
Winter Territoriality of the American Redstart in Oil Palm Plantations

College of Forest Resources and Environmental Sciences, Michigan Technological University, Houghton, MI 49931, USA
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(12), 1079; https://doi.org/10.3390/d14121079
Submission received: 7 November 2022
/
Revised: 30 November 2022
/
Accepted: 5 December 2022
/
Published: 7 December 2022
(This article belongs to the Special Issue Avian Ecology and Diversity, Population Monitoring and Conservation: Recent Advances and New Challenges)
Abstract
:Aspects of territorial behavior of Nearctic-neotropical migratory birds during the nonbreeding period are poorly studied. Information about territoriality, site persistence, between-year site fidelity, and territory sizes are not available for most birds, especially in tropical agroecosystems. Given the rapid expansion of oil palm in the neotropics, determining how oil palm affects the territorial behaviors of overwintering migratory birds is an important line of inquiry with conservation implications. The American Redstart (Setophaga ruticilla) is considered a model species for the study of population dynamics in the neotropics; however, territory size for American Redstart has only been assessed in native habitats. In this study, we outfitted individual redstarts with radio tags, across two winter seasons, to determine variation in territory sizes across oil palm plantations and native forest patches in the State of Tabasco, Mexico. Average redstart territory size was 0.29 ha in oil palm plantations and 0.17 ha in native forest. Albeit presenting larger territories in oil palm plantations, which could indicate poorer habitat quality, the difference between both habitats was not statistically significant. Our results demonstrate, for the first time, that American Redstarts hold territories in oil palm plantations and that territory size may serve as an important indicator of relative habitat quality for redstart populations in tropical working landscapes.
1. Introduction
Most Nearctic-neotropical migratory birds tend to adopt territorial behavior during the breeding season as well as between-year site fidelity to breeding territories [1]. Using the same site likely provides energetic advantages due to previous knowledge of the area, predator avoidance, and increased foraging success [2]. Relative to territoriality on the breeding grounds, the degree of territorial behaviors on the wintering grounds is more diverse, where some species routinely defend territories, others are facultatively territorial, while some exhibit no territoriality at all [3,4,5,6].
One neotropical migrant subject to a relatively large amount of study during the non-breeding season is the American Redstart (Setophaga ruticilla) which provided one of the earliest examples of carry-over effects, where habitat quality on the wintering grounds influenced reproductive success on the breeding grounds [7,8]. In the tropics, redstarts display territoriality and adult male dominance, whereby adult males often exclude females and young males from high-quality habitats like mangroves [7,9,10,11,12] and primary and secondary forests [13,14,15]. Subordinate redstarts are often relegated to suboptimal habitats [16] where they suffer from decreased fitness, characteristic of an ideal despotic distribution [17]. Moreover, experiments demonstrated that females and young males colonize the territories of dominant males after their removal from mangroves and dry limestone forest sites in Jamaica resulting in improved fitness [9,11].
Given that despotic individuals competitively exclude conspecifics from high quality habitats, measures of density could be misleading indicators of habitat quality, as high quality habitats may, in theory, have lower densities at small spatial scales [18]. Alternative indicators of habitat quality include distributional and individual measures, such as sex and age ratios, indices of fitness (e.g., body condition index, fat, and muscle scores), spring departure date, seasonal survival rate, site persistence, rate of transient behavior, and territory size. Specifically, transient behavior occurs when otherwise territorial birds do not hold territories thereby acting as “floaters” within a wintering population [19]. Transient American Redstarts tend to be females and young individuals of both sexes [12,20,21] and are more common in poorer quality habitats [22,23,24]. Additionally, individuals in high-quality habitats move less distance to acquire food resources thereby resulting in an inverse correlation between habitat quality and territory size [25,26,27].
Although American Redstarts tend to select undisturbed forest habitats during the winter [28], populations will use a diversity of other habitats, from forest edges, young secondary forests, and regenerating pasture [29,30] as well as shade coffee plantations in Puerto Rico, Jamaica and Venezuela [14,20,31], cacao plantations in Mexico [32], citrus farms in Jamaica [31], and oil palm plantations in southern Mexico [33,34]. Oil palm cultivation is a rapidly expanding agroecosystem in the neotropics where it often replaces secondary forest or pasture [35]. To our knowledge, no information is available regarding territory size of American Redstarts in oil palm plantations. Understanding how territory size varies between forest and oil palm plantation habitats can provide insight into the value of oil palm to wintering birds; such information informs long-term conservation strategies for migratory species in human-modified landscapes [36]. In this study, we aimed to: (1) determine if American Redstarts held territories in oil palm plantations, (2) estimate territory size, and (3) compare territory size between oil palm and native forest habitats. Given territoriality and dominance hierarchies exhibited by redstarts in other parts of their winter range, we predicted that birds would maintain territories in both habitats and that individuals in lower quality habitat (i.e., oil palm plantations) would exhibit larger territories relative to higher quality forest (i.e., native forest), due to lower food resource availability.
2. Materials and Methods
This work took place in southeast Mexico in the state of Tabasco where the original moist tropical forest has largely been converted to cattle pasture, known as “potreros”, as well as crops like banana, corn, hardwood, and most recently oil palm plantations [37,38]. This process resulted in just a few secondary forest remnants in lower elevations. The climate in the region is characterized by an average temperature of 24° to 28 °C and precipitation ranging from 2500 to 4000 mm [39]. We collected data during two winter seasons, 2017–2018 and 2018–2019, and most of the fieldwork took place during the stationary migratory period (November–February) in the municipalities of Jalapa, Tacotalpa, and Teapa. Study areas included one secondary native forest site (17.6495° N, −92.8146° W) and three oil palm plantations (17.5788° N, −92.8526° W; 17.6992° N, −92.9488° W; 17.6473° N, −92.8304° W). The secondary forest patch was 60 ha and isolated from other woodlands; oil palm plantations ranged from 46 to 84 ha (mean 63.3), were characterized by similar age (~12 years), and presented no adjacent contact with native forest.
Mist-netting was used to capture birds to assess habitat quality through various indicators, including species abundance, community composition, fat loads, mass, and age and sex segregation [15,34]. Concurrently with bird captures (see Oliveira et al. [34]), we outfitted 16 American Redstarts with 0.3 g radio telemetry transmitters (Advanced Telemetry Systems model no. 2414) attached using eyelash glue on the trimmed lower back of the birds. To increase the chance that we were putting transmitters on territorial individuals, we only attached the tags on birds recaptured at least once on the same mist-net line. Tracked individuals were also color banded with a unique color combination to facilitate identification by resighting.
After release, birds were relocated using handheld radio receivers (ATS model 410) and a three-element Yagi antenna (ATS model 148-152). Because American Redstart relocation methodology was mainly constrained by the 14-day battery life of the transmitter, we based our tracking on burst sampling [40]. However, we spaced the interval between relocations every 5 min to give individuals enough time to move to any point in their territory. Cooper and colleagues [41] suggested that because American Redstarts have relatively small territories in the winter, 30-s intervals between locations is sufficient for sampling independence. We tracked tagged birds until we could confirm a location visually or until the signal strength was sufficient that we were confidently within 10 m of the bird and recorded a GPS point (Universal Transverse Mercator). For birds without visual confirmation, we used a compass to determine an azimuth towards the individual based on radio signal to project the proximate point. Locations were recorded using a Garmin GPS unit (model GPSMAP 64). To minimize GPS error, we only recorded coordinates when the device showed an accuracy of ≤5 m. We relocated the individuals no less than three days apart and collected no more than five points per day. We stopped tracking individuals when the transmitter battery died, accidentally fell from the bird’s back, no signal was located in the territory or within a 150 m radius from the last location.
We estimated territory size for individuals with at least 20 GPS relocations using a minimum convex polygon calculated by the package “adehabitatHR” [42] in program R [43]. Our sample size was constrained by the number of field assistants trained for radio telemetry, distance among sites, and the transmitters’ battery life. Although there is no consensus for a minimum sample size when using the minimum convex polygon, the number of relocations can influence the territory size estimation as the method is sensitive to sample sizes and outliers [44]. Differences in territory size between oil palm plantations and native forest was compared using a t-test in program R [43]. Normality of the dataset’s distribution was assessed prior to conducting the t-test [45].
3. Results
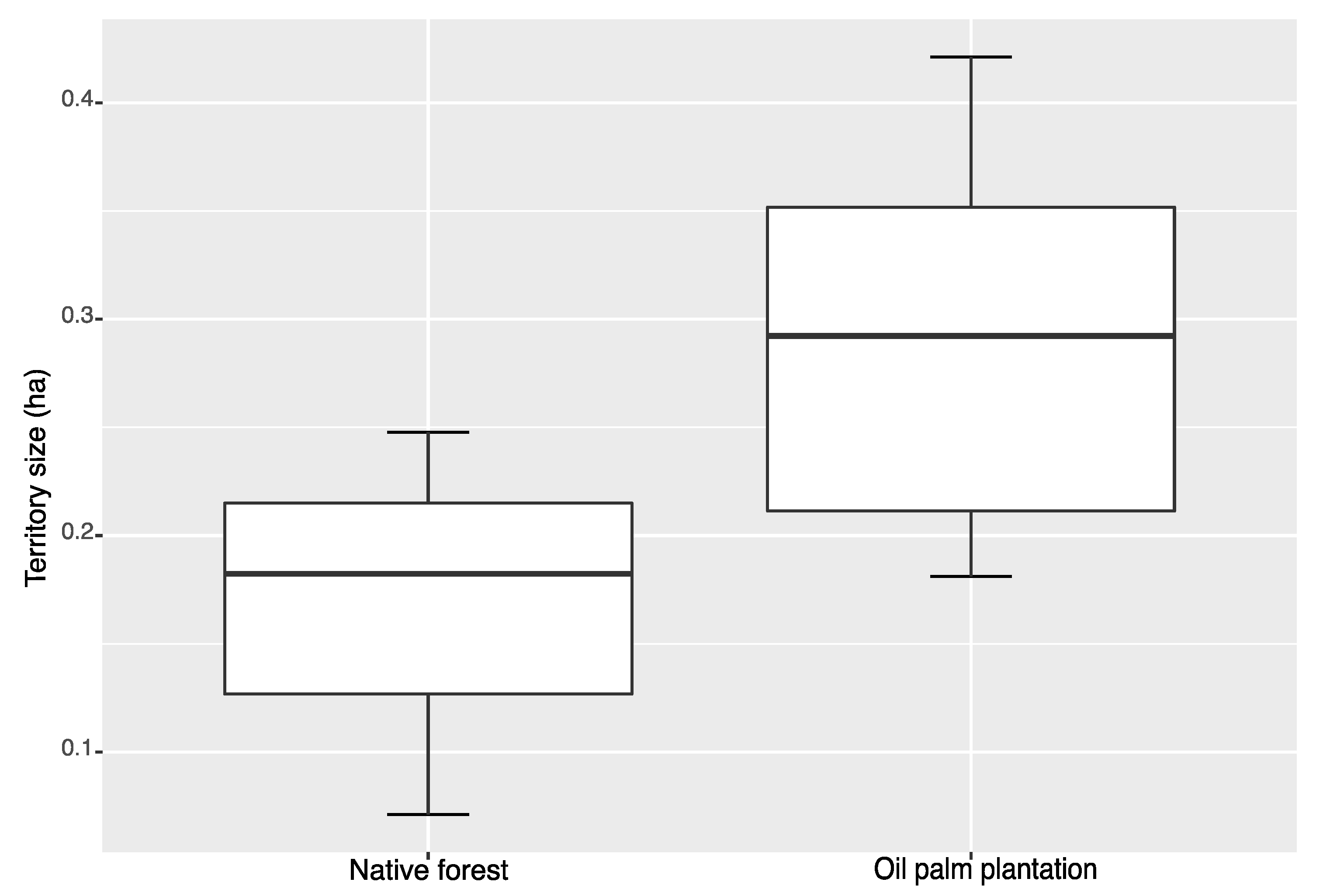
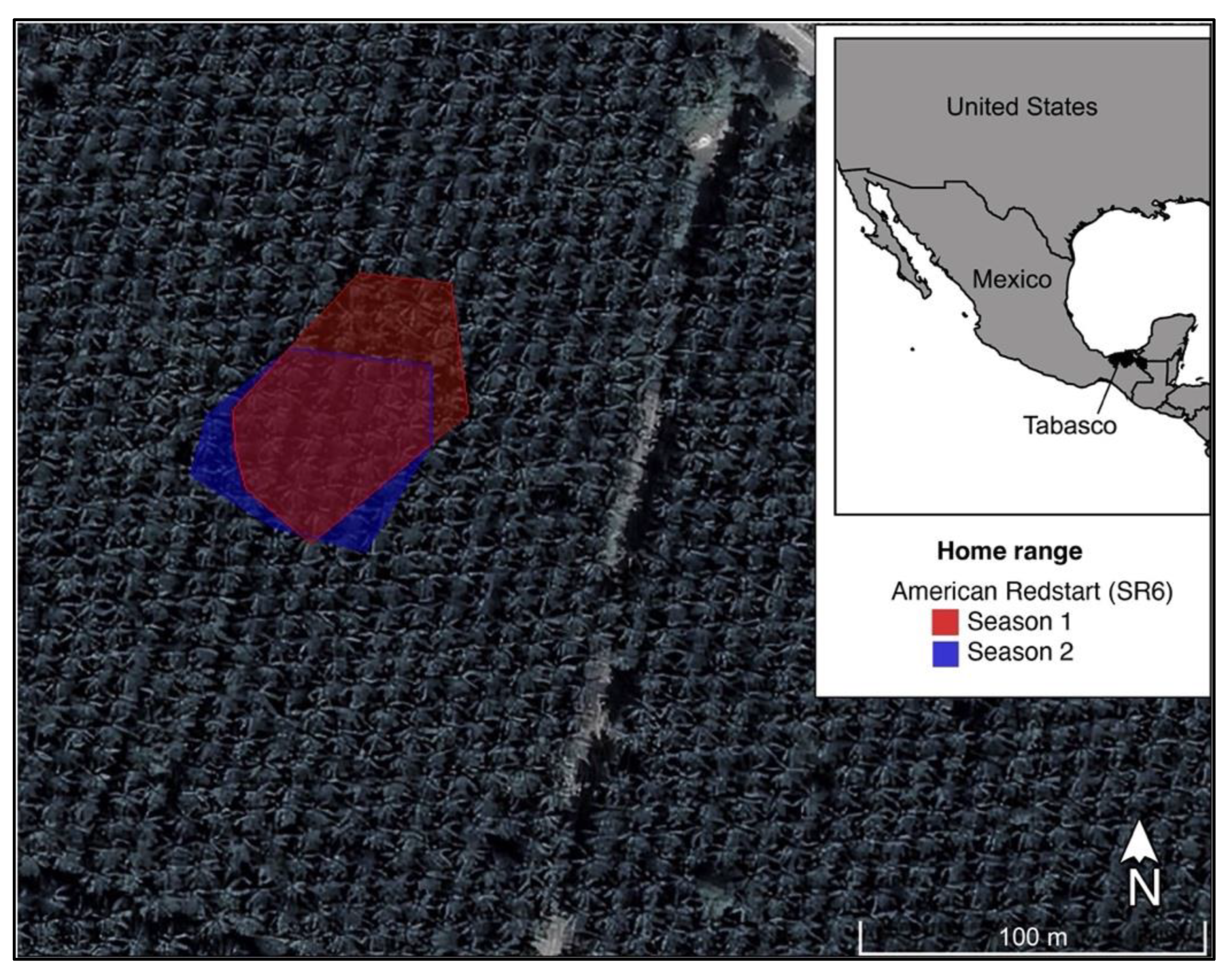
From the 16 individuals outfitted with radios, we were able to collect a minimum of 20 GPS point relocations (mean 30.7) for 10 birds. No females were captured at our native forest site. This way, we opted to use only male redstarts in our analysis, thereby controlling for sex variation. We estimated territory size for seven individuals in oil palm plantations and three individuals in the native forest patch. Overall territory size, including both native forest and oil palm plantations, was 0.25 ha (±0.033 SE). Territory size ranged from 0.18 to 0.42 ha (mean 0.29 ± 0.035 SE) in oil palm plantations, and from 0.07 to 0.24 ha (mean 0.17 ± 0.052 SE) in native forest (Table 1). Our results suggested that, on average, American Redstarts had larger territories in oil palm plantations than in native forest patches (Figure 1); however, the difference was not significant (t-value = −1.93, p = 0.09). Two individuals captured during the first season were recovered in the second season, both in oil palm plantations. The rate of inter-annual site fidelity could not be assessed with the relatively small sample size; however, we did estimate the territory size of one individual in both seasons. The bird was a young (first-cycle formative, FCF, following WRP; [46]) male during the first season and held a 0.34 ha territory, while in our second field season the territory was similarly sized at 0.29 ha (Figure 2). Additionally, although a year apart, the two home ranges included significant spatial overlap based on the minimum convex polygons (Figure 3).
4. Discussion
Although considered a suboptimal habitat for the American Redstarts in the wintering grounds, oil palm plantations were still commonly used whereby multiple individuals established territories [15,34]. Winter territoriality and sexual segregation have been shown to exist in American Redstarts overwintering in more natural forested habitats in the neotropics [9,31]. Our study is the first to identify territorial behavior in a non-native monoculture surrounded by other human-modified land covers, broadening our understanding of the diversity of habitats used by migratory birds on the wintering grounds.
Because they have been shown to arrange themselves according to an ideal despotic distribution, the density of redstarts at small spatial scales should not be used to assess habitat quality. This is a common misconception, especially for generalist birds with broader habitat tolerances [21]. In the case of the American Redstart, prey availability, influenced in part by humidity, play an important role in habitat quality for individuals inhabiting different forest types [41,47]. In Venezuelan shade coffee [14], citrus orchards in Jamaica [48], and oil palm plantations in Mexico [34,49], the observed higher densities of Redstarts, when compared with native vegetation could lead one to believe that these agroecosystems provide similar or higher quality habitats compared to native forest. Further assessments in citrus and oil palm plantations, using distributional and individual indicators, showed that these monocultures likely serve as poorer habitats for American Redstarts, as suggested by significantly lower body condition, sex and age segregation, and differences in territory size ([15,48], Figure 1).
Our overall territory size estimates, including native forest and oil palm plantations, was 0.25 ha, similar to the values reported for native vegetation in Jamaica, 0.25 and 0.3 ha [50,51] and Puerto Rico, 0.39 ha [23]. The mean territory size in the Tabascan native forest (0.17 ha) was also close to the value reported by Cooper et al. [41] in Jamaica, 0.16 ha. The area defended by territorial birds can be used as a proximate cue to assess food availability [25,52,53], and ultimately habitat quality. Our results suggest that, although not statistically significant, American Redstart territory size tended to be larger in oil palm plantations than in native forests. Despite a small sample size, we observed a pattern suggesting that territory size varies between oil palm plantations and native forests, warranting further studies that directly assess habitat quality (e.g., food abundance and survival). This pattern of higher quality habitats resulting in smaller territories has been repeatedly shown in other Neotropical migrants. For example, overwintering Wilson’s Warblers (Cardellina pusilla) in Veracruz, Mexico, defended smaller territories in protected forests than in disturbed lowland forest-covered sites [54]. On the breeding grounds, Red-eyed Vireos in West Virginia defended smaller territories in areas with more caterpillars [55]. Likewise, Ovenbirds in eastern Tennessee, held smaller territories in sites with higher prey abundance [52]. Conversely, sedentary Ovenbirds did not exhibit transient behavior, expand territory size, or move their territories when their food supply was experimentally reduced in Jamaica [56]. Similarly, in an experiment with food availability reduction in Jamaica, the American Redstarts showed no increase in territory size to acquire more food resources but instead became floaters (non-territorial) [41]. This suggests that other mechanisms may also influence territory size, such as agonistic interactions with adjacent sedentary individuals.
Inter-season winter site fidelity has been observed among American Redstarts [4,20,21,28,48,50,57,58]. Our work is the first to describe this behavior in oil palm plantations with the recovery of two individuals captured in the first winter season. Furthermore, the territory overlap exhibited by one individual monitored in two consecutive seasons suggests that American Redstarts exhibit high site fidelity even in what appear to be relatively poor-quality habitats, like the oil palm plantations [15]. Between-year site fidelity is considered advantageous as a result of individuals returning to a familiar site [1,2,59]. The rate of between-year return of wintering birds is expected to be highly correlated with food abundance and predictability, thus serving as an index of habitat quality [7,60,61,62]. Further long-term studies assessing populations in oil palm plantations would be helpful in determining what drives the habitat quality for the American Redstarts and whether these drivers could be managed. Such understanding could be used to assess the viability of the populations using this and other human-modified landscapes.
According to Sherry and Holmes [28], the American Redstart’s preference for less disturbed forested habitats in most of its winter distribution suggests that human land use change in the wintering range has contributed to the documented decline of 12% over the last 45 years [63]. Because Nearctic-neotropical migratory birds overwinter in a region much smaller than the breeding grounds [64], the loss of quality habitat during the winter is especially troubling. Evidence indicates that some habitats occupied by redstarts in the Neotropics seem to be saturated due to elevated density [9,28,41,65]. Rushing et al. [66] suggested that reproductive success may ultimately be limited by the quality of the winter habitat. This emphasizes the importance of territory estimates, especially in agroecosystems, since the availability of high quality habitat may limit the population growth of the species.
Author Contributions
Conceptualization, S.L.O. and D.J.F.; methodology, S.L.O., D.J.F. and J.D.W.; data collection, S.L.O.; data analysis, S.L.O. and J.D.W.; data curation, S.L.O.; writing—original draft preparation, S.L.O.; writing—review and editing, J.D.W. and D.J.F.; project administration, S.L.O.; funding acquisition, D.J.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation—NSF, grant number 1243444, and the U.S. Department of Agriculture McIntire-Stennis Cooperative Forestry Research Program.
Institutional Review Board Statement
The study was approved by the Institutional Animal Care and Use Committee (IACUC).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available in a repository following the final acceptance for publication.
Acknowledgments
We thank Ena E. M. Zayas, Stefan L. A. Weis, Cesar J. V. Navarrete, Julio Cesar G. Castillo, and Lízbeth Y. Moo Culebro for the logistic support provided in Mexico. We also thank Ruth A. Regnet, Marco Antonio Torrez-Pérez, and Ingrid Tello-López for their valuable work as field technicians. We are grateful for the support provided by the Wheaton College (MA) and the College of Forest Resources and Environmental Science—Ecosystem Science Center (ESC) to cover travel and fieldwork commute costs, as well as the Michigan Technological University Graduate School for the Finishing Fellowship grant. Finally, we thank the editor and the anonymous reviewers for contributing to this manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gauthreaux, S.A.J. The Ecology and Evolution of Avian Migration Systems. In Avian Biology; Farmer, D.S., King, J.R., Eds.; Academic Press: New York, NY, USA, 1982; pp. 93–167. [Google Scholar]
- Greenwood, P.J.; Harvey, P.H. The Natal and Breeding Dispersal of Birds. Annu. Rev. Ecol. Syst. 1982, 13, 1–21. [Google Scholar] [CrossRef]
- Rappole, J.H.; Warner, D.W. Ecological Aspects of Migrant Bird Behavior in Veracruz, Mexico. In Migrant Birds in the Neotropics: Ecology, Behavior, Distribution, and Conservation; Smithsonian Institution Press: Washington, DC, USA, 1980; pp. 353–393. [Google Scholar]
- Mcneil, R. Winter Resident Repeats and Returns of Austral and Boreal Migrant Birds Banded in Venezuela. J. Field Ornithol. 1982, 53, 125–132. [Google Scholar]
- Greenberg, R. The Winter Exploitation Systems of Bay-Breasted and Chestnut-Sided Warblers in Panama; University of California Publications in Zoology: Berkeley, CA, USA, 1984; pp. 1–107. [Google Scholar]
- Albert, S.; Wolfe, J.D.; Kellerman, J.; Sherry, T.; Stutchbury, B.J.M.; Bayly, N.J.; Ruiz-Sánchez, A. Habitat Ecology of Nearctic–Neotropical Migratory Landbirds on the Nonbreeding Grounds. Condor 2020, 122, duaa055. [Google Scholar] [CrossRef]
- Marra, P.P.; Hobson, K.A.; Holmes, R.T.; Dallos, P.; Geister, C.D.; David, C. Terms and Conditions Linking Winter and Summer Events in a Migratory Bird by Using Stable-Carbon Isotopes. Science 1998, 282, 1884–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, D.R.; Marra, P.P.; Kyser, T.K.; Sherry, T.W.; Ratcliffe, L.M. Tropical Winter Habitat Limits Reproductive Success on the Temperate Breeding Grounds in a Migratory Bird. Proc. R. Soc. B Biol. Sci. 2004, 271, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marra, P.; Sherry, T.W.; Holmes, R.T. Territorial Exclusion by a Long-Distance Migrant Warbler in Jamaica: A Removal Experiment with American Redstarts (Setophaga ruticilla). Auk 1993, 110, 565–572. [Google Scholar] [CrossRef]
- Marra, P.P. The Role of Behavioral Dominance in Structuring Patterns of Habitat Occupancy in a Migrant Bird during the Nonbreeding Season. Behav. Ecol. 2000, 11, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Studds, C.E.; Marra, P.P. Nonbreeding Habitat Occupancy and Population Processes: An Upgrade Experiment with a Migratory Bird. Ecology 2005, 86, 2380–2385. [Google Scholar] [CrossRef] [Green Version]
- Peele, A.M.; Marra, P.M.; Sillett, T.S.; Sherry, T.W. Combining Survey Methods to Estimate Abundance and Transience of Migratory Birds among Tropical Nonbreeding Habitats. Auk 2015, 132, 926–937. [Google Scholar] [CrossRef]
- Ornat, A.L.; Greenberg, R. Sexual Segregation by Habitat in Migratory Warblers in Quintana Roo, Mexico. Auk 1990, 107, 539–543. [Google Scholar]
- Bakermans, M.H.; Vitz, A.C.; Rodewald, A.D.; Rengifo, C.G. Migratory Songbird Use of Shade Coffee in the Venezuelan Andes with Implications for Conservation of Cerulean Warbler. Biol. Conserv. 2009, 142, 2476–2483. [Google Scholar] [CrossRef]
- Oliveira, S.L.; Flaspohler, D.J.; Knowlton, J.L.; Wolfe, J.D. Do Oil Palm Plantations Provide Quality Habitat for Migratory Birds? A Case Study from Mexico. Ecol. Indic. 2022, 139, 108964. [Google Scholar] [CrossRef]
- Newton, I. The Migration Ecology of Birds; Academy Press: Oxford, UK, 2008; ISBN 978-0-12-517367-4. [Google Scholar]
- Fretwell, S.D.; Lucas, H.L., Jr. On Territorial Behavior and Other Factors Influencing Habitat Distribution in Birds. Acta Biotheor. 1969, 19, 16–36. [Google Scholar] [CrossRef]
- Van Horne, V. Density as a Misleading Indicator of Habitat Quality. J. Wildl. Manag. 1983, 47, 893. [Google Scholar] [CrossRef]
- Brown, D.R.; Long, J.A. What Is a Winter Floater? Causes, Consequences, and Implications for Habitat Selection. Condor 2007, 109, 548–565. [Google Scholar] [CrossRef]
- Wunderle, J.M.; Latta, S.C. Winter Site Fidelity of Nearctic Migrants in Shade Coffee Plantations of Different Sizes in the Dominican Republic. Auk 2000, 117, 596–614. [Google Scholar] [CrossRef]
- Peele, A.M. Population Regulation of a Long-Distance Migratory Bird Population in the Non-Breeding Season: A Test of Buffer and Crowding Effects. Ph.D. Thesis, Tulane University, New Orleans, LA, USA, 2015. [Google Scholar]
- Lefebvre, G.; Poulin, B.; McNeil, R. Spatial and Social Behaviour of Nearctic Warblers Wintering in Venezuelan Mangroves. Can. J. Zool. 1994, 72, 757–764. [Google Scholar] [CrossRef]
- Toms, J.D. Non-Breeding Competition between Migrant American Redstarts (Setophaga ruticilla) and Resident Adelaide’s Warblers (Dendroica adelaidae) in the Guanica Biosphere Reserve, Southwest Puerto Rico. Ph.D. Thesis, University of Missouri, Columbia, MO, USA, 2011. [Google Scholar]
- Toms, J.D. Linking Behavior and Community Ecology: Interspecific Aggression Provides Evidence for Competition Between a Migrant and Resident Warbler. Ethology 2013, 119, 1057–1066. [Google Scholar] [CrossRef]
- Stenger, J. Food Habits and Available Food of Ovenbirds in Relation to Territory Size. Auk 1958, 75, 335–347. [Google Scholar]
- Brown, J.L. The Evolution of Diversity in Avian Territorial Systems. Wilson Bull. 1964, 76, 160–169. [Google Scholar]
- Wilson, E.O. Sociobiology; Harvard University Press: Cambridge, MA, USA, 1975. [Google Scholar]
- Sherry, T.W.; Holmes, R.T. Winter Habitat Quality, Population Limitation, and Conservation of Neotropical Nearctic Migrant Birds. Ecology 1996, 77, 36–48. [Google Scholar] [CrossRef]
- Sherry, T.W.; Holmes, R.T.; Pyle, P.; Patten, M.A. American Redstart (Setophaga ruticilla). In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NJ, USA, 2020. [Google Scholar]
- Reid, J.L.; Harris, J.B.C.; Martin, L.J.; Barnett, J.R.; Zahawi, R.A. Distribution and Abundance of Nearctic–Neotropical Songbird Migrants in a Forest Restoration Site in Southern Costa Rica. J. Trop. Ecol. 2008, 24, 685–688. [Google Scholar] [CrossRef] [Green Version]
- Parrish, J.D.; Sherry, T.W. Sexual Habitat Segregation by American Redstarts Wintering in Jamaica: Importance of Resource Seasonality. Auk 1994, 111, 38–49. [Google Scholar] [CrossRef]
- Greenberg, R.; Bichier, P.; Angón, A.C. The Conservation Value for Birds of Cacao Plantations with Diverse Planted Shade in Tabasco, Mexico. Anim. Conserv. 2000, 3, 105–112. [Google Scholar] [CrossRef]
- Moo-Culebro, L.Y. Avifauna Asociada a Plantaciones de Palma Africana, Acahual y Matriz Circundante de la Sierra Tabasqueña. Master’s Thesis, Universidad Juáres Autônoma de Tabasco, Villahermosa, Mexico, 2018. [Google Scholar]
- Oliveira, S.L.; Flaspohler, D.J.; Knowlton, J.L.; Webster, C.R.; Wolfe, J.D. Migratory Bird Community Structure in Oil Palm (Elaies guineensis) Plantations and Native Forest Fragments in Southern Mexico. J. Field Ornithol. 2021, 92, 1–17. [Google Scholar] [CrossRef]
- Foster, W.A.; Snaddon, J.L.; Turner, E.C.; Fayle, T.M.; Cockerill, T.D.; Farnon Ellwood, M.D.; Broad, G.R.; Chung, A.Y.C.; Eggleton, P.; Khen, C.V.; et al. Establishing the Evidence Base for Maintaining Biodiversity and Ecosystem Function in the Oil Palm Landscapes of South East Asia. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3277–3291. [Google Scholar] [CrossRef] [Green Version]
- Hostetler, J.A.; Sillett, T.S.; Marra, P.P. Full-Annual-Cycle Population Models for Migratory Birds. Auk 2015, 132, 433–449. [Google Scholar] [CrossRef] [Green Version]
- Tudela, F. La Modernización Forzada del Trópico: El Caso de Tabasco; Colegio de México: Mexico City, Mexico, 1992. [Google Scholar]
- Arriaga-Weiss, S.L.; Calmé, S.; Kampichler, C. Bird Communities in Rainforest Fragments: Guild Responses to Habitat Variables in Tabasco, Mexico. Biodivers. Conserv. 2008, 17, 173–190. [Google Scholar] [CrossRef]
- INEGI–Instituto Nacional de Estadística y Geografía. Anuario Estadístico y Geográfico de Tabasco; INEGI: Aguascalientes, Mexico, 2015. [Google Scholar]
- Barg, J.J.; Jones, J.; Robertson, R.J. Describing Breeding Territories of Migratory Passerines: Suggestions for Sampling, Choice of Estimator, and Delineation of Core Areas. J. Anim. Ecol. 2005, 74, 139–149. [Google Scholar] [CrossRef]
- Cooper, N.W.; Sherry, T.W.; Marra, P.P. Experimental Reduction of Winter Food Decreases Body Condition and Delays Migration in a Long-Distance Migratory Bird. Ecology 2015, 96, 1933–1942. [Google Scholar] [CrossRef]
- Calenge, C. The Package “Adehabitat” for the R Software: A Tool for the Analysis of Space and Habitat Use by Animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Hansteen, T.L.; Andreassen, H.P.; Ims, R.A. Effects of Spatiotemporal Scale on Autocorrelation and Home Range Estimators. J. Wildl. Manag. 1997, 61, 280. [Google Scholar] [CrossRef]
- Hogg, R.V.; Craig, A.T. Introduction to Mathematical Statistics, 4th ed.; Macmillan: New York, NY, USA, 1978; ISBN 978-0-02-355710-1. [Google Scholar]
- Wolfe, J.D.; Ryder, T.B.; Pyle, P. Using Molt Cycles to Categorize the Age of Tropical Birds: An Integrative New System. J. Field Ornithol. 2010, 81, 186–194. [Google Scholar] [CrossRef]
- Studds, C.E.; Marra, P.P. Linking Fluctuations in Rainfall to Nonbreeding Season Performance in a Long-Distance Migratory Bird, Setophaga ruticilla. Clim. Res. 2007, 35, 115–122. [Google Scholar] [CrossRef]
- Johnson, M.D.; Sherry, T.W.; Holmes, R.T.; Marra, P.P. Assessing Habitat Quality for a Migratory Songbird Wintering in Natural and Agricultural Habitats. Conserv. Biol. 2006, 20, 1433–1444. [Google Scholar] [CrossRef]
- Bennett, R.; Leuenberger, W.; Leja, B.B.B.; Cáceres, A.S.; Johnson, K.; Larkin, J. Conservation of Neotropical Migratory Birds in Tropical Hardwood and Oil Palm Plantations. PLoS ONE 2018, 13, e0210293. [Google Scholar] [CrossRef]
- Holmes, R.T.; Sherry, T.W.; Reitsma, L. Population Structure, Territoriality and Overwinter Survival of Two Migrant Warbler Species in Jamaica. Condor 1989, 91, 545. [Google Scholar] [CrossRef]
- Cooper, N.W.; Sherry, T.W.; Marra, P.P. Modeling Three-Dimensional Space Use and Overlap in Birds. Auk 2014, 131, 681–693. [Google Scholar] [CrossRef]
- Smith, T.M.; Shugart, H.H. Territory Size Variation in the Ovenbird: The Role of Habitat Structure. Ecology 1987, 68, 695–704. [Google Scholar] [CrossRef]
- Stanley, C.Q.; Dudash, M.R.; Ryder, T.B.; Shriver, W.G.; Marra, P.P. Variable Tropical Moisture and Food Availability Underlie Mixed Winter Space-Use Strategies in a Migratory Songbird. Proc. R. Soc. B 2021, 288, 20211220. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, A.; Renton, K.; Rueda-Hernández, R. Winter Habitat Disturbance Influences Density and Territory Size of a Neotropical Migratory Warbler. J. Ornithol. 2017, 158, 63–73. [Google Scholar] [CrossRef]
- Marshall, M.R.; Cooper, R.J. Territory Size of a Migratory Songbird in Response to Caterpillar Density and Foliage Structure. Ecology 2004, 85, 432–445. [Google Scholar] [CrossRef]
- Brown, D.R.; Sherry, T.W. Alternative Strategies of Space Use and Response to Resource Change in a Wintering Migrant Songbird. Behav. Ecol. 2008, 19, 1314–1325. [Google Scholar] [CrossRef] [Green Version]
- Holmes, R.T.; Sherry, T.W. Site Fidelity of Migratory Warblers in Temperate Breeding and Neotropical Wintering Areas: Implications for Population Dynamics, Habitat Selection, and Conservation. In Ecology and Conservation of Neotropical Migrant Landbirds; Smithsonian Institute Press: Washington, DC, USA, 1992; pp. 563–575. [Google Scholar]
- Faaborg, J.R.; Arendt, W.J. Population Sizes and Philopatry of Winter Resident Warblers in Puerto Rico. J. Field Ornithol. 1984, 14, 151–166. [Google Scholar]
- Stamps, J.A.; Krishnan, V.V.; Reid, M.L. Search Costs and Habitat Selection by Dispersers. Ecology 2005, 86, 510–518. [Google Scholar] [CrossRef]
- Johnson, M.D.; Sherry, T.W. Effects of Food Availability on the Distribution of Migratory Warblers among Habitats in Jamaica. J. Anim. Ecol. 2001, 70, 546–560. [Google Scholar] [CrossRef] [Green Version]
- Latta, S.C.; Faaborg, J.R. Demographic and Population Responses of Cape May. America 2002, 83, 2502–2515. [Google Scholar]
- Wunderle, J.M.; Lebow, P.K.; White, J.D.; Currie, D. Sex and Age Differences in Site Fidelity, Food Resource Tracking, and Body Condition of Wintering Kirtland’s Warblers (Setophaga kirtlandii) in the Bahamas. Ornithol. Monogr. 2014, 80, 1–62. [Google Scholar] [CrossRef]
- Rosenberg, K.V.; Kennedy, J.A.; Dettmers, R.; Ford, R.P.; Reynolds, D.; Alexander, J.D.; Beardmore, C.J.; Blancher, P.J.; Bogart, R.E.; Butcher, G.S.; et al. Partners In Flight Landbird Conservation Plan: 2016 Revision for Canada and Continental United States. Partn. Flight Sci. Comm. 2016, 119, 96–103. [Google Scholar]
- Terborgh, J. Where Have All the Birds Gone? Essays on the Biology and Conservation of Birds That Migrate to the American Tropics; Princeton University Press: Princeton, NJ, USA, 1989. [Google Scholar]
- Marra, P.; Cohen, E.B.; Loss, S.R.; Rutter, J.E.; Tonra, C.M. A Call for Full Annual Cycle Research in Animal Ecology. Biol. Lett. 2015, 11, 20150552. [Google Scholar] [CrossRef]
- Rushing, C.S.; Marra, P.P.; Dudash, M.R. Winter Habitat Quality but Not Long-Distance Dispersal—Influences Apparent Reproductive Success in a Migratory Bird. Ecology 2016, 97, 1218–1227. [Google Scholar] [CrossRef]
Figure 1.
Boxplot of mean showing territory size of American Redstarts (Setophaga ruticilla) in oil palm plantations and in native forest in Tabasco, Mexico. Lines represent, in bottom-up order: minimum, first quartile, median, third quartile, and maximum.
Figure 1.
Boxplot of mean showing territory size of American Redstarts (Setophaga ruticilla) in oil palm plantations and in native forest in Tabasco, Mexico. Lines represent, in bottom-up order: minimum, first quartile, median, third quartile, and maximum.

Figure 2.
Pictures of the first year male American Redstart (SR6) captures in the first (a) and the second (b) wintering season, in oil palm plantations, Tabasco, Mexico.
Figure 2.
Pictures of the first year male American Redstart (SR6) captures in the first (a) and the second (b) wintering season, in oil palm plantations, Tabasco, Mexico.

Figure 3.
Oil palm plantation territory of the same American Redstart (SR6) on two consecutive wintering seasons, in the state of Tabasco, Mexico. The first season is represented by the transparent red polygon (0.34 ha) and the second season by transparent blue (0.29 ha).
Figure 3.
Oil palm plantation territory of the same American Redstart (SR6) on two consecutive wintering seasons, in the state of Tabasco, Mexico. The first season is represented by the transparent red polygon (0.34 ha) and the second season by transparent blue (0.29 ha).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Territory size estimates of territorial American Redstarts (Setophaga ruticilla) during the non-breeding season in Tabasco, Mexico. Two estimates for the individual SR6 are reported, each one for a different season.
Table 1.
Territory size estimates of territorial American Redstarts (Setophaga ruticilla) during the non-breeding season in Tabasco, Mexico. Two estimates for the individual SR6 are reported, each one for a different season.
| Bird | Territory Size (ha) | Vegetation | Sex | Age |
|---|---|---|---|---|
| SR1 | 0.421 | Oil palm | Male | Adult |
| SR2 | 0.181 | Oil palm | Male | Young |
| SR3 | 0.071 | Native forest | Male | Adult |
| SR4 | 0.248 | Native forest | Male | Adult |
| SR5 | 0.182 | Native forest | Male | Adult |
| SR6 | 0.344 | Oil palm | Male | Young |
| SR7 | 0.360 | Oil palm | Male | Young |
| SR8 | 0.241 | Oil palm | Male | Adult |
| SR6 | 0.292 | Oil palm | Male | Adult |
| SR14 | 0.182 | Oil palm | Male | Young |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Oliveira, S.L.; Flaspohler, D.J.; Wolfe, J.D. Winter Territoriality of the American Redstart in Oil Palm Plantations. Diversity 2022, 14, 1079. https://doi.org/10.3390/d14121079
AMA Style
Oliveira SL, Flaspohler DJ, Wolfe JD. Winter Territoriality of the American Redstart in Oil Palm Plantations. Diversity. 2022; 14(12):1079. https://doi.org/10.3390/d14121079
Chicago/Turabian StyleOliveira, Samuel L., David J. Flaspohler, and Jared D. Wolfe. 2022. "Winter Territoriality of the American Redstart in Oil Palm Plantations" Diversity 14, no. 12: 1079. https://doi.org/10.3390/d14121079
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.