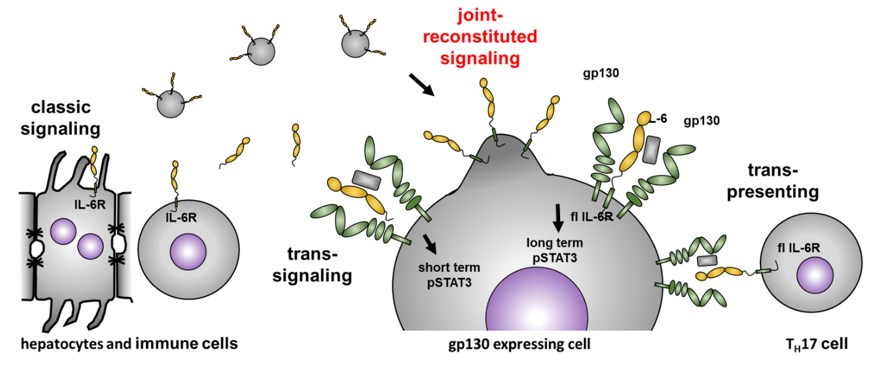
Joint Reconstituted Signaling of the IL-6 Receptor via Extracellular Vesicles

 , , , and
, , , and
Abstract
:
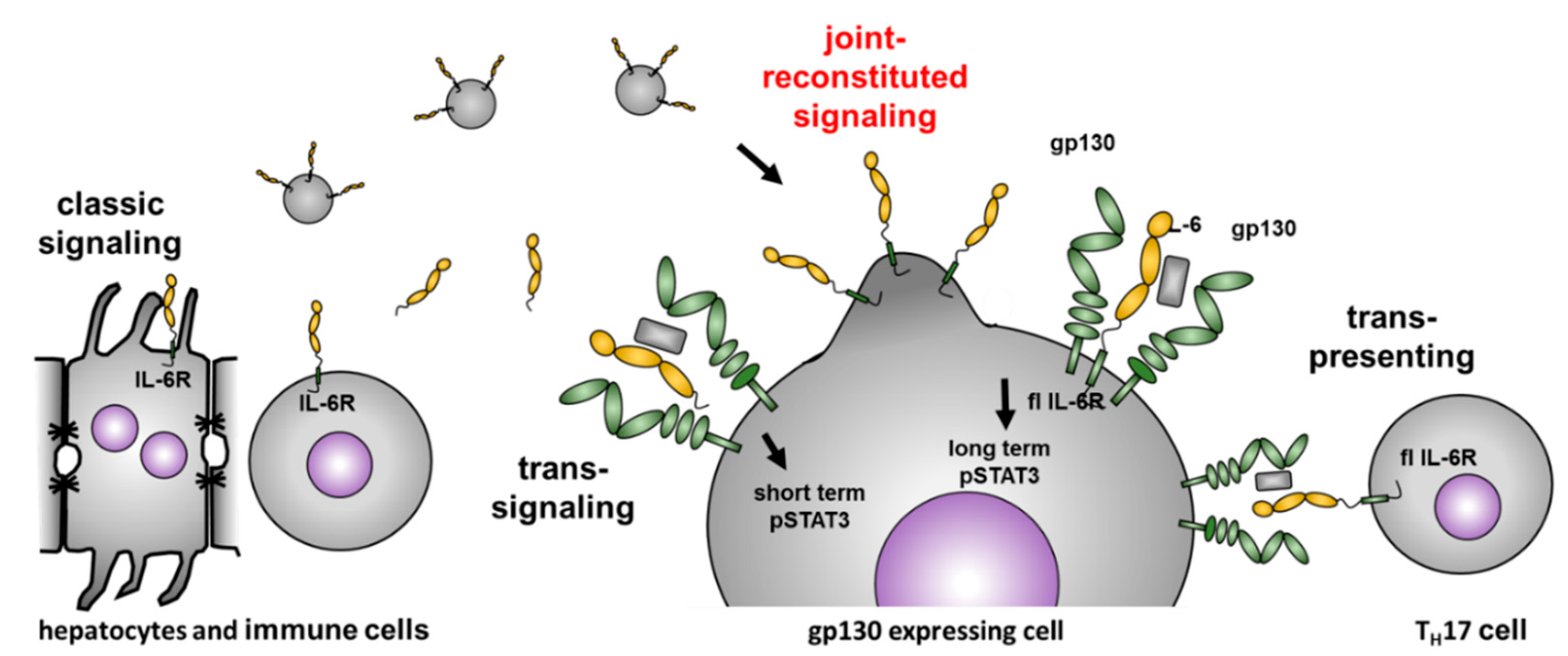
1. Introduction
2. Materials and Methods
2.1. Transient Cell Transfection, Cell Lysis, and Western Blot
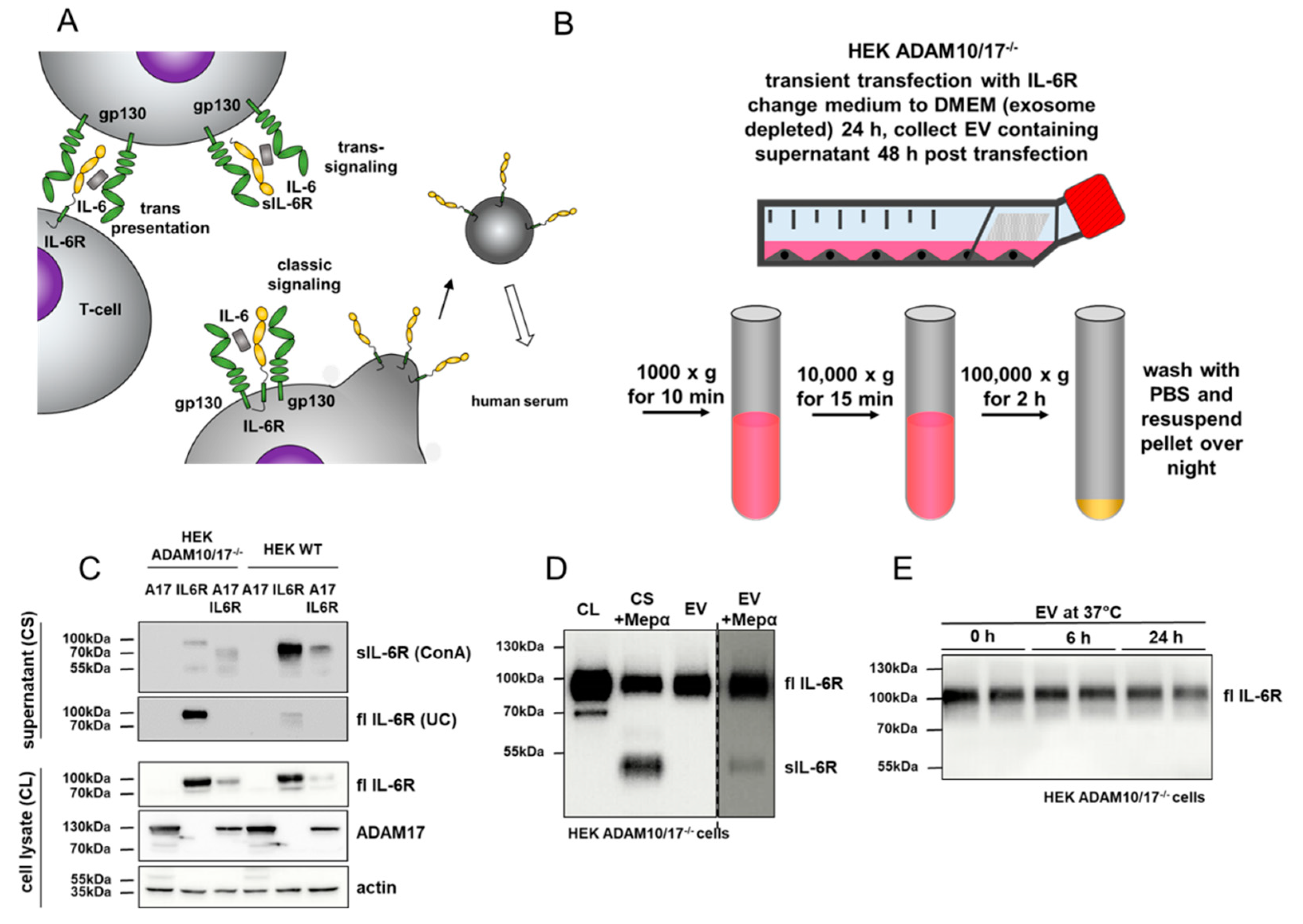
2.2. Purification of IL-6 Receptor Containing EVs From Cell Culture Supernatants
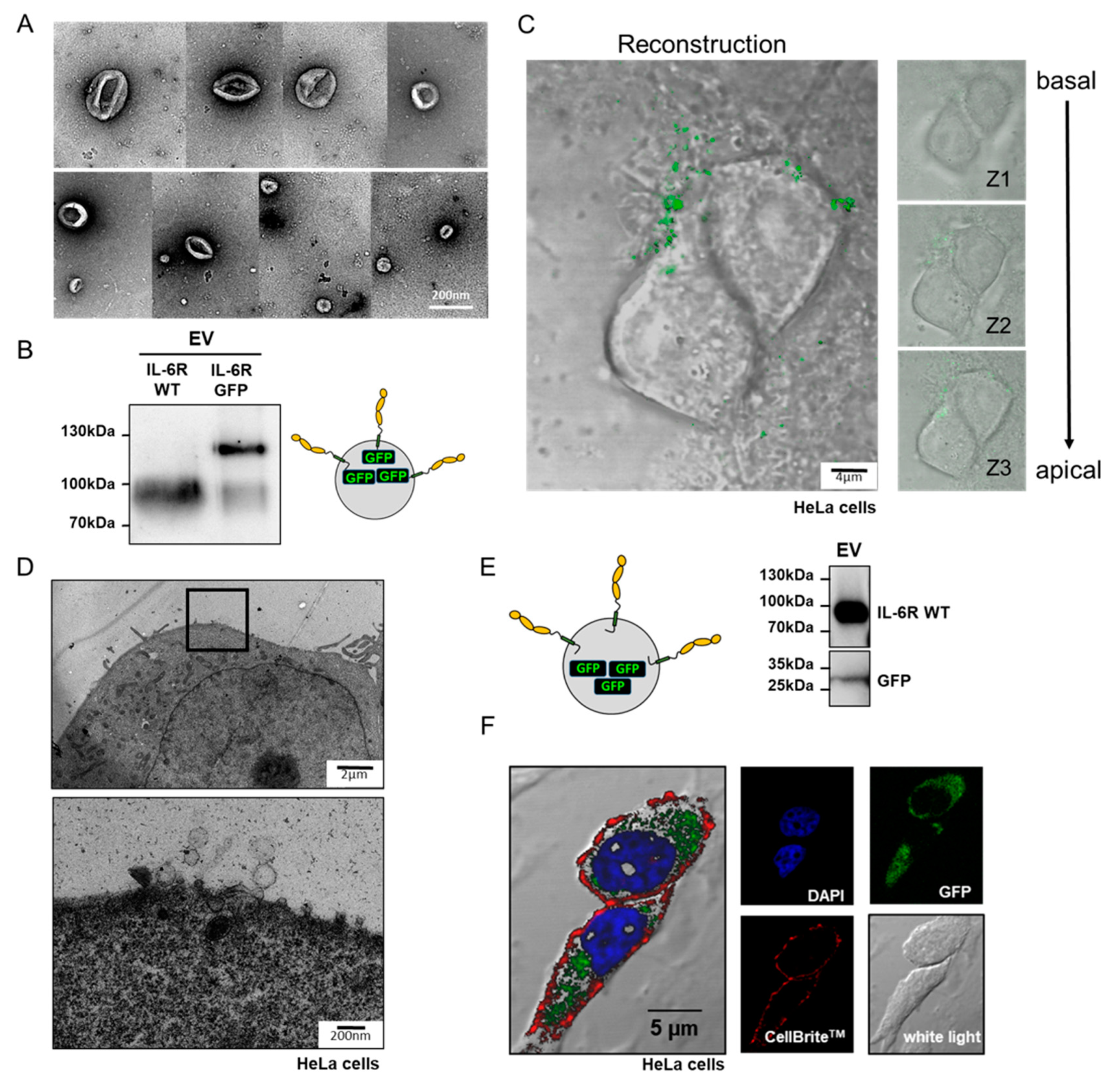
2.3. Transmission Electron Microscopy of EV and Cells
2.4. Confocal Laser Scanning Microscopy, z-Stacking, and Imaging
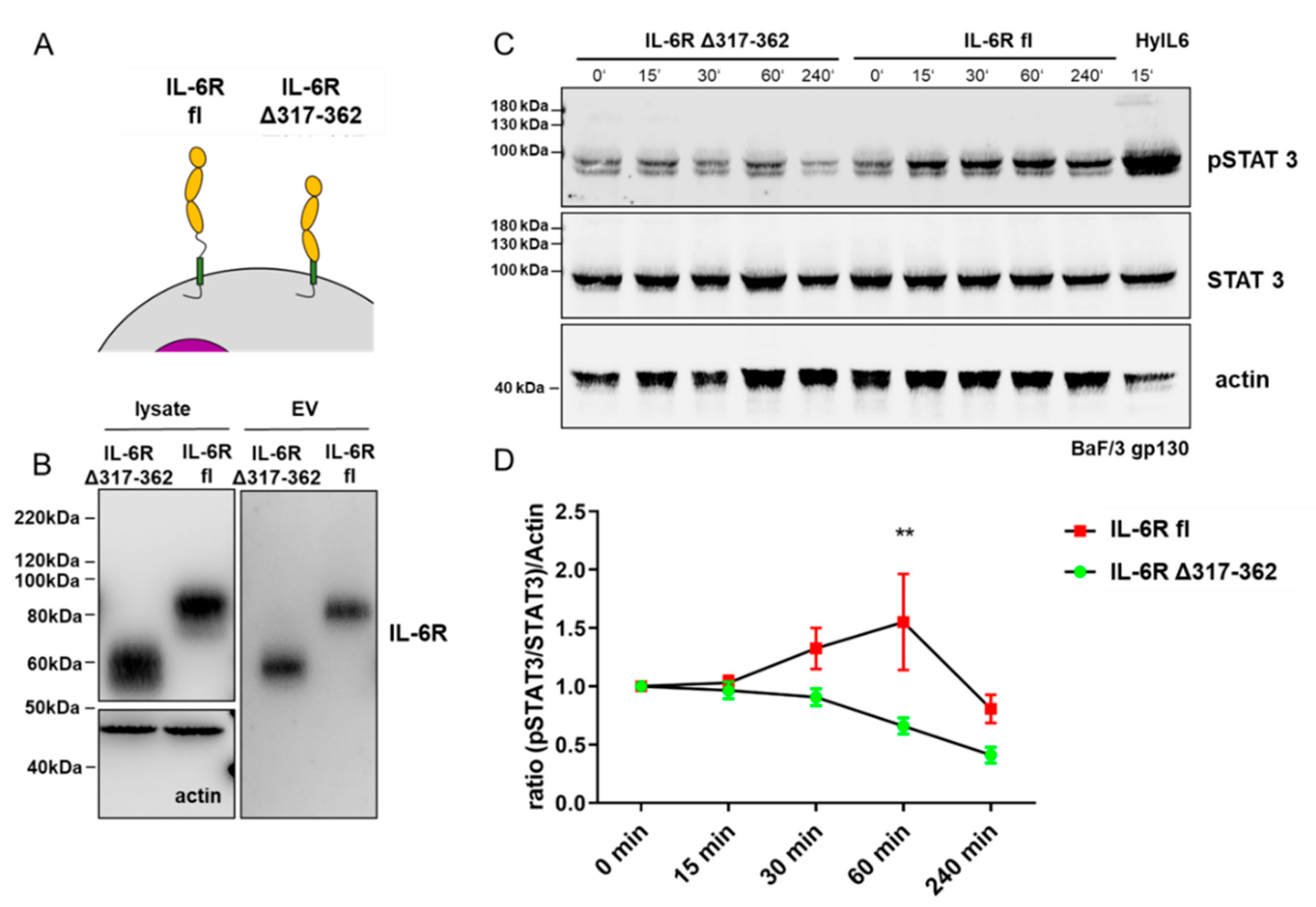
2.5. Ba/F3 gp130 Cell pSTAT3 Experiments
2.6. Statistical Analysis
3. Results
3.1. Full-Length IL-6 Receptor is Present on Extracellular Vesicles and Prone to Ectodomain Shedding
3.2. Isolated IL-6R Carrying Extracellular Vesicles Interact With the Plasma Membrane of Living Cells
3.3. Full-Length IL-6 Receptor on Extracellular Vesicles Induces Long-Term STAT3 Phosphorylation in Target Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smolen, J.S.; Schoels, M.M.; Nishimoto, N.; Breedveld, F.C.; Burmester, G.R.; Dougados, M.; Emery, P.; Ferraccioli, G.; Gabay, C.; Gibofsky, A.; et al. Consensus statement on blocking the effects of interleukin-6 and in particular by interleukin-6 receptor inhibition in rheumatoid arthritis and other inflammatory conditions. Ann. Rheum. Dis. 2012, 72, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, M.; Kamimura, D.; Hirano, T. Pleiotropy and Specificity: Insights from the Interleukin 6 Family of Cytokines. Immunology 2019, 50, 812–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbers, C.; Heink, S.; Korn, T.; Rose-John, S. Interleukin-6: Designing specific therapeutics for a complex cytokine. Nat. Rev. Drug Discov. 2018, 17, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Fu, E.; Lou, H.; Mao, X.; Yan, B.; Tong, F.; Sun, J.; Wei, L. IL-6 induced M1 type macrophage polarization increases radiosensitivity in HPV positive head and neck cancer. Cancer Lett. 2019, 456, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.-S.; Tseng, H.-Y.; Chen, Y.-A.; Shen, P.-C.; Al Haq, A.T.; Chen, L.-M.; Tung, Y.-C.; Hsu, H.-L. MCT-1/miR-34a/IL-6/IL-6R signaling axis promotes EMT progression, cancer stemness and M2 macrophage polarization in triple-negative breast cancer. Mol. Cancer 2019, 18, 42. [Google Scholar] [CrossRef] [Green Version]
- Peters, M.; Odenthal, M.; Schirmacher, P.; Blessing, M.; Fattori, E.; Ciliberto, G.; Buschenfelde, K.H.M.Z.; Rose-John, S. Soluble IL-6 receptor leads to a paracrine modulation of the IL-6-induced hepatic acute phase response in double transgenic mice. J. Immunol. 1997, 159, 1474–1481. [Google Scholar]
- Dai, J.; Lin, D.; Zhang, J.; Habib, P.; Smith, P.; Murtha, J.; Fu, Z.; Yao, Z.; Qi, Y.; Keller, E.T. Chronic alcohol ingestion induces osteoclastogenesis and bone loss through IL-6 in mice. J. Clin. Investig. 2000, 106, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Tamura, T.; Udagawa, N.; Takahashi, N.; Miyaura, C.; Tanaka, S.; Yamada, Y.; Koishihara, Y.; Ohsugi, Y.; Kumaki, K.; Taga, T. Soluble interleukin-6 receptor triggers osteoclast formation by interleukin 6. Proc. Natl. Acad. Sci. USA 1993, 90, 11924–11928. [Google Scholar] [CrossRef] [Green Version]
- Lane, T.; Gillmore, J.D.; Wechalekar, A.D.; Hawkins, P.N.; Lachmann, H.J. Therapeutic blockade of interleukin-6 by tocilizumab in the management of AA amyloidosis and chronic inflammatory disorders: A case series and review of the literature. Clin. Exp. Rheumatol. 2015, 33, 46–53. [Google Scholar]
- Puchner, A.; Blueml, S. IL-6 blockade in chronic inflammatory diseases. Wien. Med. Wochenschr. 2014, 165, 14–22. [Google Scholar] [CrossRef]
- Landskron, G.; De La Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic Inflammation and Cytokines in the Tumor Microenvironment. J. Immunol. Res. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotchkiss, R.S.; Moldawer, L.L.; Opal, S.M.; Reinhart, K.; Turnbull, I.R.; Vincent, J.-L. Sepsis and septic shock. Nat. Rev. Dis. Prim. 2016, 2, 16046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heink, S.; Yogev, N.; Garbers, C.; Herwerth, M.; Aly, L.; Gasperi, C.; Husterer, V.; Croxford, A.L.; Möller-Hackbarth, K.; Bartsch, H.S.; et al. Trans-presentation of IL-6 by dendritic cells is required for the priming of pathogenic TH17 cells. Nat. Immunol. 2016, 18, 74–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morra, D.E.; Pierson, S.K.; Shilling, D.; Nemat, S.; Appiani, C.; Guilfoyle, M.; Tendler, C.; Van Rhee, F.; Fajgenbaum, D.C. Predictors of response to anti-IL6 monoclonal antibody therapy (siltuximab) in idiopathic multicentric Castleman disease: Secondary analyses of phase II clinical trial data. Br. J. Haematol. 2019, 184, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luxembourger, C.; Ruyssen-Witrand, A.; Ladhari, C.; Rittore, C.; Degboé, Y.; Maillefert, J.-F.; Gaudin, P.; Marotte, H.; Wendling, D.; Jorgensen, C.; et al. A single nucleotide polymorphism of IL6-receptor is associated with response to tocilizumab in rheumatoid arthritis patients. Pharmacogenomics J. 2019, 19, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Genovese, M.C.; Van Der Heijde, D.; Lin, Y.; John, G.S.; Wang, S.; Van Hoogstraten, H.; Gómez-Reino, J.J.; Kivitz, A.; Maldonado-Cocco, J.A.; Seriolo, B.; et al. Long-term safety and efficacy of sarilumab plus methotrexate on disease activity, physical function and radiographic progression: 5 years of sarilumab plus methotrexate treatment. RMD Open 2019, 5, e000887. [Google Scholar] [CrossRef] [Green Version]
- Fleischmann, R.; Genovese, M.C.; Lin, Y.; John, G.S.; Van Der Heijde, D.; Wang, S.; Gomez-Reino, J.J.; Maldonado-Cocco, J.A.; Stanislav, M.; Kivitz, A.J.; et al. Long-term safety of sarilumab in rheumatoid arthritis: An integrated analysis with up to 7 years’ follow-up. Rheumatology 2019. [Google Scholar] [CrossRef] [Green Version]
- Kameda, H.; Wada, K.; Takahashi, Y.; Hagino, O.; Van Hoogstraten, H.; Graham, N.; Tanaka, Y. Sarilumab monotherapy or in combination with non-methotrexate disease-modifying antirheumatic drugs in active rheumatoid arthritis: A Japan phase 3 trial (HARUKA). Mod. Rheumatol. 2019, 30, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Rose-John, S. The Soluble Interleukin 6 Receptor: Advanced Therapeutic Options in Inflammation. Clin. Pharmacol. Ther. 2017, 102, 591–598. [Google Scholar] [CrossRef]
- Schumacher, N.; Schmidt, S.; Schwarz, J.; Dohr, D.; Lokau, J.; Scheller, J.; Garbers, C.; Chalaris, A.; Rose-John, S.; Rabe, B. Circulating Soluble IL-6R but Not ADAM17 Activation Drives Mononuclear Cell Migration in Tissue Inflammation. J. Immunol. 2016, 197, 3705–3715. [Google Scholar] [CrossRef]
- Schumacher, N.; Meyer, D.; Mauermann, A.; Von Der Heyde, J.; Wolf, J.; Schwarz, J.; Knittler, K.; Murphy, G.; Michalek, M.; Garbers, C.; et al. Shedding of Endogenous Interleukin-6 Receptor (IL-6R) Is Governed by A Disintegrin and Metalloproteinase (ADAM) Proteases while a Full-length IL-6R Isoform Localizes to Circulating Microvesicles. J. Biol. Chem. 2015, 290, 26059–26071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, P.; Boll, I.; Rothaug, M.; Schumacher, N.; Schmidt, F.; Wichert, R.; Schneppenheim, J.; Lokau, J.; Pickhinke, U.; Koudelka, T.; et al. Meprin Metalloproteases Generate Biologically Active Soluble Interleukin-6 Receptor to Induce Trans-Signaling. Sci. Rep. 2017, 7, 44053. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.; Schmidt, F.; Prox, J.; Zunke, F.; Pietrzik, C.; Lucius, R.; Becker-Pauly, C. Calcium negatively regulates meprin β activity and attenuates substrate cleavage. FASEB J. 2015, 29, 3549–3557. [Google Scholar] [CrossRef] [PubMed]
- Riethmueller, S.; Ehlers, J.C.; Lokau, J.; Düsterhöft, S.; Knittler, K.; Dombrowsky, G.; Grotzinger, J.; Rabe, B.; Rose-John, S.; Garbers, C. Cleavage Site Localization Differentially Controls Interleukin-6 Receptor Proteolysis by ADAM10 and ADAM17. Sci. Rep. 2016, 6, 25550. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.; Himmels, P.; Weiß, S.; Decker, T.-M.; Markl, J.; Gatterdam, V.; Tampé, R.; Bartholomäus, P.; Dietrich, U.; Duerr, R. Antigenic and 3D structural characterization of soluble X4 and hybrid X4-R5 HIV-1 Env trimers. Retrovirology 2014, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera–a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Jostock, T.; Müllberg, J.; Özbek, S.; Atreya, R.; Blinn, G.; Voltz, N.; Fischer, M.; Neurath, M.F.; Rose-John, S. Soluble gp130 is the natural inhibitor of soluble interleukin-6 receptor transsignaling responses. J. Biol. Inorg. Chem. 2001, 268, 160–167. [Google Scholar] [CrossRef]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef]
- Mülberg, J.; Schooltink, H.; Stoyan, T.; Günther, M.; Graeve, L.; Buse, J.B.; Mackiewicz, A.; Heinrich, P.C.; Rose-John, S. The soluble interleukin-6 receptor is generated by shedding. Eur. J. Immunol. 1993, 23, 473–480. [Google Scholar] [CrossRef]
- Riethmueller, S.; Somasundaram, P.; Ehlers, J.C.; Hung, C.-W.; Flynn, C.M.; Lokau, J.; Agthe, M.; Düsterhöft, S.; Zhu, Y.; Grötzinger, J.; et al. Proteolytic Origin of the Soluble Human IL-6R In Vivo and a Decisive Role of N-Glycosylation. PLoS Biol. 2017, 15, e2000080. [Google Scholar] [CrossRef]
- Schäffler, H.; Li, W.; Helm, O.; Krüger, S.; Böger, C.; Peters, F.; Röcken, C.; Sebens, S.; Lucius, R.; Becker-Pauly, C.; et al. The cancer-associated meprin β variant G32R provides an additional activation site and promotes cancer cell invasion. J. Cell Sci. 2019, 132, jcs220665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, S.; Pfeuffer, S.; Arnold, P.; Treitz, C.; Aden, K.; Ebsen, H.; Falk-Paulsen, M.; Gisch, N.; Fazio, A.; Kuiper, J.; et al. Prdx4 limits caspase-1 activation and restricts inflammasome-mediated signaling by extracellular vesicles. EMBO J. 2019, 38, e101266. [Google Scholar] [CrossRef] [PubMed]
- Van Der Pol, E.; Coumans, F.A.W.; Grootemaat, A.E.; Gardiner, C.; Sargent, I.L.; Harrison, P.; Sturk, A.; Van Leeuwen, T.G.; Nieuwland, R. Particle size distribution of exosomes and microvesicles determined by transmission electron microscopy, flow cytometry, nanoparticle tracking analysis, and resistive pulse sensing. J. Thromb. Haemost. 2014, 12, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Schaper, F.; Rose-John, S. Interleukin-6: Biology, signaling and strategies of blockade. Cytokine Growth Factor Rev. 2015, 26, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Lokau, J.; Garbers, C. Activating Mutations of the gp130/JAK/STAT Pathway in Human Diseases. In Advances in Protein Chemistry and Structural Biology; Academic Press: Waltham, MA, USA, 2019; Volume 116, pp. 283–309. [Google Scholar]
- Garbers, C.; Aparicio-Siegmund, S.; Rose-John, S. The IL-6/gp130/STAT3 signaling axis: Recent advances towards specific inhibition. Curr. Opin. Immunol. 2015, 34, 75–82. [Google Scholar] [CrossRef]
- Sansone, P.; Bromberg, J. Targeting the interleukin-6/Jak/stat pathway in human malignancies. J. Clin. Oncol. 2012, 30, 1005–1014. [Google Scholar] [CrossRef] [Green Version]
- Baran, P.; Nitz, R.; Grötzinger, J.; Scheller, J.; Garbers, C. Minimal Interleukin 6 (IL-6) Receptor Stalk Composition for IL-6 Receptor Shedding and IL-6 Classic Signaling. J. Biol. Chem. 2013, 288, 14756–14768. [Google Scholar] [CrossRef] [Green Version]
- Garbers, C.; Thaiss, W.; Jones, G.; Waetzig, G.H.; Lorenzen, I.; Guilhot, F.; Lissilaa, R.; Ferlin, W.G.; Grötzinger, J.; Jones, S.A.; et al. Inhibition of Classic Signaling Is a Novel Function of Soluble Glycoprotein 130 (sgp130), Which Is Controlled by the Ratio of Interleukin 6 and Soluble Interleukin 6 Receptor*. J. Biol. Chem. 2011, 286, 42959–42970. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.; Goldschmitt, J.; Peschel, C.; Brakenhoff, J.P.G.; Kallen, K.-J.; Wollmer, A.; Grötzinger, J.; Rose-John, S. A bioactive designer cytokine for human hematopoietic progenitor cell expansion. Nat. Biotechnol. 1997, 15, 142–145. [Google Scholar] [CrossRef]
- Billing, U.; Jetka, T.; Nortmann, L.; Wundrack, N.; Komorowski, M.; Waldherr, S.; Schaper, F.; Dittrich, A. Robustness and Information Transfer within IL-6-induced JAK/STAT Signalling. Commun. Biol. 2019, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Hurwitz, S.N.; Meckes, D. Extracellular Vesicle Integrins Distinguish Unique Cancers. Proteomes 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedele, C.; Singh, A.; Zerlanko, B.J.; Iozzo, R.V.; Languino, L.R. The αvβ6Integrin Is Transferred Intercellularly via Exosomes. J. Biol. Chem. 2015, 290, 4545–4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Mark, M.T.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, T.; Lv, H.; Lv, G.; Li, T.; Wang, C.; Han, Q.; Yu, L.; Su, B.; Guo, L.; Huang, S.; et al. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat. Commun. 2018, 9, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio-Siegmund, S.; Garbers, Y.; Flynn, C.M.; Waetzig, G.H.; Gouni-Berthold, I.; Krone, W.; Berthold, H.K.; Laudes, M.; Rose-John, S.; Garbers, C. The IL-6-neutralizing sIL-6R-sgp130 buffer system is disturbed in patients with type 2 diabetes. Am. J. Physiol. Metab. 2019, 317, E411–E420. [Google Scholar] [CrossRef]
- Groth, E.; Pruessmeyer, J.; Babendreyer, A.; Schumacher, J.; Pasqualon, T.; Dreymueller, D.; Higashiyama, S.; Lorenzen, I.; Grötzinger, J.; Cataldo, D.; et al. Stimulated release and functional activity of surface expressed metalloproteinase ADAM17 in exosomes. Biochim. Biophys. Acta 2016, 1863, 2795–2808. [Google Scholar] [CrossRef]
- Arenaccio, C.; Chiozzini, C.; Columba-Cabezas, S.; Manfredi, F.; Affabris, E.; Baur, A.; Federico, M. Exosomes from Human Immunodeficiency Virus Type 1 (HIV-1)-Infected Cells License Quiescent CD4+ T Lymphocytes To Replicate HIV-1 through a Nef- and ADAM17-Dependent Mechanism. J. Virol. 2014, 88, 11529–11539. [Google Scholar] [CrossRef] [Green Version]
- Padro, C.J.; Shawler, T.M.; Gormley, M.G.; Sanders, V.M. Adrenergic regulation of IgE involves modulation of CD23 and ADAM10 expression on exosomes. J. Immunol. 2013, 191, 5383–5397. [Google Scholar] [CrossRef] [Green Version]
- Mathews, J.A.; Gibb, D.R.; Chen, B.-H.; Scherle, P.; Conrad, D. CD23 Sheddase A Disintegrin and Metalloproteinase 10 (ADAM10) Is Also Required for CD23 Sorting into B Cell-derived Exosomes. J. Biol. Chem. 2010, 285, 37531–37541. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Host Species | Dilution | Retailer |
|---|---|---|---|
| α-IL-6 receptor (4/11) | Mouse | 1:1000 | In house |
| α-STAT3 (124H6) | Mouse | 1:1000 | Cell Signaling |
| α-pSTAT3 (D3A7) | Rabbit | 1:1000 | Cell Signaling |
| α-GFP (D5.1) | Rabbit | 1:1000 | Cell Signaling |
| α-actin | Rabbit | 1:5000 | Sigma-Aldrich |
| α-myc (ADAM17-myc) | Mouse | 1:1000 | Cell Signaling |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arnold, P.; Lückstädt, W.; Li, W.; Boll, I.; Lokau, J.; Garbers, C.; Lucius, R.; Rose-John, S.; Becker-Pauly, C. Joint Reconstituted Signaling of the IL-6 Receptor via Extracellular Vesicles. Cells 2020, 9, 1307. https://doi.org/10.3390/cells9051307
Arnold P, Lückstädt W, Li W, Boll I, Lokau J, Garbers C, Lucius R, Rose-John S, Becker-Pauly C. Joint Reconstituted Signaling of the IL-6 Receptor via Extracellular Vesicles. Cells. 2020; 9(5):1307. https://doi.org/10.3390/cells9051307
Chicago/Turabian StyleArnold, Philipp, Wiebke Lückstädt, Wenjia Li, Inga Boll, Juliane Lokau, Christoph Garbers, Ralph Lucius, Stefan Rose-John, and Christoph Becker-Pauly. 2020. "Joint Reconstituted Signaling of the IL-6 Receptor via Extracellular Vesicles" Cells 9, no. 5: 1307. https://doi.org/10.3390/cells9051307