Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine
by
 , , , and
, , , and
Karolina Wydra
1, 
Dawid Gawliński
1,
Kinga Gawlińska
1,
Małgorzata Frankowska
1,
Dasiel O. Borroto-Escuela
2,3,4,
Kjell Fuxe
2 and
Małgorzata Filip
1,* 1
Department of Drug Addiction Pharmacology, Maj Institute of Pharmacology Polish Academy of Sciences, 31-343 Kraków, Poland
2
Department of Neuroscience, Karolinska Institutet, 171 77 Stockholm, Sweden
3
Section of Physiology, Department of Biomolecular Science, University of Urbino, Campus Scientifico Enrico Mattei, Via Ca’ Le Suore 2, 61029 Urbino, Italy
4
Observatorio Cubano de Neurociencias, Grupo Bohío-Estudio, Zayas 50, Yaguajay 62100, Cuba
*
Author to whom correspondence should be addressed.
Cells 2020, 9(6), 1372; https://doi.org/10.3390/cells9061372
Submission received: 5 May 2020
/
Revised: 28 May 2020
/
Accepted: 28 May 2020
/
Published: 1 June 2020
(This article belongs to the Special Issue Adenosine Receptors: From Cell Biology to Human Diseases)
{kind=link}
{kind=link}
Abstract
:Several psychoactive drugs can evoke substance use disorders (SUD) in humans and animals, and these include psychostimulants, opioids, cannabinoids (CB), nicotine, and alcohol. The etiology, mechanistic processes, and the therapeutic options to deal with SUD are not well understood. The common feature of all abused drugs is that they increase dopamine (DA) neurotransmission within the mesocorticolimbic circuitry of the brain followed by the activation of DA receptors. D2 receptors were proposed as important molecular targets for SUD. The findings showed that D2 receptors formed heteromeric complexes with other GPCRs, which forced the addiction research area in new directions. In this review, we updated the view on the brain D2 receptor complexes with adenosine (A)2A receptors (A2AR) and discussed the role of A2AR in different aspects of addiction phenotypes in laboratory animal procedures that permit the highly complex syndrome of human drug addiction. We presented the current knowledge on the neurochemical in vivo and ex vivo mechanisms related to cocaine use disorder (CUD) and discussed future research directions for A2AR heteromeric complexes in SUD.
1. Introduction
From the perspective of public health, substance use disorders (SUD; commonly called drug addictions) are unsolved issues [1]. Several drugs can lead to addictive behavior in humans and animals,and these include psychostimulants, opioids, cannabinoids (CB), nicotine, and alcohol. The etiology and mechanistic processes,as well as the therapeutic options to deal with SUD, are neithercomplete nor well-understood (e.g., [2]). Thus, researchers must search for the neurobiological bases of development to uncover efficient treatments for this disorder, which affects the global population. SUD is a chronic brain disorder, in which, after the initial behavioral spectrum of feeling (well-being, pleasure, and euphoria), compulsive drug-seeking and drug-taking behaviors appear despite the negative consequences, and relapses, accompanied by psychic, somatic, and vegetative disturbances, are triggered after drug abstinence [3]. These behavioral outcomes are realized through distinct effector mechanisms, including neurotransmitter transporters, ion channels, and receptor proteins.
It is well-established that drugs of abuse have common features that trigger addictive potential, i.e., they increase dopamine (DA) neurotransmission within the mesocorticolimbic circuitry of the brain from the ventral tegmental area, the nucleus accumbens, and the prefrontal cortex. Increased DA neurotransmission leads to indirect activation of five types of DA receptors, among which, D2 receptors have been the most widely studied as molecular targets for SUD. The findingsshowed that D2 receptors formed heteromeric complexes with other G protein-coupled receptors(GPCRs), while such heteromersdemonstrated pharmacology and functions distinct from the constituent receptors,which forced the addiction research to explore new directions.
Here, we update the view on the brain D2 receptor complexes with adenosine (A)2A receptors (A2AR), the role of A2AR in different aspects of addiction phenotypes in laboratory animal procedures that permit the highly complex syndrome of human drug addiction (drug-induced reward, discrimination, sensitization, seeking behavior, and withdrawal symptoms). At the end, we present the current knowledge on the neurochemical in vivo and the ex vivo mechanisms related to cocaine use disorder (CUD) and discuss future research directions for A2AR heteromeric complexes in SUD.
2. A2ARs and Their Localization in the Brain
The distribution of A2ARs in the mammalian brain on the level of transcript and protein was demonstrated ex vivo using reverse transcriptase-polymerase chain reaction (RT-PCR), Northern blotting, Western blotting, ELISA, in situ hybridization, immunohistochemistry, autoradiography, and radioligand binding, while position emission tomography (PET) studies were employed for the in vivo analyses.
Several studies indicated the differential expression of A2ARs in mammalian brains. Studies performed with techniques to detect mRNA encodingA2ARsdemonstrated, in rodents, the highest receptor transcript level in the striatal areas, while lower levelswere detected extrastriatally in the hippocampus, hypothalamus, thalamus, cerebral cortex, amygdala, thalamic nuclei, and the substantia nigra [4,5,6,7,8,9,10,11]. A similar pattern of A2AR mRNA expression was observed in the human brain with high levels in the nucleus caudatus, while much lower levels were found in the optical cortex, amygdala, hippocampus, substantia nigra, and the cerebellum [12]. The highest protein density was reported for the striatum (dorsal and ventral parts), olfactory tubercle, and globus pallidus in the mammalian brain [8,9,13,14,15].
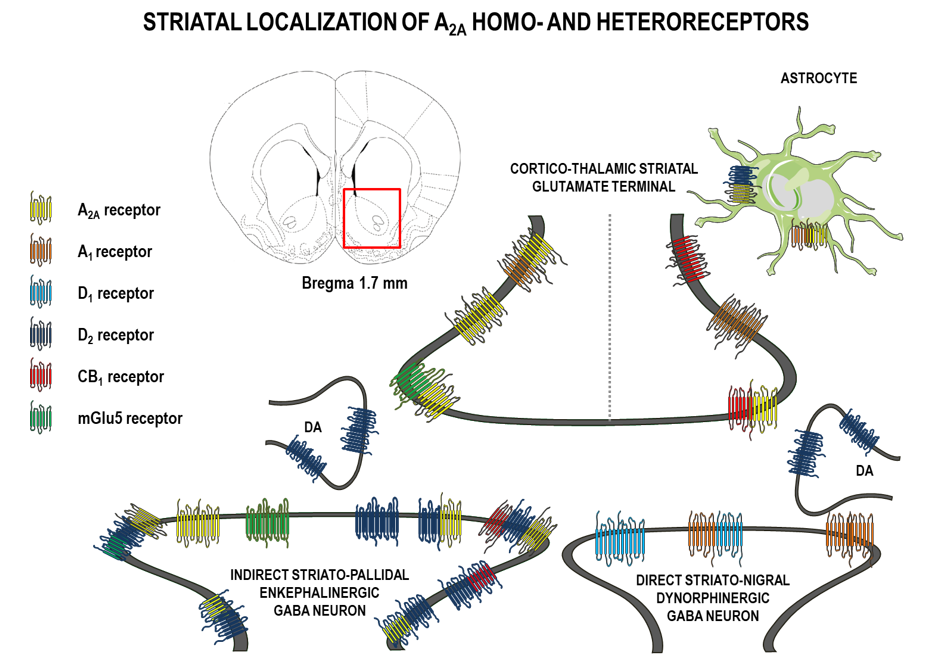
Using anatomical, electrophysiological, and biochemical approaches researchers demonstrated that A2AR in brainswerelocalized on neurons and glia cells (including astrocytes and microglia) [7,8,9,16,17]. On neurons, these receptors occurredpostsynaptically (on dendrites and dendritic spines), presynaptically (on terminals of axon collaterals), and extrasynaptically (on somas) [7,8,9,18,19] (Figure 1).
In the striatum, A2ARs were localized on neurons (ca. 90%) and about 3% were on astrocytes [9,20,21,22,23]. On striatal neurons, about 70% of A2ARs were located postsynaptically, 23% presynaptically, and 3% extrasynaptically [9,21].
Postsynaptically-expressed striatal A2ARs were mainly localized to the gamma-aminobutric acid (GABA)ergic medium-sized spiny neurons (MSN) of the indirect pathway projecting to the external segmentofthe globus pallidus. These latter neurons also expressed a high density of DA D2 receptors and enkephalin [7,9,10,13,14,24,25]. Inversely, neurons of the direct striato-nigral pathway (which selectively expressed DA D1 receptors and the peptide dynorphin) did not contain a significant level of A2ARs [13]. Postsynaptically localized A2ARs were found around dopaminergic synapses [21]. In the striatum, a smaller amount of A2ARs were localized presynaptically mainly on cortico-thalamic glutaminergic terminals that contacted medium-sized spiny neurons of the GABAergic direct and indirect pathways [9,19,26]. Striatal A2ARs were located presynaptically on cholinergic neurons that modulated acetylcholine release [27,28,29].
Using double immunofluorescence, co-immunoprecipitation, resonance energy transfer techniques (a sequential resonance energy transfer (SRET) and bimolecular fluorescence complementation plus bioluminescence response energy transfer (BRET)), and in vivo proximity ligation assay (PLA), research indicated that A2ARs occur in dimers:They form either homodimers (A2AR-A2AR) or heterodimers with other metabotropic GPCRs.
In the hippocampus,the following occurred:
In the striatum, the following existed:
- homodimeric A2AR-A2AR complexes on the cell surface, as shown using BRETassays [31];
A1-A2Aisoreceptor complexes were identified in striatal glutamate nerve terminals at the presynaptic level [36]. On striatal glutamate nerve terminals, there may also exist A2A-D2 heterodimers in balance with A1-A2A-D2 trimeric heteroreceptor complexes.
In the striatum in vivo and in vitro A2ARs and mGlu5 receptor complexes were detected inter alia extrasynaptically on glutamatergic terminals and GABA dendrites, where they play a role in local plasticity and in modulation of the activity of GABA striato–pallidal neurons [26,37]. Activation of A2ARs and mGlu5 receptors resulted in decreased affinity of D2 for DA [38], increased striatal c-Fos expression [37] and cyclic adenosine monophosphate (cAMP) formation, and striatal DA- and cAMP-regulated neuronal phosphoprotein DARP-32 phosphorylation [39]. In addition, presynaptic interactions between A2ARs and mGlu5 receptors on cortico–striatal glutamatergic nerveterminals may also contribute to the described interaction by synergistic regulation of glutamate release [26].
In the striatum, A2ARs and cannabinoid CB1 receptors may also form heteromeric complexes and, in this way, A2AR activation facilitated CB1 receptor signaling [40,41]. There is also evidence for A2A-CB1-D2 and A2A-D2-mGlu5 receptor heteromers in transfected cells [42,43] and such interactions appeared at both pre- and postsynaptic levels to control the neurotransmission and signaling in different brain structures. The existence of A2A-CB1-D2 receptor mosaics was found in the terminal parts of the dorsal striato–pallidal GABA neurons, in the dendritic regions of the ventral striato–pallidal GABA neurons, and in the cortico–striatal glutamate terminals [42,44,45,46]. The A2AR-mGlu5 receptor complexes existedinter alia on the glutamate terminals, forming synapses on the striato–pallidal GABA neurons [26,36,47].
Flajolet, Greengard, and colleagues discovered FGFR1and A2AR complexes [48,49] as well as FGFR1-A2A-D2 heterocomplexes in the striatum [50]. In the FGFR1-A2A-D2heterocomplex, the adenosine-induced activation of A2AR may then enhance signaling over FGFR1 via an allosteric receptor–receptor interaction, increasing the structural plasticity and repair.
A2ARs can heteromerize also with D2, D3, and D4 receptors [34,51,52]. The D3 receptor in the brain is mainly found in the ventral striatum, unlike the D2 receptor, which is located in high densities in both the ventral and dorsal striatum. Rivera et al. demonstrated that the D4 receptor was enriched in the striosomes and the matrix of the dorsal striatum [53]. Borroto-Escuela et al. demonstrated A2A-D4 heteroreceptor complexes in the dorsal striatum, especially in the striosomes [34] using an in situ PLA assay [33].
- The first recognized and best known heteromeric interaction was found between the A2ARs and D2 receptors, by which the activation of A2A (Gs/olf-coupled) receptors interfered with coupling of D2 receptors to Gi isoforms of G proteins [54,55,56]. A2A-D2 heteroreceptor complexes with antagonistic receptor–receptor interaction were found to exist on:
- the cortico–striatal glutamate nerve terminals, where the A2AR protomer inhibited the D2 receptor protomer-induced inhibition of glutamate release [47];
- striatal cholinergic interneurons [57];and
The striatal A2AR-D2 receptor heterocomplexes were first shown by Trifilieff et al. [60] using anin situ PLA assay and, later, by Borroto-Escuela and his group [61].
Based on a mathematical and bioinformatic approach,Tarakanov and Fuxededuced a set of triplet homologies (triplet puzzle) contributing to the formation of the receptor interface [62]. Using thein vivo PLA assay technique,theydemonstrated a high expression of A2A-D2-Sigma1 heterocomplexes was indicated to exist in the ventral striatum [60,61,63].
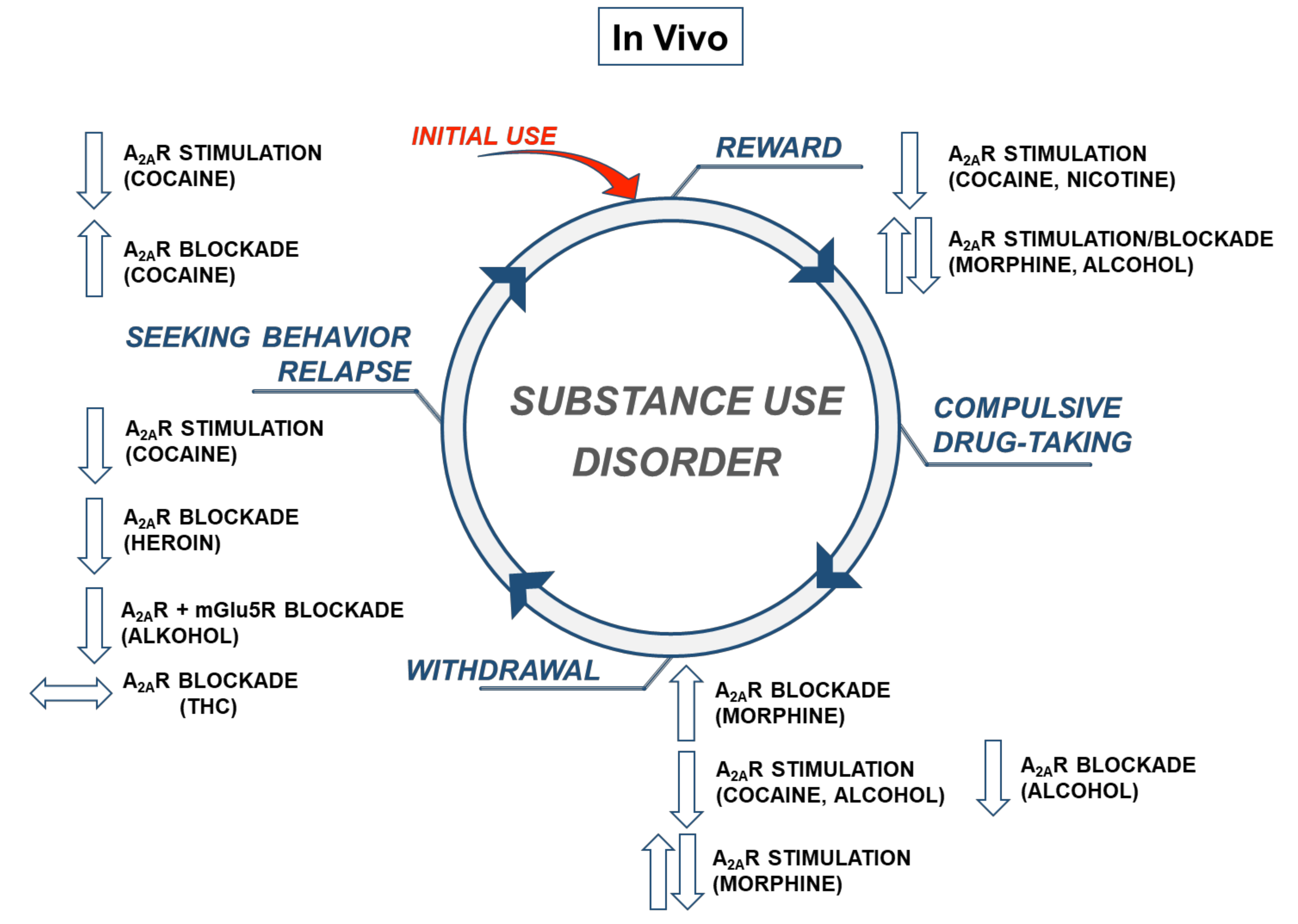
3. A2AR and Behavioral Actions to Cocaine and Other Drugs of Abuse—In Vivo Evidence
Numerous preclinical studies confirmed functional interactions between A2ARs and drugs of abuse with different mechanisms of action on the central nervous system. Using classical pharmacological tools (agonists or antagonists of A2ARs), as well as animal genetic models (overexpression or knockout (KO) of A2ARs) studies demonstrated an involvement of these receptors in the locomotor response, drug discrimination, reward-seeking behavior, and withdrawal syndrome associated with the use of psychostimulants (cocaine, amphetamine, methamphetamine, and 3,4-methylenedioxymethamphetamine (MDMA)), nicotine, opioids (morphine and heroin), cannabinoids (∆9-tetrahydrocannabinol (THC)), and ethanol.
3.1. Locomotor Responses to Drugs of Abuse
Several studies indicated a role of A2ARs in the locomotor action of drugs of abuse. Thus, studies using A2AR agonists (2-[(2-aminoethylamino)carbonylethylphenylethylamino]-5′-N-ethylcarboxamidoadenosine (APEC) and (4-[2-[[6-Amino-9-(N-ethyl-b-d-ribofuranuronamidosyl)-9H-purin-2-yl]amino]ethyl]benzene-propanoic acid hydrochloride (CGS 21680)) consistently indicated the inhibitory effect of activation of these receptors in psychostimulant-induced locomotor hyperactivity. The administration of APEC (0.01 mg/kg) [64] or CGS 21680 (0.1–2.0 mg/kg) [65] decreased locomotor response to acute amphetamine treatment, while CGS 21680 (0.03–3.0 mg/kg) attenuated such behavior of methamphetamine in rats [65,66]. In non-habituated mice, CGS 21680 (0.01–0.5 mg/kg) inhibited both amphetamine- and cocaine-induced hyperlocomotion; however, in the case of amphetamine, this agonist was effective only the highest used dose [67]. Locomotor hyperactivation induced by acute cocaine administration was decreased by CGS 21680 (0.2 mg/kg) also in well-habituated rats [68]. Supporting the pharmacological studies, A2AR overexpressing rats—as opposed to wild-type controls—did not induce an increase in locomotion after a single cocaine injection [69].
In contrast to the stimulation of A2ARs, pharmacological antagonism enhanced psychostimulant-induced hyperlocomotion. The A2AR antagonists 3,7-dimethyl-1-propar-gylxanthine (DMPX; 3.0–6.0 mg/kg) [70] and (3-(3-hydroxypropyl)-8-(m-methoxystyryl)-7-methyl-1-propargylxanthine phosphate disodium salt (MSX-3; 5.0–25.0 mg/kg) [68] increased the locomotor hyperactivation after acute cocaine or amphetamine administration, respectively. In a recent study, Haynes et al. [71] used presynaptic 2-(2-furanyl)-7-[3-(4-methoxyphenyl)propyl]-7H-pyrazolo[4,3-e-][1,2,4]triazolo[1,5-c]pyrimidin-5-amine (SCH 442416; 1.0 mg/kg) and postsynaptic 8-[(E)-2-(3,4-dimethoxyphenyl)etheyl]-1,3-diethyl-7-methylpurine-2,6-dione (KW 6002; 1.0 mg/kg) A2AR antagonists and showed that only a postsynaptic blockade of these receptors increased the locomotor activity induced by acute cocaine treatment in habituated rats.
Animal genetic models did not explicitly confirm the above pharmacological observations. Thus, Chen et al. [72] demonstrated that locomotor responses to amphetamine and cocaine were attenuated in mice lacking A2ARs, while Wright et al. [73] did not observe changes in locomotion after cocaine administration in A2AR KO animals while hyperactivity following chronic methamphetamine was attenuated.
To explain the differences between pharmacological inhibition and genetic elimination, as well as better understanding of the role A2ARs in the modulation of psychostimulant hyperactivity, a study was conducted in which cocaine-induced hyperactivity was enhanced in striatum-specific A2AR KO mice (selective deletions of A2ARs only in the neurons of the striatum) and attenuated in forebrain-specific A2AR KO mice (deletions A2ARs in the neurons of striatum, cerebral cortex, and hippocampus). In addition, the administration of a selective A2AR antagonist—KW 6002 (3.3 mg/kg)—into striatum-specific A2AR KO mice attenuated the cocaine effects, in contrast to the enhanced cocaine effects observed in wild-type mice. Therefore, these results highlight the opposing modulation in locomotor responses to cocaine—the stimulatory role of A2ARs in the extrastriatalneurons and the inhibitory action of A2ARs in the striatal neurons [74].
Research has demonstrated that A2AR activity affects rodent behavioral responses not only after acute psychostimulant administration but also with chronic exposure. CGS 21680 (0.2 mg/kg), given during the development of cocaine sensitization, reduced the locomotor response to cocaine challenge dose following 5-day withdrawal. The inhibition of hyperlocomotion augmentation in rats after acute CGS 21680 (0.2 mg/kg) administration before a cocaine challenge was also observed [68]. In A2AR overexpressing animals, a challenge with cocaine evoked a slight increase in locomotor activity in comparison to wild-type rats [69]. Taken together, the stimulation/increased activity of A2ARs protected against the development and expression of cocaine sensitization. Shimazoe et al. reported, that, in addition to affecting the effects of cocaine CGS 21680 at higher doses (1.0–3.0 mg/kg), this effect reduced the development, and in the lower dose (0.1 mg/kg), the expression of methamphetamine sensitization [75].
In addition to observations related to A2ARs antagonism on acute psychostimulant injection, researchers demonstrated that MSX-3, in contrast to agonist (CGS 21680), enhanced the development and expression of cocaine sensitization [68]. On the other hand, in the case of cocaine, similar to the effects of single administration, studies showed that the behavioral effect (motor activity) differed depending on the ligand used. Hence, a postsynaptic (KW 6002; 1.0 mg/kg) A2AR antagonist had no effect on the expression of locomotor sensitization, while a presynaptic blockade with SCH 442416 (1.0 mg/kg) inhibited the expression of locomotor sensitization [71]. Another study, in turn, showed that in mice, a blockade of A2ARs with 2-(2-furanyl)-7-(2-phenylethyl)-7H-pyrazolo[4,3-e][1,2,4]triazolo[1,5-c]pyrimidin-5-amine (SCH 58261; 0.03 mg/kg) or KW 6002 (0.03 mg/kg) prevented or delayed the development of sensitization to amphetamine [76].
Similar behavioral effects (limitation development of amphetamine sensitization) were observed in mice with a global lack of A2ARs [77], in forebrain-specific A2AR conditional KO mice [76]. The enhanced motor response characteristic for the development of cocaine sensitization did not change in mice with the genetic inactivation of A2ARs [78]. The literature providedlittle and inconsistent evidence regarding the influences of A2ARs on locomotor responses to other drugs of abuse. It has only been proven that, in the case of opioids, CGS 21680 (0.1 mg/kg) significantly reduced the development of morphine hypersensitivity (with no increase in locomotor activity by the challenge morphine dose following the 7-day withdrawal) in mice induced by increasing doses or sporadic administration of this substance [79,80].
Castañé et al. [81] did not observe differences between mice lacking A2ARs and wild-type littermates in acute effects induced by morphine (similar increase in locomotor activity after morphine injection was observed) [81]. In contrast to psychostimulants and morphine, THC administration decreased locomotor activity. In a study comparing the effect of THC (5–20 mg/kg) acute treatment in A2AR KO and wild-type mice resulted in similar hyperlocomotion in both experimental groups [82]. In a pharmacological study, an antagonist of A2AR MSX-3 (3.0 mg/kg) blocked the locomotor depressant responses observed after intrastriatal cannabinoid CB1 receptor agonist (WIN 55212-2) treatment [40]. The same antagonist (MSX-3; 2.0–4.0 mg/kg) increased locomotion during ethanol consumption in the Drinking-in-the-Dark paradigm in mice [83].
One study assessed the impact of deletions of A2AR on the locomotor effects of acute alcohol administration; however, depending on the genetic background, increases in hyperactivity were noted in mice (mice generated on CD1 background) or no changes were noted (mice generated on C57BL/6J background). In addition, there was no effect of a lack of A2ARs in CD1 mice locomotor sensitization development induced by chronic ethanol administration [84]. In the use of nicotine, a pharmacological study showed that the A2AR agonist CGS 21680 (0.2–0.4 mg/kg) inhibited, and the A2AR antagonist KW 6002 (0.5 mg/kg) enhanced the acute locomotor effect of nicotine in rats.
An A2AR agonist (0.4 mg/kg), but not antagonist, modulated the development of nicotine induced locomotor sensitization (decrease in the locomotor response to the nicotine challenge), while, during evaluation, the expression of nicotine sensitization, stimulation of A2AR was reduced, and a blockade of this receptor increased the locomotor response to a nicotine challenge [85]. In turn, acute nicotine treatment induced a reduction of horizontal movements in A2AR KO mice and wild-type littermates [86].
The above data provided strong evidence for the pre-/post-synaptic and brain structure-dependent involvement of A2ARs in the locomotor response of animals to psychostimulants and morphine treatment. At the same time, more research is needed to clearly assess their involvement in the motor effects associated with the use of other substances.
3.2. Discrimination
The drug discrimination paradigm is a well-established behavioral procedure to model the subjective effects of drugs in animals. So far, the available research on A2ARs-drugs of abuse interaction demonstrated that, in rats trained to discriminate methamphetamine or cocaine from its vehicle (saline) under a fixed ratio (FR)10 schedule of food presentation, A2AR agonist CGS 21680 (0.056–0.177 mg/kg) shifted the cocaine dose–response curve markedly to the left and produced partial, but significant cocaine-lever selection in substitution tests [87]. The same authors found that CGS 21680 produced neither substitution for the methamphetamine-training stimulus response nor a change in its dose–response curve in the combination protocol. For both psychostimulants, CGS 21680 reduced animal response rates in the doses used. A pharmacological blockade of A2ARs with MSX-3 (3–56 mg/kg) significantly shifted the dose–response curves of cocaine and methamphetamine to the right as well as caused high levels of drug-lever selection for both psychostimulants in substitutions tests [87,88].
These observations were consistent with previous reports in which another A2AR antagonist, DMPX (5.6 mg/kg), in rats trained to recognize methamphetamine from saline shifted the drug dose–response curve to the left while in doses of 10–18 mg/kg, it led to generalization of the methamphetamine-training stimulus. Methamphetamine-like effects of this A2AR antagonist were blocked partially by the DA D1 receptor antagonist (R-(+)-7-chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-3-benzazepine hydrochloride (SCH-23390) and fully by the DA D2 receptor antagonist spiperone, which indicates the interaction between adenosine and dopaminergic signaling in the control of subjective effects of this drug of abuse [89].
3.3. Reward
In preclinical studies, the rewarding effects of drugs of abuse are evaluated in self-administration or conditioned place preference (CPP) paradigms. In 2001, the first report indicated that in rats trained for intravenous cocaine self-administration (0.6 mg/kg/infusion) under a FR5 schedule of reinforcement, the A2AR agonist CGS 21680 (0.1–0.4 mg/kg) reduced the number of drug infusions [90]. The reduced number of cocaine infusions was mainly due to the prolonged time of responding to the first drug infusion at the beginning of the session, and this effect was associated, among others, with the sedative effects of CGS 21680 [90]. The same agonist (0.2–0.4 mg/kg) produced a downward shift in the cocaine dose–response curve and,more importantly, CGS 21680 reduced the number of active lever presses and cocaine (0.5 mg/kg/infusion) reinforcements defined as the breaking point in the progressive ratio (PR) reinforcement schedule[91].
The latter authors found also that none of the CGS 21680 effective doses changed the inactive lever responses, which indicated the drug specificity. Wydra and colleagues [92] also pointed to the important role of the localization of A2ARs in controlling the rewarding effects of cocaine. Namely, in the intra-accumbal shell, but not the intra-infralimbic prefrontal cortex, microinjections of CGS 21680 (1.0–2.5 ng/side) dose-dependently decreased cocaine (0.25 mg/kg/infusion) self-administration. Interestingly, the CGS 21680 (0.1 mg/kg)-induced reduction of the number of presses on the active lever and the number of cocaine infusions (0.25 mg/kg/infusion) was not changed by the A2AR synthetic trans membrane (TM) 2 peptide (0.1 µM/0.5 µL/min/side) bilaterally microinjected into the nucleus accumbens [93], while the same dose of a local administration of the TM 5 peptide (which disrupts A2A-D2 heteroreceptor complexes) counteracted the inhibitory effects of CGS 21680 on cocaine self-administration [94].
Supporting the self-administration paradigm, stimulation of A2ARs with CGS 21680 (0.25–0.5 mg/kg) decreased the acquisition and expression of cocaine-induced CPP [70] or amphetamines [95], while the drug, in a dose of 0.03 mg/kg,also inhibited the development of methamphetamine-induced CPP [66]. However, a CGS 21680-induced reduction of drug reward was not observed, after stimulation of A2ARs with this agonist in low doses (0.01–0.03 mg/kg) on methamphetamine self-administration (0.05–0.1 mg/kg/infusion) under FR5 or a progressive ratio schedule of reinforcement [66].
If pharmacological stimulation of A2ARs attenuated the drug reward, the tonic activation of these receptors with selective antagonists seemednot to be linked to drug motivational or reinforcing effects. Thus, neither systemic administration of MSX-3 (3.0 mg/kg) nor systemic or local (into the nucleus accumbens or prefrontal cortex) microinjections of KW 6002 (1.0–2.5 μg/side) or SCH 58261 (1.0–2.5 µg/side) altered cocaine self-administration in squirrel monkeys [96] or rats [92].
In line with the above findings, A2AR KO animals showed no behavioral differences compared to the control mice in a cocaine–induced CPP paradigm[78]. Self-administration procedures provided evidence that, in A2AR KO mice, the cocaine reward was reduced compared to wild-type littermates under FR1 on FR3 schedules. The deletion of A2ARs resulted in a reduction in the maximum effort to obtain a cocaine infusion under a PR schedule of reinforcement [78]. Such inhibition was also observed for MDMA self-administration under an FR1 schedule of reinforcement in A2AR KO mice [97].
The above data suggested that A2ARs participate in the control of the rewarding and motivational properties of psychostimulants, the effects of CGS 21680 in animal paradigms to evaluate such actions are drugs of abuse-sensitivity, while the effects of blockade of these receptors by receptor specific ligands or via genetic modification are undefined so far.
Two studies evaluated the rewarding properties of nicotine. In the first, CGS 21680 (0.03–0.09 mg/kg) was effective in alleviating nicotine CPP, but only in male adolescent rats, suggesting a sex difference in adenosine signaling [98]. The second study showed that, in mice, the deletion of A2ARs suppressed nicotine-induced CPP compared with wild-type animals [86].
The interaction between A2ARs and opioids at the behavioral level showed that in rats CGS 21680 (0.01–0.05 mg/kg) increased the number of drug infusions during the acquisition of morphine self-administration, while the A2AR antagonist DMXP (0.25–1.0 mg/kg) reduced morphine self-administration. Interestingly, the same ligands, given repeatedly (before training sessions), had the opposite effect during the maintenance of morphine self-administration (CGS 21680 decreased, and DMXP increased the number of intravenous morphine injections) [99]. Using morphine-induced CPP, researchers demonstrated that neither CGS 21680 (0.025–0.05 mg/kg) nor SCH 58261 (0.5–1.0 mg/kg) significantly affected the expression of sensitization to the opioid in previously conditioned rats.
On the other hand, the same ligands inhibited the acquisition of sensitization to morphine-evoked CPP [100]. Supporting the last pharmacological study, the rewarding effects of morphine (5 or 10 mg/kg) in CPP were not completely observed in mice lacking A2ARs as opposed to wild-type animals [81]. Confirming this observation, the deletion of A2ARs significantly reduced morphine self-administration under the FR1 schedule and breaking point on the PR schedule, highlighting their participation in controlling the motivational properties of this opioid [101].
Other studies reported that a blockade of A2ARs in squirrel monkeys with a low dose of its antagonist, MSX-3 (1 mg/kg), caused downward shifts of the THC and anandamide (the endogenous cannabinoid with CB1 and CB2 agonistic activity) dose–response curves [96]. The higher dose (3 mg/kg) of MSX-3 shifted the above CB dose–response curves to the left, testifying to their enhancement in the rewarding effects [96]. In addition, in A2AR KO mice, a reduction in the rewarding effects of THC was observed in the CPP model [82].
In studies on the rewarding effects of ethanol, CGS 21680 (0.5–1.0 mg/kg) reduced or did not affect the consumption and ethanol preferences in a two-bottle free-choice paradigm in mice [84]. In rats, a low dose of CGS 21680 (0.065 mg/kg) increased ethanol operant self-administration within non-dependent rats while higher CGS 21680 doses (0.095 or 0.125 mg/kg)—similar to the previous findings on mice—significantly reduced operant alcohol responses under FR1 within both non-dependent and dependent rats [102]. Recently, in rats overexpressing A2ARs, no change in the ethanol drinking behavior during the acquisition/maintenance phase in the two-bottle choice paradigm was reported [103].
The findings with the selective A2AR blockade were also inconsistent. Thus, low doses of A2AR antagonist DMPX (1 mg/kg) increased, while higher doses (10–20 mg/kg) significantly reduced the operant ethanol self-administration in Long–Evans rats [104]. Similarly, in Wistar rats, DMPX (3–10 mg/kg) dose-dependently reduced ethanol operant reinforcement [105], while another A2AR blocker, SCH 58261 (2 mg/kg), reduced the response for ethanol in alcohol-preferring rats [106]. Using the “Drinking-in-the-Dark” paradigm in mice, researchers found that MSX-3 (1–4 mg/kg) did not change voluntary alcohol consumption [83], while 4-(2-[7-amino-2-(2-furyl)[1,2,4]triazolo[2,3-a][1,3,5]triazin-5-ylamino]ethyl)phenol (ZM241385; 20 mg/kg) increased the consumption and ethanol preference in a two-bottle free-choice paradigm in mice [107]. Further, the current literature with genetic animal models indicated that male and female A2AR KO mice generated on a CD1 background were characterized by increased ethanol drinking and alcohol preference compared with wild-type mice [108]. The same mice displayed a decrease ethanol-induced CPP, and, as in previous studies, consumed more ethanol with a higher preference, whereas, in the A2AR KO mice produced on a C57BL/6J background, no changes were observed [84].
Taken together, the pharmacological approach indicated that A2ARs exerted inhibitory control over the reward properties evoked by drugs of abuse.
3.4. Seeking Behavior
In the context of substance use disorder therapy, relapse or drug-seeking behavior after a period of abstinence is the most serious limitation of effective treatment. In animal models, drug-seeking behavior is assessed by re-exposing animals to a previously used drug of abuse, the drug-associated context (cue) or stressor [109].
Many literature reports provided strong conclusions on the inhibitory effects of A2ARs on cocaine-seeking behavior. Namely, the A2AR agonist CGS 21680 dose-dependently (0.03–0.3 mg/kg) inhibited cocaine-induced reinstatement (15 mg/kg) after a 7-day period of drug abstinence in Sprague–Dawley rats. At the same time, this agonist (0.03 mg/kg) also blunted cocaine-seeking behavior induced by quinpirole (a D2 dopaminergic receptor agonist) or a drug-associated cue [110].
Wydra et al. confirmed these observations in Wistar rats, in which CGS 21680 in a low dose range (0.05–0.4 mg/kg) significantly attenuated cocaine- (10 mg/kg;i.p.), quinpirole- and cue-induced reinstatement of cocaine-seeking behavior [111]. This effect may be associated with adenosine signaling in the rat nucleus accumbens, as reduced cocaine-induced (15 mg/kg), quinpirole-induced [112], or the cue plus the subthreshold dose of cocaine-induced (2.5 mg/kg) [92] reinstatement of cocaine seeking was reported following the intra-accumbal microinjection of CGS 21680 (2.5 ng/side), but not after pharmacological stimulation of the A2ARs within the prefrontal cortex [92]. The effects of the A2AR stimulation with CGS 21680 (0.3 mg/kg) in methamphetamine-seeking behavior were partially consistent with the above observations for cocaine. Namely, this agonist blunted quinpirole-induced drug-paired lever responses, but did not affect methamphetamine-seeking induced by a methamphetamine priming injection in rats [113].
Contrary to the stimulation of A2ARs, their accumbal and systemic blockade with MSX-3 (10 µg/side and 6 mg/kg, respectively) or SCH 58261 (5 µg/side) produced cocaine-seeking behavior [92,112]. In addition, intra-accumbal microinjection with MSX-3 (10 µg/side) [112], but not KW 6002 (2.5–5.0 µg/side) and SCH 58261 (2.5 µg/side) [92], potentiated the reinstatement of cocaine-seeking behavior induced only by sub-threshold doses of cocaine and quinpirole, suggesting that removing the tonic activity of A2AR enabled behaviors mediated by DA receptors [112]. Similarly, a systemic blockade of A2ARs by KW 6002 (0.25–0.5 mg/kg) or SCH 58261 (2 mg/kg) [91], or KW 6002 (1–3 mg/kg) [71] in rats extinguished from cocaine self-administration or 9-chloro-2-(2-furanyl)-[1,2,4]triazolo[1,5-c]quinazolin-5-amine (CGS 15943; 0.032–0.32 mg/kg) in baboons [114] reinstated cocaine-seeking, and this effect was eliminated by the D2-like receptor antagonist raclopride (0.1–0.4 mg/kg) [111].
In contrast, KW 6002 at low doses (0.0625 mg/kg or 0.125 mg/kg), but not SCH 58261, evoked a reinstatement of cocaine-seeking behavior induced by sub-threshold dose of cocaine (2.5 mg/kg) or the drug-associated cue, respectively [111]. However, repeated blocking of presynaptic, but not postsynaptic, A2ARs during extinction training with SCH 442416 (0.3 or 1 mg/kg) produced enduring reductions on cocaine- and quinpirole-induced reinstatement of cocaine-seeking behavior [115]. This finding was also confirmed by recent studies, in which KW 6002 (the postsynaptic A2AR antagonist), at a dose of 0.3 mg/kg, enhanced, while SCH 442416 (the presynaptic A2AR antagonist), at a dose of 1 mg/kg, reduced cocaine-induced reinstatement of cocaine seeking [71]. The above data, in addition to confirming the important role of A2ARs to control cocaine-seeking behavior, indicated the antagonistic effect of A2ARs and D2 receptors in cocaine relapse.
Few papers focused on assessing the role of A2ARs in the seeking behavior of other drugs of abuse. Thus, Yao et al. [116] reported that, in rats, a pharmacological blockade of A2ARs with systemic MSX-3 (3 mg/kg) or intra-accumbal DMPX (5 nmol) administration completely abolished reinstatement of heroin-seeking behavior induced by unconditional stimuli (heroin, 0.25 mg/kg). In subsequent studies, using mice with different genotypes, no differences were observed in the cue-induced morphine-seeking behavior after 3 weeks of drug abstinence in the home cage between A2AR KO and wild-type animals [101].
The selective A2AR antagonist, MSX-3 (1 or 3 mg/kg), as opposed to the effect on heroin-seeking behavior in rats, neither promoted the reinstatement of extinguished THC-seeking behavior, nor altered the reinstatement of drug-seeking behavior induced by priming doses of THC (10 or 40 μg/kg; i.v.) in squirrel monkeys [96].
In the case of alcohol, there was no direct evidence of A2ARs in ethanol-seeking behavior. However, co-administration of a sub-threshold doses of the A2AR antagonist SCH 58261 (0.5 mg/kg) and glutamate mGlu5 receptor antagonist 3-[(2-methyl-1,3-thiazol-4-yl)ethynyl]-pyridine (MTEP; 0.25 mg/kg) effectively blocked conditioned reinstatement of alcohol-seeking in alcohol-preferring rats extinguished from ethanol (10% v/v) self-administration [106] (Figure 2).
3.5. Withdrawal
There are several preclinical reports on the role of A2ARs in mediating drugs of abuse withdrawal syndromes. For example, O’Neill et al. [115] observed that in rats self-administering cocaine (0.5 mg/kg) under an FR1 schedule of reinforcement, the A2AR agonist CGS 21680 (0.03–0.1 mg/kg) reduced the extinction response during the few first extinction sessions, while the A2AR antagonist SCH 442416 (0.3–3.0 mg/kg) was inactive during this behavioral paradigm. In turn, a study using a brain stimulation reward task showed that the A2AR antagonist DMPX (3–10 mg/kg) reversed the threshold elevation produced by cocaine withdrawal in rats [117].
The initial evidence for the involvement of A2ARs in opioid withdrawal symptoms was inconclusive. Namely, the A2AR agonist 2-phenylaminoadenosine (CV-1808; 30 or 100 μg/kg) attenuated naloxone-precipitated withdrawal sings (i.e., teeth chattering or diarrhea) in morphine-dependent rats, but this effect was not dose-dependent [118]. Subsequent studies confirmed the inhibitory effects of the A2AR stimulation on some naloxone-precipitated withdrawal symptoms in morphine-dependent rodents. For example, CGS 21680 (0.01 mg/kg) significantly inhibited teeth chattering and forepaw treads in mice [119], and the same agonist (0.03–0.3 mg/kg) decreased in the incidence of body shakes, teeth chattering, and paw shakes in female rats [120]. In turn, a pharmacological blockade of A2ARs with DMPX (1 or 30 mg/kg) increased the incidence of body shakes during morphine withdrawal [119,120].
The inhibitory contribution of A2ARs in the expression of the physical dependence on opioids was further confirmed by studies using the A2AR KO mice. Significant increases in naloxone-induced morphine withdrawal somatic signs (the number of jumps, paw tremors, and writhes) in A2AR KO compared to wild-type mice were observed [121]. Bilbao and colleagues [122] demonstrated that, while A2AR KO mice were characterized by significant increase of diarrhea, sniffing, and the global withdrawal score, the manifestations of naloxone-precipitated morphine withdrawal syndrome were not modified in CB1/A2AR double KO mice, which may suggest opposite roles of these receptors in the morphine dependence [123]. Equally interesting, a lack of A2ARs abolished quasi-morphine withdrawal syndrome induced by the co-administration of caffeine (an A1 and A2A receptors antagonist) and naloxone in opiate-naive mice [122].
Little is known about the role of A2ARs in the control of nicotine, CB, and alcohol withdrawal signs. A separate report indicated that the deletion of A2ARs did not affect the development of the somatic signs of withdrawal induced by the nicotinic receptor antagonist mecamylamine to mice receiving repeated nicotine injections (10 or 25 mg/kg/day for 6 days) by using minipumps [86], suggesting no participation of A2ARs in nicotine withdrawal.
Few reports described A2ARs engagement in THC or alcohol withdrawal symptoms. Soria et al. [82] observed that the A2AR deletion significantly attenuated somatic manifestations (global withdrawal score and paw tremors) of CB1 receptor antagonist N-(piperidin-1-yl)-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide hydrochloride (SR141716A)-precipitated THC withdrawal in mice receiving chronic treatment with THC (20 mg/kg). In turn, alcohol studies found that the A2AR agonist CGS 21680 (0.3 mg/kg) reduced the withdrawal score in mice who chronically (14 days) consumed a liquid diet containing ethanol (6.7%, v/v) [124], while another agonist (DPMA, 0.1–5 m/kg) did not alter the anxiety-like behavior during ethanol hangovers in mice evaluated in the elevated plus maze test [125].
On the other hand, observations by El Yacoubi and colleagues [126] indicated that a blockade of A2ARs with ZM241385 (20 mg/kg) during the last 5-days of the ethanol intake attenuated ethanol withdrawal-induced seizures in wild-type mice. The handling-induced convulsions score during ethanol withdrawal was lowered in the A2AR KO mice when compared with wild-type controls, pointing to A2ARs as a potential target for the treatment of symptoms of alcohol withdrawal.
4. A2AR and Behavioral Actions to Cocaine and Other Drugs of Abuse—Ex Vivo Evidence
Several preclinical reports using different techniques (i.e., biochemical binding, autoradiography, immunoblotting,and PLA) demonstrated, in theex vivoresearch, changes of the A2AR expression in brain areas relevant to repeated exposure to cocaine and other drugs of abuse.
The first quantitative autoradiographic mapping on cocaine showed no difference in the density of A2ARs in the accumbal (core or shell) subregions, dorsal striatum, globus pallidus, and olfactory tubercle in Fischer rats after 14-daily escalating dose "binge" cocaine administration paradigm and a 14-day withdrawal [127]. In 2007, Marcellino and colleagues reported using the A2AR antagonist radioligand [3H]ZM241385 or the immunoblots that intravenous cocaine self-administration with a discrete trials schedule (4 trials/h) by 10 days evoked an up-regulation in the nucleus accumbens and a down-regulation in the posterior dorsal striatum of functional A2ARs while these changes disappeared to baseline levels after 7 days of drug withdrawal in male Sprague Dawley rats [128]. Another report coming from our laboratory demonstrated that 16 days with i.v. cocaine given to yoked Wistar rats, serving as a control to self-administering cocaine, evoked a significant increase in the A2AR density, but not affinity, in the ventral striatum, while the same amount of cocaine self-administered by rats failed.
This increase in the A2AR density was still observed following extinction training (10 days) in the yoked group but also appeared in animals previously self-administered cocaine. In contrast to the ventral striatum, no changes were found in the A2AR Bmax values, in the dorsal striatum in both groups of rats with either an active or passive history of cocaine administration and after 10-day extinction training [129]. The increased A2AR expression was also reported in the hypothalamus of male Sprague–Dawley rats subjected to 8 and 14 days of cocaine withdrawal following 7-daily s.c. cocaine injections [130].
Recently, we showed that a microinjection of synthetic peptide TM5, a part of the A2AR-D2 receptor interface, into the nucleus accumbens during cocaine self-administration, blocked in vivo the inhibitory action of the A2AR agonist CGS 21680 on cocaine self-administration, and ex vivoreduced the number of A2A-D2 heteroreceptor complexes within the nucleus accumbens, but not in the dorsal striatum [94]. Using the same PLA method, another transmembrane peptide, TM2, which did not interfere with the formation of the A2A-D2 heteroreceptor complex, failed to alter CGS 21680-induced inhibition of a cocaine reward or the number of PLA blots identifying A2A-D2 heteroreceptor complexes in the nucleus accumbens, but it reduced the number of A2A-A2Ahomoreceptor complexes in this brain structure in Sprague Dawley rats [93].
The separate report on methamphetamine indicated that experimenter-delivered drugs produced no changes in the A2ARs expression in the striatum, while methamphetamine self-administration for 14 days decreased A2ARs in the accumbens shell and increased them in the amygdala in Sprague Dawley rats [66].
In mice, opposing functional interactions between A2ARs and CB1receptors in the hippocampus, as well as the Δ9-THC-induced cognitive impairment being blocked by an A2AR antagonist, were demonstrated [131]. Such interactions might also occur in the microglia [132]; however, this must be confirmed in further studies.
The opposite findings of the involvement of morphine on the expression of striatal A2ARs indicated that 72-h s.c. delivery of drugs by implanted pump did not change the expression in mice [133], while chronic administration of morphine resulted in a decreased density and affinity of the receptors in rats [134].
Acute or prolonged ethanol exposure and short- or long-term drug withdrawal did not change the A2AR numbers and function values in the striatum in mice and rats [124,135,136,137,138,139,140]. Further, long-term voluntary alcohol drinking did not alter gene expression of the accumbal or striatal A2ARs, but significantly increased the number of PLA blots identifying A2A-D2 heteroreceptor complexes in the accumbal shell and the dorsal striatum in Wistar rats [141].
Adolescent long-term caffeine consumption in adulthood produced decreases in A2AR expression with simultaneous increases in D2 receptor and protein for DAT and DARPP-32 in the rat nucleus accumbens [142].
To summarize, repeated administration of drugs of abuse evoked changes in the A2AR expression seen mainly in the nucleus accumbens and dorsal striatum in rodents and the decreases in these receptors were linked with the local up-regulation of the D2receptor or the new A2A-D2heteroreceptor formation.
5. Neurochemical Correlates to Behavioral Findings in Rat Models of Substance Use Disorder. A Focus on Cocaine
The early work of Filip and colleagues [68] indicated that the stimulation of A2ARs can produce inhibitory effects over acute cocaine-induced increases in locomotion and over repeated cocaine-evoked expression of locomotor sensitization in habituated rats. In this paper, the neurochemical mechanism proposed was related to the existence of antagonistic intramembrane A2AR-D2 receptor interactions in the brain [143] through the formation of A2AR-D2 receptor heteromeric complexes [30]. The interface between A2AR-D2 receptor was shown to involve the C-terminal tail and the transmembrane helices [144]. More recently, based on the acute effects of cocaine in the nanomolar range (1–100 nM) in the D2 receptor-Sigma1 receptor complexes, putative A2A-D2 receptor-Sigma1 heteroreceptor complexes appeared to have relevance for the treatment of cocaine use disorder [145].
5.1. New Mechanisms of Acute Cocaine
As shown in the in vitro studies, cocaine in the nanomolar range enhanced the function of the D2 receptor [146,147], related to the existence of D2receptor-Sigma1 receptor heterocomplexes and through (i) enhancement of the Gi/o-mediated signaling of D2 receptor protomers via cocaine binding to the Sigma1 receptor and (ii) counteraction of the D2receptor protomer internalization HEK293 cells [145]. In the absence of a dominance of A2AR-D2 receptor-Sigma1 receptor complexes, the action of cocaine likely switches from inhibition to enhancement of D2 receptor protomer recognition and signaling.
So far only the dorsal striatum has been studiedin vivo after acute treatment with cocaine. Our present research demonstrated that, in rats, cocaine at a low dose (1 mg/kg)—without increasing the extracellular DA levels—selectively affected the Sigma1 receptors [148]. Further, in the brain tissue of such animals, the A2AR agonist CGS 21680 (100 nM) produced significantly larger decreases in the affinity of the high affinity state versus those observed for the saline-treated rats. In the ex vivo studies, cocaine evoked significant reductions of the low and high affinity of D2 receptors. Such effects were missing in the control rats. The cocaine induced increases in the antagonistic A2AR-D2 receptor in the dorsal striatum werelikely due to cocaine-induced increases in the formation of A2AR-D2 receptor-Sigma1 receptor higher order complexes, in which cocaine can bind to the Sigma1 receptor. The resulting allosteric changes in the D2 receptor, through the receptor-protein interaction, may strongly enhance the antagonistic allosteric A2AR-D2 receptor interactions [148]. Thus, at least in the dorsal striatum acute cocaine in low doses in vivo, not increasing extracellular DA levels, markedly increased the allosteric inhibition of D2 receptor recognition.
5.2. Cocaine Self-Administration
In rats, cocaine self-administration induced no changes in the A2AR density while a 10-day abstinence with extinction training increased the density of A2ARs in the ventral striatum [129]. These observations may be interpreted to reflect a significant and stronger increase of the A2A-D2 heteroreceptor complex in the ventral striatum in extinction vs. the maintenance of cocaine self-administration. In contrast, in the “yoked” cocaine controls, which received cocaine passively, an increase in the A2AR density was developed also in the maintenance phase of cocaine self-administration [129]. This finding may reflect an increase in the antagonistic A2AR-D2 receptor interactions linked with reduced motivation.
The changes in the A2AR density in the ventral striatum appeared to be associated with changes in the overflow of DA, GABA, and glutamate transmitters in cocaine self-administration and its extinction [149]. In fact, during the maintenance of cocaine self-administration, the DA and GABA changes in the accumbal and ventral pallidal overflows may be positively correlated to the motivational features of cocaine intake. On day 10 of extinction (cocaine-free period) reduced basal glutamate extracellular levels were found in the nucleus accumbens [149]. Thus, the panorama of transmitter changes appeared to be different in cocaine self-administration vs. day 10 of extinction. They can in part be related to differential integration of the A2AR-D2 receptor heterocomplexes in these two states through different types of allosteric receptor–receptor interactions.
Using pharmacological analysis (see above), research demonstrated that A2AR antagonists lacked effects on cocaine self-administration while the A2AR agonist also gave systemically reduced cocaine rewards and motivation [91]. Thus, there was not an endogenous tone of adenosine activating A2ARs during cocaine self-administration but upon local steady-state infusion of the A2AR agonist CGS 21680 an increase in active lever pressing was found instead. These behavioral changes were associated with increased extracellular GABA and reduced DA levels in the nucleus accumbens. Although unexpected, the neurochemical and behavioral data can be explained by the existence of several networks. In these networks, A2ARs control over the cocaine reward and the transmitter events were proposed to be caused by the A2AR agonist-induced activation of the post-junctional A2AR protomer inhibiting the D2 receptor protomer inhibitory signaling in the A2AR-D2 receptor complex in the ventral striato–pallidal GABA antireward neurons.
In this way, the reduction of D2 receptor inhibition of the above-mentioned GABA neurons can explain the increase of extracellular GABA levels observed in the nucleus accumbens[91]. As a consequence, GABA volume transmission can be enhanced, followed by the activation of the GABA receptors, by which, enhanced inhibition of DA release can develop and reach the adjacent DA terminals. Such a mechanism can mediate the reduced extracellular DA levels observed in the nucleus accumbens [91].
In 2016, the first evidence supporting the above hypothesis was obtained using CGS 21680 ex vivo, which significantly reduced the affinity of the D2-like receptors in the ventral striatum in self-administering cocaine rats vs. yoked cocaine controls [150]. An inhibitory feedback developed in cocaine self-administration through enhancing the antagonistic allosteric receptor–receptor interactions in the A2AR-D2receptorheteroreceptor complexes in the ventral striatum. In contrast to the nucleus accumbens, the inhibitory interaction in the A2AR-D2 receptor complexes in the dorsal striatum was blocked.
The enhancement of D2 receptor protomer signaling in the latter brain region may contribute to locomotor sensitization and habit forming learning. Our results demonstrated not only the differential modulation of the A2AR-D2 receptor heterocomplexes in the ventral vs. the dorsal striatum [148], but suggested that functional plasticity in striatal networks in cocaine addiction could clear-cut increase the antagonistic allosteric A2AR-D2 receptor interactions in the ventral striatum [151]. Intra-accumbalmicroinjections of CGS 21680 further emphasized the nucleus accumbens as a target for the inhibitory actions of A2AR agonists on cocaine self-administration [92]. Microinjections of the A2AR agonist into the prefrontal cortex lacked effects on cocaine rewards in self-administration.
The continued work on the neurochemistry of cocaine self-administration demonstrated significant increases in A2AR-D2 receptor and D2 receptor-Sigma1 receptor heterocomplexes in the shell region of the nucleus accumbens [152]. The increase in the D2 receptor-Sigma1 receptor complexes in this region may reflect the enhanced formation of A2AR-D2 receptor-Sigma1 receptor heterocomplexes that, in addition, can produce an increased strength of the antagonistic allosteric A2AR-D2 receptor interactions [150]. The work of Romieu et al. [153] demonstrated that cocaine could increase the density of Sigma1 receptors in the nucleus accumbens. Cocaine can also bind to the Sigma1 receptor and recruit it into the plasma membrane where the Sigma1 receptor can bind to the D2 receptor, increasing the number of D2 receptor-Sigma1 receptor complexes in the surface membrane [154,155].
The work on the role of the Sigma1 receptor in cocaine self-administration was resumed using a new pharmacological tool, the monoamine stabilizer OSU-6162, which, in low doses, is a selective Sigma1 receptor ligand with a nanomolar affinity for the receptor [156]. However, in the low dose of 5 mg/kg OSU-6162—when pretreated before cocaine—did not significantly reduce the active lever pressing and the number of cocaine infusions [157]. Only a trend for a reduction of cocaine self-administration was noted after daily injections of OSU-6162 for three days. As shown with competition binding experiments, three days of treatment with OSU-6162 did produce a substantial increase in the density of the D2 receptor-Sigma1 receptor complexes in the nucleus shell upon cocaine self-administration vs. the increase found upon treatment with the same OSU-6162 dose in yoked saline animals using ex vivo PLA.
In the same experiment, treatment with OSU-6162 in rats self-administering cocaine evoked significant increases of the A2AR-D2 receptor heterocomplexes in nucleus accumbens shell vs. the increases obtained for these complexes in the yoked saline animals. The failure to see a significant inhibition of cocaine self-administration with OSU-6162 alone may be the lack of a simultaneous A2AR stimulation of the A2AR protomer in this experiment. Furthermore, in the D2 receptor-Sigma1 receptor complex acute cocaine could enhance D2 receptor recognition and signaling with a parallel blockade of the D2 receptor internalization [145]. In fact, treatment with the A2AR agonist ex vivoin combined treatment within vivoOSU-6162 during cocaine self-administration produced a highly significant reduction of the affinity of the high affinity component of the D2 receptor vs. the reduction of affinity of this state found in yoked saline rats not receiving OSU-6162 [157].
5.3. Cocaine-Seeking
Reinstatement of cocaine seeking was substantially reduced by A2AR agonist treatment with enhanced potency to counteract a D2 receptor agonist- or cue-induced cocaine seeking. In contrast, the A2AR antagonists KW 6002 and SCH 58261 alone were able to evoke cocaine reinstatement. Preliminary data indicated that the antagonistic allosteric A2AR–D2 receptor was absent in cocaine reinstatement. Instead, 10-day cocaine abstinence with extinction training was characterized by a strong antagonistic A2AR–D2 receptor interaction as seen from a clear-cut A2AR agonist induced reduction of affinity in the D2 receptor high affinity state as studiedex vivo in the ventral striatum. Local microinjections of CGS 21680 into the nucleus accumbens, but not into the intra-prefrontal cortex, strongly reduced cocaine reinstatement, which means that the A2AR stimulation controls cocaine seeking and that such interactions depend on the nucleus accumbens [92].
5.4. The Role of the A2AR-D2Receptor Heterocomplexes in Cocaine Use and Addiction
We showed that the synthetic TM5 peptide was part of the interface of the A2AR-D2 receptor heterodimer in HEK293 cells as it blocked the appearance of the BRET signal and disrupted the rat A2AR-D2 receptor complex [144,145]. In the in vivo study, the rat synthetic TM5 peptide caused a full blockade of the inhibitory actions of the A2AR agonist on cocaine self-administration following its microinjection into the nucleus accumbens, while ex vivo it produced a disappearance of the A2AR-D2 receptor complex as well as its allosteric receptor–receptor interactions [158]. Taken together, these results gave evidence that the anti-cocaine actions of A2AR agonists were mediated through an A2AR protomeractivation of an allosteric brake on the D2 receptor protomer recognition and signaling in A2AR-D2 receptor heterocomplexes. The brake appears to be markedly enhanced by activation of the Sigma1 receptor protomer in this receptor complex [158].
At the same time, when the rat synthetic TM5 peptide blocked the inhibitory action of CGS 21680 on the cocaine reward, the rat synthetic TM2 peptide (which neither belongs to the receptor interface of the A2AR-D2 receptor heterodimer [144,159] nor participates in the interface of the A2AR-A2AR homodimer [93]) did not block the formation of the A2AR-D2 receptor heterodimer as determined with a BRET assay and did not interfere with the effects of cocaine in self-administration procedures. We reported that the nucleus accumbens microinjection with TM2 peptide reduced the formation of the A2AR homodimer, however, this had no consequences for the inhibitory actions of the A2AR agonist on cocaine self-administration [93]. Thus, disruption of the A2AR-A2AR homomer caused no effects on rat cocaine self-administration and the A2AR homomeric complex did not appear to have a critical role in cocaine use and addiction.
A transgenic approach based on using rats with the overexpression of A2AR under the regulation of the neuronal specific enolase promotor (NSEA2A) indicated an enhanced number of such receptor complexes with possible increased stoichiometry mainly in the prefrontal cortex, the hippocampal formation, the striatum, and the cerebellum [160]. Such enhancement could lead to increased A2AR signaling, which mediates the substantial inhibitory effects on nicotine behavioral readouts, such as sensitization of locomotion or conditioned locomotor activity (see above). These observations indicated that enhanced A2AR signaling in the above brain areas could also counteract nicotine use.
6. Final Conclusions
The latest preclinical findings indicated a role for A2ARs in several models to assess drug reward, withdrawal, or seeking behavior. Several rodent research models clearly demonstrated that the stimulation of A2ARs was specific to cocaine for the attenuation of drug-induced rewarding effects, withdrawal syndrome, and relapses. Unlike cocaine, no similar effects were seen for other drugs of abuse regarding reward or seeking behaviors. Concerning drug withdrawal,studies reported that the A2AR stimulation was effective for reducing opioid or alcohol withdrawal symptoms. The research information on the interaction A2AR-nicotine or A2AR-cannabinoids is still limited and does not allow us to make conclusions regarding the receptors control over the behavioral or neurochemical outcomes of the above drugs of abuse.
The interaction between A2ARs and cocaine seen at the behavioral level was associated with the antagonistic interplay within A2AR-D2 receptor complexes localized in the central striato–pallidal GABA neurons as shown ex vivoat both the molecular and behavioral levels. This observation should prompt further preclinical research on the study of antagonistic interactions of the A2AR-D2 receptors for investigations on other drugs of abuse as well asresearch in the search for safe and effective anti-cocaine A2AR agonists and/or heterobivalent ligands targeted for A2AR-D2receptors.
Author Contributions
Conceptualization: Małgorzata Filip; Writing—Original Draft Preparation: K.W.; D.G.; K.G.; Małgorzata Frankowska; D.O.B.-E.; K.F.; Małgorzata Filip; Writing—Review and Editing: K.W.; D.G.; K.G.; Małgorzata Frankowska; D.O.B.-E.; K.F.; Małgorzata Filip; Visualization: K.W.; D.G.; K.G.; Małgorzata Frankowska; D.O.B.-E.; K.F.; Małgorzata Filip; Supervision: Małgorzata Filip; Project Administration: Małgorzata Filip; Funding Acquisition: Małgorzata Filip; K.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by statutory funds of the Maj Institute of Pharmacology Polish Academy of Sciences, Krakow, Poland and by the Swedish Medical Research Council (Vetenskapsrådet 2020–2022; 04X-715), Sweden.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McLellan, A.T. Substance Misuse and Substance use Disorders: Why do they Matter in Healthcare? Trans. Am. Clin. Climatol. Assoc. 2017, 128, 112–130. [Google Scholar] [PubMed]
- Bennett, S.; Petrash, P. The Neurobiology of Substance Use Disorders: Information for Assessment and Clinical Treatment. Smith Coll. Stud. Soc. 2014, 84, 273–291. [Google Scholar] [CrossRef]
- Hasin, D.S.; O’Brien, C.P.; Auriacombe, M.; Borges, G.; Bucholz, K.; Budney, A.; Compton, W.M.; Crowley, T.; Ling, W.; Petry, N.M.; et al. DSM-5 Criteria for Substance Use Disorders: Recommendations and Rationale. Am. J. Psychiatry 2013, 170, 834–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, R.A.; Johansson, B.; van der Ploeg, I.; Sebastiao, A.M.; Ribeiro, J.A.; Fredholm, B.B. Evidence for functionally important adenosine A2a receptors in the rat hippocampus. Brain Res. 1994, 649, 208–216. [Google Scholar] [CrossRef]
- Dixon, A.K.; Gubitz, A.K.; Sirinathsinghji, D.J.; Richardson, P.J.; Freeman, T.C. Tissue distribution of adenosine receptor mRNAs in the rat. Br. J. Pharmacol. 1996, 118, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, M.F.; Williams, M. Direct autoradiographic localization of adenosine A2 receptors in the rat brain using the A2-selective agonist, [3H]CGS 21680. Eur. J. Pharmacol. 1989, 168, 243–246. [Google Scholar] [CrossRef]
- Rebola, N.; Canas, P.M.; Oliveira, C.R.; Cunha, R.A. Different synaptic and subsynaptic localization of adenosine A2A receptors in the hippocampus and striatum of the rat. Neuroscience 2005, 132, 893–903. [Google Scholar] [CrossRef]
- Rosin, D.L.; Robeva, A.; Woodard, R.L.; Guyenet, P.G.; Linden, J. Immunohistochemical localization of adenosine A2A receptors in the rat central nervous system. J. Comp. Neurol. 1998, 401, 163–186. [Google Scholar] [CrossRef]
- Rosin, D.L.; Hettinger, B.D.; Lee, A.; Linden, J. Anatomy of adenosine A2A receptors in brain: Morphological substrates for integration of striatal function. Neurology 2003, 61, S12–S18. [Google Scholar] [CrossRef]
- Svenningsson, P.; Le Moine, C.; Aubert, I.; Burbaud, P.; Fredholm, B.B.; Bloch, B. Cellular distribution of adenosine A2A receptor mRNA in the primate striatum. J. Comp. Neurol. 1998, 399, 229–240. [Google Scholar] [CrossRef]
- Svenningsson, P.; Le Moine, C.; Fisone, G.; Fredholm, B.B. Distribution, biochemistry and function of striatal adenosine A2A receptors. Prog. Neurobiol. 1999, 59, 355–396. [Google Scholar] [CrossRef]
- Peterfreund, R.A.; MacCollin, M.; Gusella, J.; Fink, J.S. Characterization and expression of the human A2a adenosine receptor gene. J. Neurochem. 1996, 66, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Schiffmann, S.N.; Jacobs, O.; Vanderhaeghen, J.J. Striatal restricted adenosine A2 receptor (RDC8) is expressed by enkephalin but not by substance P neurons: an in situ hybridization histochemistry study. J. Neurochem. 1991, 57, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Schiffmann, S.N.; Fisone, G.; Moresco, R.; Cunha, R.A.; Ferre, S. Adenosine A2A receptors and basal ganglia physiology. Prog. Neurobiol. 2007, 83, 277–292. [Google Scholar] [CrossRef] [Green Version]
- Brooks, D.J.; Doder, M.; Osman, S.; Luthra, S.K.; Hirani, E.; Hume, S.; Kase, H.; Kilborn, J.; Martindill, S.; Mori, A. Positron emission tomography analysis of [11C]KW-6002 binding to human and rat adenosine A2A receptors in the brain. Synapse 2008, 62, 671–681. [Google Scholar] [CrossRef]
- Lee, Y.C.; Chien, C.L.; Sun, C.N.; Huang, C.L.; Huang, N.K.; Chiang, M.C.; Lai, H.L.; Lin, Y.S.; Chou, S.Y.; Wang, C.K.; et al. Characterization of the rat A2A adenosine receptor gene: a 4.8-kb promoter-proximal DNA fragment confers selective expression in the central nervous system. Eur. J. Neurosci. 2003, 18, 1786–1796. [Google Scholar] [CrossRef]
- Nishizaki, T.; Nagai, K.; Nomura, T.; Tada, H.; Kanno, T.; Tozaki, H.; Li, X.X.; Kondoh, T.; Kodama, N.; Takahashi, E.; et al. A new neuromodulatory pathway with a glial contribution mediated via A(2a) adenosine receptors. Glia 2002, 39, 133–147. [Google Scholar] [CrossRef]
- Sebastião, A.M.; Ribeiro, J.A. Tuning and fine-tuning of synapses with adenosine. Curr. Neuropharmacol. 2009, 7, 180–194. [Google Scholar] [CrossRef] [Green Version]
- Quiroz, C.; Luján, R.; Uchigashima, M.; Simoes, A.P.; Lerner, T.N.; Borycz, J.; Kachroo, A.; Canas, P.M.; Orru, M.; Schwarzschild, M.A.; et al. Key modulatory role of presynaptic adenosine A2A receptors in cortical neurotransmission to the striatal direct pathway. ScientificWorldJournal 2009, 9, 1321–1344. [Google Scholar] [CrossRef] [Green Version]
- Daré, E.; Schulte, G.; Karovic, O.; Hammarberg, C.; Fredholm, B.B. Modulation of glial cell functions by adenosine receptors. Physiol. Behav. 2007, 92, 15–20. [Google Scholar] [CrossRef]
- Hettinger, B.D.; Lee, A.; Linden, J.; Rosin, D.L. Ultrastructural localization of adenosine A2A receptors suggests multiple cellular sites for modulation of GABAergic neurons in rat striatum. J. Comp. Neurol. 2001, 431, 331–346. [Google Scholar] [CrossRef]
- Matos, M.; Augusto, E.; Santos-Rodrigues, A.D.; Schwarzschild, M.A.; Chen, J.F.; Cunha, R.A.; Agostinho, P. Adenosine A2A receptors modulate glutamate uptake in cultured astrocytes and gliosomes. Glia 2012, 60, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.; Augusto, E.; Agostinho, P.; Cunha, R.A.; Chen, J.F. Antagonistic interaction between adenosine A2A receptors and Na+/K+-ATPase-α2 controlling glutamate uptake in astrocytes. J. Neurosci. 2013, 33, 18492–18502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augood, S.J.; Emson, P.C. Adenosine A2a receptor mRNA is expressed by enkephalin cells but not by somatostatin cells in rat striatum: a co-expression study. Brain Res. Mol. Brain Res. 1994, 22, 204–210. [Google Scholar] [CrossRef]
- Fink, J.S.; Weaver, D.R.; Rivkees, S.A.; Peterfreund, R.A.; Pollack, A.E.; Adler, E.M.; Reppert, S.M. Molecular cloning of the rat A2 adenosine receptor: selective co-expression with D2 dopamine receptors in rat striatum. Brain Res. Mol. Brain Res. 1992, 14, 186–195. [Google Scholar] [CrossRef]
- Rodrigues, R.J.; Alfaro, T.M.; Rebola, N.; Oliveira, C.R.; Cunha, R.A. Co-localization and functional interaction between adenosine A(2A) and metabotropic group 5 receptors in glutamatergic nerve terminals of the rat striatum. J. Neurochem. 2005, 92, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; James, S.; Reddington, M.; Richardson, P.J. Both A1 and A2a purine receptors regulate striatal acetylcholine release. J. Neurochem. 1990, 55, 31–38. [Google Scholar] [CrossRef]
- Kurokawa, M.; Kirk, I.P.; Kirkpatrick, K.A.; Kase, H.; Richardson, P.J. Inhibition by KF17837 of adenosine A2A receptor-mediated modulation of striatal GABA and ACh release. Br. J. Pharmacol. 1994, 113, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Kurokawa, M.; Koga, K.; Kase, H.; Nakamura, J.; Kuwana, Y. Adenosine A2a receptor-mediated modulation of striatal acetylcholine release in vivo. J. Neurochem. 1996, 66, 1882–1888. [Google Scholar] [CrossRef]
- Canals, M.; Marcellino, D.; Fanelli, F.; Ciruela, F.; de Benedetti, P.; Goldberg, S.R.; Neve, K.; Fuxe, K.; Agnati, L.F.; Woods, A.S.; et al. Adenosine A2A-dopamine D2 receptor-receptor heteromerization: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J. Biol. Chem. 2003, 278, 46741–46749. [Google Scholar] [CrossRef] [Green Version]
- Canals, M.; Burgueño, J.; Marcellino, D.; Cabello, N.; Canela, E.I.; Mallol, J.; Agnati, L.; Ferré, S.; Bouvier, M.; Fuxe, K.; et al. Homodimerization of adenosine A2A receptors: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J. Neurochem. 2004, 88, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Brito, I.; Romero-Fernandez, W.; Di Palma, M.; Oflijan, J.; Skieterska, K.; Duchou, J.; Van Craenenbroeck, K.; Suarez-Boomgaard, D.; Rivera, A.; et al. The G protein-coupled receptor heterodimer network (GPCR-HetNet) and its hub components. Int. J. Mol. Sci. 2014, 15, 8570–8590. [Google Scholar] [CrossRef] [PubMed]
- Borroto Escuela, D.; Hagman, B.; Woolfenden, M.; Pinton, L.; Jiménez-Beristain, A.; Oflijan, J.; Narváez Peláez, M.; Palma, M.; Feltmann, K.; Sartini, S.; et al. In Situ Proximity Ligation Assay to Study and Understand the Distribution and Balance of GPCR Homo- and Heteroreceptor Complexes in the Brain. In Receptor and Ion Channel Detection in the Brain; Lujan, R., Ciruela, F., Eds.; Humana Press: New York, NY, USA, 2016; pp. 109–124. [Google Scholar]
- Borroto Escuela, D.; Brito, I.; Palma, M.; Jiménez-Beristain, A.; Narváez Peláez, M.; Corrales, F.; Pita, M.; Sartini, S.; Ambrogini, P.; Lattanzi, D.; et al. On the Role of the Balance of GPCR Homo/ Heteroreceptor Complexes in the Brain. J. Adv. Neurosci. Res. 2015, 2, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Hinz, S.; Navarro, G.; Franco, R.; Müller, C.E.; Fuxe, K. Understanding the Role of Adenosine A2AR Heteroreceptor Complexes in Neurodegeneration and Neuroinflammation. Front. Neurosci. 2018, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Ciruela, F.; Casadó, V.; Rodrigues, R.J.; Luján, R.; Burgueño, J.; Canals, M.; Borycz, J.; Rebola, N.; Goldberg, S.R.; Mallol, J.; et al. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. 2006, 26, 2080–2087. [Google Scholar] [CrossRef] [Green Version]
- Ferre, S.; Karcz-Kubicha, M.; Hope, B.T.; Popoli, P.; Burgueno, J.; Gutierrez, M.A.; Casado, V.; Fuxe, K.; Goldberg, S.R.; Lluis, C.; et al. Synergistic interaction between adenosine A2A and glutamate mGlu5 receptors: implications for striatal neuronal function. Proc. Natl. Acad. Sci. USA 2002, 99, 11940–11945. [Google Scholar] [CrossRef] [Green Version]
- Ferre, S.; Popoli, P.; Rimondini, R.; Reggio, R.; Kehr, J.; Fuxe, K. Adenosine A2A and group I metabotropic glutamate receptors synergistically modulate the binding characteristics of dopamine D2 receptors in the rat striatum. Neuropharmacology 1999, 38, 129–140. [Google Scholar] [CrossRef]
- Nishi, A.; Liu, F.; Matsuyama, S.; Hamada, M.; Higashi, H.; Nairn, A.C.; Greengard, P. Metabotropic mGlu5 receptors regulate adenosine A2A receptor signaling. Proc. Natl. Acad. Sci. USA 2003, 100, 1322–1327. [Google Scholar] [CrossRef] [Green Version]
- Carriba, P.; Ortiz, O.; Patkar, K.; Justinova, Z.; Stroik, J.; Themann, A.; Muller, C.; Woods, A.S.; Hope, B.T.; Ciruela, F.; et al. Striatal adenosine A2A and cannabinoid CB1 receptors form functional heteromeric complexes that mediate the motor effects of cannabinoids. Neuropsychopharmacology 2007, 32, 2249–2259. [Google Scholar] [CrossRef] [Green Version]
- Ferré, S.; Lluís, C.; Justinova, Z.; Quiroz, C.; Orru, M.; Navarro, G.; Canela, E.I.; Franco, R.; Goldberg, S.R. Adenosine-cannabinoid receptor interactions. Implications for striatal function. Br. J. Pharmacol. 2010, 160, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Cabello, N.; Gandia, J.; Bertarelli, D.C.; Watanabe, M.; Lluis, C.; Franco, R.; Ferre, S.; Lujan, R.; Ciruela, F. Metabotropic glutamate type 5, dopamine D2 and adenosine A2a receptors form higher-order oligomers in living cells. J. Neurochem. 2009, 109, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Carriba, P.; Navarro, G.; Ciruela, F.; Ferre, S.; Casado, V.; Agnati, L.; Cortes, A.; Mallol, J.; Fuxe, K.; Canela, E.I.; et al. Detection of heteromerization of more than two proteins by sequential BRET-FRET. Nat. Methods 2008, 5, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Agnati, L.F.; Jacobsen, K.; Hillion, J.; Canals, M.; Torvinen, M.; Tinner-Staines, B.; Staines, W.; Rosin, D.; Terasmaa, A.; et al. Receptor heteromerization in adenosine A2A receptor signaling: relevance for striatal function and Parkinson’s disease. Neurology 2003, 61, S19–S23. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Dahlstrom, A.B.; Jonsson, G.; Marcellino, D.; Guescini, M.; Dam, M.; Manger, P.; Agnati, L. The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog. Neurobiol. 2010, 90, 82–100. [Google Scholar] [CrossRef] [PubMed]
- Beggiato, S.; Antonelli, T.; Tomasini, M.C.; Borelli, A.C.; Agnati, L.F.; Tanganelli, S.; Fuxe, K.; Ferraro, L. Adenosine A2A-D2 receptor-receptor interactions in putative heteromers in the regulation of the striato–pallidal gaba pathway: possible relevance for parkinson’s disease and its treatment. Curr. Protein Pept. Sci. 2014, 15, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, A.; Tscherter, A.; Belcastro, V.; Tantucci, M.; Costa, C.; Picconi, B.; Centonze, D.; Calabresi, P.; Borsini, F. Interaction of A2A adenosine and D2 dopamine receptors modulates corticostriatal glutamatergic transmission. Neuropharmacology 2007, 53, 783–789. [Google Scholar] [CrossRef]
- Flajolet, M.; Wang, Z.; Futter, M.; Shen, W.; Nuangchamnong, N.; Bendor, J.; Wallach, I.; Nairn, A.C.; Surmeier, D.J.; Greengard, P. FGF acts as a co-transmitter through adenosine A(2A) receptor to regulate synaptic plasticity. Nat. Neurosci. 2008, 11, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Ravani, A.; Tarakanov, A.O.; Brito, I.; Narvaez, M.; Romero-Fernandez, W.; Corrales, F.; Agnati, L.F.; Tanganelli, S.; Ferraro, L.; et al. Dopamine D2 receptor signaling dynamics of dopamine D2-neurotensin 1 receptor heteromers. Biochem. Biophys. Res. Commun. 2013, 435, 140–146. [Google Scholar] [CrossRef]
- Fuxe, K.; Borroto-Escuela, D.; Fisone, G.; Agnati, L.F.; Tanganelli, S. Understanding the role of heteroreceptor complexes in the central nervous system. Curr. Protein Pept. Sci. 2014, 15, 647. [Google Scholar] [CrossRef]
- Fuxe, K.; Ferré, S.; Canals, M.; Torvinen, M.; Terasmaa, A.; Marcellino, D.; Goldberg, S.R.; Staines, W.; Jacobsen, K.X.; Lluis, C.; et al. Adenosine A2A and dopamine D2 heteromeric receptor complexes and their function. J. Mol. Neurosci. 2005, 26, 209–220. [Google Scholar] [CrossRef]
- Torvinen, M.; Marcellino, D.; Canals, M.; Agnati, L.F.; Lluis, C.; Franco, R.; Fuxe, K. Adenosine A2A receptor and dopamine D3 receptor interactions: Evidence of functional A2A/D3 heteromeric complexes. Mol. Pharmacol. 2005, 67, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Cuellar, B.; Giron, F.J.; Grandy, D.K.; de la Calle, A.; Moratalla, R. Dopamine D4 receptors are heterogeneously distributed in the striosomes/matrix compartments of the striatum. J. Neurochem. 2002, 80, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferre, S.; Quiroz, C.; Woods, A.S.; Cunha, R.; Popoli, P.; Ciruela, F.; Lluis, C.; Franco, R.; Azdad, K.; Schiffmann, S.N. An update on adenosine A2A-dopamine D2 receptor interactions: implications for the function of G protein-coupled receptors. Curr. Pharm. Des. 2008, 14, 1468–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferre, S.; Quiroz, C.; Orru, M.; Guitart, X.; Navarro, G.; Cortes, A.; Casado, V.; Canela, E.I.; Lluis, C.; Franco, R. Adenosine A(2A) Receptors and A(2A) Receptor Heteromers as Key Players in Striatal Function. Front. Neuroanat. 2011, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuxe, K.; Marcellino, D.; Borroto-Escuela, D.O.; Guescini, M.; Fernandez-Duenas, V.; Tanganelli, S.; Rivera, A.; Ciruela, F.; Agnati, L.F. Adenosine-dopamine interactions in the pathophysiology and treatment of CNS disorders. CNS Neurosci. Ther. 2010, 16, e18–e42. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, A.; de Iure, A.; Di Filippo, M.; Tantucci, M.; Costa, C.; Borsini, F.; Ghiglieri, V.; Giampà, C.; Fusco, F.R.; Picconi, B.; et al. The Distinct Role of Medium Spiny Neurons and Cholinergic Interneurons in the D2/A2AReceptor Interaction in the Striatum: Implications for Parkinson’s Disease. J. Neurosci. 2011, 31, 1850. [Google Scholar] [CrossRef] [Green Version]
- Cervetto, C.; Venturini, A.; Passalacqua, M.; Guidolin, D.; Genedani, S.; Fuxe, K.; Borroto-Esquela, D.O.; Cortelli, P.; Woods, A.; Maura, G.; et al. A2A-D2 receptor-receptor interaction modulates gliotransmitter release from striatal astrocyte processes. J. Neurochem. 2017, 140, 268–279. [Google Scholar] [CrossRef]
- Cervetto, C.; Venturini, A.; Guidolin, D.; Maura, G.; Passalacqua, M.; Tacchetti, C.; Cortelli, P.; Genedani, S.; Candiani, S.; Ramoino, P.; et al. Homocysteine and A2A-D2 Receptor-Receptor Interaction at Striatal Astrocyte Processes. J. Mol. Neurosci. 2018, 65, 456–466. [Google Scholar] [CrossRef]
- Trifilieff, P.; Rives, M.L.; Urizar, E.; Piskorowski, R.A.; Vishwasrao, H.D.; Castrillon, J.; Schmauss, C.; Slättman, M.; Gullberg, M.; Javitch, J.A. Detection of antigen interactions ex vivo by proximity ligation assay: endogenous dopamine D2-adenosine A2A receptor complexes in the striatum. Biotechniques 2011, 51, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Garriga, P.; Ciruela, F.; Narvaez, M.; Tarakanov, A.O.; Palkovits, M.; Agnati, L.F.; Fuxe, K. G protein-coupled receptor heterodimerization in the brain. Methods Enzymol. 2013, 521, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Tarakanov, A.O.; Fuxe, K.G. Triplet puzzle: Homologies of receptor heteromers. J. Mol. Neurosci. 2010, 41, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Pinton, L.; O Borroto-Escuela, D.; Narváez, M.; Oflijan, J.; F Agnati, L.; Fuxe, K. Evidence for the existence of dopamine D2R and Sigma 1 allosteric receptor-receptor interaction in the rat brain: Role in brain plasticity and cocaine action. SpringerPlus 2015, 4, P37. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, S.M.; Pollack, A.E.; Schusheim, L.; Fink, J.S. Effects of selective adenosine A1 and A2a agonists on amphetamine-induced locomotion and c-Fos in striatum and nucleus accumbens. Brain Res. 1996, 707, 75–80. [Google Scholar] [CrossRef]
- Rimondini, R.; Ferre, S.; Ogren, S.O.; Fuxe, K. Adenosine A2A agonists: A potential new type of atypical antipsychotic. Neuropsychopharmacology 1997, 17, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, K.A.; Schreiner, D.C.; Levis, S.C.; O’Neill, C.E.; Bachtell, R.K. Role of adenosine receptor subtypes in methamphetamine reward and reinforcement. Neuropharmacology 2015, 89, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poleszak, E.; Malec, D. Cocaine-induced hyperactivity is more influenced by adenosine receptor agonists than amphetamine-induced hyperactivity. Pol. J. Pharmacol. 2002, 54, 359–366. [Google Scholar] [PubMed]
- Filip, M.; Frankowska, M.; Zaniewska, M.; Przegalinski, E.; Muller, C.E.; Agnati, L.; Franco, R.; Roberts, D.C.; Fuxe, K. Involvement of adenosine A2A and dopamine receptors in the locomotor and sensitizing effects of cocaine. Brain Res. 2006, 1077, 67–80. [Google Scholar] [CrossRef]
- Czyzyk, J.; Zaniewska, M.; Bader, M.; Fuxe, K.; Filip, M. Attenuated behavioral responses to acute and chronic cocaine in adenosine A2A overexpressed transgenic rats. In European Neuropsychopharmacology. Proceedings of the The 24th ECNP Congress, Paris, France, 3–7 September 2011; ECNP: Utrecht, The Netherlands; pp. S597–S598.
- Poleszak, E.; Malec, D. Adenosine receptor ligands and cocaine in conditioned place preference (CPP) test in rats. Pol. J. Pharmacol. 2002, 54, 119–126. [Google Scholar]
- Haynes, N.S.; O’Neill, C.E.; Hobson, B.D.; Bachtell, R.K. Effects of adenosine A2A receptor antagonists on cocaine-induced locomotion and cocaine seeking. Psychopharmacology (Berl). 2019, 236, 699–708. [Google Scholar] [CrossRef]
- Chen, J.F.; Beilstein, M.; Xu, Y.H.; Turner, T.J.; Moratalla, R.; Standaert, D.G.; Aloyo, V.J.; Fink, J.S.; Schwarzschild, M.A. Selective attenuation of psychostimulant-induced behavioral responses in mice lacking A(2A) adenosine receptors. Neuroscience 2000, 97, 195–204. [Google Scholar] [CrossRef]
- Wright, S.R.; Zanos, P.; Georgiou, P.; Yoo, J.H.; Ledent, C.; Hourani, S.M.; Kitchen, I.; Winsky-Sommerer, R.; Bailey, A. A critical role of striatal A2A R-mGlu5 R interactions in modulating the psychomotor and drug-seeking effects of methamphetamine. Addict. Biol. 2016, 21, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.Y.; Coelho, J.E.; Ohtsuka, N.; Canas, P.M.; Day, Y.J.; Huang, Q.Y.; Rebola, N.; Yu, L.; Boison, D.; Cunha, R.A.; et al. A critical role of the adenosine A2A receptor in extrastriatal neurons in modulating psychomotor activity as revealed by opposite phenotypes of striatum and forebrain A2A receptor knock-outs. J. Neurosci. 2008, 28, 2970–2975. [Google Scholar] [CrossRef]
- Shimazoe, T.; Yoshimatsu, A.; Kawashimo, A.; Watanabe, S. Roles of adenosine A(1) and A(2A) receptors in the expression and development of methamphetamine-induced sensitization. Eur. J. Pharmacol. 2000, 388, 249–254. [Google Scholar] [CrossRef]
- Bastia, E.; Xu, Y.H.; Scibelli, A.C.; Day, Y.J.; Linden, J.; Chen, J.F.; Schwarzschild, M.A. A crucial role for forebrain adenosine A(2A) receptors in amphetamine sensitization. Neuropsychopharmacology 2005, 30, 891–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.F.; Moratalla, R.; Yu, L.; Martin, A.B.; Xu, K.; Bastia, E.; Hackett, E.; Alberti, I.; Schwarzschild, M.A. Inactivation of adenosine A2A receptors selectively attenuates amphetamine-induced behavioral sensitization. Neuropsychopharmacology 2003, 28, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Soria, G.; Castane, A.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. The lack of A2A adenosine receptors diminishes the reinforcing efficacy of cocaine. Neuropsychopharmacology 2006, 31, 978–987. [Google Scholar] [CrossRef]
- Listos, J.; Talarek, S.; Fidecka, S. Involvement of adenosine receptor agonists on the development of hypersensitivity to acute dose of morphine during morphine withdrawal period. Pharmacol. Rep. 2008, 60, 679–685. [Google Scholar]
- Listos, J.; Talarek, S.; Poleszak, E.; Wrobel, A.; Fidecka, S. Attenuating effect of adenosine receptor agonists on the development of behavioral sensitization induced by sporadic treatment with morphine. Pharmacol. Biochem. Behav. 2011, 98, 356–361. [Google Scholar] [CrossRef]
- Castane, A.; Wells, L.; Soria, G.; Hourani, S.; Ledent, C.; Kitchen, I.; Opacka-Juffry, J.; Maldonado, R.; Valverde, O. Behavioural and biochemical responses to morphine associated with its motivational properties are altered in adenosine A(2A) receptor knockout mice. Br. J. Pharmacol. 2008, 155, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Soria, G.; Castane, A.; Berrendero, F.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Adenosine A2A receptors are involved in physical dependence and place conditioning induced by THC. Eur. J. Neurosci. 2004, 20, 2203–2213. [Google Scholar] [CrossRef]
- Fritz, B.M.; Boehm, S.L. 2nd. Adenosinergic regulation of binge-like ethanol drinking and associated locomotor effects in male C57BL/6J mice. Pharmacol. Biochem. Behav. 2015, 135, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houchi, H.; Warnault, V.; Barbier, E.; Dubois, C.; Pierrefiche, O.; Ledent, C.; Daoust, M.; Naassila, M. Involvement of A2A receptors in anxiolytic, locomotor and motivational properties of ethanol in mice. Genes Brain Behav. 2008, 7, 887–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jastrzębska, J.; Nowak, E.; Smaga, I.; Bystrowska, B.; Frankowska, M.; Bader, M.; Filip, M.; Fuxe, K. Adenosine (A)(2A)receptor modulation of nicotine-induced locomotor sensitization. A pharmacological and transgenic approach. Neuropharmacology 2014, 81, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Castane, A.; Soria, G.; Ledent, C.; Maldonado, R.; Valverde, O. Attenuation of nicotine-induced rewarding effects in A2A knockout mice. Neuropharmacology 2006, 51, 631–640. [Google Scholar] [CrossRef]
- Justinova, Z.; Ferre, S.; Segal, P.N.; Antoniou, K.; Solinas, M.; Pappas, L.A.; Highkin, J.L.; Hockemeyer, J.; Munzar, P.; Goldberg, S.R. Involvement of adenosine A1 and A2A receptors in the adenosinergic modulation of the discriminative-stimulus effects of cocaine and methamphetamine in rats. J. Pharmacol. Exp. Ther. 2003, 307, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Justinova, Z.; Ferre, S.; Barnes, C.; Wertheim, C.E.; Pappas, L.A.; Goldberg, S.R.; Le Foll, B. Effects of chronic caffeine exposure on adenosinergic modulation of the discriminative-stimulus effects of nicotine, methamphetamine, and cocaine in rats. Psychopharmacology (Berl). 2009, 203, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Munzar, P.; Justinova, Z.; Kutkat, S.W.; Ferre, S.; Goldberg, S.R. Adenosinergic modulation of the discriminative-stimulus effects of methamphetamine in rats. Psychopharmacol. Berl. 2002, 161, 348–355. [Google Scholar] [CrossRef]
- Knapp, C.M.; Foye, M.M.; Cottam, N.; Ciraulo, D.A.; Kornetsky, C. Adenosine agonists CGS 21680 and NECA inhibit the initiation of cocaine self-administration. Pharmacol. Biochem. Behav. 2001, 68, 797–803. [Google Scholar] [CrossRef]
- Wydra, K.; Golembiowska, K.; Suder, A.; Kaminska, K.; Fuxe, K.; Filip, M. On the role of adenosine (A)(2)A receptors in cocaine-induced reward: A pharmacological and neurochemical analysis in rats. Psychopharmacology (Berl). 2015, 232, 421–435. [Google Scholar] [CrossRef]
- Wydra, K.; Suder, A.; Frankowska, M.; Borroto Escuela, D.O.; Fuxe, K.; Filip, M. Effects of intra-accumbal or intra-prefrontal cortex microinjections of adenosine 2A receptor ligands on responses to cocaine reward and seeking in rats. Psychopharmacology (Berl). 2018, 235, 3509–3523. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Wydra, K.; Romero-Fernandez, W.; Zhou, Z.; Frankowska, M.; Filip, M.; Fuxe, K. A2AR Transmembrane 2 Peptide Administration Disrupts the A2AR-A2AR Homoreceptor but Not the A2AR-D2R Heteroreceptor Complex: Lack of Actions on Rodent Cocaine Self-Administration. Int. J. Mol. Sci. 2019, 20, 6100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borroto-Escuela, D.O.; Wydra, K.; Li, X.; Rodriguez, D.; Carlsson, J.; Jastrzebska, J.; Filip, M.; Fuxe, K. Disruption of A2AR-D2R Heteroreceptor Complexes After A2AR Transmembrane 5 Peptide Administration Enhances Cocaine Self-Administration in Rats. Mol. Neurobiol. 2018, 55, 7038–7048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poleszak, E.; Malec, D. Effects of adenosine receptor agonists and antagonists in amphetamine-induced conditioned place preference test in rats. Pol. J. Pharmacol. 2003, 55, 319–326. [Google Scholar] [PubMed]
- Justinova, Z.; Ferre, S.; Redhi, G.H.; Mascia, P.; Stroik, J.; Quarta, D.; Yasar, S.; Muller, C.E.; Franco, R.; Goldberg, S.R. Reinforcing and neurochemical effects of cannabinoid CB1 receptor agonists, but not cocaine, are altered by an adenosine A2A receptor antagonist. Addict. Biol. 2011, 16, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Medina, J.; Ledent, C.; Carreton, O.; Valverde, O. The A2a adenosine receptor modulates the reinforcement efficacy and neurotoxicity of MDMA. J. Psychopharmacol. 2011, 25, 550–564. [Google Scholar] [CrossRef]
- Gill, W.D.; Shelton, H.W.; Burgess, K.C.; Brown, R.W. Effects of an adenosine A2A agonist on the rewarding associative properties of nicotine and neural plasticity in a rodent model of schizophrenia. J. Psychopharmacol. 2020, 34, 137–144. [Google Scholar] [CrossRef]
- Sahraei, H.; Motamedi, F.; Khoshbaten, A.; Zarrindast, M.R. Adenosine A(2) receptors inhibit morphine self-administration in rats. Eur. J. Pharmacol. 1999, 383, 107–113. [Google Scholar] [CrossRef]
- Listos, J.; Talarek, S.; Listos, P.; Orzelska, J.; Lupina, M.; Fidecka, S. Effects of the adenosinergic system on the expression and acquisition of sensitization to conditioned place preference in morphine-conditioned rats. Naunyn Schmiedebergs Arch. Pharmacol. 2016, 389, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.M.; Short, J.L.; Cowen, M.S.; Ledent, C.; Lawrence, A.J. A differential role for the adenosine A2A receptor in opiate reinforcement vs opiate-seeking behavior. Neuropsychopharmacology 2009, 34, 844–856. [Google Scholar] [CrossRef] [Green Version]
- Houchi, H.; Persyn, W.; Legastelois, R.; Naassila, M. The adenosine A2A receptor agonist CGS 21680 decreases ethanol self-administration in both non-dependent and dependent animals. Addict. Biol. 2013, 18, 812–825. [Google Scholar] [CrossRef]
- Zaniewska, M.; Gawlinski, D.; Wyczesana, M.; Nowak, E.; Kula, K.; Maciow-Glab, M.; Jastrzebska, J.; Sadakierska-Chudy, A.; Bader, M.; Fuxe, K. Increased Ethanol Consumption and Locomotion Develop upon Ethanol Deprivation in Rats Overexpressing the Adenosine (A)2A Receptor. Neuroscience 2019, 418, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Arolfo, M.P.; Yao, L.; Gordon, A.S.; Diamond, I.; Janak, P.H. Ethanol operant self-administration in rats is regulated by adenosine A2 receptors. Alcohol. Clin. Exp. Res. 2004, 28, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Thorsell, A.; Johnson, J.; Heilig, M. Effect of the adenosine A2a receptor antagonist 3,7-dimethyl-propargylxanthine on anxiety-like and depression-like behavior and alcohol consumption in Wistar Rats. Alcohol. Clin. Exp. Res. 2007, 31, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.L.; Cowen, M.S.; Short, J.L.; Lawrence, A.J. Combined antagonism of glutamate mGlu5 and adenosine A2A receptors interact to regulate alcohol-seeking in rats. Int. J. Neuropsychopharmacol. 2008, 11, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Nam, H.W.; Hinton, D.J.; Kang, N.Y.; Kim, T.; Lee, M.R.; Oliveros, A.; Adams, C.; Ruby, C.L.; Choi, D.S. Adenosine transporter ENT1 regulates the acquisition of goal-directed behavior and ethanol drinking through A2A receptor in the dorsomedial striatum. J. Neurosci. 2013, 33, 4329–4338. [Google Scholar] [CrossRef] [Green Version]
- Naassila, M.; Ledent, C.; Daoust, M. Low ethanol sensitivity and increased ethanol consumption in mice lacking adenosine A2A receptors. J. Neurosci. 2002, 22, 10487–10493. [Google Scholar] [CrossRef] [Green Version]
- Mukhara, D.; Banks, M.L.; Neigh, G.N. Stress as a Risk Factor for Substance Use Disorders: A Mini-Review of Molecular Mediators. Front. Behav. Neurosci. 2018, 12, 309. [Google Scholar] [CrossRef] [Green Version]
- Bachtell, R.K.; Self, D.W. Effects of adenosine A2A receptor stimulation on cocaine-seeking behavior in rats. Psychopharmacol. Berl. 2009, 206, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Wydra, K.; Suder, A.; Borroto-Escuela, D.O.; Filip, M.; Fuxe, K. On the role of A(2)A and D(2) receptors in control of cocaine and food-seeking behaviors in rats. Psychopharmacol. Berl. 2015, 232, 1767–1778. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, C.E.; LeTendre, M.L.; Bachtell, R.K. Adenosine A2A receptors in the nucleus accumbens bi-directionally alter cocaine seeking in rats. Neuropsychopharmacology 2012, 37, 1245–1256. [Google Scholar] [CrossRef] [Green Version]
- Larson, T.A.; Winkler, M.C.; Stafford, J.; Levis, S.C.; O’Neill, C.E.; Bachtell, R.K. Role of dopamine D2-like receptors and their modulation by adenosine receptor stimulation in the reinstatement of methamphetamine seeking. Psychopharmacol. Berl. 2019, 236, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Weerts, E.M.; Griffiths, R.R. The adenosine receptor antagonist CGS15943 reinstates cocaine-seeking behavior and maintains self-administration in baboons. Psychopharmacol. Berl. 2003, 168, 155–163. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.E.; Hobson, B.D.; Levis, S.C.; Bachtell, R.K. Persistent reduction of cocaine seeking by pharmacological manipulation of adenosine A1 and A 2A receptors during extinction training in rats. Psychopharmacol. Berl. 2014, 231, 3179–3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; McFarland, K.; Fan, P.; Jiang, Z.; Ueda, T.; Diamond, I. Adenosine A2a blockade prevents synergy between mu-opiate and cannabinoid CB1 receptors and eliminates heroin-seeking behavior in addicted rats. Proc. Natl. Acad. Sci. USA 2006, 103, 7877–7882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, B.A.; Koob, G.F.; Markou, A. Role of adenosine A2 receptors in brain stimulation reward under baseline conditions and during cocaine withdrawal in rats. J. Neurosci. 1999, 19, 11017–11026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dionyssopoulos, T.; Hope, W.; Coupar, I.M. Effect of adenosine analogues on the expression of opiate withdrawal in rats. Pharmacol. Biochem. Behav. 1992, 42, 201–206. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Sears, M.T. Adenosine receptor agonists attenuate and adenosine receptor antagonists exacerbate opiate withdrawal signs. Psychopharmacol. Berl. 1996, 123, 64–70. [Google Scholar] [CrossRef]
- Salem, A.; Hope, W. Effect of adenosine receptor agonists and antagonists on the expression of opiate withdrawal in rats. Pharmacol. Biochem. Behav. 1997, 57, 671–679. [Google Scholar] [CrossRef]
- Bailey, A.; Davis, L.; Lesscher, H.M.; Kelly, M.D.; Ledent, C.; Hourani, S.M.; Kitchen, I. Enhanced morphine withdrawal and micro -opioid receptor G-protein coupling in A2A adenosine receptor knockout mice. J. Neurochem. 2004, 88, 827–834. [Google Scholar] [CrossRef]
- Bilbao, A.; Cippitelli, A.; Martin, A.B.; Granado, N.; Ortiz, O.; Bezard, E.; Chen, J.F.; Navarro, M.; Rodriguez de Fonseca, F.; Moratalla, R. Absence of quasi-morphine withdrawal syndrome in adenosine A2A receptor knockout mice. Psychopharmacol. Berl. 2006, 185, 160–168. [Google Scholar] [CrossRef]
- Berrendero, F.; Castane, A.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Increase of morphine withdrawal in mice lacking A2a receptors and no changes in CB1/A2a double knockout mice. Eur. J. Neurosci. 2003, 17, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.B.; Bharmal, N.H.; Leite-Morris, K.A.; Adams, W.R. Role of adenosine A1 and A2A receptors in the alcohol withdrawal syndrome. Alcohol 1999, 19, 157–162. [Google Scholar] [CrossRef]
- Prediger, R.D.; da Silva, G.E.; Batista, L.C.; Bittencourt, A.L.; Takahashi, R.N. Activation of adenosine A1 receptors reduces anxiety-like behavior during acute ethanol withdrawal (hangover) in mice. Neuropsychopharmacology 2006, 31, 2210–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Yacoubi, M.; Ledent, C.; Parmentier, M.; Daoust, M.; Costentin, J.; Vaugeois, J. Absence of the adenosine A(2A) receptor or its chronic blockade decrease ethanol withdrawal-induced seizures in mice. Neuropharmacology 2001, 40, 424–432. [Google Scholar] [CrossRef]
- Bailey, A.; Gianotti, R.; Ho, A.; Kreek, M.J. Persistent upregulation of mu-opioid, but not adenosine, receptors in brains of long-term withdrawn escalating dose "binge" cocaine-treated rats. Synapse 2005, 57, 160–166. [Google Scholar] [CrossRef]
- Marcellino, D.; Roberts, D.C.; Navarro, G.; Filip, M.; Agnati, L.; Lluis, C.; Franco, R.; Fuxe, K. Increase in A2A receptors in the nucleus accumbens after extended cocaine self-administration and its disappearance after cocaine withdrawal. Brain Res. 2007, 1143, 208–220. [Google Scholar] [CrossRef]
- Frankowska, M.; Marcellino, D.; Adamczyk, P.; Filip, M.; Fuxe, K. Effects of cocaine self-administration and extinction on D2 -like and A2A receptor recognition and D2 -like/Gi protein coupling in rat striatum. Addict. Biol. 2013, 18, 455–466. [Google Scholar] [CrossRef]
- Yang, S.L.; Han, J.Y.; Kim, Y.B.; Nam, S.Y.; Song, S.; Hong, J.T.; Oh, K.W. Increased non-rapid eye movement sleep by cocaine withdrawal: possible involvement of A2A receptors. Arch. Pharm. Res. 2011, 34, 281–287. [Google Scholar] [CrossRef]
- Aso, E.; Fernández-Dueñas, V.; López-Cano, M.; Taura, J.; Watanabe, M.; Ferrer, I.; Luján, R.; Ciruela, F. Adenosine A2A-Cannabinoid CB1 Receptor Heteromers in the Hippocampus: Cannabidiol Blunts Δ9-Tetrahydrocannabinol-Induced Cognitive Impairment. Mol. Neurobiol. 2019, 56, 5382–5391. [Google Scholar] [CrossRef] [Green Version]
- Franco, R.; Reyes-Resina, I.; Aguinaga, D.; Lillo, A.; Jiménez, J.; Raïch, I.; Borroto-Escuela, D.O.; Ferreiro-Vera, C.; Canela, E.I.; Sánchez de Medina, V.; et al. Potentiation of cannabinoid signaling in microglia by adenosine A(2A) receptor antagonists. Glia 2019, 67, 2410–2423. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Leite-Morris, K.A.; Sears, M.T. Alterations of adenosine A1 receptors in morphine dependence. Brain Res. 1994, 657, 347–350. [Google Scholar] [CrossRef]
- De Montis, M.; Devoto, P.; Meloni, D.; Saba, P.; Tagliamonte, A. Deacresed adenosine A2A receptor function in morphine dependent rats. Pharmacol. Res. 1992, 25, 232–233. [Google Scholar] [CrossRef]
- Dar, M.S.; Mustafa, S.J.; Wooles, W.R. Possible role of adenosine in the CNS effects of ethanol. Life Sci. 1983, 33, 1363–1374. [Google Scholar] [CrossRef]
- Clark, M.; Dar, M.S. In vitro autoradiographic evidence for adenosine modulation of ethanol-induced motor disturbances in rats. Alcohol Alcohol. Suppl. 1991, 1, 203–206. [Google Scholar]
- Daly, J.W.; Shi, D.; Wong, V.; Nikodijevic, O. Chronic effects of ethanol on central adenosine function of mice. Brain Res. 1994, 650, 153–156. [Google Scholar] [CrossRef]
- Jarvis, M.F.; Becker, H.C. Single and repeated episodes of ethanol withdrawal increase adenosine A1, but not A2A, receptor density in mouse brain. Brain Res. 1998, 786, 80–88. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Wallman-Johansson, A. Effects of ethanol and acetate on adenosine production in rat hippocampal slices. Pharmacol. Toxicol. 1996, 79, 120–123. [Google Scholar] [CrossRef]
- Bolewska, P.; Martin, B.I.; Orlando, K.A.; Rhoads, D.E. Sequential Changes in Brain Glutamate and Adenosine A1 Receptors May Explain Severity of Adolescent Alcohol Withdrawal after Consumption of High Levels of Alcohol. Neurosci. J. 2019, 2019, 5950818. [Google Scholar] [CrossRef] [Green Version]
- Feltmann, K.; Borroto-Escuela, D.O.; Rüegg, J.; Pinton, L.; de Oliveira Sergio, T.; Narváez, M.; Jimenez-Beristain, A.; Ekström, T.J.; Fuxe, K.; Steensland, P. Effects of Long-Term Alcohol Drinking on the Dopamine D2 Receptor: Gene Expression and Heteroreceptor Complexes in the Striatum in Rats. Alcohol. Clin. Exp. Res. 2018, 42, 338–351. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, C.E.; Levis, S.C.; Schreiner, D.C.; Amat, J.; Maier, S.F.; Bachtell, R.K. Effects of adolescent caffeine consumption on cocaine sensitivity. Neuropsychopharmacology 2015, 40, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Fuxe, K.; Ferre, S.; Zoli, M.; Agnati, L.F. Integrated events in central dopamine transmission as analyzed at multiple levels. Evidence for intramembrane adenosine A2A/dopamine D2 and adenosine A1/dopamine D1 receptor interactions in the basal ganglia. Brain Res. Brain Res. Rev. 1998, 26, 258–273. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Marcellino, D.; Narvaez, M.; Flajolet, M.; Heintz, N.; Agnati, L.; Ciruela, F.; Fuxe, K. A serine point mutation in the adenosine A2AR C-terminal tail reduces receptor heteromerization and allosteric modulation of the dopamine D2R. Biochem. Biophys. Res. Commun. 2010, 394, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Narvaez, M.; Romero-Fernandez, W.; Pinton, L.; Wydra, K.; Filip, M.; Beggiato, S.; Tanganelli, S.; Ferraro, L.; Fuxe, K. Acute Cocaine Enhances Dopamine D2R Recognition and Signaling and Counteracts D2R Internalization in Sigma1R-D2R Heteroreceptor Complexes. Mol. Neurobiol. 2019, 56, 7045–7055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraro, L.; Beggiato, S.; Marcellino, D.; Frankowska, M.; Filip, M.; Agnati, L.F.; Antonelli, T.; Tomasini, M.C.; Tanganelli, S.; Fuxe, K. Nanomolar concentrations of cocaine enhance D2-like agonist-induced inhibition of the K+-evoked [3H]-dopamine efflux from rat striatal synaptosomes: a novel action of cocaine. J. Neural Transm. (Vienna) 2010, 117, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, L.; Frankowska, M.; Marcellino, D.; Zaniewska, M.; Beggiato, S.; Filip, M.; Tomasini, M.C.; Antonelli, T.; Tanganelli, S.; Fuxe, K. A novel mechanism of cocaine to enhance dopamine d2-like receptor mediated neurochemical and behavioral effects. An in vivo and in vitro study. Neuropsychopharmacology 2012, 37, 1856–1866. [Google Scholar] [CrossRef] [Green Version]
- Romero-Fernandez, W.; Zhou, Z.; Beggiato, S.; Wydra, K.; Filip, M.; Tanganelli, S.; Borroto-Escuela, D.O.; Ferraro, L.; Fuxe, K. Acute cocaine treatment enhances the antagonistic allosteric adenosine A2A-dopamine D2 receptor-receptor interactions in rat dorsal striatum without increasing significantly extracellular dopamine levels. Pharmacol. Rep. 2020, 72, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Wydra, K.; Golembiowska, K.; Zaniewska, M.; Kaminska, K.; Ferraro, L.; Fuxe, K.; Filip, M. Accumbal and pallidal dopamine, glutamate and GABA overflow during cocaine self-administration and its extinction in rats. Addict. Biol. 2013, 18, 307–324. [Google Scholar] [CrossRef]
- Pintsuk, J.; Borroto-Escuela, D.O.; Pomierny, B.; Wydra, K.; Zaniewska, M.; Filip, M.; Fuxe, K. Cocaine self-administration differentially affects allosteric A2A-D2 receptor-receptor interactions in the striatum. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 2016, 144, 85–91. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Wydra, K.; Pintsuk, J.; Narvaez, M.; Corrales, F.; Zaniewska, M.; Agnati, L.F.; Franco, R.; Tanganelli, S.; Ferraro, L.; et al. Understanding the Functional Plasticity in Neural Networks of the Basal Ganglia in Cocaine Use Disorder: A Role for Allosteric Receptor-Receptor Interactions in A2A-D2 Heteroreceptor Complexes. Neural Plast. 2016, 2016, 4827268. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Narvaez, M.; Wydra, K.; Pintsuk, J.; Pinton, L.; Jimenez-Beristain, A.; Di Palma, M.; Jastrzebska, J.; Filip, M.; Fuxe, K. Cocaine self-administration specifically increases A2AR-D2R and D2R-sigma1R heteroreceptor complexes in the rat nucleus accumbens shell. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 2017, 155, 24–31. [Google Scholar] [CrossRef]
- Romieu, P.; Meunier, J.; Garcia, D.; Zozime, N.; Martin-Fardon, R.; Bowen, W.D.; Maurice, T. The sigma1 (sigma1) receptor activation is a key step for the reactivation of cocaine conditioned place preference by drug priming. Psychopharmacol. Berl. 2004, 175, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Kourrich, S.; Su, T.P.; Fujimoto, M.; Bonci, A. The sigma-1 receptor: roles in neuronal plasticity and disease. Trends Neurosci. 2012, 35, 762–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, G.; Moreno, E.; Bonaventura, J.; Brugarolas, M.; Farre, D.; Aguinaga, D.; Mallol, J.; Cortes, A.; Casado, V.; Lluis, C.; et al. Cocaine inhibits dopamine D2 receptor signaling via sigma-1-D2 receptor heteromers. PLoS ONE 2013, 8, e61245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahlholm, K.; Arhem, P.; Fuxe, K.; Marcellino, D. The dopamine stabilizers ACR16 and (-)-OSU6162 display nanomolar affinities at the sigma-1 receptor. Mol. Psychiatry 2013, 18, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Wydra, K.; Zhou, Z.; Suder, A.; Filip, M.; Fuxe, K. OSU-6162, a Sigma1R Ligand in Low Doses, Can Further Increase the Effects of Cocaine Self-Administration on Accumbal D2R Heteroreceptor Complexes. Neurotox. Res. 2020, 37, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Wydra, K.; Filip, M.; Fuxe, K. A2AR-D2R Heteroreceptor Complexes in Cocaine Reward and Addiction. Trends Pharmacol. Sci. 2018, 39, 1008–1020. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Rodriguez, D.; Romero-Fernandez, W.; Kapla, J.; Jaiteh, M.; Ranganathan, A.; Lazarova, T.; Fuxe, K.; Carlsson, J. Mapping the Interface of a GPCR Dimer: A Structural Model of the A2A Adenosine and D2 Dopamine Receptor Heteromer. Front. Pharmacol. 2018, 9, 829. [Google Scholar] [CrossRef] [Green Version]
- Gimenez-Llort, L.; Schiffmann, S.N.; Shmidt, T.; Canela, L.; Camon, L.; Wassholm, M.; Canals, M.; Terasmaa, A.; Fernandez-Teruel, A.; Tobena, A.; et al. Working memory deficits in transgenic rats overexpressing human adenosine A2A receptors in the brain. Neurobiol. Learn Mem. 2007, 87, 42–56. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic diagram showing the neuronal and astrocytic localization of adenosine (A)2A homo-, heterodimer, and oligomer complexes with dopamine (D), cannabinoid (CB), and metabotropic glutamate receptors (mGlu) on glutamatergic input from the cortex and thalamus and dopaminergic (DA) input from the ventral tegmental area and both enkephalinstriato–pallidal and dynorfinstriato-nigral gamma-aminobutyric acid (GABA) neurons in the rat striatum. To simplify, cholinergic interneurons are not included.
Figure 1.
Schematic diagram showing the neuronal and astrocytic localization of adenosine (A)2A homo-, heterodimer, and oligomer complexes with dopamine (D), cannabinoid (CB), and metabotropic glutamate receptors (mGlu) on glutamatergic input from the cortex and thalamus and dopaminergic (DA) input from the ventral tegmental area and both enkephalinstriato–pallidal and dynorfinstriato-nigral gamma-aminobutyric acid (GABA) neurons in the rat striatum. To simplify, cholinergic interneurons are not included.

Figure 2.
The diagram showing in vivobehavioral effects of stimulation or blockade of the adenosine (A)2A receptors in the cycle of substance use disorders. Up and down arrows indicate an increase or decrease in behavioral responses, respectively. Double-sided horizontal arrows indicate mixed effects on behavioral responses. mGlu5—metabotropic glutamatergic receptors type 5; THC—tetrahydrocannabinol.
Figure 2.
The diagram showing in vivobehavioral effects of stimulation or blockade of the adenosine (A)2A receptors in the cycle of substance use disorders. Up and down arrows indicate an increase or decrease in behavioral responses, respectively. Double-sided horizontal arrows indicate mixed effects on behavioral responses. mGlu5—metabotropic glutamatergic receptors type 5; THC—tetrahydrocannabinol.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wydra, K.; Gawliński, D.; Gawlińska, K.; Frankowska, M.; Borroto-Escuela, D.O.; Fuxe, K.; Filip, M. Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine. Cells 2020, 9, 1372. https://doi.org/10.3390/cells9061372
AMA Style
Wydra K, Gawliński D, Gawlińska K, Frankowska M, Borroto-Escuela DO, Fuxe K, Filip M. Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine. Cells. 2020; 9(6):1372. https://doi.org/10.3390/cells9061372
Chicago/Turabian StyleWydra, Karolina, Dawid Gawliński, Kinga Gawlińska, Małgorzata Frankowska, Dasiel O. Borroto-Escuela, Kjell Fuxe, and Małgorzata Filip. 2020. "Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine" Cells 9, no. 6: 1372. https://doi.org/10.3390/cells9061372
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.