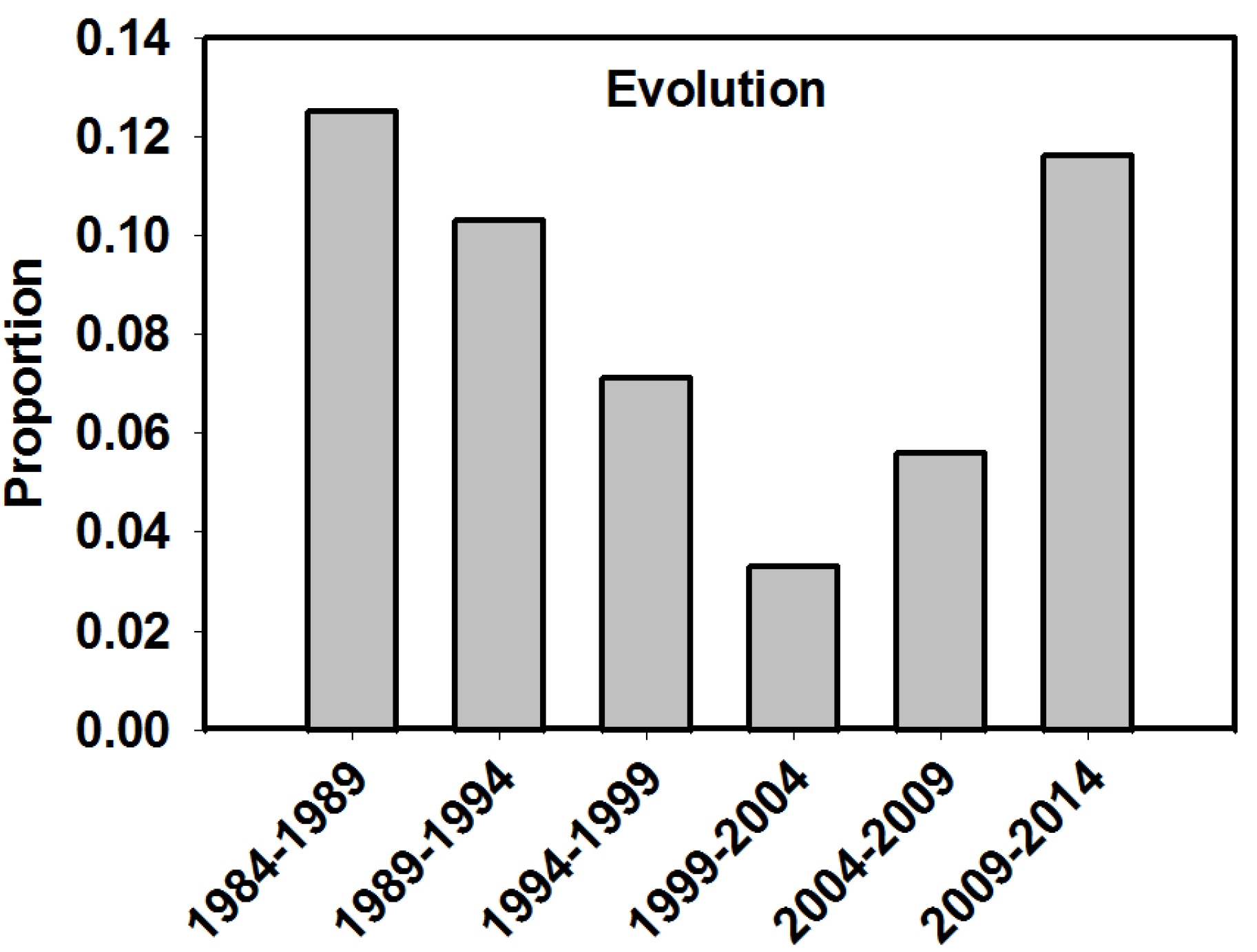
| Evolution? | Binary | Did the study address evolution as a driver of biodiversity? | |
| Multiscale? | Binary | Did the study explore multiscale issues in biodiversity (e.g., local vs. regional controls)? | |
| Anthropogenic? | Binary | Did the study focus on human influences over biodiversity patterns (e.g., climate change, mining)? | |
| Metacommunity? | Binary | Did the study mention the term “metacommunity”? | |
| At least 1 of the “Big 4” metacommunity paradigms mentioned? | Binary | Did the study refer to any of the four metacommunity paradigms described by Leibold et al. (2004)? | |
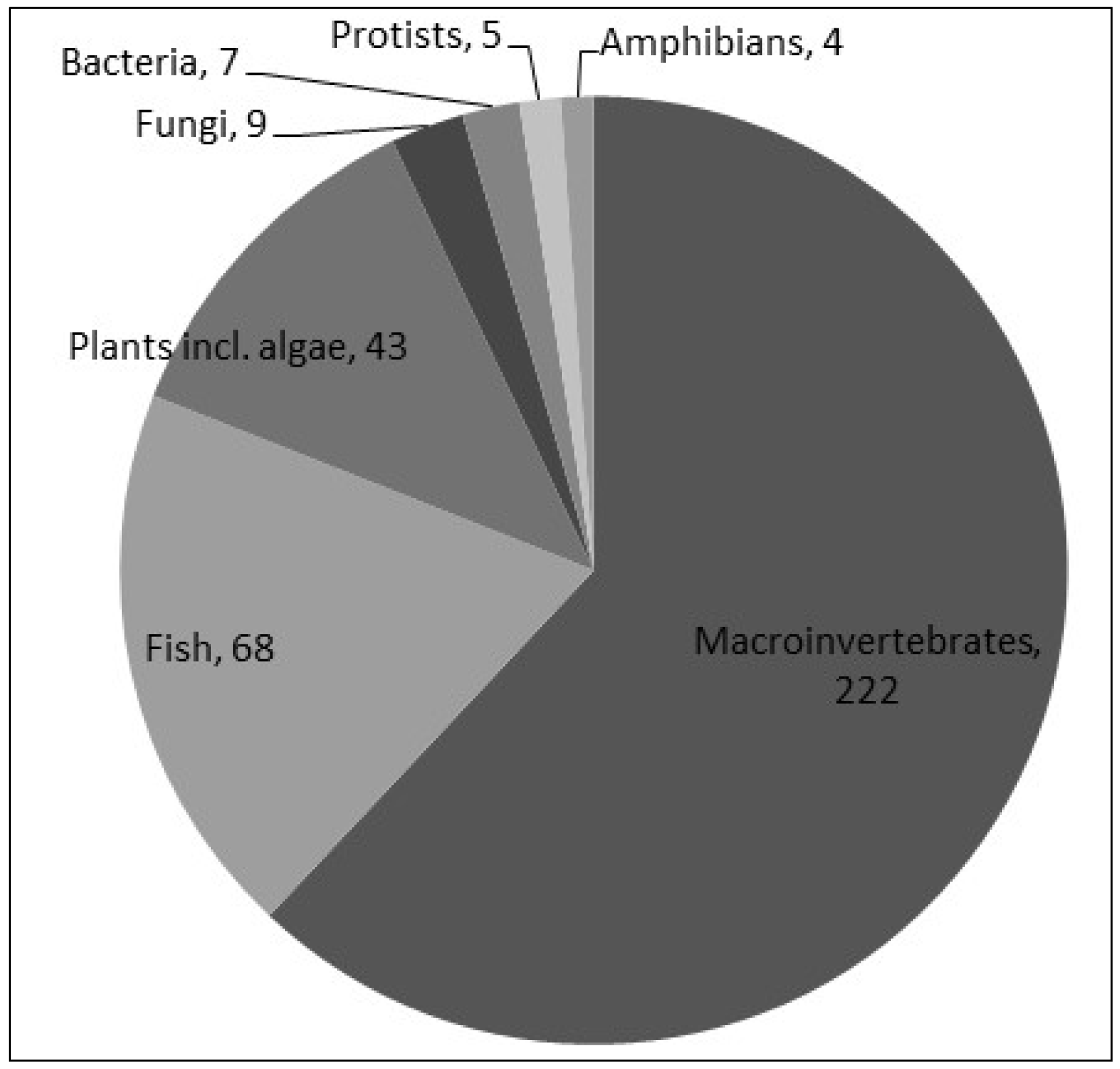
| Organism? | Text (e.g., “macroinvertebrate”) | Type of organisms for which biodiversity was quantified | |
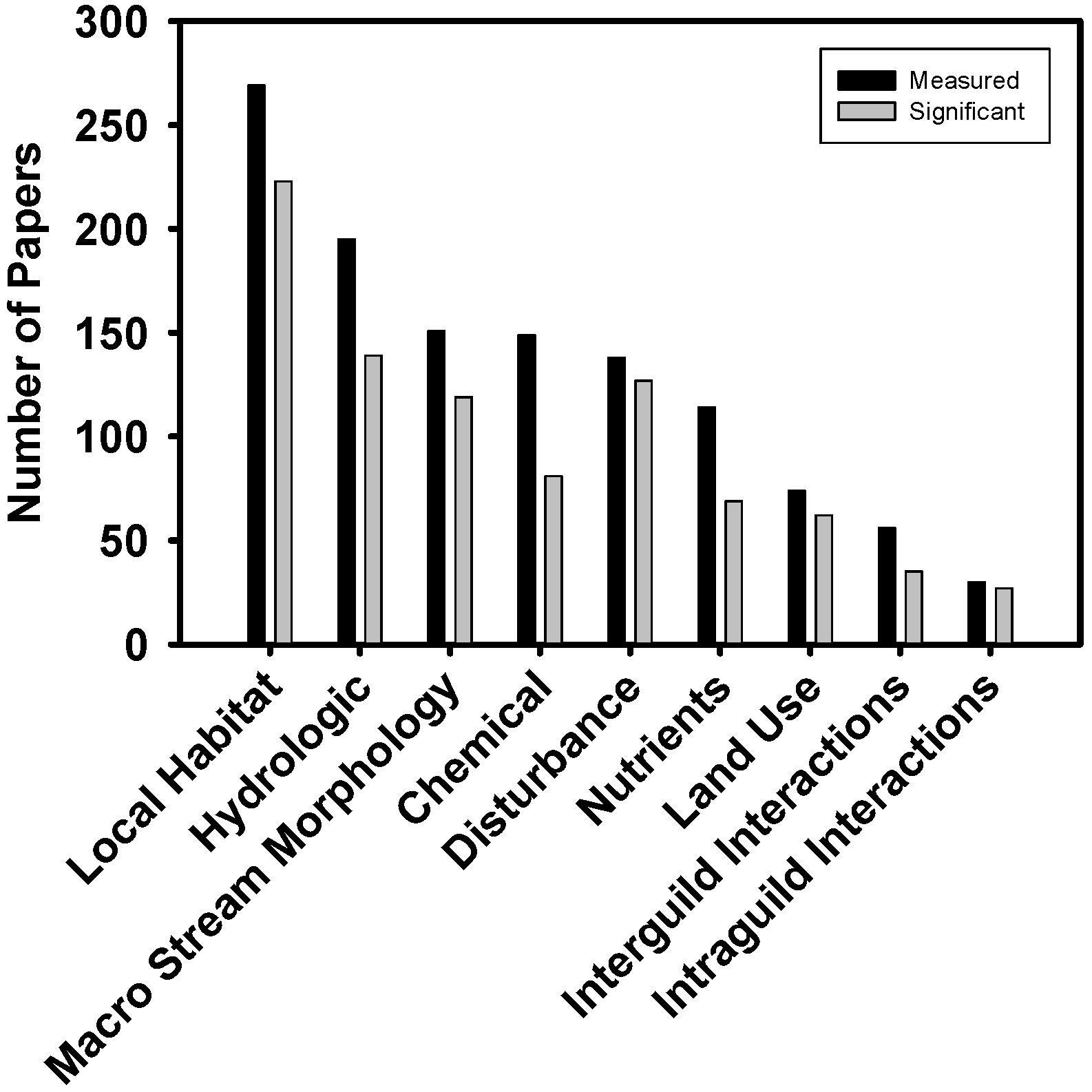
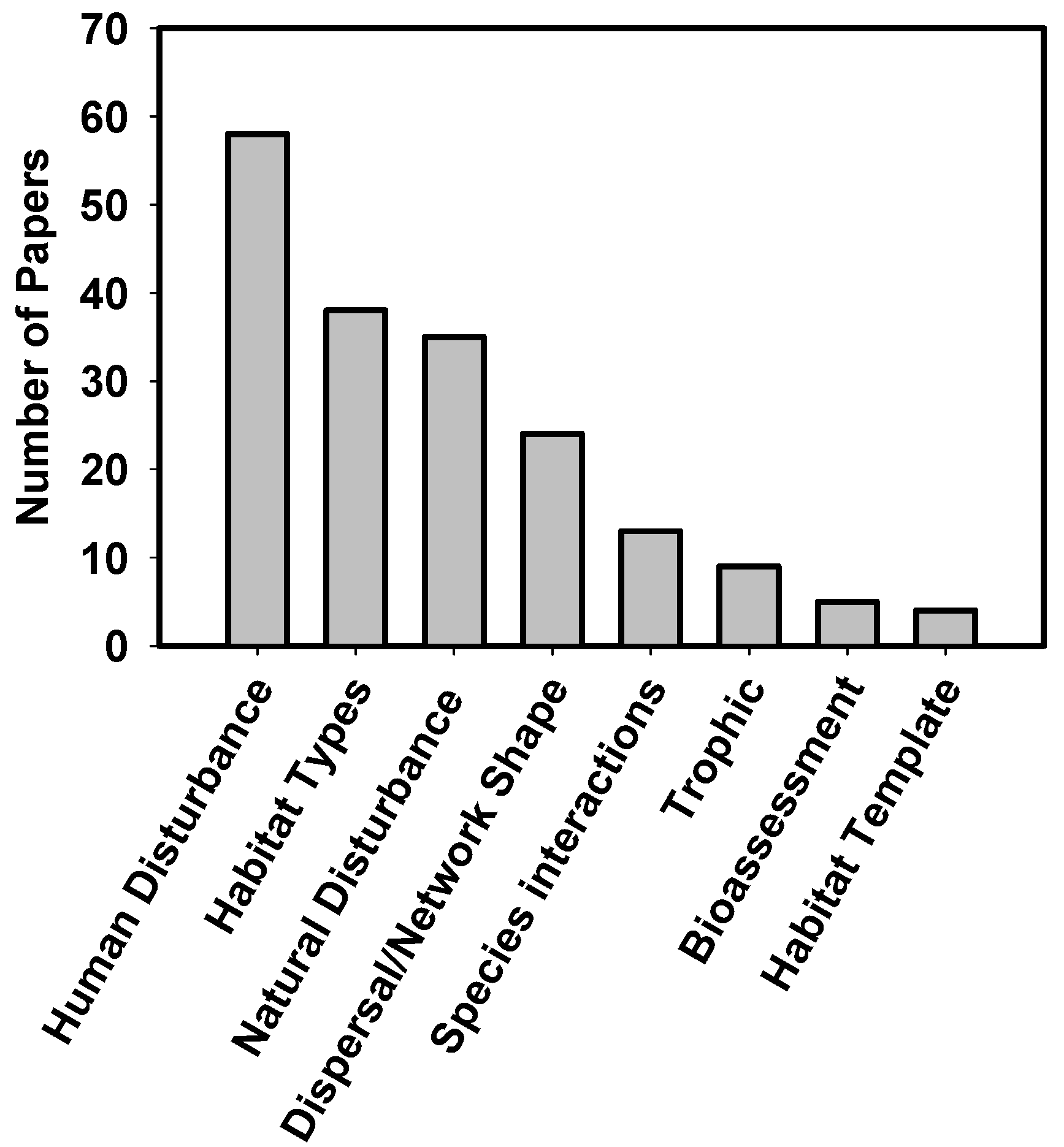
| Predictors of biodiversity | Theme | Text | A brief description of the study theme |
| Chemical | {0, 1, 2} * | Water chemistry variables (e.g., pH, conductivity, non-nutrient solute concentrations) | |
| Disturbance | {0, 1, 2} * | Was disturbance quantified to predict biodiversity? (e.g., heavy metal concentrations caused by mining operations) | |
| Hydrologic | {0, 1, 2} * | Measures of local hydrology (e.g., discharge, hydroperiod) | |
| Land use/cover | {0, 1, 2} * | Land cover variables (e.g., land use categories quantified using GIS) | |
| Local Habitat | {0, 1, 2} * | Physical reach scale variables (e.g., temperature, substrate complexity) | |
| Macro-scale stream morphology | {0, 1, 2} * | Broad scale descriptions of stream, river, or catchment (e.g., stream order, catchment size) | |
| Nutrients | {0, 1, 2} * | Nutrient concentrations or availability (e.g., N or P concentrations, nutrient diffusing substrates) | |
| Species interactions-INTRAguild | {0, 1, 2} * | Were intra-guild biotic interactions invoked to describe biodiversity (e.g., competitive exclusion)? | |
| Species interactions-INTERguild | {0, 1, 2} * | Were inter-guild biotic interactions invoked to describe biodiversity (e.g., food availability, predation) | |
| Other | {0, 1, 2} * | Other predictors of biodiversity not described above |
3. Results and Discussion








4. Discussion
4.1. What are the Major Questions that Investigators Asked with Regard to Diversity in Lotic Systems?
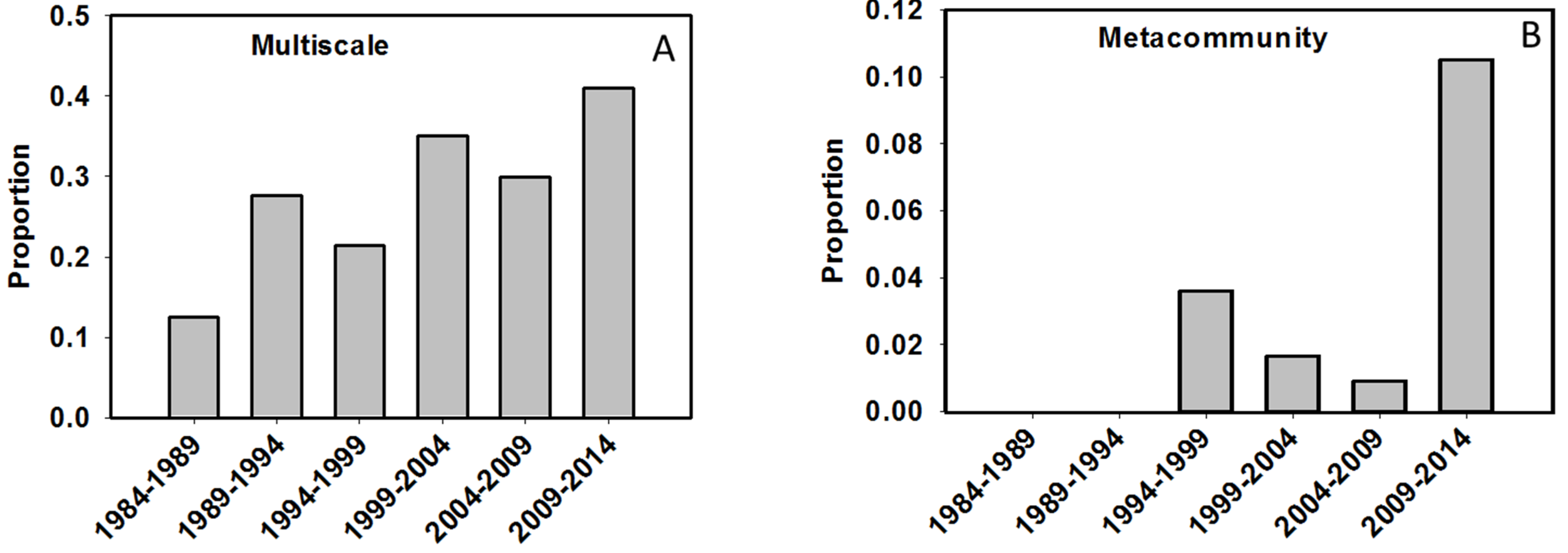
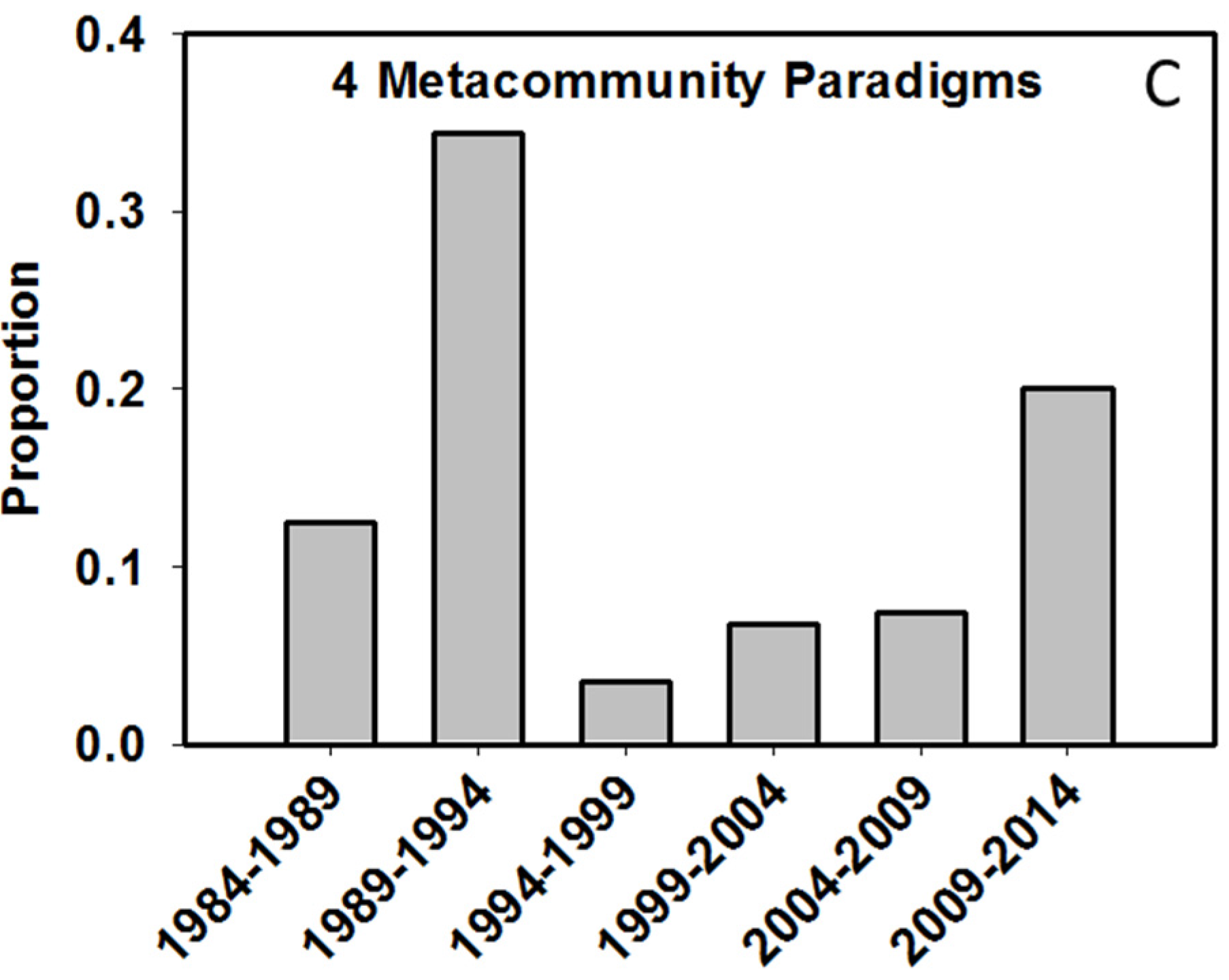
4.2. At What Spatial Scales are Researchers Investigating Biodiversity?
5. Conclusions
Acknowledgments
Author Contributions
Supplementary Materials
Conflicts of Interest
References and Notes
- Darwin, C. On the Origins of Species by Means of Natural Selection; Murray: London, UK, 1859. [Google Scholar]
- Haeckel, E.H. Generelle Morphologie der Organismen Allgemeine Grundzuge der Organischen Formen-Wissenschaft, Mechanisch Begrundet Durch die von Charles Darwin Reformirte Descendenz-Theorie von Ernst Haeckel: Allgemeine Entwickelungsgeschichte der Organismen Kritische Grundzuge der Mechanischen Wissenschaft von den Entstehenden Formen der Organismen, Begrundet Durch die Descendenz-Theorie; Verlag von Georg Reimer: Berlin, Germany, 1866; Volume 2. (In Germany) [Google Scholar]
- Schulze, E.-D.; Mooney, H.A. Biodiversity and Ecosystem Function: With 22 Tables; Springer: Ann Arbor, MI, USA, 1994; Volume 99. [Google Scholar]
- Cottingham, K.L.; Brown, B.L.; Lennon, J.T. Biodiversity may regulate the temporal variability of ecological systems. Ecol. Lett. 2001, 4, 72–85. [Google Scholar] [CrossRef]
- Tilman, D.; Downing, J. Biodiversity and stability in grasslands. In Ecosystem Management; Springer: New York, NY, USA, 1996; pp. 3–7. [Google Scholar]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Costanza, R.; d’Arge, R.; Groot, R.D.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world's ecosystem services and natural capital. Nature 1997, 387, 254–260. [Google Scholar]
- Daily, G.C.; Alexander, S.; Erlich, P.R.; Goulder, L.; Lubchenco, J.; Matson, P.A.; Mooney, H.A.; Postel, S.; Schneider, S.H.; Tilman, D.; et al. Ecosystem services: Benefits supplied to human societies by natural ecosystems. Issues Ecol. 1997, 2, 1–16. [Google Scholar]
- Ostfeld, R.S.; Keesing, F. Effects of host diversity on infectious disease. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 157–182. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Allan, J.D.; Flecker, A.S. Biodiversity conservation in running waters. Bioscience 1993, 43, 32–42. [Google Scholar] [CrossRef]
- Ward, J.V. Riverine landscapes: Biodiversity patterns, disturbance regimes, and aquatic conservation. Biol. Conserv. 1998, 83, 269–278. [Google Scholar] [CrossRef]
- Sheldon, A.L. Conservation of stream fishes: Patterns of diversity, rarity, and risk. Conserv. Biol. 1988, 2, 149–156. [Google Scholar] [CrossRef]
- Peterson, B.J.; Wollheim, W.M.; Mulholland, P.J.; Webster, J.R.; Meyer, J.L.; Tank, J.L.; Martí, E.; Bowden, W.B.; Valett, H.M.; Hershey, A.E. Control of nitrogen export from watersheds by headwater streams. Science 2001, 292, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Poff, N.L.; Olden, J.D.; Merritt, D.M.; Pepin, D.M. Homogenization of regional river dynamics by dams and global biodiversity implications. Proc. Natl. Acad. Sci. USA 2007, 104, 5732–5737. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D. Asian river fishes in the anthropocene: Threats and conservation challenges in an era of rapid environmental change. J. Fish Biol. 2011, 79, 1487–1524. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.L.; Swan, C.M.; Auerbach, D.A.; Campbell Grant, E.H.; Hitt, N.P.; Maloney, K.O.; Patrick, C. Metacommunity theory as a multispecies, multiscale framework for studying the influence of river network structure on riverine communities and ecosystems. J. N. Am. Benthol. Soc. 2011, 30, 310–327. [Google Scholar] [CrossRef]
- Heino, J.; Peckarsky, B.L. Integrating behavioral, population and large-scale approaches for understanding stream insect communities. Curr. Opin. Insect Sci. 2014, 2, 7–13. [Google Scholar] [CrossRef]
- Resh, V.H.; Brown, A.V.; Covich, A.P.; Gurtz, M.E.; Li, H.W.; Minshall, G.W.; Reice, S.R.; Sheldon, A.L.; Wallace, J.B.; Wissmaar, R.C. The role of disturbance in stream ecology. J. N. Am. Benthol. Soc 1988, 7, 433–455. [Google Scholar] [CrossRef]
- Leigh, C.; Stewart-Koster, B.; Sheldon, F.; Burford, M.A. Understanding multiple ecological responses to anthropogenic disturbance: Rivers and potential flow regime change. Ecol. Appl. 2011, 22, 250–263. [Google Scholar] [CrossRef]
- Astorga, A.; Heino, J.; Luoto, M.; Muotka, T. Freshwater biodiversity at regional extent: Determinants of macroinvertebrate taxonomic richness in headwater streams. Ecography 2011, 34, 705–713. [Google Scholar] [CrossRef]
- Leung, A.S.L.; Dudgeon, D. Scales of spatiotemporal variability in macroinvertebrate abundance and diversity in monsoonal streams: Detecting environmental change. Freshw. Biol. 2011, 56, 1193–1208. [Google Scholar] [CrossRef]
- Davidson, T.A.; Mackay, A.W.; Wolski, P.; Mazebedi, R.; Murray-Hudson, M.; Todd, M. Seasonal and spatial hydrological variability drives aquatic biodiversity in a flood-pulsed, sub-tropical wetland. Freshw. Biol. 2012, 57, 1253–1265. [Google Scholar] [CrossRef]
- Robinson, C.T.; Uehlinger, U.; Monaghan, M.T. Effects of a multi-year experimental flood regime on macroinvertebrates downstream of a reservoir. Aquat. Sci. 2003, 65, 210–222. [Google Scholar] [CrossRef]
- Brown, B.L.; Lawson, R.L. Habitat heterogeneity and activity of an omnivorous ecosystem engineer control stream community dynamics. Ecology 2010, 91, 1799–1810. [Google Scholar] [CrossRef] [PubMed]
- Fagan, W.F. Connectivity, fragmentation, and extinction risk in dendritic metapopulations. Ecology 2002, 83, 3243–3249. [Google Scholar] [CrossRef]
- Blanchet, F.G.; Legendre, P.; Maranger, R.; Monti, D.; Pepin, P. Modelling the effect of directional spatial ecological processes at different scales. Oecologia 2011, 166, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.L.; Swan, C.M. Dendritic network structure constrains metacommunity properties in riverine ecosystems. J. Anim. Ecol. 2010, 79, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.H.C.; Lowe, W.H.; Fagan, W.F. Living in the branches: Population dynamics and ecological processes in dendritic networks. Ecol. Lett. 2007, 10, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Grönroos, M.; Heino, J.; Siqueira, T.; Landeiro, V.L.; Kotanen, J.; Bini, L.M. Metacommunity structuring in stream networks: Roles of dispersal mode, distance type, and regional environmental context. Ecolo. Evol. 2013, 3, 4473–4487. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Flecker, A.S.; Hoeinghaus, D.J. Patch dynamics and environmental heterogeneity in lotic ecosystems. J. N. Am. Benthol. Soc. 2010, 29, 84–99. [Google Scholar] [CrossRef]
- Heino, J. A macroecological perspective of diversity patterns in the freshwater realm. Freshw. Biol. 2011, 56, 1703–1722. [Google Scholar] [CrossRef]
- Heino, J.; Muotka, T.; Paavola, R. Determinants of macroinvertebrate diversity in headwater streams: Regional and local influences. J. Anim. Ecol. 2003, 72, 425–434. [Google Scholar] [CrossRef]
- Finn, D.S.; Bonada, N.; Múrria, C.; Hughes, J.M. Small but mighty: Headwaters are vital to stream network biodiversity at two levels of organization. J. N. Am. Benthol. Soc. 2011, 30, 963–980. [Google Scholar] [CrossRef]
- Hitt, N.P.; Angermeier, P.L. Fish community and bioassessment responses to stream network position. J. N. Am. Benthol. Soc. 2011, 30, 296–309. [Google Scholar] [CrossRef]
- Swan, C.M.; Brown, B.L. Using rarity to infer how dendritic network structure shapes biodiversity in riverine communities. Ecography 2014, 37, 993–1001. [Google Scholar] [CrossRef]
- Sokol, E.R.; Benfield, E.F.; Belden, L.K.; Valett, H.M. The assembly of ecological communities inferred from taxonomic and functional composition. Am. Nat. 2011, 177, 630–644. [Google Scholar] [CrossRef] [PubMed]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The flood pulse concept in river-floodplain systems. Can. Spec. Publ. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Ward, J.V.; Stanford, J.A. The serial discontinuity concept of lotic ecosystems. In Dynamics of Lotic Ecosystems; Fontaine, T.D., III, Bartell, S.M., Eds.; Ann Arbor Science: Ann Arbor, MI, USA, 1983; pp. 29–42. [Google Scholar]
- Ward, J. The four-dimensional nature of lotic ecosystems. J. N. Am. Benthol. Soc. 1989, 8, 2–8. [Google Scholar] [CrossRef]
- Heino, J.; Schmera, D.; Erős, T. A macroecological perspective of trait patterns in stream communities. Freshw. Biol. 2013, 58, 1539–1555. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, N.; Izard, J.; Klatt, C.; Zhou, J.; Aronson, E. The unseen world: Environmental microbial sequencing and identification methods for ecologists. Front. Ecol. Environ. 2014, 12, 224–231. [Google Scholar] [CrossRef]
- Simberloff, D.; von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invas. 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Møller, A.P.; Jennions, M.D. Testing and adjusting for publication bias. Trends Ecol. Evol. 2001, 16, 580–586. [Google Scholar] [CrossRef]
- Johnson, M.T.; Stinchcombe, J.R. An emerging synthesis between community ecology and evolutionary biology. Trends Ecol. Evol. 2007, 22, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Finn, D.S.; Poff, N.L. Examining spatial concordance of genetic and species diversity patterns to evaluate the role of dispersal limitation in structuring headwater metacommunities. J. N. Am. Benthol. Soc. 2011, 30, 273–283. [Google Scholar] [CrossRef]
- Lowe, W.; McPeek, M. Can natural selection maintain long-distance dispersal? Insight from a stream salamander system. Evol. Ecol. 2012, 26, 11–24. [Google Scholar]
- Levin, S.A. The problem of pattern and scale in ecology. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Avois-Jacquet, C.; Tuomisto, H. Dissecting the spatial structure of ecological data at multiple scales. Ecology 2004, 85, 1826–1832. [Google Scholar] [CrossRef]
- Cooper, S.D.; Diehl, S.; Kratz, K.; Sarnelle, O. Implications of scale for patterns and processes in stream ecology. Austr. J. Ecol. 1998, 23, 27–40. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar]
- Chase, J.M. Community assembly: When should history matter? Oecologia 2003, 136, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.M. Towards a really unified theory for metacommunities. Funct. Ecol. 2005, 19, 182–186. [Google Scholar] [CrossRef]
- Cottenie, K. Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E. Naturalists, natural history, and the nature of biological diversity. Am. Nat. 2012, 179, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Heino, J. Metacommunity patterns of highly diverse stream midges: Gradients, chequerboards, and nestedness, or is there only randomness? Ecol. Entomol. 2005, 30, 590–599. [Google Scholar] [CrossRef]
- Heino, J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biol. Rev. 2013, 88, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Patrick, C.J.; Swan, C.M. Reconstructing the assembly of a stream-insect metacommunity. J. N. Am. Benthol. Soc. 2011, 30, 259–272. [Google Scholar] [CrossRef]
- Siqueira, T.; Bini, L.M.; Roque, F.O.; Marques Couceiro, S.R.; Trivinho-Strixino, S.; Cottenie, K. Common and rare species respond to similar niche processes in macroinvertebrate metacommunities. Ecography 2012, 35, 183–192. [Google Scholar] [CrossRef]
- Auerbach, D.A.; Poff, N.L. Spatiotemporal controls of simulated metacommunity dynamics in dendritic networks. J. N. Am. Benthol. Soc. 2011, 30, 235–251. [Google Scholar] [CrossRef]
- Funk, A.; Schiemer, F.; Reckendorfer, W. Metacommunity structure of aquatic gastropods in a river floodplain: The role of niche breadth and drift propensity. Freshw. Biol. 2013, 58, 2505–2516. [Google Scholar] [CrossRef]
- Göthe, E.; Angeler, D.G.; Sandin, L. Metacommunity structure in a small boreal stream network. J. Anim. Ecol. 2013, 82, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.M. Fish species richness and incidence patterns in isolated and connected stream pools: Effects of pool volume and spatial position. Oecologia 1997, 110, 560–566. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tornwall, B.; Sokol, E.; Skelton, J.; Brown, B.L. Trends in Stream Biodiversity Research since the River Continuum Concept. Diversity 2015, 7, 16-35. https://doi.org/10.3390/d7010016
Tornwall B, Sokol E, Skelton J, Brown BL. Trends in Stream Biodiversity Research since the River Continuum Concept. Diversity. 2015; 7(1):16-35. https://doi.org/10.3390/d7010016
Chicago/Turabian StyleTornwall, Brett, Eric Sokol, James Skelton, and Bryan L. Brown. 2015. "Trends in Stream Biodiversity Research since the River Continuum Concept" Diversity 7, no. 1: 16-35. https://doi.org/10.3390/d7010016