Photosynthetic Pigments Changes of Three Phenotypes of Picocyanobacteria Synechococcus sp. under Different Light and Temperature Conditions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
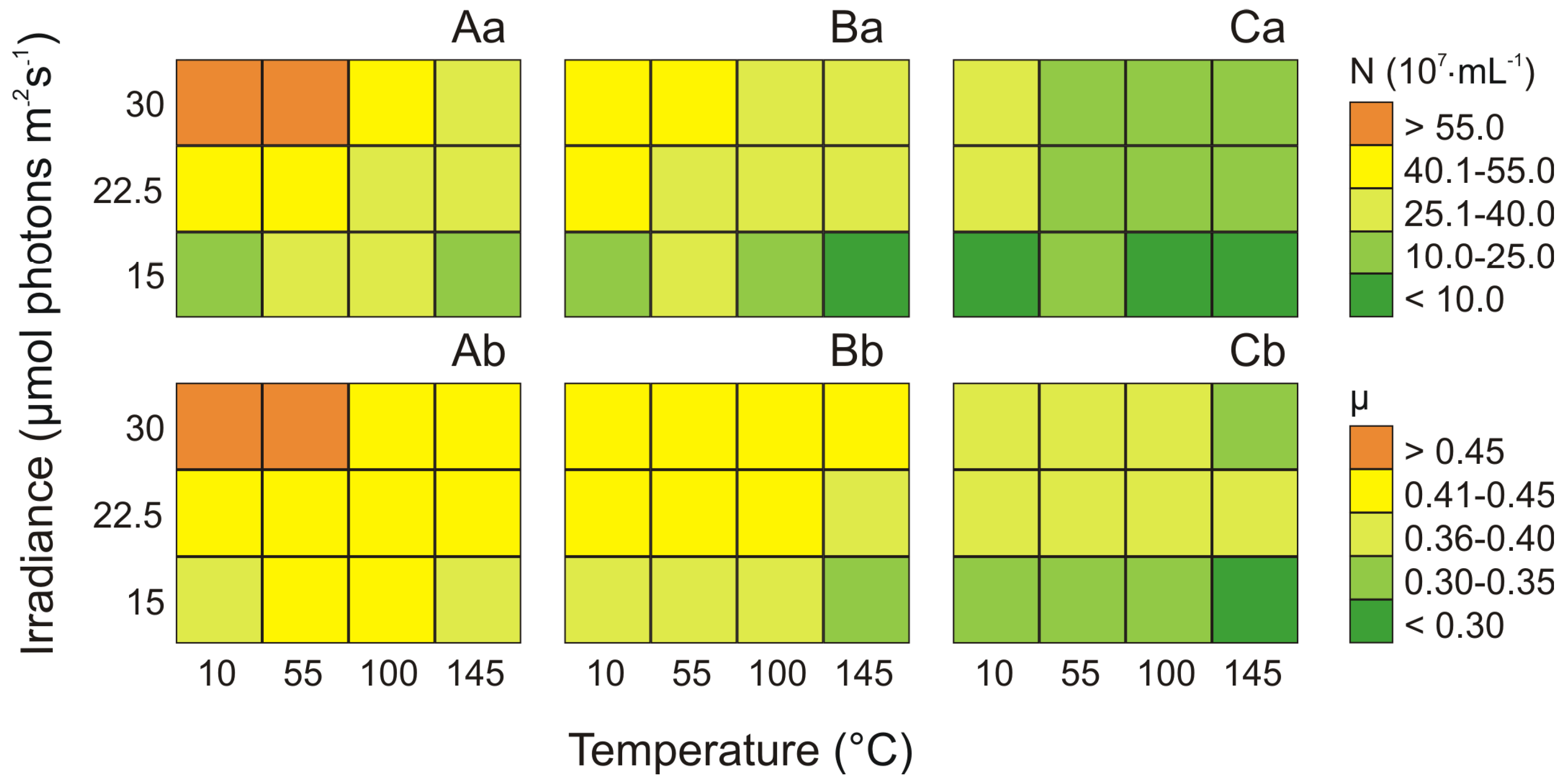
2.1. The Cell Concentration and the Growth Rate of Three Synechococcus sp. Phenotypes under Different Culture Conditions
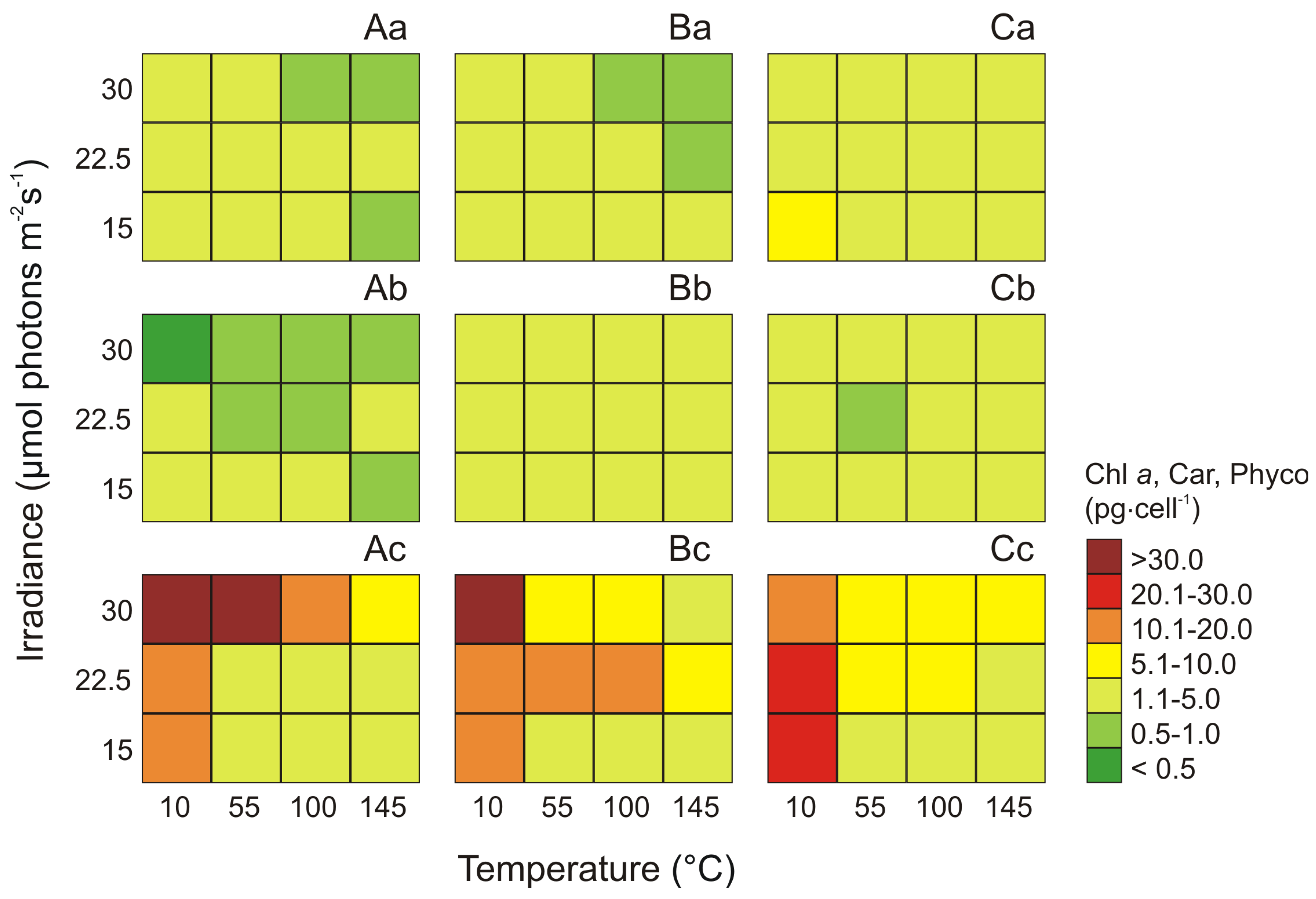
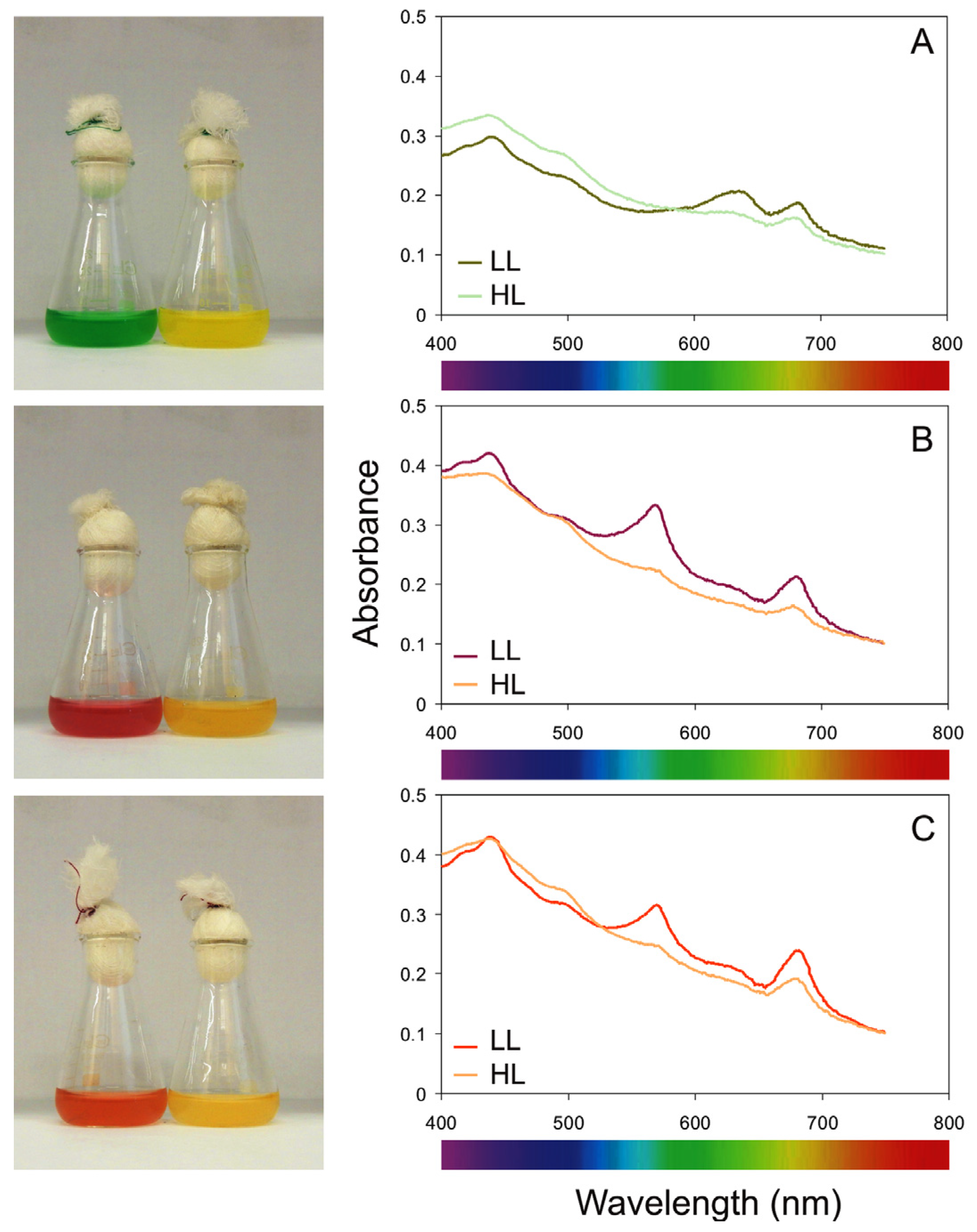
2.2. The Total Pigments Content for Three Phenotypes of the Genus Synechococcus
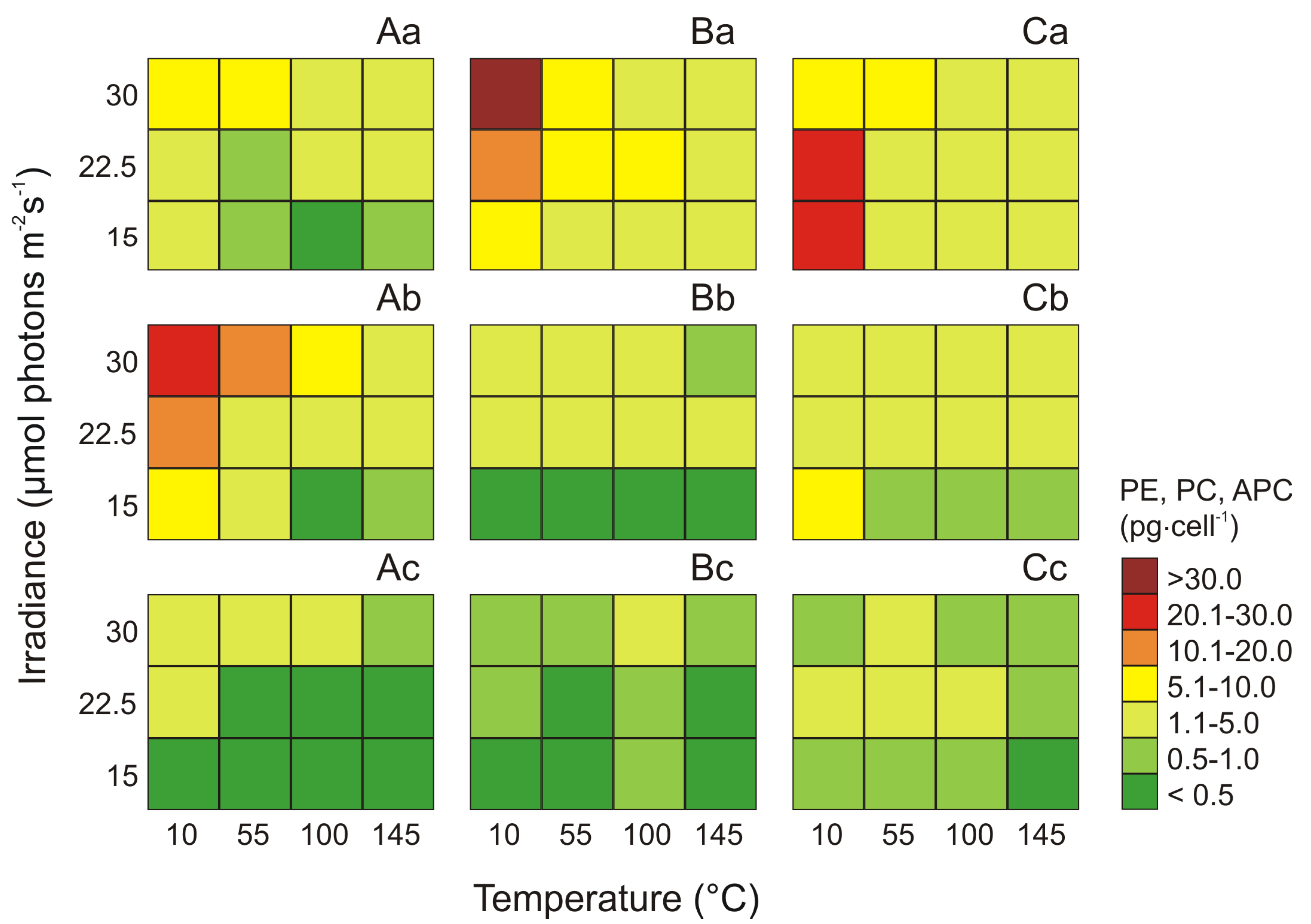
2.3. Effect of Irradiance and Temperature on Phycocyanin, Phycoerythrin, and Allophycocyanin Content
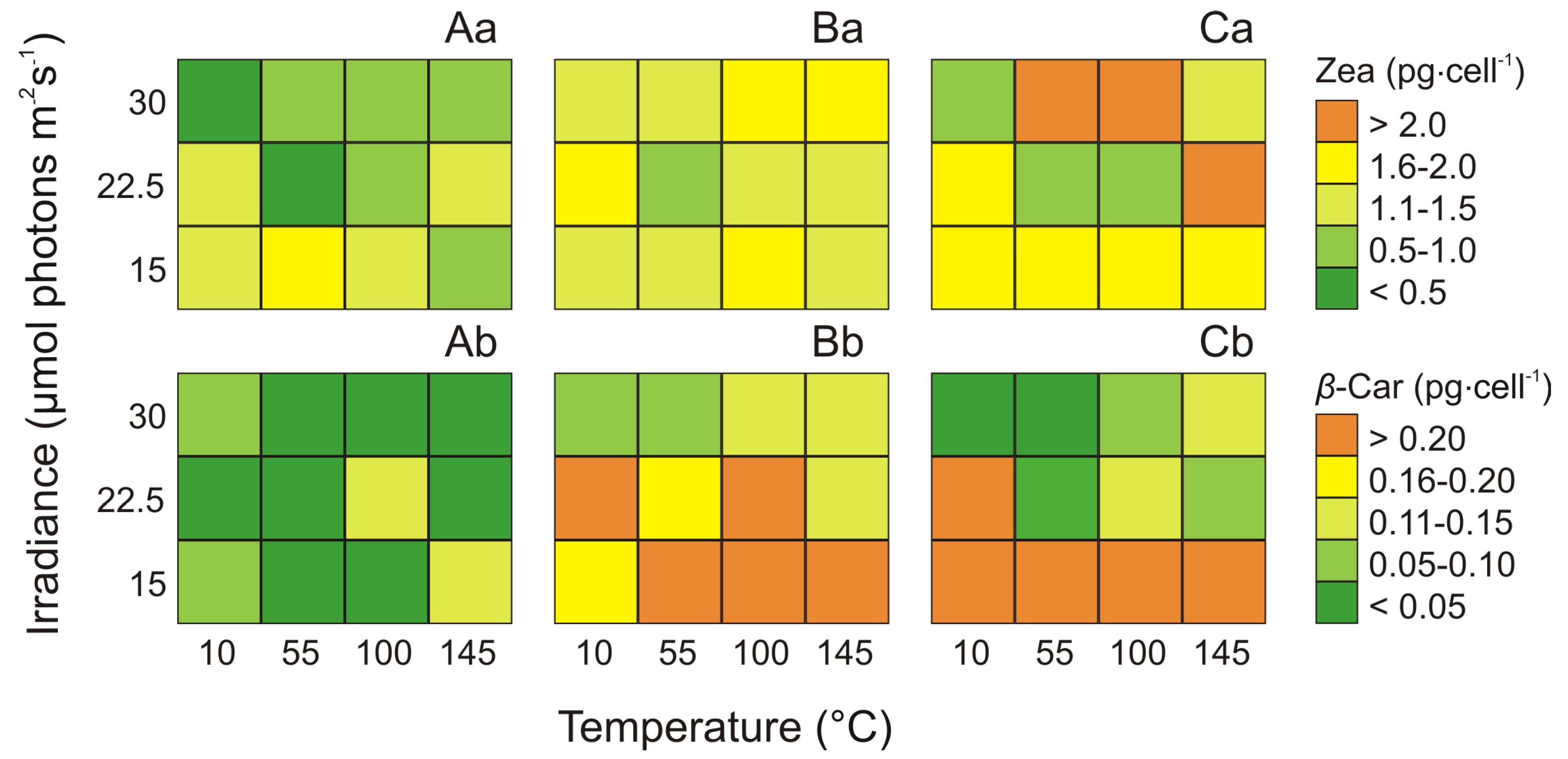
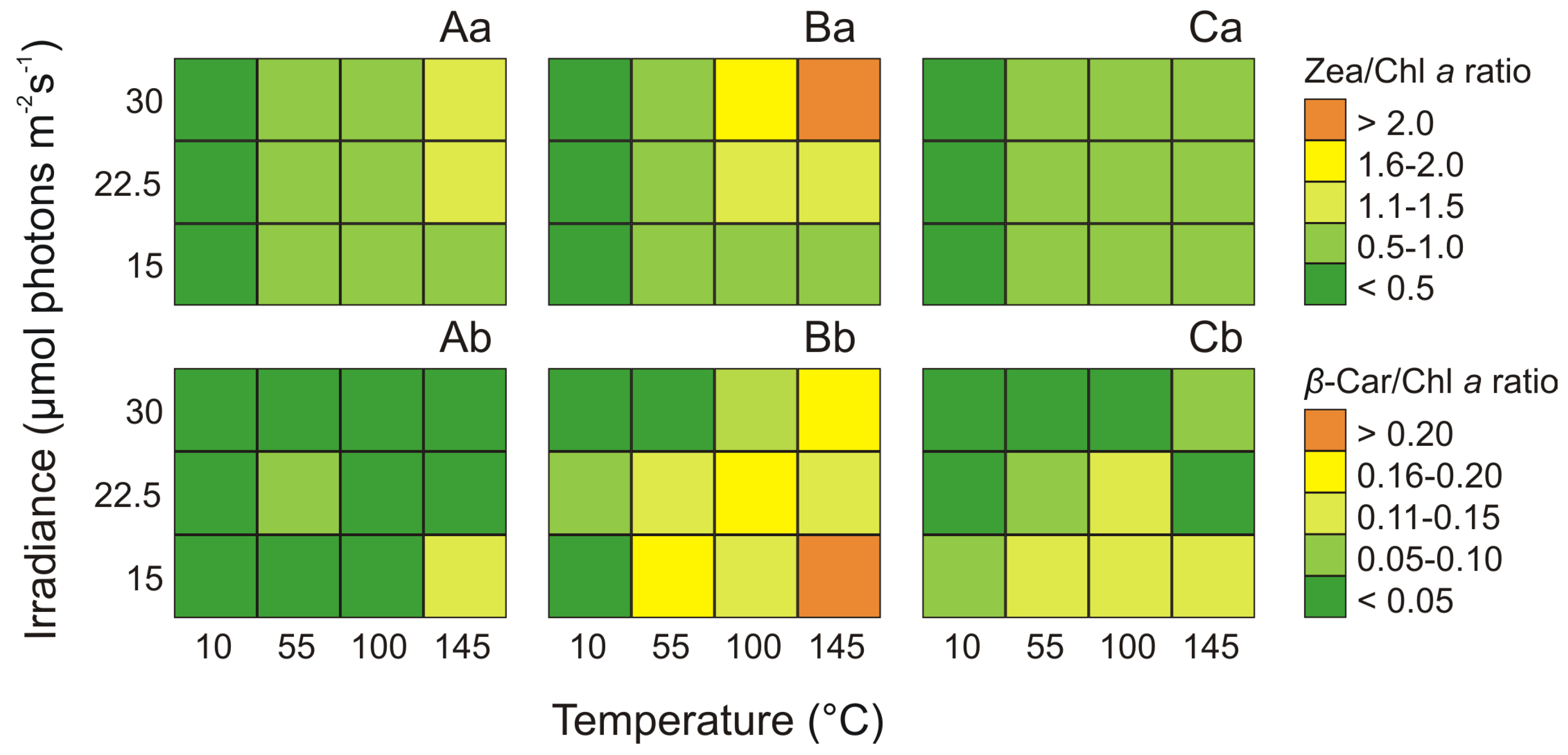
2.4. Effect of Irradiance and Temperature on Zeaxanthin and β-carotene
2.5. Effect of Irradiance and Temperature on Pigments Ratios
3. Discussion
3.1. Occurrence and Abundance of Picocyanobacteria under Changing Irradiance and Temperature Conditions
3.2. Changes in Pigments Content and Pigment Ratios under Different Irradiance and Temperature Conditions
4. Materials and Methods
4.1. Culture Conditions
4.2. Calculation of Cell Density and Growth Rates
4.3. Determination of the Chlorophyll and Carotenoids Content
4.4. Determination of the Phycobiliproteins Content
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, P.W.; Sieburth, J.M. Chroococcoid cyanobacteria in the sea: A ubiquitous and diverse phototrophic biomass. Limnol. Oceanogr. 1979, 24, 928–935. [Google Scholar] [CrossRef]
- Waterbury, J.B.; Watson, S.W.; Guillard, R.R.L.; Brand, L.E. Widespread occurrence of a unicellular, marine, planktonic cyanobacterium. Nature 1979, 277, 293–294. [Google Scholar] [CrossRef]
- Stockner, J.G. Phototrophic picoplankton: An overview from marine and freshwater ecosystems. Limnol. Oceanogr. 1988, 33, 765–775. [Google Scholar] [CrossRef]
- Jasser, I.; Lehtovaara, A.; Arvola, L. Seasonality and coexistence of autotrophic pico- and nanoplankton and zooplankton in three boreal lakes. Internationale Vereinigung für Theoretische und Angewandte Limnologie Verhandlungen 2006, 29, 1413–1416. [Google Scholar] [CrossRef]
- Mühling, M.; Woolven-Allen, J.; Murrell, J.C.; Joint, I. Improved group-specific PCR primers for denaturing gradient gel electrophoresis analysis of the genetic diversity of complex microbial communities. ISME J. 2008, 2, 379–392. [Google Scholar] [CrossRef]
- Guidi, L.; Chaffron, S.; Bittner, L.; Eveillard, D.; Larhlimi, A.; Roux, S.; Darzi, Y.; Audic, S.; Berline, L.; Brum, J.R.; et al. Plankton networks driving carbon export in the oligotrophic ocean. Nature 2016, 532, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Worden, A.Z.; Wilken, S. A plankton bloom shifts as the ocean warms. Science 2016, 354, 287–288. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincon, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.; Lomas, M.W.; Veneziano, D. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, Z.; Jing, Z.; Zhou, L.; Li, G.; Ke, Z.; Jiang, X.; Liu, J.; Liu, H.; Tan, Y. Synechococcus bloom in the Pearl River Estuary and adjacent coastal area–With special focus on flooding during wet seasons. Sci. Total Environ. 2019, 692, 769–783. [Google Scholar] [CrossRef]
- Oziel, L.; Neukermans, G.; Ardyna, M.; Lancelot, C.; Tison, J.-L.; Wassmann, P.; Sirven, J.; Ruiz-Pino, D.; Gascard, J.-C. Role for Atlantic inflows and sea ice loss on shifting phytoplankton blooms in the Barents Sea. J. Geophys. Res. Oceans. 2017, 122, 5121–5139. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, M.L.; Doré, H.; Garczarek, L.; Seuthe, L.; Müller, O.; Sandaa, R.A.; Bratbak, G.; Larsen, A. Synechococcus in the Atlantic Gateway to the Arctic Ocean. Front. Mar. Sci. 2016, 3, 191. [Google Scholar] [CrossRef] [Green Version]
- Dutkiewicz, S.; Morris, J.J.; Follows, M.J.; Scott, J.; Levitan, O.; Dyhrman, S.T.; Berman-Frank, I. Impact of ocean acidification on the structure of future phytoplankton communities. Nat. Clim. Change 2015, 5, 1002–1006. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Cieszyńska, A.; Maculewicz, J.; Latała, A. Ecophysiological characteristics of red, green, and brown strains of the Baltic picocyanobacterium Synechococcus sp.-a laboratory study. Biogeosciences 2018, 15, 6257–6276. [Google Scholar] [CrossRef] [Green Version]
- Stransky, H.; Hager, A. Das Carotinoid-muster und die Verbreitung des lichtinduzierten Xanthophyllcyclus in verschiedenen Algenklassen. Cyanophyceae und Rhodophyceae. Arch. Mikrobiol. 1970, 72, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, L.; Lauridsen, T.L.; Krogh, G.; Jørgensen, T. Identification and quantification of phytoplankton groups in lakes using new pigment ratios―a comparison between pigment analysis by HPLC and microscopy. Freshw. Biol. 2006, 51, 1474–1485. [Google Scholar] [CrossRef]
- Kana, T.M.; Glibert, P.M. Zeaxanthin and B-carotene in Synechococcus WH7803 respond differently to irradiance. Limnol. Oceanogr. 1988, 33, 1623–1627. [Google Scholar]
- Ostrowska, M.; Woźniak, B.; Dera, J. Modelled quantum yields and energy efficiency of fluorescence, photosynthesis and heat production by phytoplankton in the World Ocean. Oceanologia 2012, 54, 565–610. [Google Scholar] [CrossRef]
- Hauschild, C.A.; McMurter, H.J.G.; Pick, F.R. Effect of spectral quality on growth and pigmentation of picocyanobacteria. J. Phycol. 1991, 27, 698–702. [Google Scholar] [CrossRef]
- Cornejo, J.; Beale, S.I. Phycobilin biosynthetic reactions in extracts of cyanobacteria. Photosynth. Res. 1997, 51, 223–230. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Barreiro Felpeto, A.; Latała, A. Allelopathic and bloom-forming picocyanobacteria in a changing world. Toxins 2018, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Konarzewska, Z.; Śliwińska-Wilczewska, S.; Barreiro Felpeto, A.; Vasconcelos, V.; Latała, A. Assessment of the Allelochemical Activity and Biochemical Profile of Different Phenotypes of Picocyanobacteria from the Genus Synechococcus. Mar. Drugs 2020, 18, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Six, C.; Thomas, J.C.; Garczarek, L.; Ostrowski, M.; Dufresne, A.; Blot, N.; Scanlan, D.J.; Partensky, F. Diversity and evolution of phycobilisomes in marine Synechococcus spp.: A comparative genomics study. Genome Biol. 2007, 8, R259. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp, T.H.; Schouten, D.; Doeleman, M.; Wollenzien, U.; Huisman, J.; Stal, L.J. Colorful microdiversity of Synechococcus strains (picocyanobacteria) isolated from the Baltic Sea. ISME J. 2009, 3, 34–408. [Google Scholar] [CrossRef] [PubMed]
- Callieri, C. Synechococcus plasticity under environmental changes. FEMS Microbiol. Lett. 2017, 364, fnx229. [Google Scholar] [CrossRef]
- Roy, S.; Llewellyn, C.; Egeland, E.S.; Johnsen, G. Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Cambridge University Press: Cambridge, UK, 2011; p. 784. [Google Scholar]
- Morris, I.; Glover, H. Physiology of photosynthesis by marine coccoid Cyanobacteria -some ecological implications. Limnol. Oceanogr. 1981, 26, 957–961. [Google Scholar] [CrossRef]
- Antia, N.J.; Cheng, J.Y. The survival of axenic cultures of marine planktonic algae from prolonged exposure to darkness at 20 °C. Phycologia 1970, 9, 179–183. [Google Scholar] [CrossRef]
- Antia, N.J. Effects of temperature on the darkness survival of marine microplanktonic algae. Microb. Ecol. 1976, 3, 41–54. [Google Scholar] [CrossRef]
- Platt, T.; Subba-Rao, D.V.; Irwin, B. Photosynthesis of picoplankton in the oligotrophic ocean. Nature 1983, 301, 702–704. [Google Scholar] [CrossRef]
- Cai, H.; Wang, K.; Huang, S.; Jiao, N.; Chen, F. Distinct patterns of picocyanobacterial communities in winter and summer in the Chesapeake Bay. Appl. Environ. Microbiol. 2010, 76, 2955–2960. [Google Scholar] [CrossRef] [Green Version]
- Ernst, A. Cyanobacterial picoplankton from Lake Constance. Isolation by fluorescence characteristics. J. Plankton Res. 1991, 13, 1307–1312. [Google Scholar] [CrossRef]
- Jasser, I.; Callieri, C. Picocyanobacteria. In Handbook on Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 19–27. [Google Scholar]
- Glover, H.E. The physiology and ecology of marine Cyanobacteria, Synechococcus spp. In Advances in Applied Microbiology; Jannasch, H.W., Williams Leb, P.J., Eds.; Academic Press: New York, USA, 1985; pp. 49–107. [Google Scholar]
- Vörös, L.; Callieri, C.; Katalin, V.; Bertoni, R. Freshwater picocyanobacteria along a trophic gradient and light quality range. Hydrobiologia 1998, 369, 117–125. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Wang, P.; Zhang, S. Multiple effects of environmental factors on algal growth and nutrient thresholds for harmful algal blooms: Application of response surface methodology. Environ. Model. Assess. 2016, 21, 247–259. [Google Scholar] [CrossRef]
- Jasser, I.; Arvola, L. Potential effects of abiotic factors on the abundance of autotrophic picoplankton in four boreal lakes. J. Plankton Res. 2003, 25, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Jasser, I. The relationship between autotrophic picoplankton (APP)-the smallest autotrophic component of food web and the trophic status and depth of lakes. Ecohydrol. Hydrobiol. 2006, 6, 69–77. [Google Scholar] [CrossRef]
- Gławdel, M.; Mackiewicz, T.; Witek, Z. Composition and abundance of picoplankton in the coastal zone of the Gulf of Gdańsk. Oceanol. Stud. 1999, 28, 17–30. [Google Scholar]
- Barlow, R.G.; Alberte, R.S. Photosynthetic characteristic of phycoerythrin-containing marine Synechococcus spp. I. Responses to growth photon flux density. Mar. Biol. 1985, 86, 63–74. [Google Scholar] [CrossRef]
- Glover, H.E.; Campbell, L.; Prézelin, B.B. Contribution of Synechococcus spp. to size-fraction primary productivity in three waters masses in the Northwest Atlantic Ocean. Mar. Biol. 1986, 91, 193–203. [Google Scholar] [CrossRef]
- Campbell, L.; Carpenter, E.J. Diel patterns of cell division in marine Synechococcus spp. (Cyanobacteria): Use of the frequency of dividing cell technique to measure growth rate. Mar. Ecol. Prog. Ser. 1986, 32, 139–148. [Google Scholar] [CrossRef]
- Kana, T.M.; Glibert, P.M. Effect of irradiances up to 2000 μmol E·m−2·s−1 on marine Synechococcus WH7803-I. Growth, pigmentation, and cell composition. Deep-Sea Res. 1987, 34, 479–495. [Google Scholar] [CrossRef]
- Kana, T.M.; Glibert, P.M. Effect of irradiances up to 2000 μmol E·m−2·s−1 on marine Synechococcus WH7803-II. Photosynthetic responses and mechanisms. Deep-Sea Res. 1987, 34, 497–516. [Google Scholar] [CrossRef]
- Jodłowska, S.; Śliwińska, S. Effects of light intensity and temperature on the photosynthetic irradiance response curves and chlorophyll fluorescence in three picocyanobacterial strains of Synechococcus. Photosynthetica 2014, 52, 223–232. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Eviron. Microb. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Wilde, E.W.; Tilly, L.J. Structural characteristics of algal communities in thermally altered artificial streams. Hydrobiologia 1981, 76, 57–63. [Google Scholar] [CrossRef]
- Noaman, N.H.; Fattah, A.; Khaleafa, M.; Zaky, S.H. Factors affecting antimicrobial activity of Synechococcus leopoliensis. Microbiol. Res. 2004, 159, 395–402. [Google Scholar] [CrossRef]
- Robarts, R.D.; Zohary, T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. N. Z. J. Mar. Freshwat. Res. 1987, 21, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Weisse, T. The microbial food-web and its sensitivity to eutrophication and contaminant enrichment: Across-system overview. Int. Rev. Ges. Hydrobiol. 1991, 76, 327–338. [Google Scholar] [CrossRef]
- Kuosa, H. Picoplanktonic algae in the northern Baltic Sea: Seasonal dynamics and flagellate grazing. Mar. Ecol. Prog. Ser. 1991, 73, 269–276. [Google Scholar] [CrossRef]
- Kononen, K.; Kuparinen, J.; Mäkelä, K.; Laanemets, J.; Pavelson, J.; Nômmann, S. Initiation of cyanobacterial blooms in a frontal region at the entrance to the Gulf of Finland, Baltic Sea. Limnol. Ocean. 1996, 41, 98–112. [Google Scholar] [CrossRef]
- Stal, L.J.; Albertano, P.; Bergman, B.; Bröckel, K.; Gallon, J.R.; Hayes, P.K.; Sivonen, K.; Walsby, A.E.; BASIC: Baltic Sea cyanobacteria. An investigation of the structure and dynamics of water blooms of cyanobacteria in the Baltic Sea―responses to a changing environment. Cont. Shelf Res. 2003, 23, 1695–1714. [Google Scholar]
- Hanelt, D.; Wiencke, C.; Bischof, K. Photosynthesis in Algae. In Advances in Photosynthesis and Respiration; Larkum, A.W.D., Douglas, S.E., Raven, J.A., Eds.; Springer Science & Business Media: Berlin, Germany, 2003; Volume 14, p. 417. [Google Scholar]
- Stomp, M.; Huisman, J.; Stal, L.J.; Matthijs, H.C. Colorful niches of phototrophic microorganisms shaped by vibrations of the water molecule. ISME J. 2007, 1, 271–282. [Google Scholar] [CrossRef]
- Millie, D.F.; Ingram, D.A.; Dionigi, C.P. Pigment and photosynthetic responses of Oscillatoria agardhii (Cyanophyta) to photon flux density and spectral quality. J. Phycol. 1990, 26, 660–666. [Google Scholar] [CrossRef]
- Prézelin, B.B. Light reactions in photosynthesis. In Physiological Bases of Phytoplankton Ecology; Platt, T., Ed.; Canadian Bulletin of Fisheries and Aquatic Sciences: Ottawa, ON, Canada, 1981; pp. 1–43. [Google Scholar]
- Paerl, H.W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol. Oceanogr. 1988, 33, 823–847. [Google Scholar] [CrossRef]
- Carter, J.R. Diatom from the Devil’s Hole Cave, Fife; Nova Hedwigia: Scotland, UK, 1971; pp. 657–681. [Google Scholar]
- Lund, I.W.G. The ecology of the freshwater phytoplankton. Biol. Rev. Cambridge Philosoph. Soc. 1965, 40, 231–293. [Google Scholar] [CrossRef]
- Foy, R.H.; Gibson, C.E. Photosynthetic characteristics of planktonic blue-green algae: Changes in photosynthetic capacity and pigmentation of Oscillatoria redekei Van Goor under high and low light. Br. Phycol. J. 1982, 17, 183–193. [Google Scholar] [CrossRef]
- Post, A.F. Transient state characteristics of adaptation to changes in light conditions for the cyanobacterium Oscillatoria agardhii. I. Pigmentation and photosynthesis. Arch. Microbiol. 1986, 145, 353–357. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Murphy, L.S.; Foss, P.; Liaaen-Jensen, S. Synechococcus spp. as likely zeaxanthin-dominant ultraphytoplankton in the North Atlantic. Limnol. Oceanogr. 1985, 30, 412–414. [Google Scholar] [CrossRef]
- Gieskes, W.W.; Kraay, G.W. Floristic and physiological differences between the shallow and the deep nanophytoplankton community in the euphotic zone of the open tropical Atlantic revealed by HPLC analysis of pigments. Mar. Biol. 1986, 91, 567–576. [Google Scholar] [CrossRef]
- Dubinsky, Z.; Falkowski, P.J.; Wyman, K. Light harvesting and utilization by phytoplankton. Plant. Cell Physiol. 1986, 27, 1335–1349. [Google Scholar] [CrossRef]
- Neale, P.J.; Melis, A. Algal photosynthetic membrane complex and the photosynthesis-irradiance curve: A comparison of light-adaptation responses in Chlamydomonas reinhardtii (Chlorophyta). J. Phycol. 1986, 22, 531–538. [Google Scholar] [CrossRef]
- Sukenik, A.; Bennett, J.; Falkowski, P.G. Light saturated photosynthesis-limitation by electron transport or carbon-fixation? Biochim. Biophys. Acta. 1987, 891, 205–215. [Google Scholar] [CrossRef]
- Tang, E.P.Y.; Vincent, W.F. Effects of daylength and temperature on the growth and photosynthesis of an Arctic cyanobacterium, Schizothrix calcicola (Oscillatoriaceae). Eur. J. Phycol. 2000, 35, 263–272. [Google Scholar] [CrossRef]
- Rae, R.; Vincent, W.F. Phytoplankton production in subarctic lake and river ecosystems: Development of a photosynthesis-temperature-irradiance model. J. Plankton Res. 1998, 20, 1293–1312. [Google Scholar] [CrossRef]
- Young, A.J. Factors that affect the carotenoids composition of higher plants and algae. In Carotenoids in Photosynthesis; Young, A.J., Britton, G., Eds.; Chapman and Hall: London, UK, 1993; pp. 160–205. [Google Scholar]
- Young, A.J.; Frank, H.A. Energy transfer reactions involving carotenoids: Quenching of chlorophyll fluorescence. J. Photochem. Photobiol. B 1996, 36, 3–15. [Google Scholar] [CrossRef]
- Paerl, H.W.; Tucker, J.; Bland, P.T. Carotenoid enhancement and its role in maintaining blue-green algal (Microcystis aeruginosa) surface blooms. Limnol. Oceanogr. 1983, 28, 847–857. [Google Scholar]
- Paerl, H.W. Cyanobacterial carotenoids: Their roles in maintaining optimal photosynthetic production among aquatic bloom-forming genera. Oecologia 1984, 61, 143–149. [Google Scholar] [CrossRef]
- Jones, L.W.; Myers, J. Pigment variations in Anacystis nidulans induced by light of selected wavelengths. J. Phycol. 1965, 1, 7–15. [Google Scholar] [CrossRef]
- Glover, H.E.; Keller, M.D.; Spinrad, R.W. The effects of light quality on photosynthesis and growth of marine eukaryotic and prokaryotic phytoplankton clones. J. Exp. Mar. Biol. Ecol. 1987, 105, 137–159. [Google Scholar] [CrossRef]
- Tilzer, M.M. Light-dependence of photosynthesis and growth in cyanobacteria: Implications for their dominance in eutrophic lakes. New Zeal. J. Mar. Fresh. Res. 1987, 21, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Fahnenstiel, G.L.; Carrick, H.J.; Rogers, C.E.; Sicko-Goad, L. Red fluorescing phototrophic picoplankton in the Laurentian Great Lakes: What are they and what are they doing? Int. Rev. Ges. Hydrobiol. 1991, 76, 603–616. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Cohen-Bazire, G.; Kunisawa, R.; Mandel, M. Purification and properties of unicellular blue-green algae (order Chroococcales). Bact. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef] [Green Version]
- Guillard, R.R.; Sieracki, M.S. Counting cells in cultures with the light microscope. In Algal Culturing Techniques; Andersen, R.R., Ed.; Elsevier Academic Press: Burlington, VT, USA, 2005; pp. 239–252. [Google Scholar]
- Llewellyn, C.A.; Mantoura, R.F.C. The rapid determination of algal chlorophyll and carotenoid pigments and their breakdown products in natural waters by reverse-phase high-performance liquid chromatography. Anal. Chem. Acta. 1983, 151, 297–314. [Google Scholar]
- Jodłowska, S.; Latała, A. Simultaneous separation of chlorophylls and carotenoids by RP-HPLC in some algae and cyanobacteria from the Southern Baltic. Oceanol. Hydrobiol. Stud. 2003, 32, 81–89. [Google Scholar]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in freshwater blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.A.; Guglielmi, G.; Tandeau de Marsac, N.; Casteta, A.-M.; Cohen-Bazire, G. The structure of cyanobacterial phycobilisomes: A model. Arch. Microbiol. 1979, 123, 113–127. [Google Scholar] [CrossRef]
- Fisher, R.A.; Yates, F. Statistical Tables for Biological, Agricultural and Medical Research, 6th ed.; Olivier and Boyd: Edinburgh, UK, 1963; p. 146. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Śliwińska-Wilczewska, S.; Konarzewska, Z.; Wiśniewska, K.; Konik, M. Photosynthetic Pigments Changes of Three Phenotypes of Picocyanobacteria Synechococcus sp. under Different Light and Temperature Conditions. Cells 2020, 9, 2030. https://doi.org/10.3390/cells9092030
Śliwińska-Wilczewska S, Konarzewska Z, Wiśniewska K, Konik M. Photosynthetic Pigments Changes of Three Phenotypes of Picocyanobacteria Synechococcus sp. under Different Light and Temperature Conditions. Cells. 2020; 9(9):2030. https://doi.org/10.3390/cells9092030
Chicago/Turabian StyleŚliwińska-Wilczewska, Sylwia, Zofia Konarzewska, Kinga Wiśniewska, and Marta Konik. 2020. "Photosynthetic Pigments Changes of Three Phenotypes of Picocyanobacteria Synechococcus sp. under Different Light and Temperature Conditions" Cells 9, no. 9: 2030. https://doi.org/10.3390/cells9092030