
Antigenotoxic Effect of Ascorbic Acid and Resveratrol in Erythrocytes of Ambystoma mexicanum, Oreochromis niloticus and Human Lymphocytes Exposed to Glyphosate

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
Chemicals and Reagents
2.2. Obtaining Individuals and Blood Cells
2.2.1. Oreochromis Niloticus
2.2.2. Ambystoma Mexicanum
2.2.3. Human Blood Cells
2.3. Preparation of Concentrations and Treatment
2.3.1. Evaluation of Genetic Damage in Blood Cells Exposed to EMS and Glyphosate
2.3.2. Evaluation of Antigenotoxic Activity
2.4. Comet Test
Comet Observation and Counting
2.5. Statistic Analysis
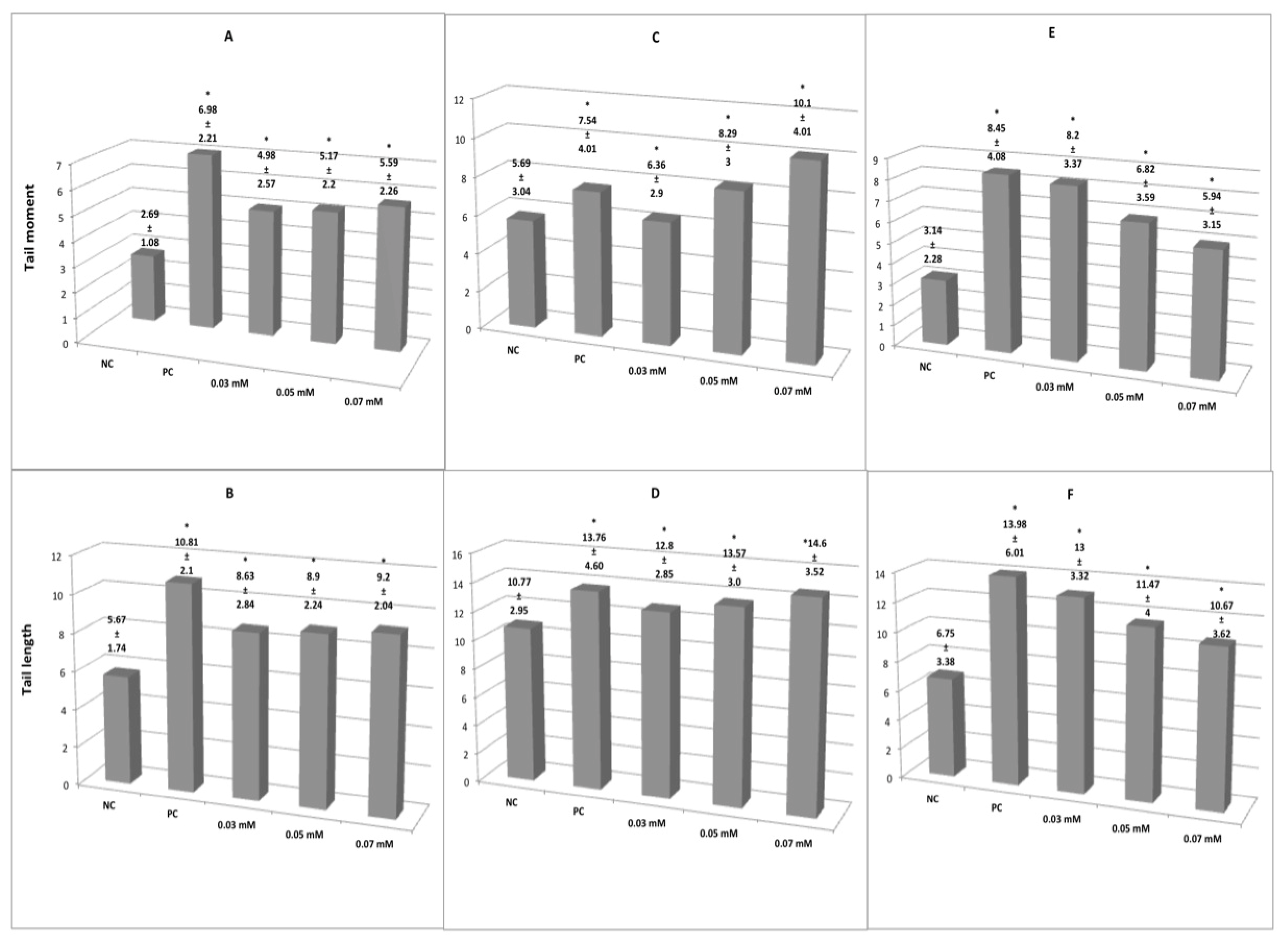
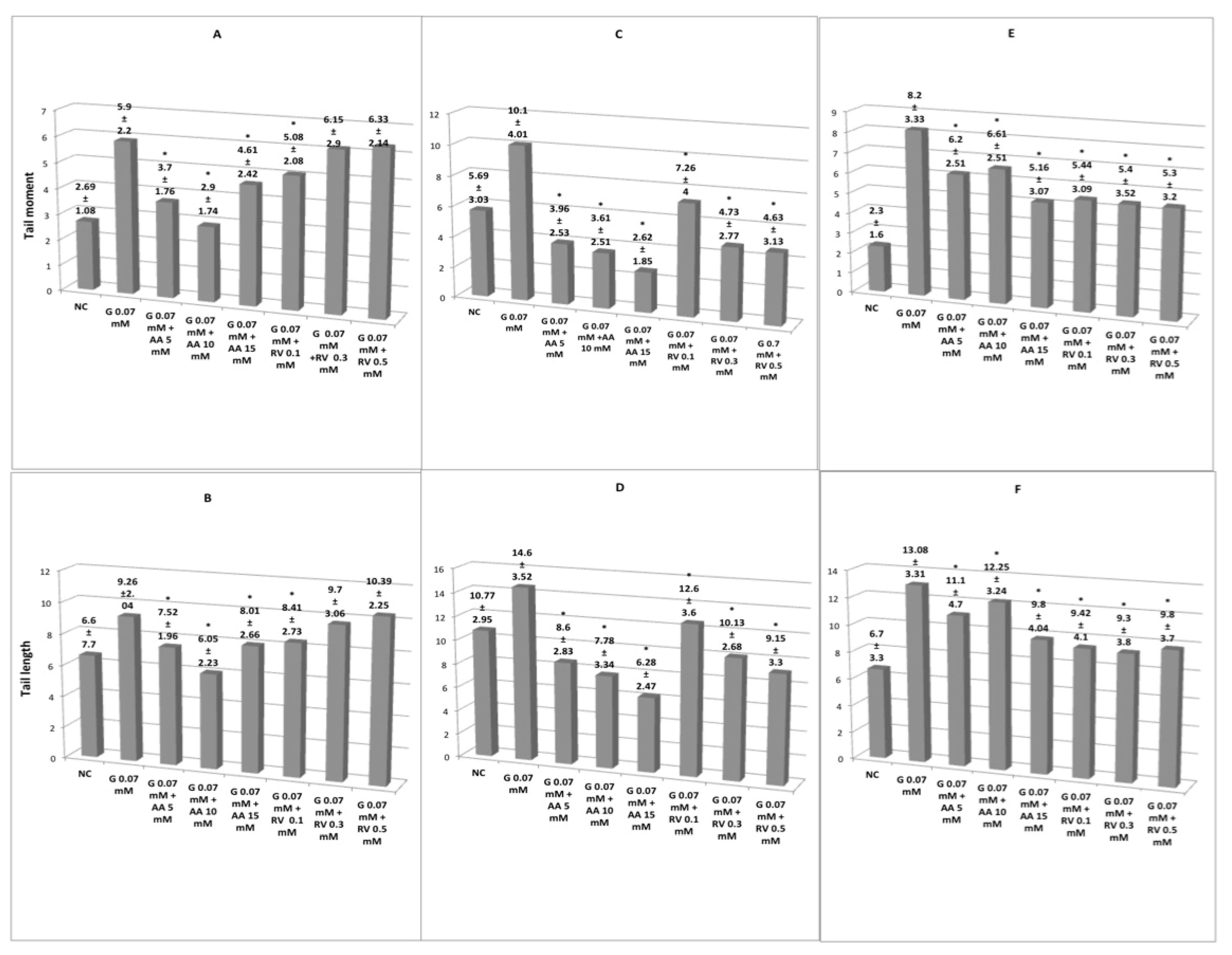
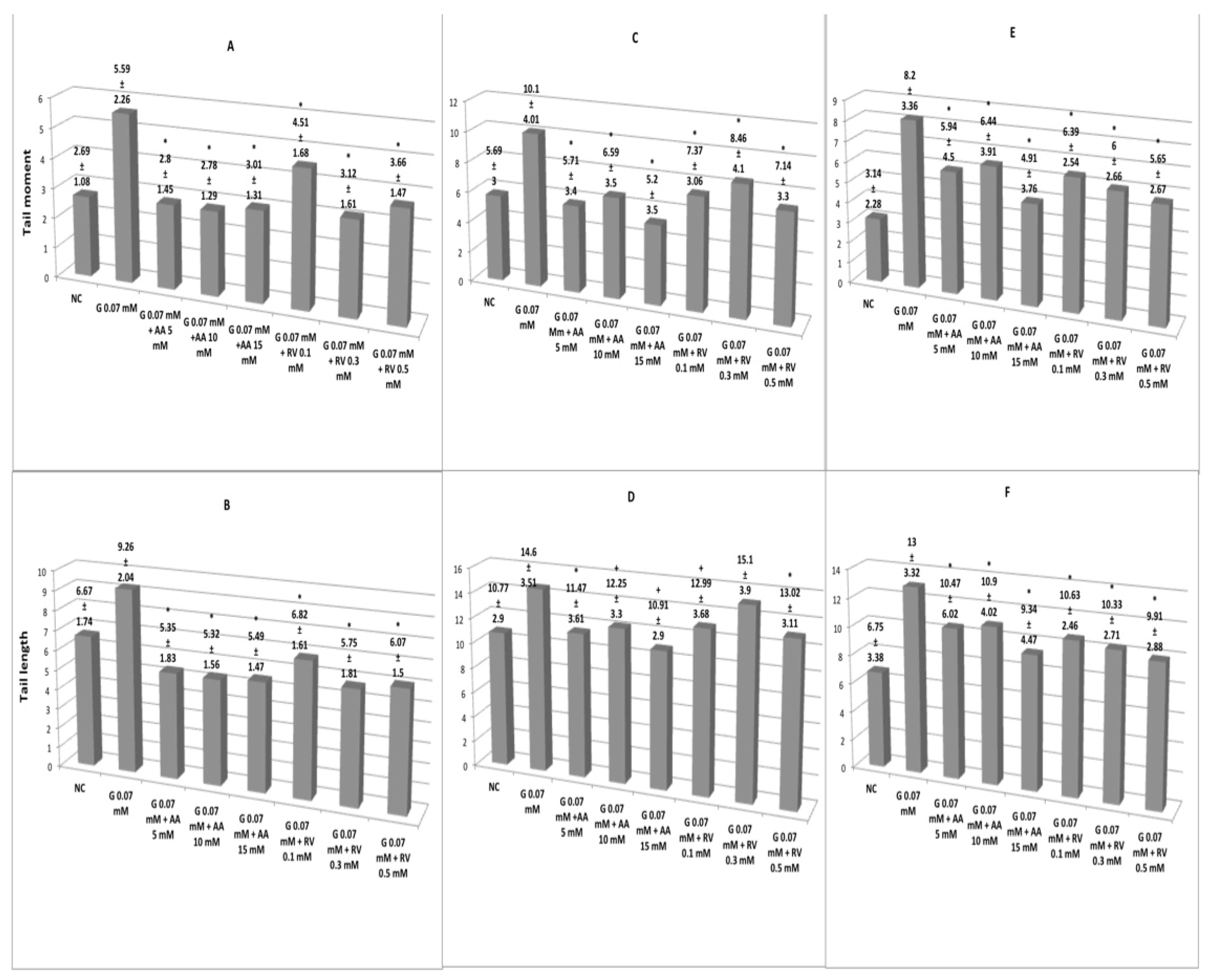
3. Results
3.1. Genotoxicity Induced by Glyphosate
3.2. Antigenotoxic Activity of Ascorbic Acid and Resveratrol in Erythrocytes and Lymphocytes Exposed to Glyphosate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatkowska, M.; Reszka, E.; Woźniak, K.; Jabłońska, E.; Michałowicz, J.; Bukowska, B. DNA damage and methylation induced by glyphosate in human peripheral blood mononuclear cells (in vitro study). Food Chem. Toxicol. 2017, 105, 93–98. [Google Scholar] [CrossRef] [PubMed]
- De Lapuente, J.; Lourenço, J.; Mendo, S.A.; Borràs, M.; Martins, M.G.; Costa, P.M.; Pacheco, M. The Comet Assay and its applications in the field of ecotoxicology: A mature tool that continues to expand its perspectives. Front. Genet. 2015, 6, 180. [Google Scholar] [CrossRef] [PubMed]
- Vischetti, C.; Casucci, C.; De Bernardi, A.; Monaci, E.; Tiano, L.; Marcheggiani, F.; Ciani, M.; Comitini, F.; Marini, E.; Taskin, E.; et al. Sub-Lethal Effects of Pesticides on the DNA of Soil Organisms as Early Ecotoxicological Biomarkers. Front. Microbiol. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Richmond, M.E. Glyphosate: A review of its global use, environmental impact, and potential health effects on humans and other species. J. Environ. Stud. Sci. 2018, 8, 416–434. [Google Scholar] [CrossRef]
- Gill, J.P.K.; Sethi, N.; Mohan, A.; Datta, S.; Girdhar, M. Glyphosate toxicity for animals. Environ. Chem. Lett. 2018, 16, 401–426. [Google Scholar] [CrossRef]
- Tsai, W.T. Trends in the use of glyphosate herbicide and its relevant regulations in Taiwan: A water contaminant of increasing concern. Toxics 2019, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Matozzo, V.; Fabrello, J.; Marin, M.G. The effects of glyphosate and its commercial formulations to marine invertebrates: A review. J. Mar. Sci. Eng. 2020, 8, 399. [Google Scholar] [CrossRef]
- Hong, Y.; Yang, X.; Huang, Y.; Yan, G.; Cheng, Y. Assessment of the oxidative and genotoxic effects of the glyphosate-based herbicide roundup on the freshwater shrimp, Macrobrachium nipponensis. Chemosphere 2018, 210, 896–906. [Google Scholar] [CrossRef]
- Alvarez-Moya, C.; Silva, M.R.; Valdez-Ramírez, C.; Gallardo, D.G.; León-Sánchez, R.; Aguirre, A.C.; Velasco, A.F. Comparison of the in vivo and in vitro genotoxicity of glyphosate isopropylamine salt in three different organisms. Genet. Mol. Biol. 2014, 37, 105–110. [Google Scholar] [CrossRef]
- Santos, W.S.; Gonzalez, P.; Cormier, B.; Mazzella, N.; Bonnaud, B.; Morin, S.; Clérandeau, C.; Morin, B.; Cachot, J. A glyphosate-based herbicide induces sub-lethal effects in early life stages and liver cell line of rainbow trout, Oncorhynchus mykiss. Aquat. Toxicol. 2019, 216, 105291. [Google Scholar] [CrossRef] [PubMed]
- Meza-Joya, F.L.; Ramírez-Pinilla, M.P.; Fuentes-Lorenzo, J.L. Toxic, cytotoxic, and genotoxic effects of a glyphosate formulation (Roundup®SL-Cosmoflux®411F) in the direct-developing frog Eleutherodactylus johnstonei. Environ. Mol. Mutagen. 2013, 54, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Soloneski, S.; Ruiz de Arcaute, C.; Larramendy, M.L. Genotoxic effect of a binary mixture of dicamba- and glyphosate-based commercial herbicide formulations on Rhinella arenarum (Hensel, 1867) (Anura, Bufonidae) late-stage larvae. Environ. Sci. Pollut. Res. 2016, 23, 17811–17821. [Google Scholar] [CrossRef]
- Carvalho, W.F.; Franco, F.C.; Godoy, F.R.; Folador, D.; Avelar, J.B.; Nomura, F.; Cruz, A.D.; Sabóia-Morais, S.M.T.; Bastos, R.P.; Silva, D. Evaluation of Genotoxic and Mutagenic Effects of Glyphosate Roundup Original ® in Dendropsophus minutus Peters, 1872 Tadpoles. S. Am. J. Herpetol. 2018, 13, 220–229. [Google Scholar] [CrossRef]
- Poletta, G.L.; Larriera, A.; Kleinsorge, E.; Mudry, M.D. Genotoxicity of the herbicide formulation Roundup® (glyphosate) in broad-snouted caiman (Caiman latirostris) evidenced by the Comet assay and the Micronucleus test. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2009, 672, 95–102. [Google Scholar] [CrossRef] [PubMed]
- González, H.M.; Zamora, E. Manual Básico para el Cuidado en Cautiverio del Axolote de Xochimilco (Ambystoma mexicanum), 1st ed.; Instituto de Biología: Mexico City, Mexico, 2014; pp. 18–24. [Google Scholar]
- Schaumburg, L.G.; Siroski, P.A.; Poletta, G.L.; Mudry, M.D. Genotoxicity induced by Roundup® (Glyphosate) in tegu lizard (Salvator merianae) embryos. Pestic. Biochem. Physiol. 2016, 130, 71–78. [Google Scholar] [CrossRef]
- Hussain, R.; Ali, F.; Rafique, A.; Ghaffar, A.; Jabeen, G.; Rafay, M.; Liaqat, S.; Khan, I.; Malik, R.; Kasib Khan, M.; et al. Exposure to Sub-Acute Concentrations of Glyphosate Induce Clinico-Hematological, Serum Biochemical and Genotoxic Damage in Adult Cockerels. Pak. Vet. J. 2019, 39, 181–186. [Google Scholar] [CrossRef]
- Rossi, L.F.; Luaces, J.P.; Palermo, A.M.; Merani, M.S.; Mudry, M.D. Cytogenetic damage in peripheral blood cultures of Chaetophractus villosus exposed in vivo to a glyphosate formulation (Roundup). Ecotoxicol. Environ. Saf. 2018, 157, 121–127. [Google Scholar] [CrossRef]
- Milić, M.; Žunec, S.; Micek, V.; Kašuba, V.; Mikolić, A.; Lovaković, B.T.; Semren, T.Ž.; Pavičić, I.; Čermak, A.M.M.; Pizent, A.; et al. Oxidative stress, cholinesterase activity, and DNA damage in the liver, whole blood, and plasma of Wistar rats following a 28-day exposure to glyphosate. Arh. Hig. Rada Toksikol. 2018, 69, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Townsend, M.; Peck, C.; Meng, W.; Heaton, M.; Robison, R.; O’Neill, K. Evaluation of various glyphosate concentrations on DNA damage in human Raji cells and its impact on cytotoxicity. Regul. Toxicol. Pharmacol. 2017, 85, 79–85. [Google Scholar] [CrossRef]
- Mesnage, R.; Defarge, N.; Spiroux de Vendômois, J.; Séralini, G.E. Potential toxic effects of glyphosate and its commercial formulations below regulatory limits. Food Chem. Toxicol. 2015, 84, 133–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.A.; Maturano, E.; Etchegoyen, A.; Difilippo, F.S.; Maclean, B. Association between Cancer and Environmental Exposure to Glyphosate. Int. J. Clin. Med. 2017, 8, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Andreotti, G.; Koutros, S.; Hofmann, J.N.; Sandler, D.P.; Lubin, J.H.; Lynch, C.F.; Lerro, C.C.; De Roos, A.J.; Parks, C.G.; Alavanja, M.C.; et al. Glyphosate Use and Cancer Incidence in the Agricultural Health Study. J. Natl. Cancer Inst. 2018, 110, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Milesi, M.M.; Lorenz, V.; Pacini, G.; Repetti, M.R.; Demonte, L.D.; Varayoud, J.; Luque, E.H. Perinatal exposure to a glyphosate-based herbicide impairs female reproductive outcomes and induces second-generation adverse effects in Wistar rats. Arch. Toxicol. 2018, 92, 2629–2643. [Google Scholar] [CrossRef]
- Avila-Vazquez, M.; Difilippo, F.S.; Lean, B.; Maturano, E.; Etchegoyen, A. Environmental Exposure to Glyphosate and Reproductive Health Impacts in Agricultural Population of Argentina. J. Environ. Prot. 2018, 9, 241–253. [Google Scholar] [CrossRef] [Green Version]
- George, J.; Prasad, S.; Mahmood, Z.; Shukla, Y. Studies on glyphosate-induced carcinogenicity in mouse skin: A proteomic approach. J. Proteom. 2010, 73, 951–964. [Google Scholar] [CrossRef]
- Bai, S.H.; Ogbourne, S.M. Glyphosate: Environmental contamination, toxicity and potential risks to human health via food contamination. Environ. Sci. Pollut. Res. 2016, 23, 18988–19001. [Google Scholar] [CrossRef]
- Gajski, G.; Žegura, B.; Ladeira, C.; Pourrut, B.; Del Bo, C.; Novak, M.; Sramkova, M.; Milić, M.; Gutzkow, K.B.; Costa, S.; et al. The comet assay in animal models: From bugs to whales—(Part 1 Invertebrates). Mutat. Res. Rev. Mutat. Res. 2019, 779, 82–113. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga-Gonzales, G. Sistemas de la detección de daño genético. In Genética, Ambiente y Salud, 3rd ed.; Álvarez-Moya, C., Ed.; Editorial universitário; Universidad de Guadalajara: Guadalajara, Mexico, 2013; Volume 1, pp. 99–119. [Google Scholar]
- Uno, Y.; Kojima, H.; Omori, T.; Corvi, R.; Honma, M.; Schechtman, L.M.; Tice, R.R.; Beevers, C.; De Boeck, M.; Burlinson, B.; et al. JaCVAM-organized international validation study of the in vivo rodent alkaline comet assay for detection of genotoxic carcinogens: II. Summary of definitive validation study results. Mutat. Res.Genet. Toxicol. Environ. Mutagen. 2015, 786, 45–76. [Google Scholar] [CrossRef]
- Santo, G.D.; Grotto, A.; Boligon, A.A.; Da Costa, B.; Rambo, C.L.; Fantini, E.A.; Sauer, E.; Lazzarotto, L.M.V.; Bertoncello, K.T.; Júnior, O.T.; et al. Protective effect of Uncaria tomentosa extract against oxidative stress and genotoxicity induced by glyphosate-Roundup® using zebrafish (Daniorerio) as a model. Environ. Sci. Pollut. Res. 2018, 25, 11703–11715. [Google Scholar] [CrossRef] [PubMed]
- Çavuşoǧlu, K.; Yapar, K.; Oruç, E.; Yalçin, E. Protective effect of ginkgo biloba L. leaf extract against glyphosate toxicity in swiss albino mice. J. Med. Food. 2011, 14, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Romero-Jiménez, M.; Campos-Sánchez, J.; Analla, M.; Muñoz-Serrano, A.; Alonso-Moraga, Á. Genotoxicity and anti-genotoxicity of some traditional medicinal herbs. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2005, 585, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Branco, C.S.; Garcez, M.E.; Pasqualotto, F.F.; Erdtman, B.; Salvador, M. Resveratrol and ascorbic acid prevent DNA damage induced by cryopreservation in human semen. Cryobiology 2010, 60, 235–237. [Google Scholar] [CrossRef]
- Nguyen, T.; Talbi, H.; Hilali, A.; Anthonissen, R.; Maes, A.; Verschaeve, L. In vitro toxicity, genotoxicity and antigenotoxicity of Nigella sativa extracts from different geographic locations. S. Afr. J. Bot. 2019, 126, 132–141. [Google Scholar] [CrossRef]
- Siddique, Y.H.; Beg, T.; Afzal, M. Antigenotoxic effects of ascorbic acid against megestrol acetate-induced genotoxicity in mice. Hum. Exp. Toxicol. 2005, 24, 121–127. [Google Scholar] [CrossRef]
- Ranjini, A.; Manonmani, H.K. Resveratrol protects acrylamide / glycidamide induced genotoxicity in hepatic cell line. Pharma. Innov. J. 2019, 8, 56–64. Available online: https://www.thepharmajournal.com/archives/2019/vol8issue1/PartB/7-12-46-567.pdf (accessed on 1 March 2021).
- Ikpeme, E. Efficacy of Ascorbic Acid in Reducing Glyphosate-Induced Toxicity in Rats. Br. Biotechnol. J. 2012, 2, 157–168. [Google Scholar] [CrossRef]
- NOM-062-ZOO-1999 Norma Oficial Mexicana, 1999. Especificaciones Técnicas para la Producción, Cuidado y Uso de los Animales de Laboratorio. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación: Mexico City, Mexico. Available online: https://www.fmvz.unam.mx/fmvz/principal/archivos/062ZOO.PDF (accessed on 11 February 2021).
- Close, B.; Banister, K.; Baumans, V.; Bernoth, E.M.; Bromage, N.; Bunyan, J.; Erhardt, W.; Flecknell, P.; Gregory, N.; Hackbarth, H.; et al. Recomendaciones para la Eutanasia de los Animales de Experimentación: Parte 2. 1996. Available online: https://sea.umh.es/files/2011/07/eutanasia2.pdf (accessed on 19 March 2021).
- Barriga-Vallejo, C.; Aguilera, C.; Cruz, J.; Banda-Leal, J.; Lazcano, D.; Mendoza, R. Ecotoxicological Biomarkers in Multiple Tissues of the Neotenic Ambystoma spp. for a Non-lethal Monitoring of Contaminant Exposure in Wildlife and Captive Populations. Water. Air. Soil Pollut. 2016, 228, 1–11. [Google Scholar] [CrossRef]
- Oliveira, R.J.; Ribeiro, L.R.; da Silva, A.F.; Matuo, R.; Mantovani, M.S. Evaluation of antimutagenic activity and mechanisms of action of β-glucan from barley, in CHO-k1 and HTC cell lines using the micronucleus test. Toxicol. Vitr. 2006, 20, 1225–1233. [Google Scholar] [CrossRef]
- Herek, J.S.; Vargas, L.; Rinas Trindade, S.A.; Rutkoski, C.F.; Macagnan, N.; Hartmann, P.A.; Hartmann, M.T. Genotoxic effects of glyphosate on Physalaemus tadpoles. Environ. Toxicol. Pharmacol. 2021, 81, 103516. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, E.; Sicińska, P.; Michałowicz, J.; Woźniak, K.; Reszka, E.; Huras, B.; Zakrzewski, J.; Bukowska, B. The mechanism of DNA damage induced by Roundup 360 PLUS, glyphosate and AMPA in human peripheral blood mononuclear cells—Genotoxic risk assessement. Food Chem. Toxicol. 2018, 120, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Farthing, T.S.; Muir, J.P.; Brady, J.A. Three Bermudagrass-suppression techniques have little effect on soil-nutrient availability and microbial communities 200 days after application. Appl. Soil Ecol. 2020, 145, 103352. [Google Scholar] [CrossRef]
- Solomon, K.R.; Thompson, D.G. Ecological risk assessment for aquatic organisms from over-water uses of glyphosate. J. Toxicol. Environ. Health Part B Crit. Rev. 2003, 6, 289–324. [Google Scholar] [CrossRef] [PubMed]
- Nagy, K.; Tessema, R.A.; Budnik, L.T.; Ádám, B. Comparative cyto- and genotoxicity assessment of glyphosate and glyphosate-based herbicides in human peripheral white blood cells. Environ. Res. 2019, 179, 108851. [Google Scholar] [CrossRef]
- Martini, C.N.; Gabrielli, M.; María Codesido, M.; Del Vila, M.C. Glyphosate-based herbicides with different adjuvants are more potent inhibitors of 3T3-L1 fibroblast proliferation and differentiation to adipocytes than glyphosate alone. Comp. Clin. Path. 2016, 25, 607–613. [Google Scholar] [CrossRef]
- Robles-Mendoza, C.; García-Basilio, C.; Cram-Heydrich, S.; Hernández-Quiroz, M.; Vanegas-Pérez, C. Organophosphorus pesticides effect on early stages of the axolotl Ambystoma mexicanum (Amphibia: Caudata). Chemosphere 2009, 74, 703–710. [Google Scholar] [CrossRef]
- Alvarez-Moya, C.; Santerre-Lucas, A.; Zúñiga-González, G.; Torres-Bugarín, O.; Padilla-Camberos, E.; Feria-Velasco, A. Evaluation of genotoxic activity of maleic hydrazide, ethyl methane sulfonate, and N-nitroso diethylamine in Tradescantia. Sal. Pub. Mex. 2001, 43, 6–12. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0036-36342001000600007 (accessed on 17 May 2021). [CrossRef]
- Kumar, D.; Salian, S.R.; Kalthur, G.; Uppangala, S.; Kumari, S.; Challapalli, S.; Chandraguthi, S.G.; Jain, N.; Krishnamurthy, H.; Kumar, P. Association between sperm DNA integrity and seminal plasma antioxidat levels in health workers occupationally exposed to ionizing radiation. Environ. Res. 2014, 132, 297–304. [Google Scholar] [CrossRef]
- Johnson, A.A.; Naaldijk, Y.; Hohaus, C.; Meisel, H.J.; Krystel, L.; Stolzing, A. Protective effects of alpha phenyl-tert-butyl nitrone and ascorbic acid in human adipose derived mesenchymal stem cells from differently aged donors. Aging 2017, 9, 340–352. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, L.C.L.; Soares, R.E.P.; Moreira, V.R.; Pontes, R.L.; Castelo-Branco, P.V.; Ferreira Pereira, S.R. Genistein and ascorbic acid reduce oxidative stress-derived DNA damage induced by the antileishmanial meglumine antimoniate. Antimicrob. Agents Chemother. 2018, 62, e00456-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, S.; Lee, J.; Jo, H.; Scholten, J.; Willingham, R.; Nicoll, J.; Baswan, S.M. Chrysanthemum morifolium extract and ascorbic acid-2-glucoside (Aa2g) blend inhibits uva-induced delayed cyclobutane pyrimidine dimer (cpd) production in melanocytes. Clin. Cosmet. Investig. Dermatol. 2019, 12, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Kašuba, V.; Milić, M.; Rozgaj, R.; Kopjar, N.; Mladinić, M.; Žunec, S.; Želježić, D. Effects of low doses of glyphosate on DNA damage, cell proliferation and oxidative stress in the HepG2 cell line. Environ. Sci. Poll. Res. 2017, 24, 19267–19281. [Google Scholar] [CrossRef]
- Jiraungkoorskul, W.; Sahaphong, S.; Kangwanrangsan, N.; Zakaria, S. The protective influence of ascorbic acid against the genotoxicity of waterborne lead exposure in Nile tilapia Oreochromis niloticus (L.). J. Fish. Bio. 2008, 73, 355–366. [Google Scholar] [CrossRef]
- Dos Santos, P.C.; Dutra, J.C.V.; Delarmelina, J.M.E.; Tose, L.V.; Romão, W.M.; Jamal, C.S.; França, H.S.; Do Carmo Pimentel Batitucci, M. Coriandrum sativum grown under organic or chemical fertilizer effectively prevents DNA damage: Preliminary phytochemical screening, flavonoid content, ESI (-) FT-ICR MS, in vitro antioxidant and in vivo (mice bone marrow) antimutagenic activity against cyclophosphamide. Asian Pac. J. Trop. Biomed. 2018, 8, 292–301. [Google Scholar] [CrossRef]
- Almeida, I.V.; Düsman, E.; Mattge, G.I.; Toledo, F.; Reusing, A.F.; Vicentini, V.E.P. In vivo antimutagenic activity of the medicinal plants Pfaffia glomerata (Brazilian ginseng) and Ginkgo biloba. Genet. Mol. Res. 2017, 16, 16039785. [Google Scholar] [CrossRef]
- De La Lastra, A.C.; Villegas, I. Resveratrol as an antioxidant and pro-oxidant agent: Mechanisms and clinical implications, in: Biochemical Society Transactions. Biochem Soc. Trans. 2007, 35, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Sotler, R.; Poljšak, B.; Dahmane, R.; Jukić, T.; Pavan Jukić, D.; Rotim, C.; Trebše, P.; Starc, A. Pro-oxidant activities of antioxidants and their impact on health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef]
- Jia, R.; Li, Y.; Cao, L.; Du, J.; Zheng, T.; Qian, H.; Gu, Z.; Jeney, G.; Xu, P.; Yin, G. Antioxidative, anti-inflammatory and hepatoprotective effects of resveratrol on oxidative stress-induced liver damage in tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 215, 56–66. [Google Scholar] [CrossRef]
- Zheng, L.F.; Wei, Q.Y.; Cai, Y.J.; Fang, J.G.; Zhou, B.; Yang, L.; Liu, Z.L. DNA damage induced by resveratrol and its synthetic analogues in the presence of Cu (II) ions: Mechanism and structure-activity relationship. Free Radic. Biol. Med. 2006, 41, 1807–1816. [Google Scholar] [CrossRef]
- Medina, I.; Alcántara, D.; González, M.J.; Torres, P.; Lucas, R.; Roque, J.; Plou, F.J.; Morales, J.C. Antioxidant activity of resveratrol in several fish lipid matrices: Effect of acylation and glucosylation. J. Agric. Food Chem. 2010, 58, 9778–9786. [Google Scholar] [CrossRef]
- Turkmen, R.; Birdane, Y.O.; Demirel, H.H.; Kabu, M.; Ince, S. Protective effects of resveratrol on biomarkers of oxidative stress, biochemical and histopathological changes induced by sub-chronic oral glyphosate-based herbicide in rats. Toxicol. Res. 2019, 8, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Giordo, R.; Nasrallah, G.K.; Al-Jamal, O.; Paliogiannis, P.; Pintus, G. Resveratrol inhibits oxidative stress and prevents mitochondrial damage induced by zinc oxide nanoparticles in zebrafish (Daniorerio). Int. J. Mol. Sci. 2020, 21, 3838. [Google Scholar] [CrossRef]
- Chen, C.; Jiang, X.; Hu, Y.; Zhang, Z. The protective role of resveratrol in the sodium arsenite-induced oxidative damage via modulation of intracellular gsh homeostasis. Biol. Trace Elem. Res. 2013, 155, 119–131. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Moya, C.; Sámano-León, A.G.; Reynoso-Silva, M.; Ramírez-Velasco, R.; Ruiz-López, M.A.; Villalobos-Arámbula, A.R. Antigenotoxic Effect of Ascorbic Acid and Resveratrol in Erythrocytes of Ambystoma mexicanum, Oreochromis niloticus and Human Lymphocytes Exposed to Glyphosate. Curr. Issues Mol. Biol. 2022, 44, 2230-2242. https://doi.org/10.3390/cimb44050151
Alvarez-Moya C, Sámano-León AG, Reynoso-Silva M, Ramírez-Velasco R, Ruiz-López MA, Villalobos-Arámbula AR. Antigenotoxic Effect of Ascorbic Acid and Resveratrol in Erythrocytes of Ambystoma mexicanum, Oreochromis niloticus and Human Lymphocytes Exposed to Glyphosate. Current Issues in Molecular Biology. 2022; 44(5):2230-2242. https://doi.org/10.3390/cimb44050151
Chicago/Turabian StyleAlvarez-Moya, Carlos, Alexis Gerardo Sámano-León, Mónica Reynoso-Silva, Rafael Ramírez-Velasco, Mario Alberto Ruiz-López, and Alma Rosa Villalobos-Arámbula. 2022. "Antigenotoxic Effect of Ascorbic Acid and Resveratrol in Erythrocytes of Ambystoma mexicanum, Oreochromis niloticus and Human Lymphocytes Exposed to Glyphosate" Current Issues in Molecular Biology 44, no. 5: 2230-2242. https://doi.org/10.3390/cimb44050151