
Neural Marker of Habituation at 5 Months of Age Associated with Deferred Imitation Performance at 12 Months: A Longitudinal Study in the UK and The Gambia


Abstract
:1. Introduction
1.1. Development of Deferred Imitation Abilities across Infancy
1.2. Imitation Behaviours across Cultures
1.3. Neural Markers Associated with Deferred Imitation
1.4. Cross-Cultural Development of Early Neural Markers of Habituation and Novelty Detection
1.5. Aims and Hypotheses
2. Method
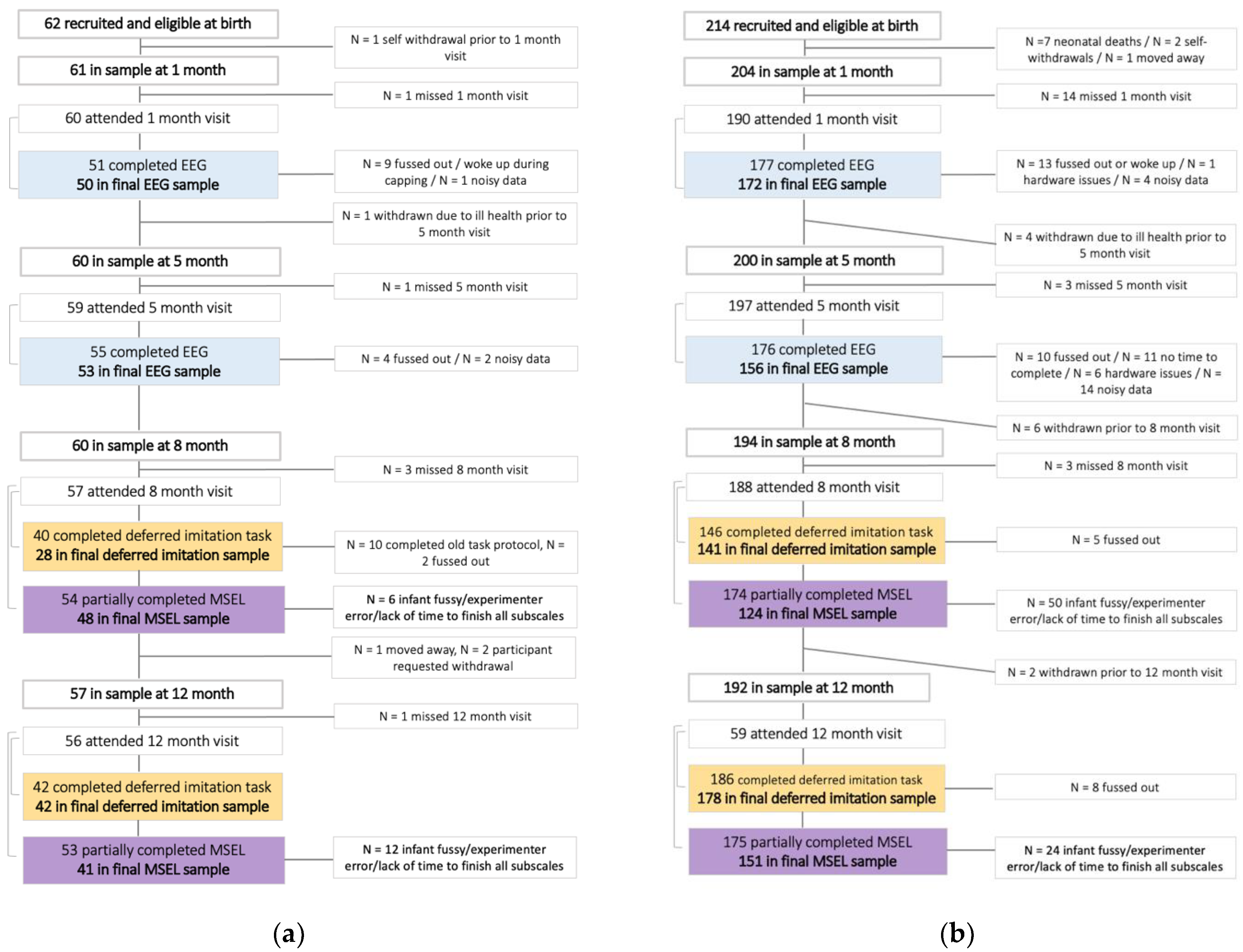
2.1. Participants
2.2. ERP Study
2.2.1. Procedure
2.2.2. Pre-Processing
2.3. Deferred Imitation
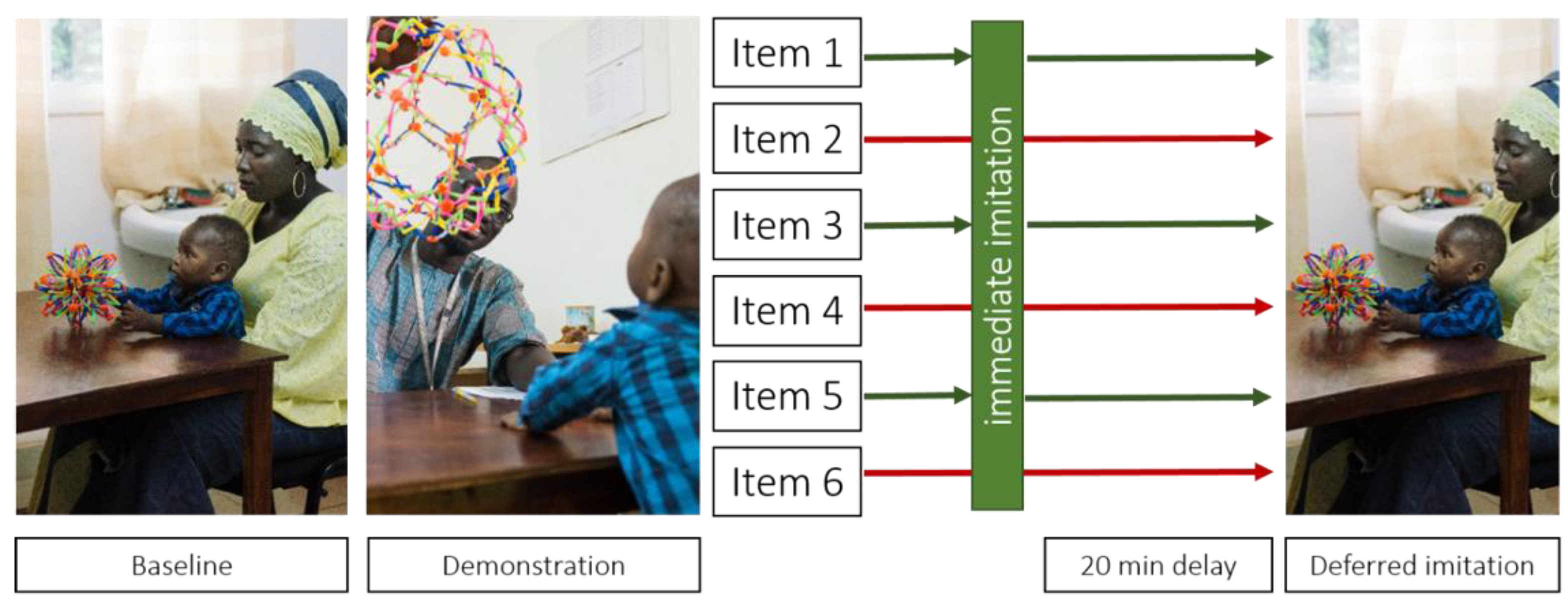
2.3.1. Procedure
2.3.2. Task Design and Reliability Assessments
2.3.3. Scoring and Outcome Variables
2.4. Mullen Scales of Early Learning (MSEL)
MSEL Scoring and Outcome Variables
2.5. Statistical Analyses
3. Results
3.1. Data Rejection and Sample Characteristics
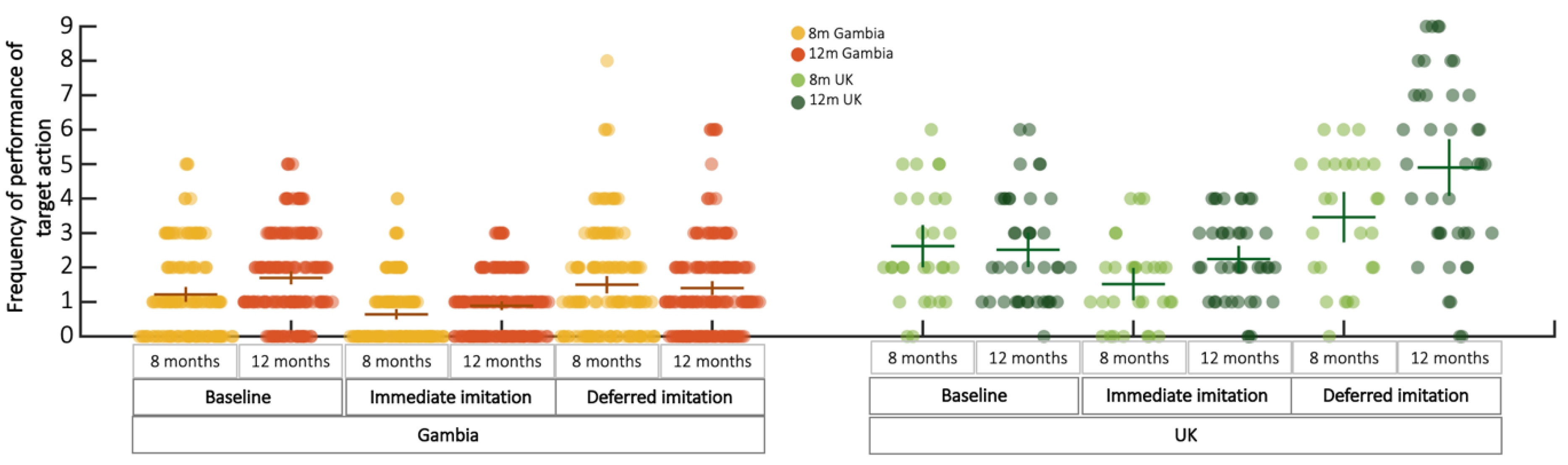
3.2. Deferred Imitation Behavioural Results
4. Discussion
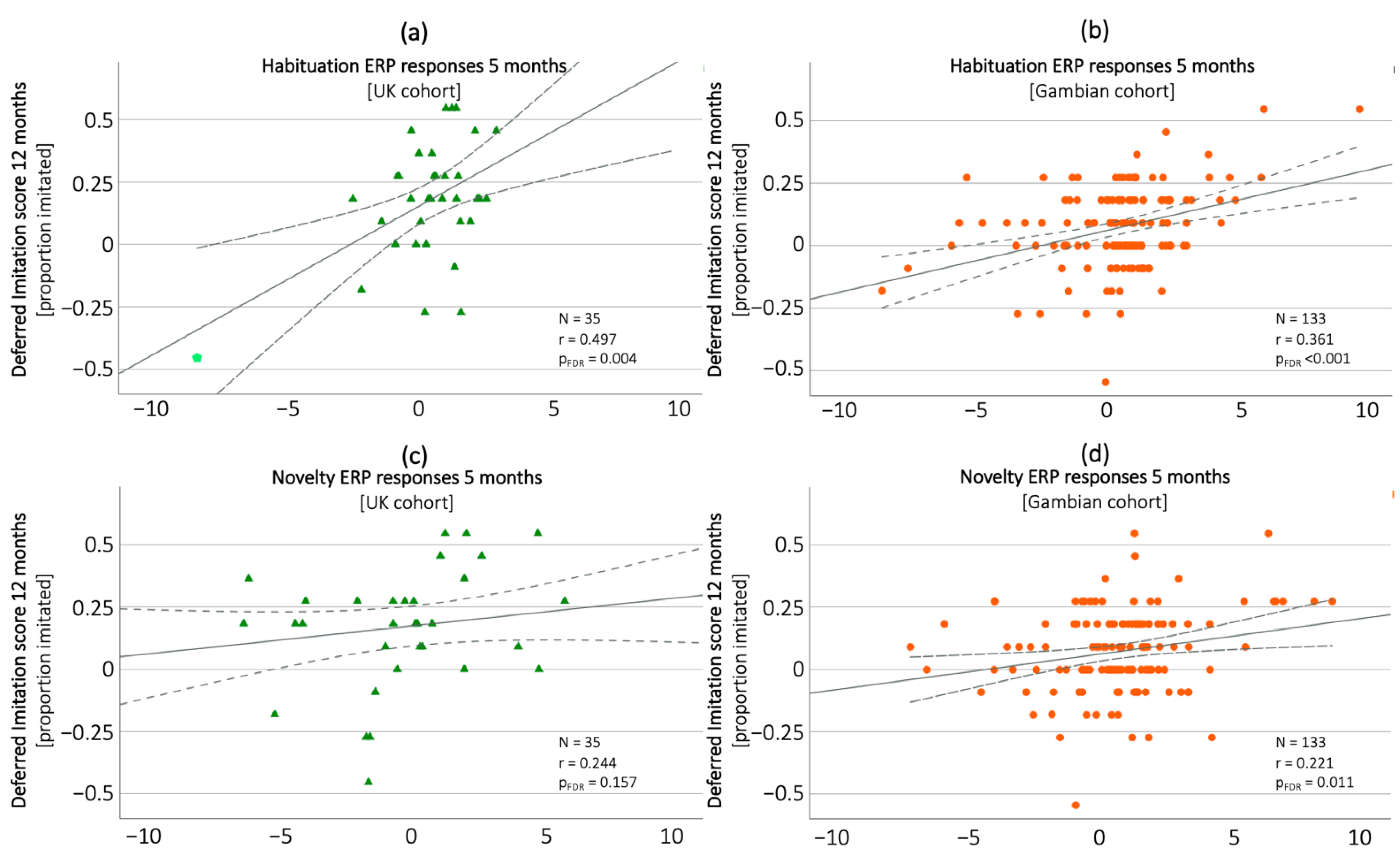
4.1. Developmental Changes in Deferred Imitation and Association with ERP Markers
4.2. Domain-Specific Links of Deferred Imitation with ERP Markers and MSEL Scales
4.3. Strengths and Limitations
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bandura, A. Social-learning theory of identificatory processes. Handb. Social. Theory Res. 1969, 213, 262. [Google Scholar]
- Kawai, M. Newly-acquired pre-cultural behavior of the natural troop of Japanese monkeys on Koshima Islet. Primates 1965, 6, 1–30. [Google Scholar] [CrossRef]
- Rumbaugh, D.M.; Savage-Rumbaugh, E.S. Language in comparative perspective. In Animal Learning and Cognition; Mackintosh, N.J., Ed.; Academic Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Piaget, J. Play, Dreams and Imitation in Childhood; Routledge: San Diego, CA, USA, 2013. [Google Scholar]
- De Haan, M.; Mishkin, M.; Baldeweg, T.; Vargha-Khadem, F. Is deferred imitation a test of cognitive recall? Trends Neurosci. 2006, 7, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Heimann, M.; Nordqvist, E.; Rudner, M.; Johansson, M.; Lindgren, M. Associative learning measured with ERP predicts deferred imitation using a strict observation only design in 14 to 15 month old children. Scand. J. Psychol. 2013, 54, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Meltzoff, A.N. Immediate and deferred imitation in fourteen-and twenty-four-month-old infants. Child Dev. 1985, 56, 62–72. [Google Scholar]
- Meltzoff, A.N. Infant imitation and memory: Nine-month-olds in immediate and deferred tests. Child Dev. 1988, 59, 217. [Google Scholar] [CrossRef]
- Meltzoff, A.N.; Moore, M.K. Newborn infants imitate adult facial gestures. Child Dev. 1983, 54, 702–709. [Google Scholar] [CrossRef]
- Jones, E.J.; Herbert, J.S. Exploring memory in infancy: Deferred imitation and the development of declarative memory. Infant Child Dev. Int. J. Res. Pract. 2006, 15, 195–205. [Google Scholar] [CrossRef]
- Barr, R.; Dowden, A.; Hayne, H. Developmental changes in deferred imitation by 6-to 24-month-old infants. Infant Behav. Dev. 1996, 19, 159–170. [Google Scholar] [CrossRef]
- Carver, L.J.; Bauer, P.J.; Nelson, C.A. Associations between infant brain activity and recall memory. Dev. Sci. 2000, 3, 234–246. [Google Scholar] [CrossRef]
- Bauer, P.J.; Wenner, J.; Dropik, P.L.; Wewerka, S.S. Parameters of remembering and forgetting in the transition from infancy to early childhood. Monogr. Soc. Res. Child Dev. 2000, 65, i–vi, 1–204. [Google Scholar] [PubMed]
- Hayne, H.; Boniface, J.; Barr, R. The development of declarative memory in human infants: Age-related changes in deffered imitation. Behav. Neurosci. 2000, 114, 77. [Google Scholar] [CrossRef] [PubMed]
- Learmonth, A.E.; Lamberth, R.; Rovee-Collier, C. Generalization of deferred imitation during the first year of life. J. Exp. Child Psychol. 2004, 88, 297–318. [Google Scholar] [CrossRef] [PubMed]
- Klein, P.J.; Meltzoff, A.N. Long-term memory, forgetting, and deferred imitation in 12-month-old infants. Dev. Sci. 1999, 2, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Meltzoff, A. The role of imitation in understanding persons and developing theory of mind. In Understanding Other Minds: Perspectives from Autism; Oxford University Press: New York, NY, USA, 1993; pp. 335–366. [Google Scholar]
- Barr, R.; Hayne, H. The effect of event structure on imitation in infancy: Practice makes perfect? Infant Behav. Dev. 1996, 19, 253–257. [Google Scholar] [CrossRef]
- Hayne, H.; MacDonald, S.; Barr, R. Developmental changes in the specificity of memory over the second year of life. Infant Behav. Dev. 1997, 20, 233–245. [Google Scholar] [CrossRef]
- Fivush, R.; Kuebli, J.; Clubb, P.A. The structure of events and event representations: A developmental analysis. Child Dev. 1992, 63, 188–201. [Google Scholar] [CrossRef]
- Hudson, J.; Nelson, K. Repeated encounters of a similar kind: Effects of familiarity on children’s autobiographic memory. Cogn. Dev. 1986, 1, 253–271. [Google Scholar] [CrossRef]
- Murachver, T.; Pipe, M.E.; Gordon, R.; Owens, J.L. Hooked on scripts: Generalized event memories acquired through direct experience and stories. In Proceedings of the Biennial Meeting of the Society for Research in Child Development, New Orleans, LA, USA, 25–28 March 1993. [Google Scholar]
- Bauer, P.J.; Hertsgaard, L.A.; Wewerka, S.S. Effects of experience and reminding on long-term recall in infancy: Remembering not to forget. J. Exp. Child Psychol. 1995, 59, 260–298. [Google Scholar] [CrossRef]
- Bauer, P.J.; Mandler, J.M. One thing follows another: Effects of temporal structure on 1-to 2-year-olds’ recall of events. Dev. Psychol. 1989, 25, 197. [Google Scholar] [CrossRef]
- Bauer, P.J.; Shore, C.M. Making a memorable event: Effects of familiarity and organization on young children’s recall of action sequences. Cogn. Dev. 1987, 2, 327–338. [Google Scholar] [CrossRef]
- Eckerman, C.O.; Whitehead, H. How toddler peers generate coordinated action: A cross-cultural exploration. Early Educ. Dev. 1999, 10, 241–266. [Google Scholar] [CrossRef]
- Nielsen, M.; Tomaselli, K. Overimitation in Kalahari Bushman children and the origins of human cultural cognition. Psychol. Sci. 2010, 21, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Goertz, C.; Lamm, B.; Graf, F.; Kolling, T.; Knopf, M.; Keller, H. Deferred imitation in 6-month-old German and Cameroonian Nso infants. J. Cogn. Educ. Psychol. 2011, 10, 44–55. [Google Scholar] [CrossRef]
- Graf, F.; Borchert, S.; Lamm, B.; Goertz, C.; Kolling, T.; Fassbender, I.; Teubert, M.; Vierhaus, M.; Freitag, C.; Spangler, S.; et al. Imitative learning of Nso and German infants at 6 and 9 months of age: Evidence for a cross-cultural learning tool. J. Cross Cult. Psychol. 2014, 45, 47–61. [Google Scholar] [CrossRef]
- Teiser, J.; Lamm, B.; Boning, M.; Graf, F.; Gudi, H.; Goertz, C.; Fassbender, I.; Freitag, C.; Spangler, S.; Teubert, M.; et al. Deferred imitation in 9-month-olds: How do model and task characteristics matter across cultures? Int. J. Behav. Dev. 2014, 38, 247–254. [Google Scholar] [CrossRef]
- Correa-Chávez, M.; Rogoff, B. Children’s attention to interactions directed to others: Guatemalan mayan and european american patterns. Dev. Psychol. 2009, 45, 630. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, A.C.; Gwaltney, A.; Raspa, M.; Okoniewski, K.C.; Berry-Kravis, E.; Botteron, K.N.; Budimirovic, D.; Hazlett, H.C.; Hessl, D.; Losh, M.; et al. Emergence of developmental delay in infants and toddlers with an FMR1 mutation. Pediatrics 2021, 147, e2020011528. [Google Scholar] [CrossRef]
- Yaari, M.; Mankuta, D.; Gadassi, A.H.; Friedlander, E.; Bar-Oz, B.; Eventov-Friedman, S.; Maniv, N.; Zucker, D.; Yirmiya, N. Early developmental trajectories of preterm infants. Res. Dev. Disabil. 2018, 81, 12–23. [Google Scholar] [CrossRef]
- Bauer, P.J.; Wiebe, S.A.; Carver, L.J.; Waters, J.M.; Nelson, C.A. Developments in long-term explicit memory late in the first year of life: Behavioral and electrophysiological indices. Psychol. Sci. 2003, 14, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Bauer, P.J.; Wiebe, S.A.; Carver, L.J.; Lukowski, A.F.; Haight, J.C.; Waters, J.M.; Nelson, C.A. Electrophysiological indexes of encoding and behavioral indexes of recall: Examining relations and developmental change late in the first year of life. Dev. Neuropsychol. 2006, 29, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Morasch, K.C.; Bell, M.A. Patterns of brain-electrical activity during declarative memory performance in 10-month-old infants. Brain Cogn. 2009, 71, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Nordqvist, E.; Rudner, M.; Johansson, M.; Lindgren, M.; Heimann, M. The relationship between deferred imitation, associative memory, and communication in 14-months-old children. Behavioral and electrophysiological indices. Front. Psychol. 2015, 6, 260. [Google Scholar] [CrossRef] [Green Version]
- Katus, L.; Mason, L.; Milosavljevic, B.; McCann, S.; Rozhko, M.; Moore, S.E.; Elwell, C.E.; Lloyd-Fox, S.; de Haan, M.; Drammeh, S.; et al. ERP markers are associated with neurodevelopmental outcomes in 1–5 month old infants in rural Africa and the UK. NeuroImage 2020, 210, 116591. [Google Scholar] [CrossRef]
- Lloyd-Fox, S.; Blasi, A.; McCann, S.; Rozhko, M.; Katus, L.; Mason, L.; Austin, T.; Moore, S.E.; Elwell, C.E.; The BRIGHT Project Team. Habituation and novelty detection fNIRS brain responses in 5-and 8-month-old infants: The Gambia and UK. Dev. Sci. 2019, 22, e12817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunn, P.G.; Northrop-Clewes, C.A.; Downes, R.M. Intestinal permeability, mucosal injury, and growth faltering in Gambian infants. Lancet 1991, 338, 907–910. [Google Scholar] [CrossRef]
- Kushnerenko, E.; Winkler, I.; Horváth, J.; Näätänen, R.; Pavlov, I.; Fellman, V.; Huotilainen, M. Processing acoustic change and novelty in newborn infants. Eur. J. Neurosci. 2007, 26, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Milosavljevic, B.; Vellekoop, P.; Maris, H.; Halliday, D.; Drammeh, S.; Sanyang, L.; Darboe, M.K.; Elwell, C.; Moore, S.E.; Lloyd-Fox, S. Adaptation of the Mullen Scales of Early Learning for use among infants aged 5-to 24-months in rural Gambia. Dev. Sci. 2019, 22, e12808. [Google Scholar] [CrossRef] [Green Version]
- Mullen, E.M. Mullen Scales of Early Learning; American Guidance Service: Circle Pines, MN, USA, 1995; pp. 58–64. [Google Scholar]
- Delehanty, A.D.; Stronach, S.; Guthrie, W.; Slate, E.; Wetherby, A.M. Verbal and nonverbal outcomes of toddlers with and without autism spectrum disorder, language delay, and global developmental delay. Autism Dev. Lang. Impair. 2018, 3, 2396941518764764. [Google Scholar] [CrossRef] [Green Version]
- Jeffreys, H. Theory of Probability, 3rd ed.; MR0187257; Oxford University Press: Oxford, UK, 1961; p. 432. [Google Scholar]
- Carver, L.J.; Bauer, P.J. The dawning of a past: The emergence of long-term explicit memory in infancy. J. Exp. Psychol. Gen. 2001, 130, 726. [Google Scholar] [CrossRef]
- Otte, R.; Winkler, I.; Braeken, M.; Stekelenburg, J.; van der Stelt, O.; Bergh, B.R.V.D. Detecting violations of temporal regularities in waking and sleeping two-month-old infants. Biol. Psychol. 2013, 92, 315–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Heuvel, M.I.; Otte, R.A.; Braeken, M.A.; Winkler, I.; Kushnerenko, E.; Bergh, B.R.V.D. Differences between human auditory event-related potentials (AERPs) measured at 2 and 4 months after birth. Int. J. Psychophysiol. 2015, 97, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adlam, A.-L.R.; Vargha-Khadem, F.; Mishkin, M.; de Haan, M. Deferred imitation of action sequences in developmental amnesia. J. Cogn. Neurosci. 2005, 17, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infant Characteristics | ||||||||
|---|---|---|---|---|---|---|---|---|
| Cohort Gambia | Cohort UK | |||||||
| Included | Excluded | Included | Excluded | |||||
| Sex (% female) | 49.9 | 50 | 46.3 | 55 | ||||
| 1 month | 5 months | 1 month | 5 months | |||||
| Included | Excluded | Included | Excluded | Included | Excluded | Included | Excluded | |
| X ± SD | X ± SD | X ± SD | X ± SD | X ± SD | X ± SD | X ± SD | X ± SD | |
| Age (days) | 42.22 ± 25.99 | 47.43 ± 32.51 | 159.51 ± 9.78 | 162.7 ± 12.85 | 32.77 ± 5.29 | 35.0 ± 6.74 | 155.74 ± 6.95 | 155.89 ± 5.77 |
| Weight (kg) | 4.23 ± 0.60 | 4.31 ± 0.54 | 6.84 ± 0.77 | 6.78 ± 0.89 | 4.35 ± 0.52 | 4.38 ± 0.58 | 7.19 ± 0.92 | 7.07 ± 0.85 |
| Length (cm) | 53.01 ± 2.08 | 53.08 ± 1.86 | 64.07 ± 2.06 | 64.21 ± 2.76 | 53.93 ± 2.17 | 54. 17 ± 1.93 | 64.42 ± 2.19 | 64.56 ± 2.32 |
| Head circumference (cm) | 36.58 ± 1.17 | 36.74 ± 1.19 | 41.26 ± 1.29 | 41.31 ± 1.46 | 37.34 ± 1.21 | 37.96 ± 1.02 | 42.84 ± 1.24 | 43.26 ± 1.15 |
| Weight-for-age | −0.55 ± 0.95 | −0.43 ± 0.85 | −0.61 ± 0.95 | −0.75 ± 1.09 | −0.14 ± 0.81 | −0.25 ± 1.02 | −0.16 ± 1.12 | −0.18 ± 0.95 |
| Length-for-age | −0.93 ± 0.96 | −0.56 ± 0.86 | −0.60 ± 0.92 | −0.631 ± 1.92 | −0.30 ± 1.07 | −0.32 ± 0.97 | −0.38 ± 0.97 | −0.23 ± 1.13 |
| Head circumference for age | −0.59 ± 0.88 | −0.47 ± 0.944 | −0.74 ± 0.91 | −0.78 ± 0.99 | 0.57 ± 0.90 | 0.67 ± 0.72 | 0.56 ± 0.94 | 0.99 ± 0.69 |
| Weight-for-length | 0.40 ± 1.08 | 0.14 ± 1.05 | −0.23 ± 1.06 | −0.37 ± 0.96 | 0.15 ± 1.06 | −0.28 ± 1.09 | 0.17 ± 1.22 | 0.04 ± 0.95 |
| Mullen Scales of Early Learning—VDQ | not collected | not collected | 12.45 ± 2.71 | 12.91 ± 1.95 | not collected | not collected | 11.16 ± 2.09 | 11.41 ± 1.37 |
| Mullen Scales of Early Learning—PDQ | not collected | not collected | 14.74 ± 2.79 | 14.7 ± 2.245 | not collected | not collected | 14.17 ± 2.43 | 14.87 ± 1.59 |
| Maternal Characteristics at Birth | ||||
|---|---|---|---|---|
| Cohort Gambia | Cohort UK | |||
| Included | Excluded | Included | Excluded | |
| Maternal age | 29.29 ± 6.55 | 29.95 ± 6.723 | 33.07 ± 2.99 | 32.81 ± 2.91 |
| Parity | 4.38 ± 2.39 | 4.38 ± 2.92 | 1.25 ± 0.49 | 1.45 ± 0.61 |
| Gestational age | 39.84 ± 1.93 | 39.42 ± 1.97 | 40.37 ± 1.29 | 39.98 ± 1.36 |
| Cohort Gambia | Cohort UK | |||
|---|---|---|---|---|
| 8 Months | 12 Months | 8 Months | 12 Months | |
| n | 144 | 182 | 29 | 41 |
| X ± SD | X ± SD | X ± SD | X ± SD | |
| Baseline | 1.500 ± 1.537 | 1.401 ± 1.390 | 2.621 ± 1.613 | 2.512 ± 1.583 |
| Immediate | 0.640 ± 0.910 | 0.890 ± 0.879 | 1.517 ± 1.242 | 2.244 ± 1.220 |
| Deferred | 1.217 ± 1.294 | 1.698 ± 1.202 | 3.462 ± 1.861 | 4.902 ± 2.615 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katus, L.; Milosavljevic, B.; Rozhko, M.; McCann, S.; Mason, L.; Mbye, E.; Touray, E.; Moore, S.E.; Elwell, C.E.; Lloyd-Fox, S.; et al. Neural Marker of Habituation at 5 Months of Age Associated with Deferred Imitation Performance at 12 Months: A Longitudinal Study in the UK and The Gambia. Children 2022, 9, 988. https://doi.org/10.3390/children9070988
Katus L, Milosavljevic B, Rozhko M, McCann S, Mason L, Mbye E, Touray E, Moore SE, Elwell CE, Lloyd-Fox S, et al. Neural Marker of Habituation at 5 Months of Age Associated with Deferred Imitation Performance at 12 Months: A Longitudinal Study in the UK and The Gambia. Children. 2022; 9(7):988. https://doi.org/10.3390/children9070988
Chicago/Turabian StyleKatus, Laura, Bosiljka Milosavljevic, Maria Rozhko, Samantha McCann, Luke Mason, Ebrima Mbye, Ebou Touray, Sophie E. Moore, Clare E. Elwell, Sarah Lloyd-Fox, and et al. 2022. "Neural Marker of Habituation at 5 Months of Age Associated with Deferred Imitation Performance at 12 Months: A Longitudinal Study in the UK and The Gambia" Children 9, no. 7: 988. https://doi.org/10.3390/children9070988