Desialylation of Sonic-Hedgehog by Neu2 Inhibits Its Association with Patched1 Reducing Stemness-Like Properties in Pancreatic Cancer Sphere-forming Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Flow Cytometry
2.4. Real-Time PCR analysis
2.5. Immunoblotting
2.6. Glycoprotein Analysis
2.7. Detection of Linkage-Specific Sialic Acids
2.8. Transfection
2.9. Sphere Formation Assays
2.10. Confocal Microscopy
2.11. Cell Viability Assay
2.12. Patient Samples
2.13. Immunohistochemistry
2.14. Co-Immunoprecipitation
2.15. In Vivo Tumorigenicity
2.16. Statistical Analysis
3. Results
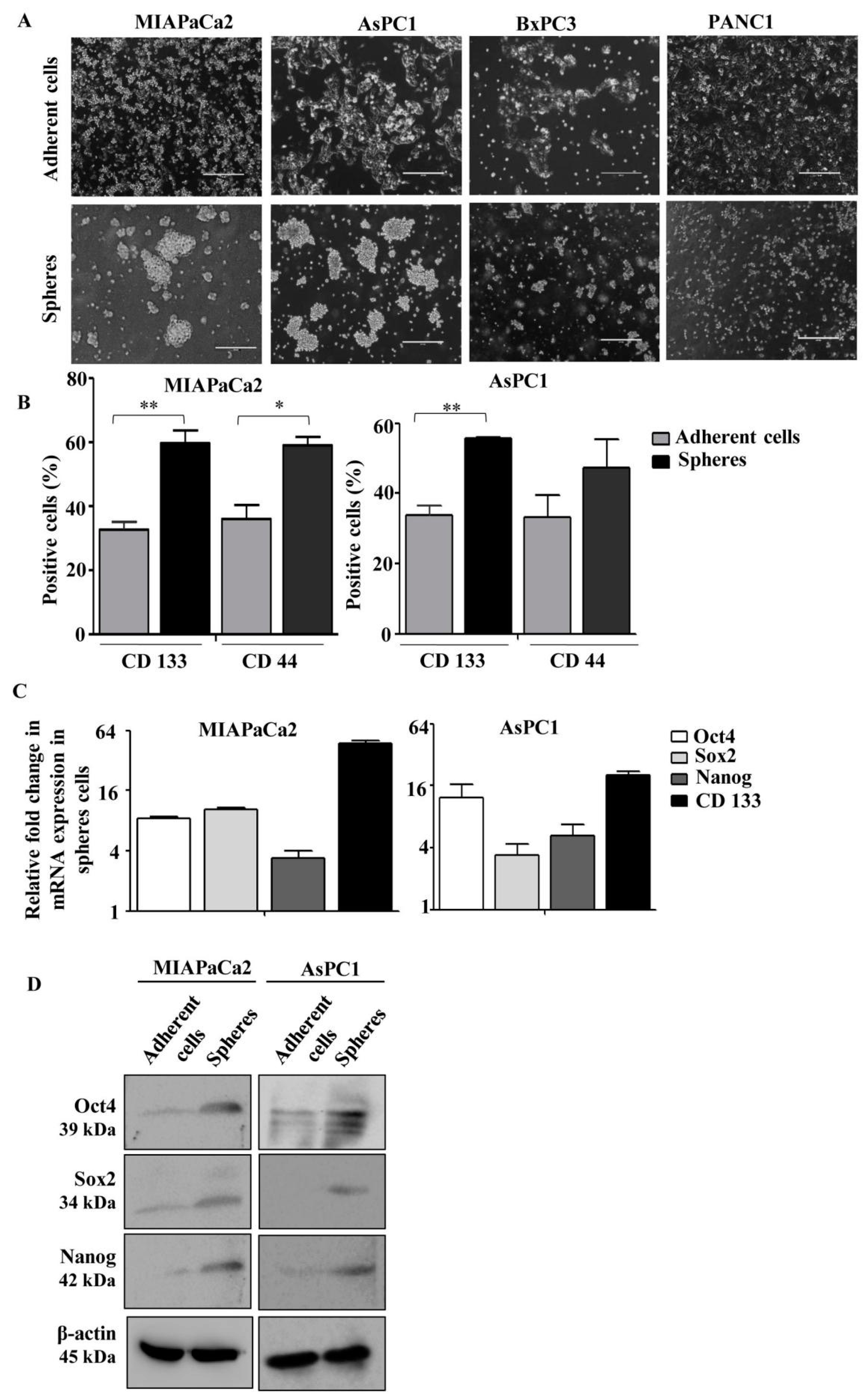
3.1. Generation and Characterization of Pancreatic Cancer Sphere-Forming Cells (PCS) from an Array of Pancreatic Cancer Cell Lines
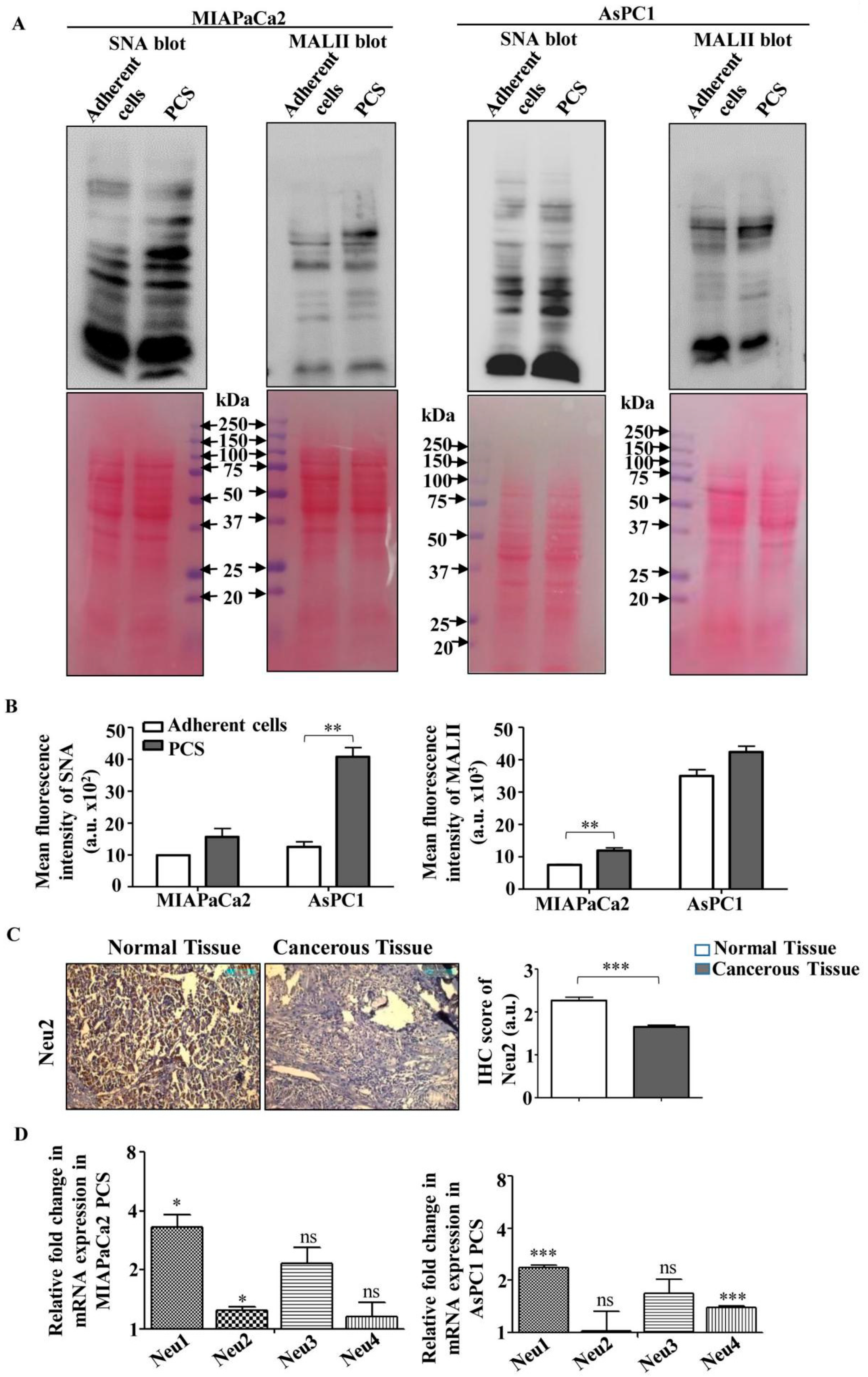
3.2. Higher Sialylation and Lower Sialidase (Neu2) Expression in PCS
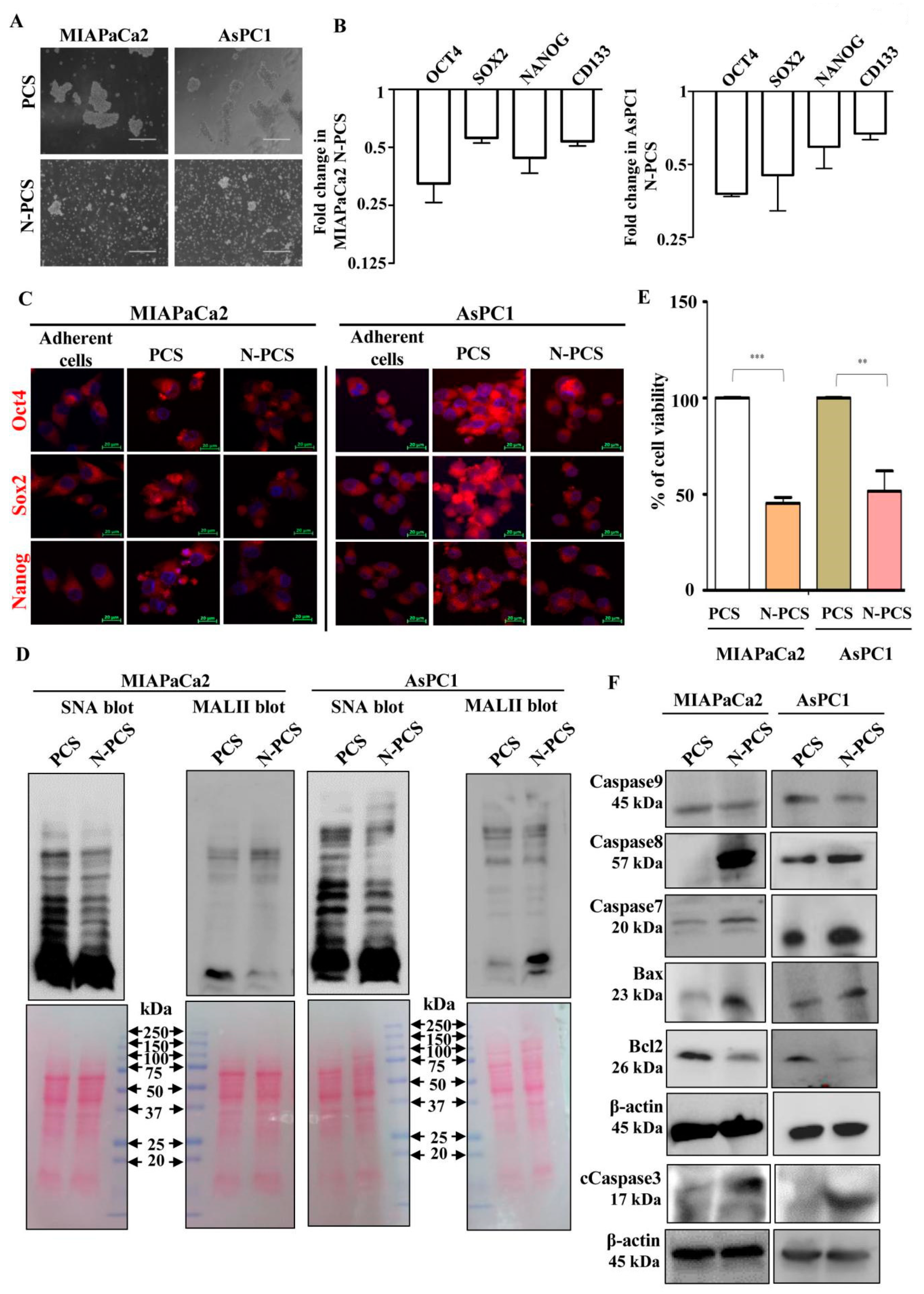
3.3. Neu2 Overexpression Reduces Stemness-like Properties in PCS
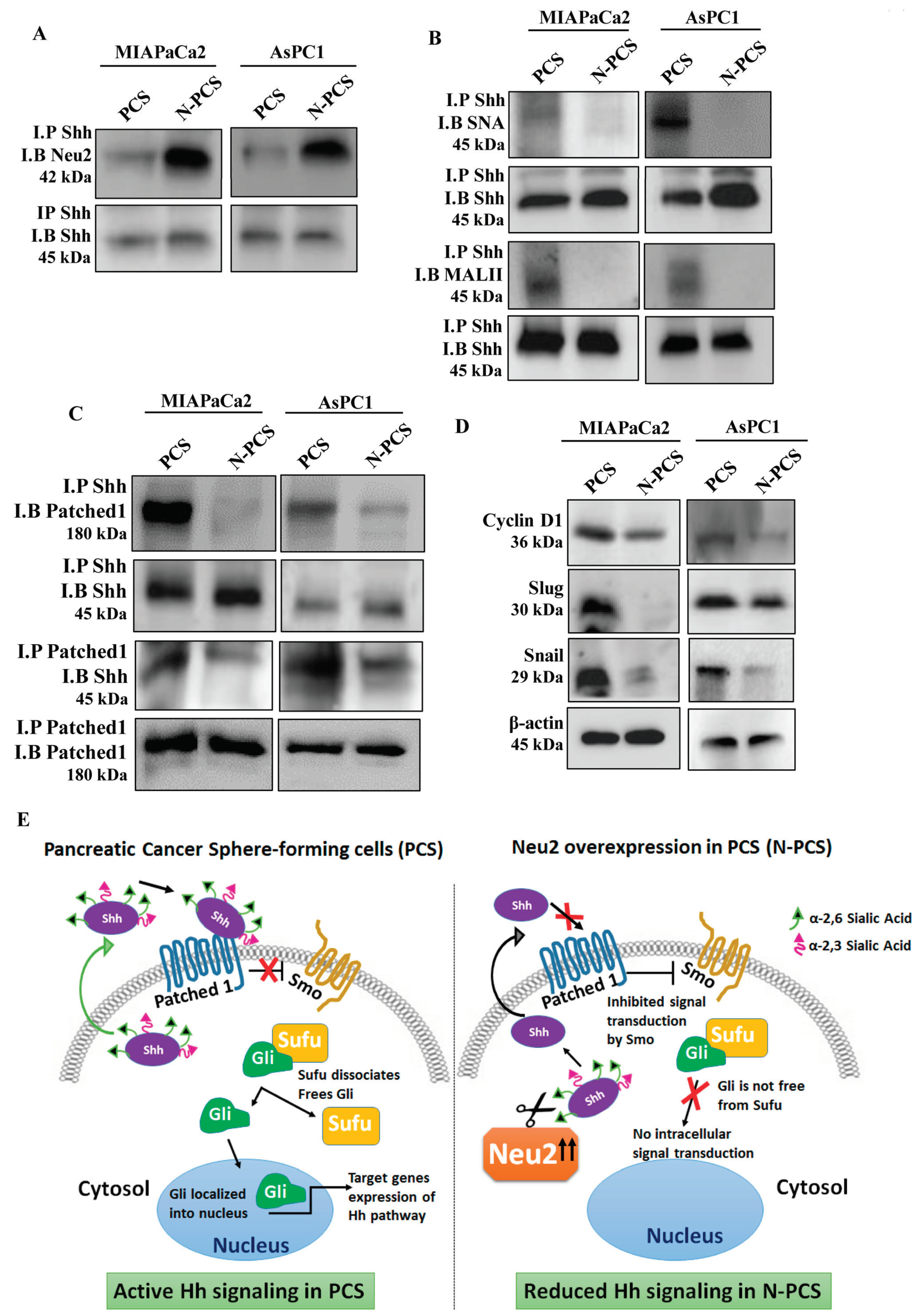
3.4. Reduction of Sialylation and Activation of Apoptosis in N-PCS
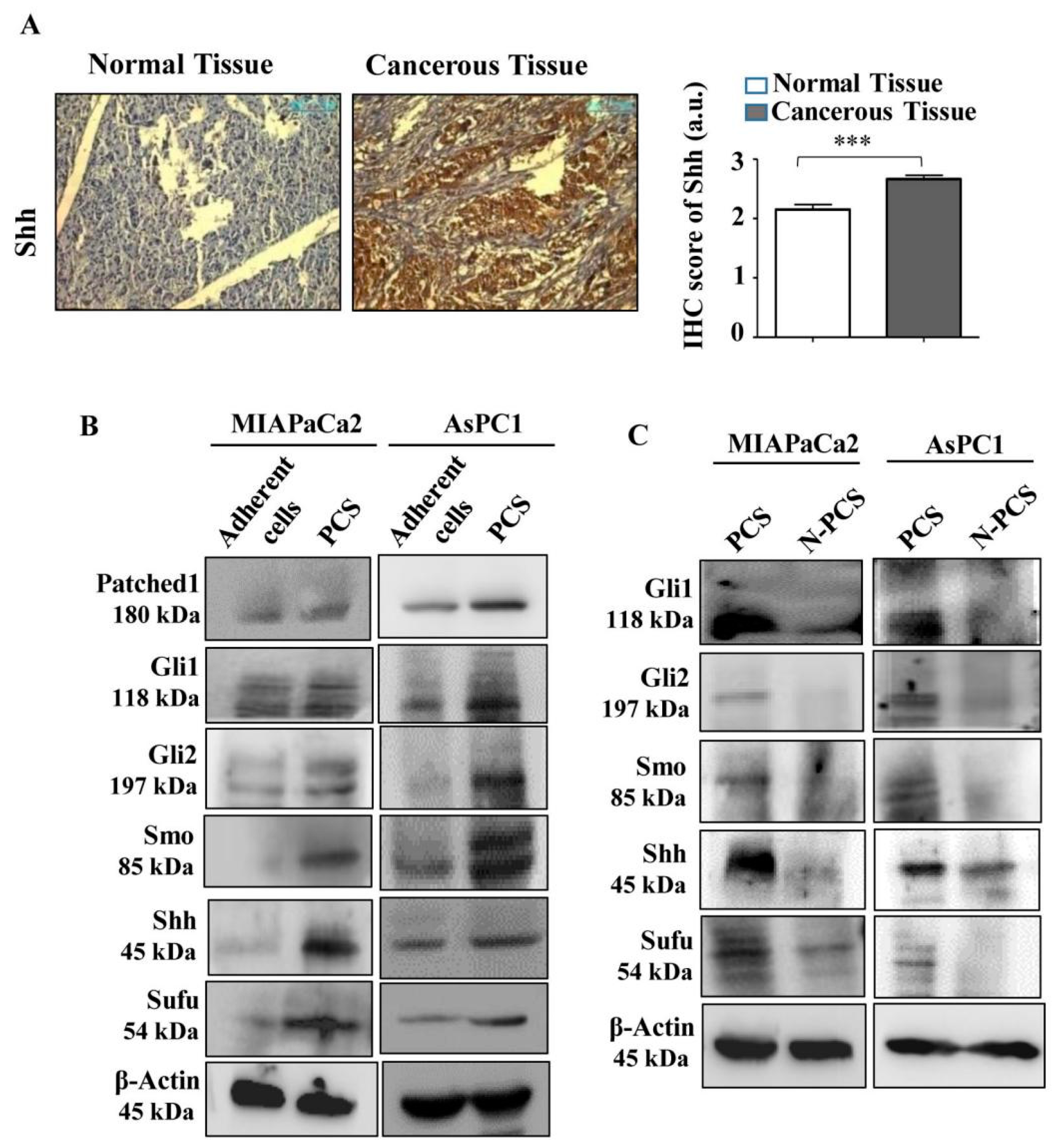
3.5. Neu2 Overexpression in PCS Diminishes Hedgehog Pathway (Hh) Activity
3.6. Association of Neu2 with Sonic Hedgehog (Shh) Causes Its Desialylation and Deactivation
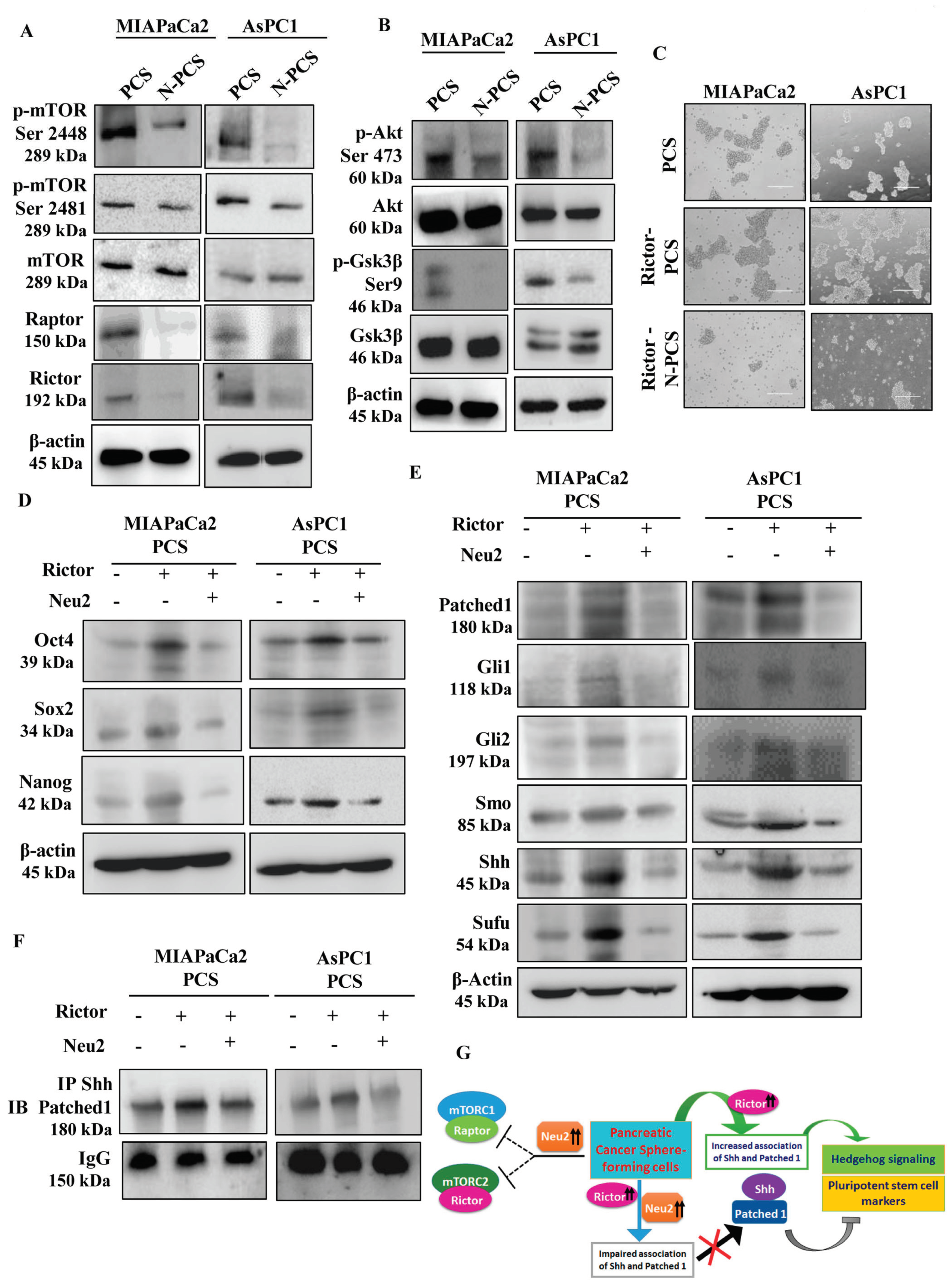
3.7. Neu2 Modulates Hedgehog Pathway (Hh) Activity by Downregulating mTORC2 Formation
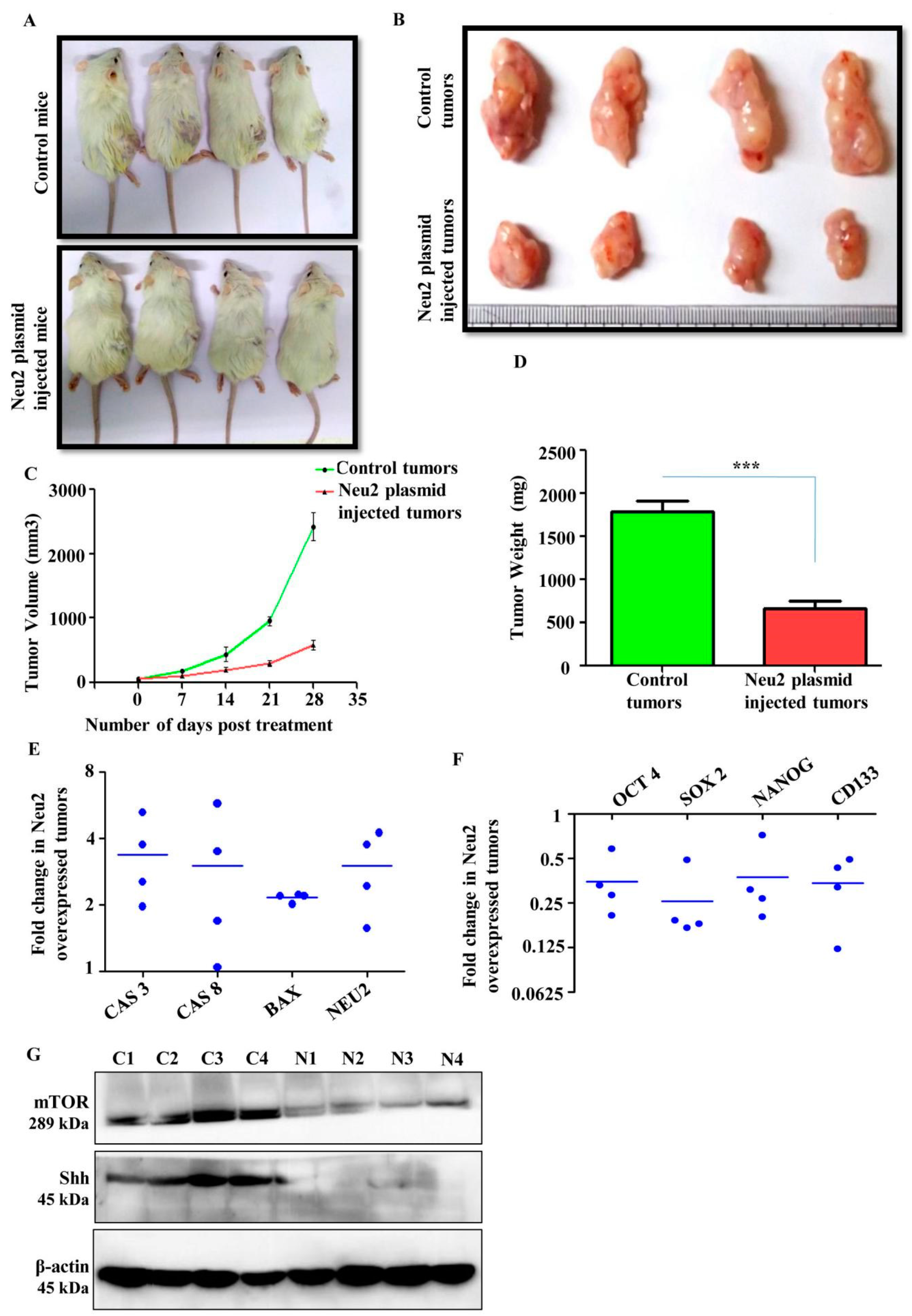
3.8. Neu2 Overexpression Reduced Tumor Growth in Xenograft Model
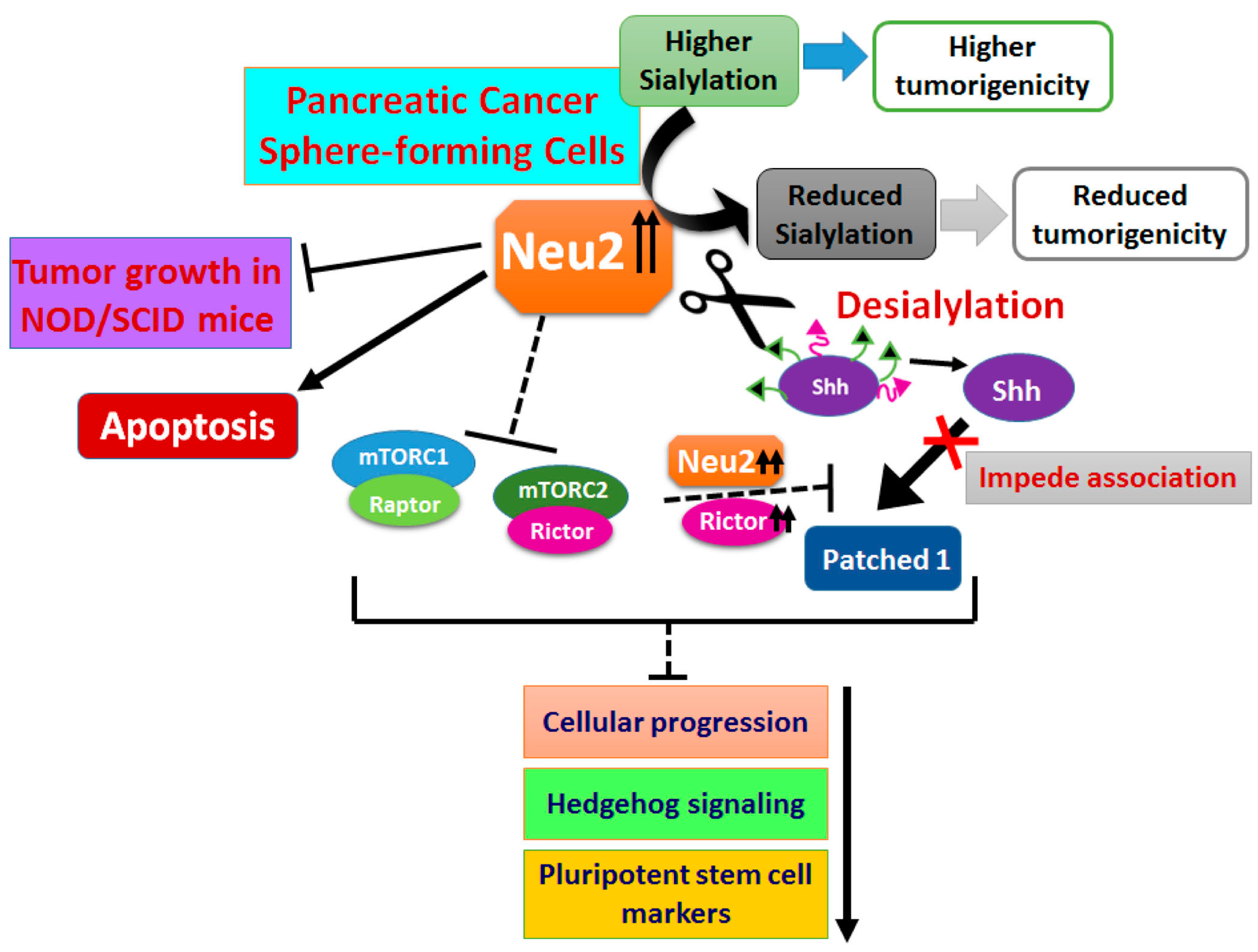
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aponte, P.M.; Caicedo, A. Stemness in Cancer: Stem Cells, Cancer Stem Cells, and Their Microenvironment. Stem Cells Int. 2017, 2017, 5619472. [Google Scholar] [CrossRef] [PubMed]
- Lathia, J.D.; Liu, H. Overview of Cancer Stem Cells and Stemness for Community Oncologists. Target Oncol. 2017, 12, 387–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Bhattacharya, K.; Mandal, C. Nutritional stress reprograms dedifferention in glioblastoma multiforme driven by PTEN/Wnt/Hedgehog axis: A stochastic model of cancer stem cells. Cell Death Discov. 2018, 4, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carballo, G.B.; Honorato, J.R.; de Lopes, G.P.F.; Spohr, T.C.L.S.E. A highlight on Sonic hedgehog pathway. Cell Commun. Signal. 2018, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.N.; Fu, J.; Nall, D.; Rodova, M.; Shankar, S.; Srivastava, R.K. Inhibition of sonic hedgehog pathway and pluripotency maintaining factors regulate human pancreatic cancer stem cell characteristics. Int. J. Cancer 2012, 131, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, Y.; Matsubara, S.; Ding, Q.; Tsukasa, K.; Yoshimitsu, M.; Kosai, K.; Takao, S. Efficient elimination of pancreatic cancer stem cells by hedgehog/GLI inhibitor GANT61 in combination with mTOR inhibition. Mol. Cancer 2016, 15, 49. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Du, Y.; Yang, Z.; He, L.; Wang, Y.; Hao, L.; Ding, M.; Yan, R.; Wang, J.; Fan, Z. GALNT1-Mediated Glycosylation and Activation of Sonic Hedgehog Signalling Maintains the Self-Renewal and Tumor-Initiating Capacity of Bladder Cancer Stem Cells. Cancer Res. 2016, 76, 1273–1283. [Google Scholar] [CrossRef] [Green Version]
- Zuo, M.; Rashid, A.; Churi, C.; Vauthey, J.N.; Chang, P.; Li, Y.; Hung, M.C.; Li, D.; Javle, M. Novel therapeutic strategy targeting the Hedgehog signalling and mTOR pathways in biliary tract cancer. Br. J. Cancer 2015, 112, 1042–1051. [Google Scholar] [CrossRef] [Green Version]
- Maiti, S.; Mondal, S.; Satyavarapu, E.M.; Mandal, C. mTORC2 regulates hedgehog pathway activity by promoting stability to Gli2 protein and its nuclear translocation. Cell Death Dis. 2017, 8, e2926. [Google Scholar] [CrossRef] [Green Version]
- Mandal, C.; Chatterjee, M.; Sinha, D. Investigation of 9-O-acetylated sialoglycoconjugates in childhood acute lymphoblastic leukaemia. Br. J. Haematol. 2000, 110, 801–812. [Google Scholar] [CrossRef]
- Mandal, C.; Mandal, C.; Chandra, S.; Schauer, R.; Mandal, C. Regulation of O-acetylation of sialic acids by sialate-O-acetyltransferase and sialate-O-acetylesterase activities in childhood acute lymphoblastic leukemia. Glycobiology 2012, 22, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, E.; Bonten, E.; D’Azzo, A.; Bresciani, R.; Venerando, B.; Borsani, G.; Schauer, R.; Tettamanti, G. Sialidases in Vertebrates: A Family Of Enzymes Tailored For Several Cell Functions*. Adv. Carbohyd. Chem. Biochem. 2010, 64, 403–479. [Google Scholar]
- Mandal, C.; Srinivasan, G.V.; Chowdhury, S.; Chandra, S.; Mandal, C.; Schauer, R.; Mandal, C. High level of sialate-O-acetyltransferase activity in lymphoblasts of childhood acute lymphoblastic leukaemia (ALL): Enzyme characterization and correlation with disease status. Glycoconj J. 2009, 26, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wei, A.; Zhang, H.; Chen, X.; Wang, L.; Zhang, H.; Yu, X.; Yuan, Q.; Zhang, J.; Wang, S. α2,6-Sialylation mediates hepatocellular carcinoma growth in vitro and in vivo by targeting the Wnt/β-catenin pathway. Oncogenesis 2017, 6, e343. [Google Scholar] [CrossRef]
- Miyagi, T.; Yamaguchi, K. Mammalian sialidases: Physiological and pathological roles in cellular functions. Glycobiology 2012, 22, 880–896. [Google Scholar] [CrossRef] [Green Version]
- Tringali, C.; Lupo, B.; Anastasia, L.; Papini, N.; Monti, E.; Bresciani, R.; Tettamanti, G.; Venerando, B. Expression of sialidase Neu2 in leukemic K562 cells induces apoptosis by impairing Bcr-Abl/Src kinases signalling. J Biol Chem. 2007, 282, 14364–14372. [Google Scholar] [CrossRef] [Green Version]
- Sawada, M.; Moriya, S.; Saito, S.; Shineha, R.; Satomi, S.; Yamori, T.; Tsuruo, T.; Kannagi, R.; Miyagi, T. Reduced sialidase expression in highly metastatic variants of mouse colon adenocarcinoma 26 and retardation of their metastatic ability by sialidase overexpression. Int. J. Cancer 2002, 97, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Nath, S.; Mandal, C.; Chatterjee, U.; Mandal, C. Association of cytosolic sialidase Neu2 with plasma membrane enhances Fas-mediated apoptosis by impairing PI3K-Akt/mTOR-mediated pathway in pancreatic cancer cells. Cell Death Dis. 2018, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Dutta, D.; Samanta, S.K.; Bhattacharya, K.; Pal, B.C.; Li, J.; Datta, K.; Mandal, C.; Mandal, C. Oxidative inhibition of Hsp90 disrupts the super-chaperone complex and attenuates pancreatic adenocarcinoma in vitro and in vivo. Int. J. Cancer 2013, 132, 695–706. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Mandal, C.; Sangwan, R.; Mandal, C. Coupling G2/M arrest to the Wnt/β-catenin pathway restrains pancreatic adenocarcinoma. Endocr. Relat. Cancer 2014, 21, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Mandal, C.; Sarkar, S.; Chatterjee, U.; Schwartz-Albiez, R.; Mandal, C. Disialoganglioside GD3-synthase over expression inhibits survival and angiogenesis of pancreatic cancer cells through cell cycle arrest atS-phase and disruption of integrin-β1-mediated anchorage. Int. J. Biochem. Cell Bio. 2014, 53, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Mandal, C.; Tringali, C.; Mondal, S.; Anastasia, L.; Chandra, S.; Venerando, B.; Mandal, C. Down regulation of membrane-bound Neu3 constitutes a new potential marker for childhood acute lymphoblastic leukemia and induces apoptosis suppression of neoplastic cells. Int J Cancer 2010, 126, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, J.; Roy, S.; Mandal, C. Modulation of TLR4 Sialylation Mediated by a Sialidase Neu1 and Impairment of Its Signaling in Leishmania donovani Infected Macrophages. Front Immunol. 2019, 10, 2360. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, K.; Maiti, S.; Mandal, C. PTEN negatively regulates mTORC2 formation and signalling in grade IV glioma via Rictor hyperphosphorylation at Thr1135 and direct the mode of action of an mTORC1/2 inhibitor. Oncogenesis 2016, 5, e227. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, S.; Sato, I.; Wada, T.; Yamaguchi, K.; Li, Y.; Li, D.; Zhao, X.; Ueno, S.; Aoki, H.; Tochigi, T.; et al. Plasma membrane-associated sialidase (NEU3) regulates progression of prostate cancer to androgen-independent growth through modulation of androgen receptor signalling. Cell Death Differ. 2012, 19, 170–179. [Google Scholar] [CrossRef]
- Li, S.; Zhang, X.; Xia, X. Regression of Tumor Growth and Induction of Long-Term Antitumor Memory by Interleukin 12 Electro-Gene Therapy. J. Natl. Cancer Inst. 2002, 94, 10. [Google Scholar] [CrossRef]
- Hattori, Y. Delivery of Plasmid DNA into Tumors by Intravenous Injection of PEGylated Cationic Lipoplexes into Tumor-Bearing Mice. Pharmacol. Pharm. 2016, 7, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, S.; Ding, Q.; Miyazaki, Y.; Kuwahata, T.; Tsukasa, K.; Takao, S. mTOR plays critical roles in pancreatic cancer stem cells through specific and stemness-related functions. Sci. Rep. 2013, 3, 3230. [Google Scholar] [CrossRef] [Green Version]
- Gkountakos, A.; Pilotto, S.; Mafficini, A.; Vicentini, C.; Simbolo, M.; Milella, M.; Tortora, G.; Scarpa, A.; Bria, E.; Corbo, V. Unmasking the impact of Rictor in cancer: Novel insights of mTORC2 complex. Carcinogenesis 2018, 39, 971–980. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.C.; Shyr, Y.M.; Liao, W.Y.; Chen, T.H.; Wang, S.E.; Lu, P.C.; Lin, P.Y.; Chen, Y.B.; Mao, W.Y.; Han, H.Y.; et al. Elevation of β-galactoside α2,6-sialyltransferase 1 in a fructose responsive manner promotes pancreatic cancer metastasis. Oncotarget 2017, 8, 7691–7709. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Ghosh, S.; Bandyopadhyay, S.; Mandal, C.; Bandhyopadhyay, S.; Bhattacharya, K.D.; Mandal, C. Differential expression of 9-O-acetylated sialoglycoconjugates on leukemic blasts: A potential tool for long-term monitoring of children with acute lymphoblastic leukemia. Int. J. Cancer 2004, 111, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Razi, E.; Radak, M.; Mahjoubin-Tehran, M.; Talebi, S.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; Sharifi, H.; Mousavi, N.; Sarvizadeh, M.; et al. Cancer stem cells as therapeutic targets of pancreatic cancer. Fundam. Clin. Pharmacol. 2020, 34, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Barkeer, S.; Chugh, S.; Batra, S.K.; Ponnusamy, M.P. Glycosylation of Cancer Stem Cells: Function in Stemness, Tumorigenesis, and Metastasis. Neoplasia 2018, 20, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.J.; Holdbrooks, A.T.; Chakraborty, A.; Grizzle, W.E.; Landen, C.N.; Buchsbaum, D.J.; Conner, M.G.; Arend, R.C.; Yoon, K.J.; Klug, C.A. The Tumor-Associated Glycosyltransferase ST6Gal-I Regulates Stem Cell Transcription Factors and Confers a Cancer Stem Cell Phenotype. Cancer Res. 2016, 76, 3978–3988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.C.; Stein, J.W.; Lynch, C.L.; Tran, H.T.; Lee, C.Y.; Coleman, R.; Hatch, A.; Antontsev, V.G.; Chy, H.S.; O’Brien, C.M. Glycosyltransferase ST6GAL1 contributes to the regulation of pluripotency in human pluripotent stem cells. Sci. Rep. 2015, 5, 13317. [Google Scholar] [CrossRef] [Green Version]
- Fanzani, A.; Colombo, F.; Giuliani, R.; Preti, A. Marchesini S. Cytosolic sialidase Neu2 upregulation during PC12 cells differentiation. FEBS Lett. 2004, 566, 178–182. [Google Scholar] [CrossRef]
- Wang, F.; Ma, L.; Zhang, Z.; Liu, X.; Gao, H.; Zhuang, Y.; Yang, P.; Kornmann, M.; Tian, X.; Yang, Y. Hedgehog Signalling Regulates Epithelial-Mesenchymal Transition in Pancreatic Cancer Stem-Like Cells. J Cancer. 2016, 7, 408–417. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nath, S.; Mondal, S.; Butti, R.; Prasanna Gunasekaran, V.; Chatterjee, U.; Halder, A.; Kundu, G.C.; Mandal, C. Desialylation of Sonic-Hedgehog by Neu2 Inhibits Its Association with Patched1 Reducing Stemness-Like Properties in Pancreatic Cancer Sphere-forming Cells. Cells 2020, 9, 1512. https://doi.org/10.3390/cells9061512
Nath S, Mondal S, Butti R, Prasanna Gunasekaran V, Chatterjee U, Halder A, Kundu GC, Mandal C. Desialylation of Sonic-Hedgehog by Neu2 Inhibits Its Association with Patched1 Reducing Stemness-Like Properties in Pancreatic Cancer Sphere-forming Cells. Cells. 2020; 9(6):1512. https://doi.org/10.3390/cells9061512
Chicago/Turabian StyleNath, Shalini, Susmita Mondal, Ramesh Butti, Vinoth Prasanna Gunasekaran, Uttara Chatterjee, Aniket Halder, Gopal C. Kundu, and Chitra Mandal. 2020. "Desialylation of Sonic-Hedgehog by Neu2 Inhibits Its Association with Patched1 Reducing Stemness-Like Properties in Pancreatic Cancer Sphere-forming Cells" Cells 9, no. 6: 1512. https://doi.org/10.3390/cells9061512