Ethanolic Extract of Hippocampus abdominalis Exerts Anti-Melanogenic Effects in B16F10 Melanoma Cells and Zebrafish Larvae by Activating the ERK Signaling Pathway


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Preparation and Compositional Analysis of EEHA
2.3. Cell Culture
2.4. Cell Viability
2.5. Flow Cytometry Analysis
2.6. Measurement of Extracellular and Intracellular Melain Content
2.7. Measurement of Tyrosinase Activity
2.8. cAMP Assay
2.9. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.10. Protein Extraction and Western Blot Analysis
2.11. Anti-Melanogenic Effect in Zebrafish Larvae
2.12. Statistical Analysis
3. Results
3.1. Compositional Analysis of EEHA
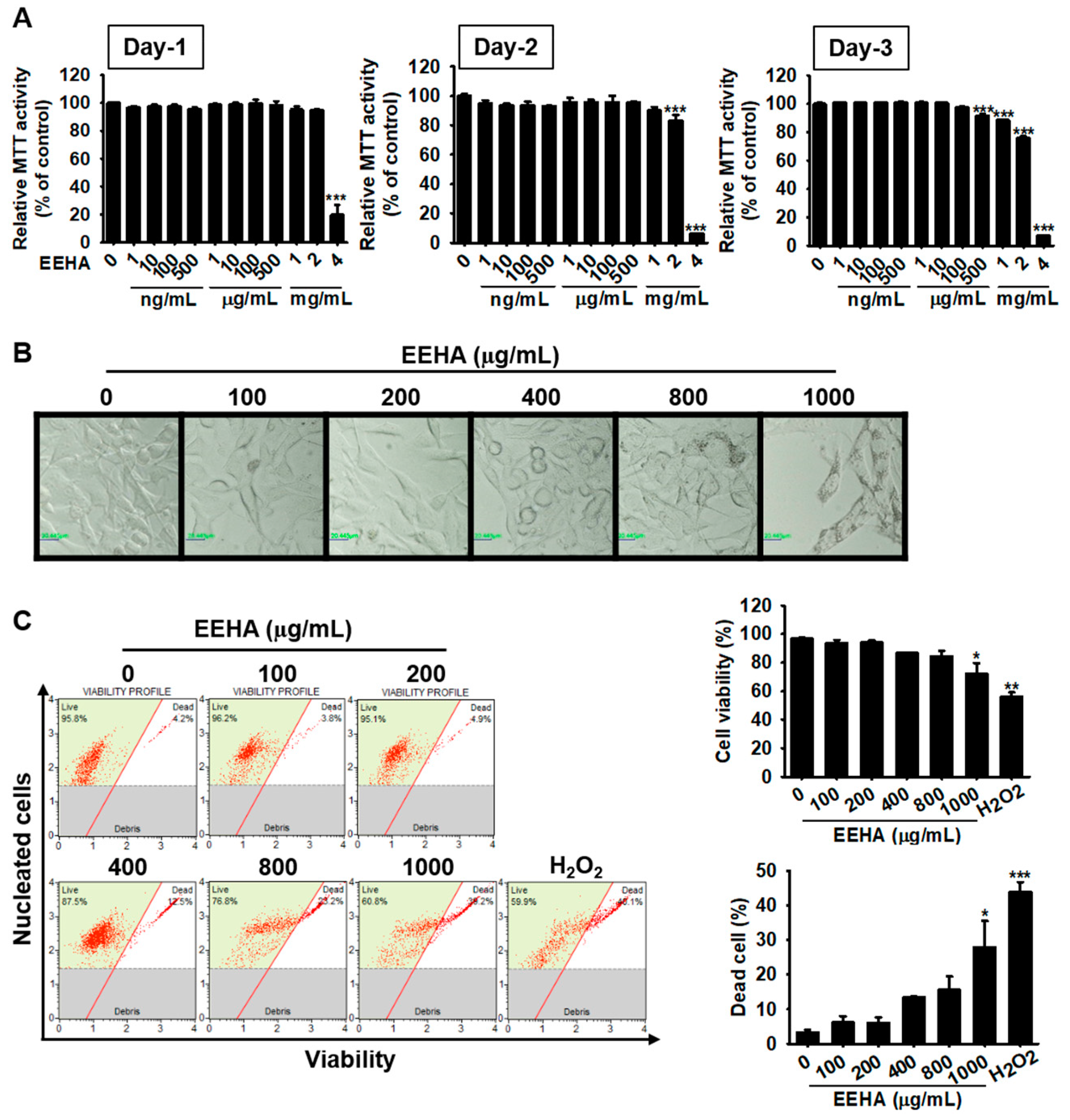
3.2. High Concentrations of EEHA Decrease B16F10 Melanoma Cell Viability
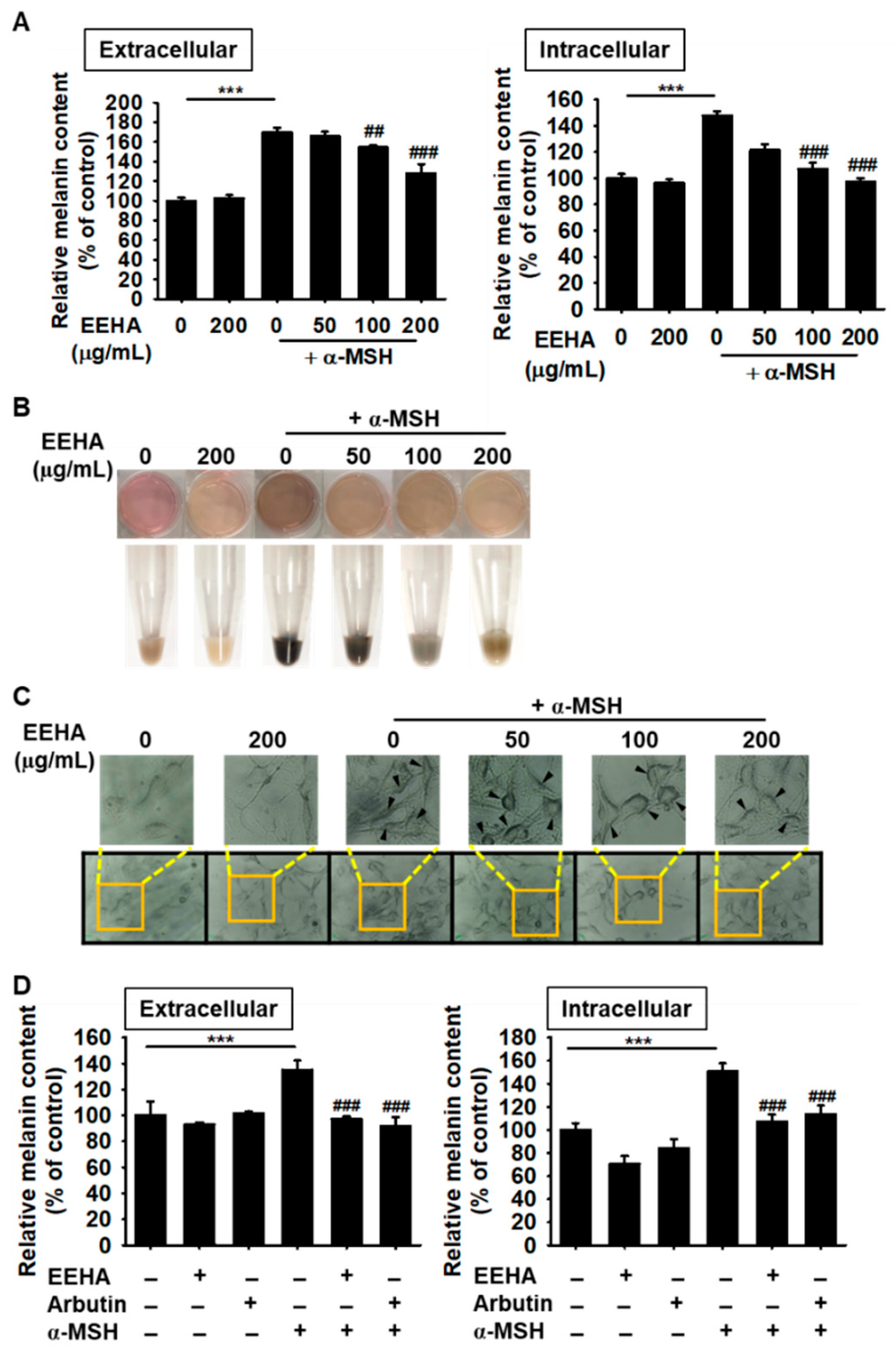
3.3. EEHA Inhibits α-MSH-Induced Melanogenesis in B16F10 Melanoma Cells
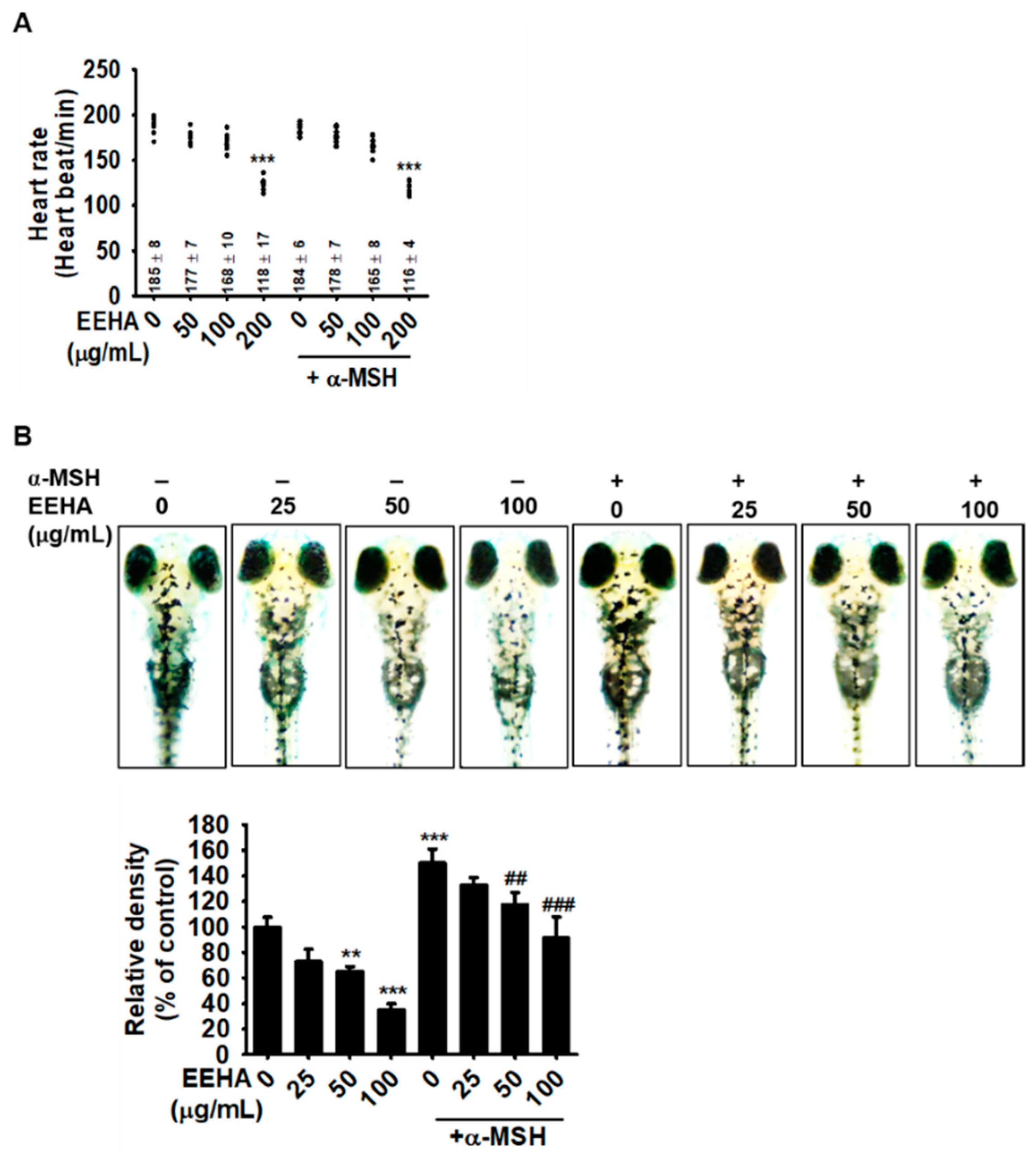
3.4. EEHA Attenuates Melanogenesis in Zebrafish Larvae
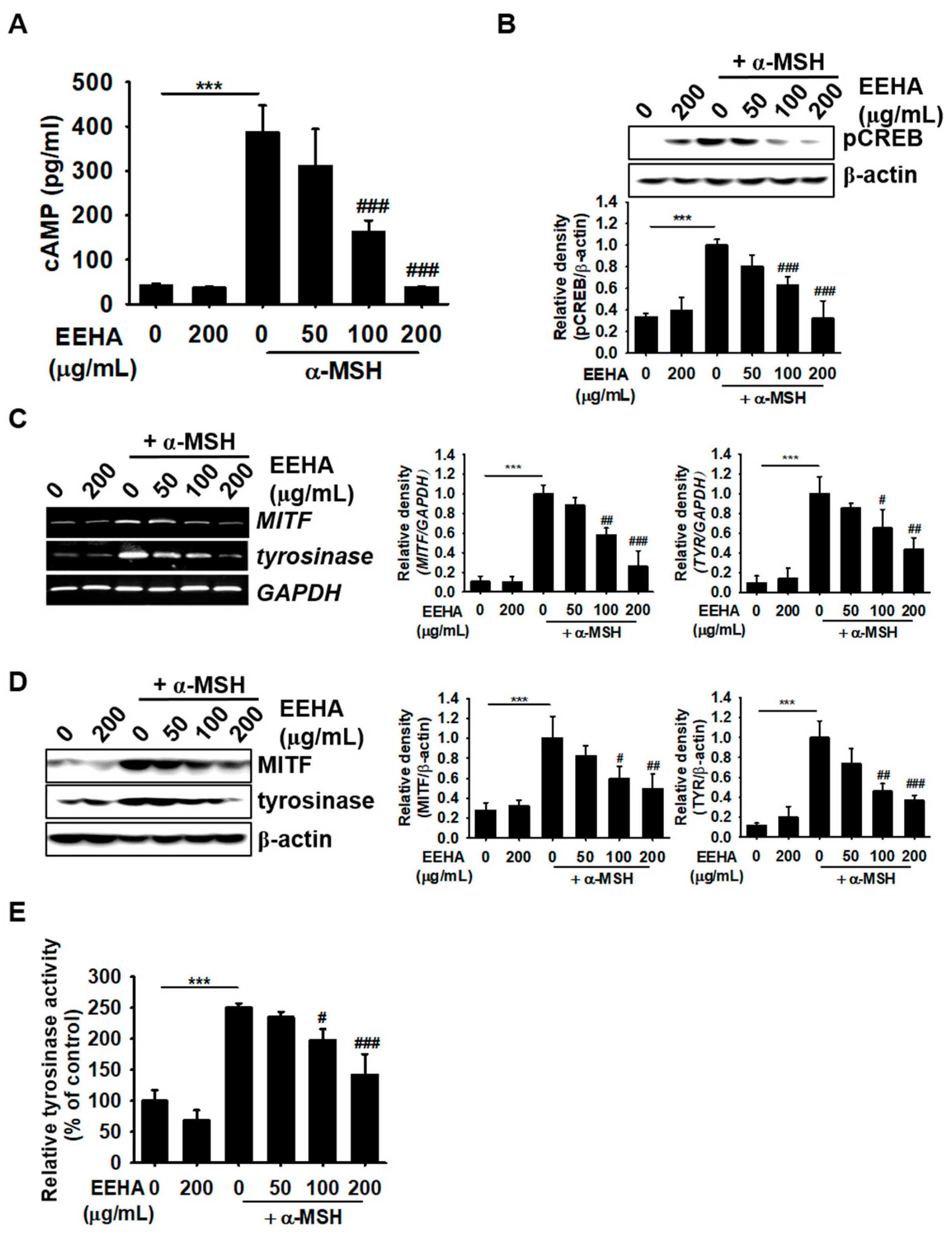
3.5. EEHA Inhibits MITF and Tyrosinase Expression in B16F10 Melanoma Cells by Suppressing the cAMP Signaling Pathway
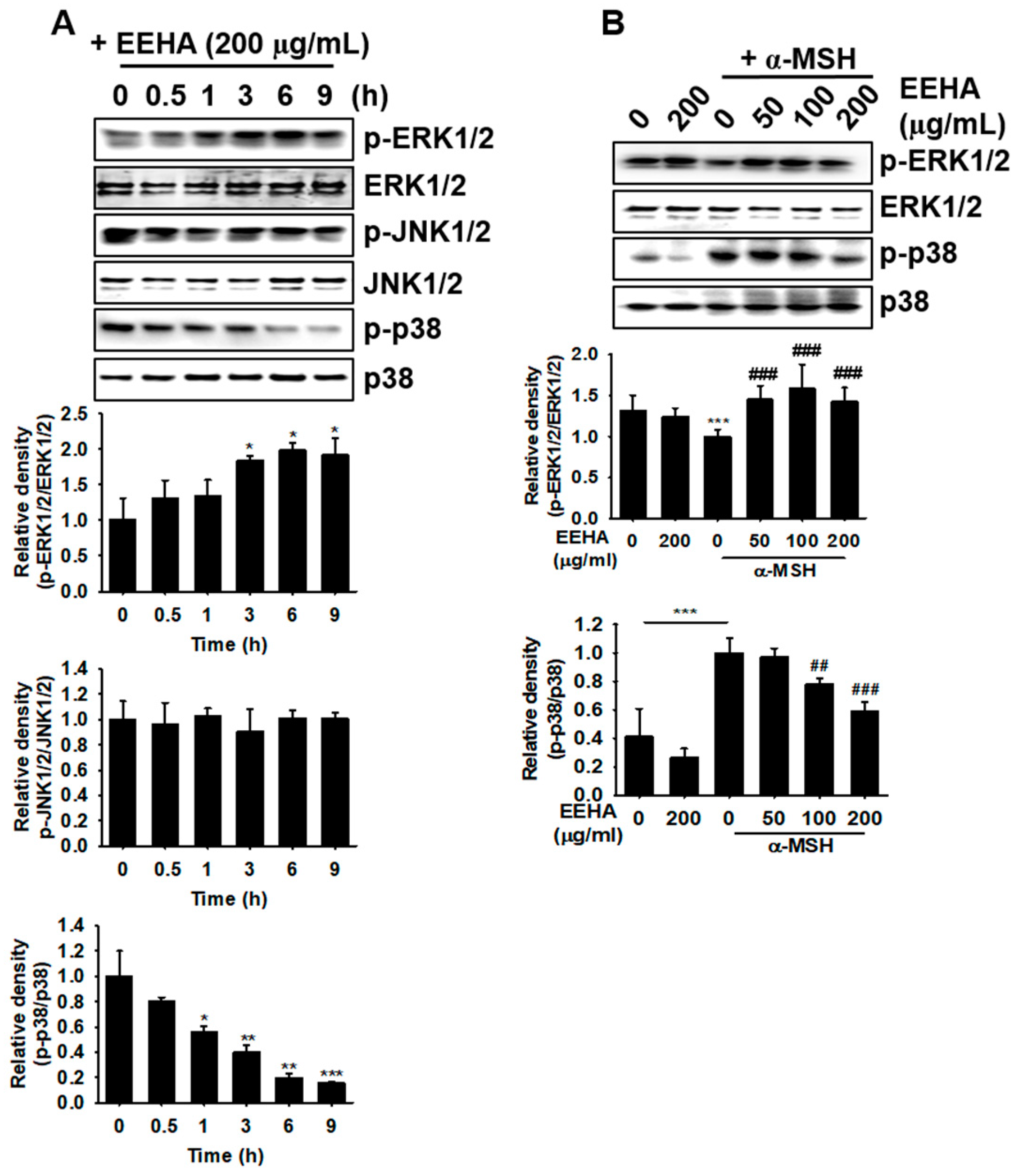
3.6. EEHA Increases ERK Phosphorylation and p38 MAPK Dephosphorylation
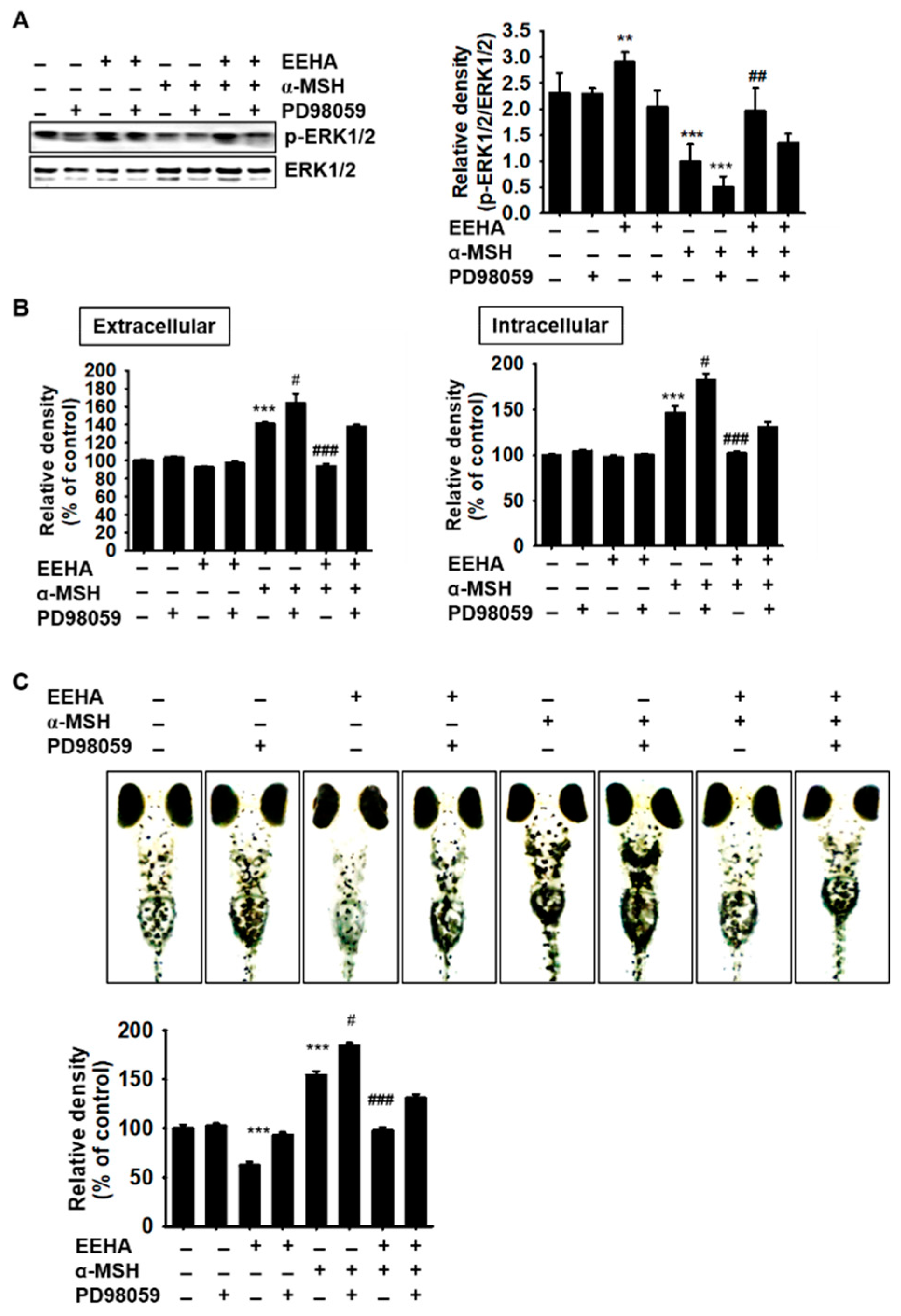
3.7. Inhibition of ERK Reverses EEHA-Mediated Anti-Melanogenesis in B16F10 Melanoma Cells and Zebrafish Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- D’Ischia, M.; Wakamatsu, K.; Cicoira, F.; Di Mauro, E.; Garcia-Borron, J.C.; Commo, S.; Galván, I.; Ghanem, G.; Kenzo, K.; Meredith, P.; et al. Melanins and melanogenesis: From pigment cells to human health and technological applications. Pigment Cell Melanoma Res. 2015, 28, 520–544. [Google Scholar] [CrossRef] [Green Version]
- Nicolaidou, E.; Katsambas, A.D. Pigmentation disorders: Hyperpigmentation and hypopigmentation. Clin. Dermatol. 2014, 32, 66–72. [Google Scholar] [CrossRef]
- Speeckaert, R.; Van Gele, M.; Speeckaert, M.M.; Lambert, J.; van Geel, N. The biology of hyperpigmentation syndromes. Pigment Cell Melanoma Res. 2014, 27, 512–524. [Google Scholar] [CrossRef]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaventure, J.; Domingues, M.J.; Larue, L. Cellular and molecular mechanisms controlling the migration of melanocytes and melanoma cells. Pigment Cell Melanoma Res. 2013, 26, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ferrer, A.; Rodriguez-Lopez, J.N.; Garcia-Canovas, F.; Garcia-Carmona, F. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta 1995, 1247, 1–11. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Recent development of signaling pathways inhibitors of melanogenesis. Cell. Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef]
- Chen, L.; Wang, X.; Huang, B. The genus Hippocampus—A review on traditional medicinal uses, chemical constituents and pharmacological properties. J. Ethnopharmacol. 2015, 162, 104–111. [Google Scholar] [CrossRef]
- Xu, D.H.; Wang, L.H.; Mei, X.T.; Li, B.J.; Lv, J.L.; Xu, S.B. Protective effects of seahorse extracts in a rat castration and testosterone-induced benign prostatic hyperplasia model and mouse oligospermatism model. Environ. Toxicol. Pharmacol. 2014, 37, 679–688. [Google Scholar] [CrossRef]
- Kumaravel, K.; Ravichandran, S.; Balasubramanian, T.; Siva Subramanian, K.; Bilal, A. Antimicrobial effect of five seahorse species from the Indian coast. Br. J. Pharmacol. Toxicol. 2010, 1, 62–66. [Google Scholar]
- Kumaravel, K.; Ravichandran, S.; Balasubramanian, T.; Sonneschein, L. Seahorses—A source of traditional medicine. Nat. Prod. Res. 2012, 26, 2330–2334. [Google Scholar] [CrossRef]
- Kang, N.; Kim, S.-Y.; Rho, S.; Ko, J.-Y.; Jeon, Y.-J. Anti-fatigue activity of a mixture of seahorse (Hippocampus abdominalis) hydrolysate and red ginseng. Fish. Aquat. Sci. 2017, 20, 3. [Google Scholar] [CrossRef] [Green Version]
- Woods, C.M.C. Improving initial survival in cultured seahorses, Hippocampus abdominalis Leeson, 1827 (Teleostei: Syngnathidae). Aquaculture 2000, 190, 377–388. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.; Lee, Y.C. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef]
- Tsuboi, T.; Kondoh, H.; Hiratsuka, J.; Mishima, Y. Enhanced melanogenesis induced by tyrosinase gene-transfer increases boron-uptake and killing effect of boron neutron capture therapy for amelanotic melanoma. Pigment Cell Res. 1998, 11, 275–282. [Google Scholar] [CrossRef]
- Lin, V.C.-H.; Ding, H.-Y.; Kuo, S.-Y.; Chin, L.-W.; Wu, J.-Y.; Chang, T.-S. Evaluation of in vitro and in vivo depigmenting activity of raspberry ketone from fheum officinale. Int. J. Mol. Sci. 2011, 12, 4819–4835. [Google Scholar] [CrossRef] [Green Version]
- Jani, R.A.; Nag, S.; Setty, S. Visualization of intracellular tyrosinase activity in vitro. Biol. Protocol. 2016, 6, 1794. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Ahn, C.B.; Yoon, N.Y.; Nam, K.H.; Kim, Y.K.; Je, J.Y. Protective effect of enzymatic hydrolysates from seahorse (Hippocampus abdominalis) against H2O2-mediated human umbilical vein endothelial cell injury. Biomed. Pharmacother. 2018, 108, 103–110. [Google Scholar] [CrossRef]
- Muthuramalingam, K.; Kim, J.H.; Jeon, Y.J.; Rho, S.; Kim, Y.M.; Cho, M. Effects of sea horse (Hippocampus abdominalis)-derived protein hydrolysate on skeletal muscle development. J. Appl. Biol. Chem. 2017, 60, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.J. 60 YEARS OF POMC: The proopiomelanocortin gene: Discovery, deletion and disease. J. Mol. Endocrinol. 2016, 56, T27–T37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, C.I.; Setaluri, V. Cyclic AMP (cAMP) signaling in melanocytes and melanoma. Arch. Biochem. Biophys. 2014, 563, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.; Lundegaard, P.R.; Reynolds, N.L.; Dorin, J.R.; Porteous, D.J.; Jackson, I.J.; Patton, E.E. mc1r Pathway regulation of zebrafish melanosome dispersion. Zebrafish 2008, 5, 289–295. [Google Scholar] [CrossRef]
- Chen, W.; Shi, X.; Ren, L.; Wang, K.; Liang, X.; Chen, L.; Sun, W.; Guo, W.; Yang, S. Transcript expression profiles of stria vascularis in Mitf-m knockout mice. Int. J. Clin. Exp. Med. 2017, 10, 2241–2251. [Google Scholar]
- Yi, X.; Zhao, G.; Zhang, H.; Guan, D.; Meng, R.; Zhang, Y.; Yang, Q.; Jia, H.; Dou, K.; Liu, C.; et al. MITF-siRNA Formulation Is a Safe and Effective Therapy for Human Melasma. Mol. Ther. 2011, 19, 362–371. [Google Scholar] [CrossRef]
- Kumari, S.; Thng, S.T.G.; Verma, N.K.; Gautam, H.K. Melanogenesis inhibitors. Acta Derm. Venereol. 2018, 98, 924–931. [Google Scholar] [CrossRef] [Green Version]
- Hachiya, A.; Kobayashi, A.; Ohuchi, A.; Takema, Y.; Imokawa, G. The paracrine role of stem cell factor/c-kit signaling in the activation of human melanocytes in ultraviolet-B-induced pigmentation. J. Investig. Dermatol. 2001, 116, 578–586. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Malek, Z.; Swope, V.B.; Suzuki, I.; Akcali, C.; Harriger, M.D.; Boyce, S.T.; Urabe, K.; Hearing, V.J. Mitogenic and melanogenic stimulation of normal human melanocytes by melanotropic peptides. Proc. Natl. Acad. Sci. USA 1995, 92, 1789–1793. [Google Scholar] [CrossRef] [Green Version]
- Nishio, T.; Usami, M.; Awaji, M.; Shinohara, S.; Sato, K. Dual effects of acetylsalicylic acid on ERK signaling and Mitf transcription lead to inhibition of melanogenesis. Mol. Cell. Biochem. 2016, 412, 101–110. [Google Scholar] [CrossRef]
- Saha, B.; Singh, S.K.; Sarkar, C.; Bera, R.; Ratha, J.; Tobin, D.J.; Bhadra, R. Activation of the Mitf promoter by lipid-stimulated activation of p38-stress signalling to CREB. Pigment Cell Res. 2006, 19, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Widlund, H.R.; Horstmann, M.A.; Price, E.R.; Cui, J.; Lessnick, S.L.; Wu, M.; He, X.; Fisher, D.E. Beta-catenin-induced melanoma growth requires the downstream target Microphthalmia-associated transcription factor. J. Cell Biol. 2002, 158, 1079–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EEHA | (%) |
|---|---|
| Protein | 81.8 ± 1.2 |
| Ash | 12.2 ± 0.2 |
| Carbohydrate | 4.6 ± 1.2 |
| Lipid | 3.5 ± 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molagoda, I.M.N.; Choi, Y.H.; Lee, S.; Sung, J.; Lee, C.R.; Lee, H.G.; Lim, J.; Lee, K.-J.; Jeon, Y.-J.; Ma, J.; et al. Ethanolic Extract of Hippocampus abdominalis Exerts Anti-Melanogenic Effects in B16F10 Melanoma Cells and Zebrafish Larvae by Activating the ERK Signaling Pathway. Cosmetics 2020, 7, 1. https://doi.org/10.3390/cosmetics7010001
Molagoda IMN, Choi YH, Lee S, Sung J, Lee CR, Lee HG, Lim J, Lee K-J, Jeon Y-J, Ma J, et al. Ethanolic Extract of Hippocampus abdominalis Exerts Anti-Melanogenic Effects in B16F10 Melanoma Cells and Zebrafish Larvae by Activating the ERK Signaling Pathway. Cosmetics. 2020; 7(1):1. https://doi.org/10.3390/cosmetics7010001
Chicago/Turabian StyleMolagoda, Ilandarage Menu Neelaka, Yung Hyun Choi, Seungheon Lee, Jiwon Sung, Cho Rong Lee, Hyo Geun Lee, Jongho Lim, Kyeong-Jun Lee, You-Jin Jeon, Jeongin Ma, and et al. 2020. "Ethanolic Extract of Hippocampus abdominalis Exerts Anti-Melanogenic Effects in B16F10 Melanoma Cells and Zebrafish Larvae by Activating the ERK Signaling Pathway" Cosmetics 7, no. 1: 1. https://doi.org/10.3390/cosmetics7010001