The Influence of Palmatine Isolated from Berberis sibirica Radix on Pentylenetetrazole-Induced Seizures in Zebrafish

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Quantitative Structure–Activity Relationship (QSAR) Studies
2.2. Extraction
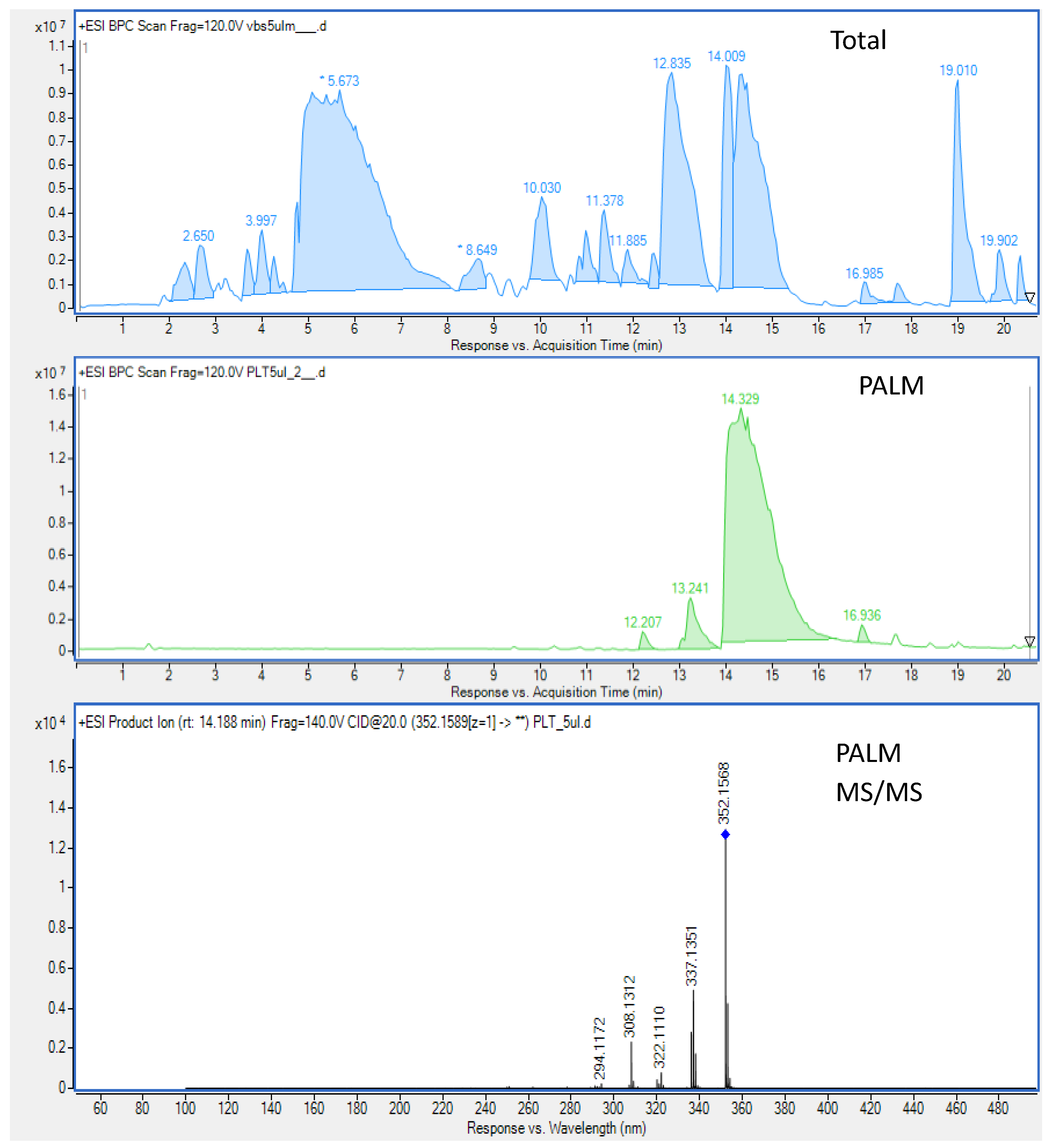
2.3. Chromatographic Analysis of the Extract by HPLC-ESI-Q-TOF-MS
2.4. Centrifugal Partition Chromatography (CPC)-Based Fractionation of the Extract
2.5. Drugs and Reagents
2.6. Zebrafish Maintenance and Breeding
2.7. EEG Analysis in Zebrafish
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
- gapdh_f_5′GTGGAGTCTACTGGTGTCTTC3′,
- gapdh_r_5′GTGCAGGAGGCATTGCTTACA3′;
- c-fos_f_5′CCGATACACTGCAAGCTGA 3′,
- c-fos_r_ 5′TGCGGCGAGGATGAACTCTA3′;
- bdnf_f_5′AGCTGAAGAGACAACTTGC3′,
- bdnf_r_5′CCATAGTAACGAACAGGAT3′.
2.9. Behavioral Analysis of Anticonvulsant Activity in Zebrafish
2.10. Statistical Analysis
3. Results
3.1. Quantitative Structure–Activity Relationship (QSAR) Studies for the BBB Permeation of PALM
n = 13, R2 = 73.59%, SE = 0.276
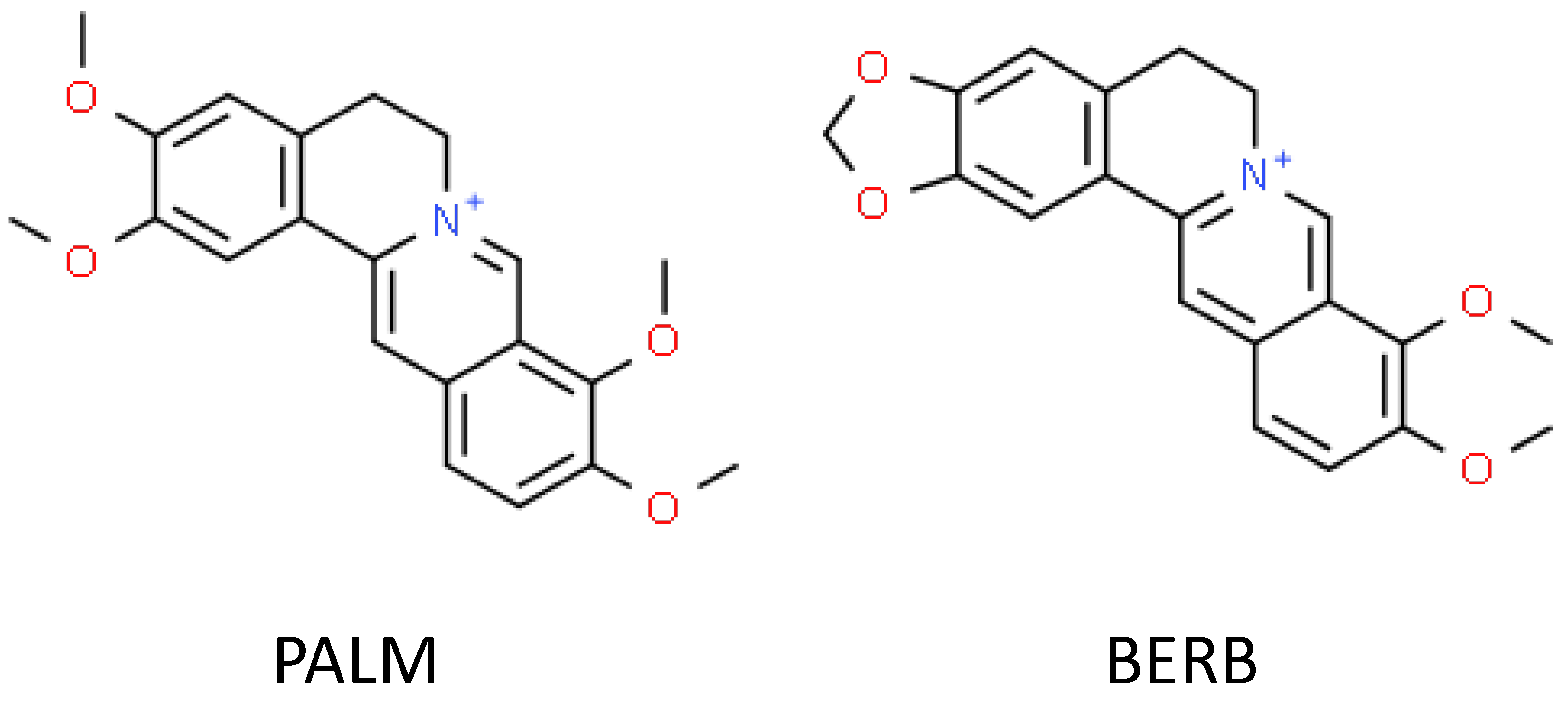
3.2. Isolation of PALM and BERB from Plant Material
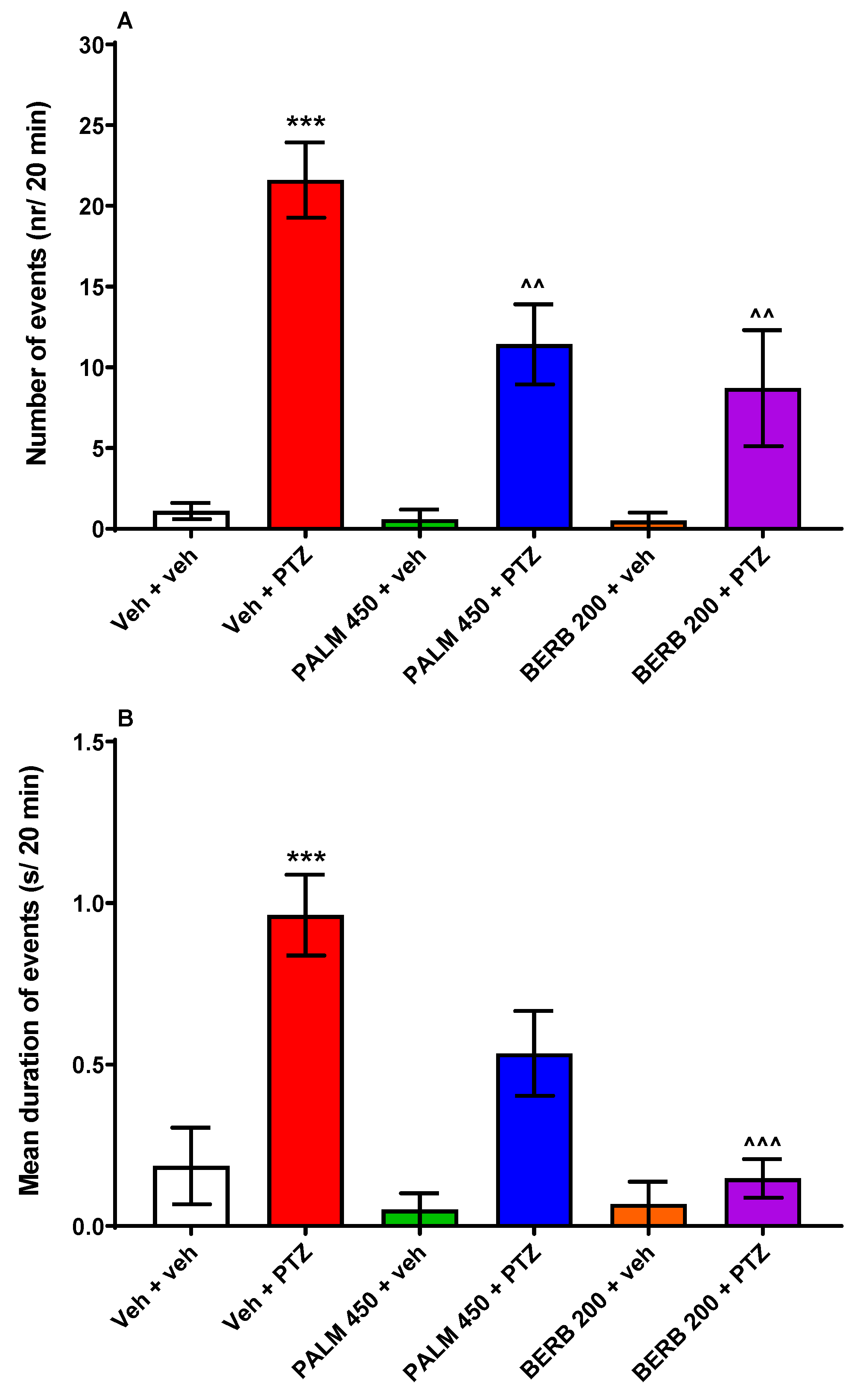
3.3. The Influence of PALM or BERB on PTZ-Induced EEG Discharges in Zebrafish
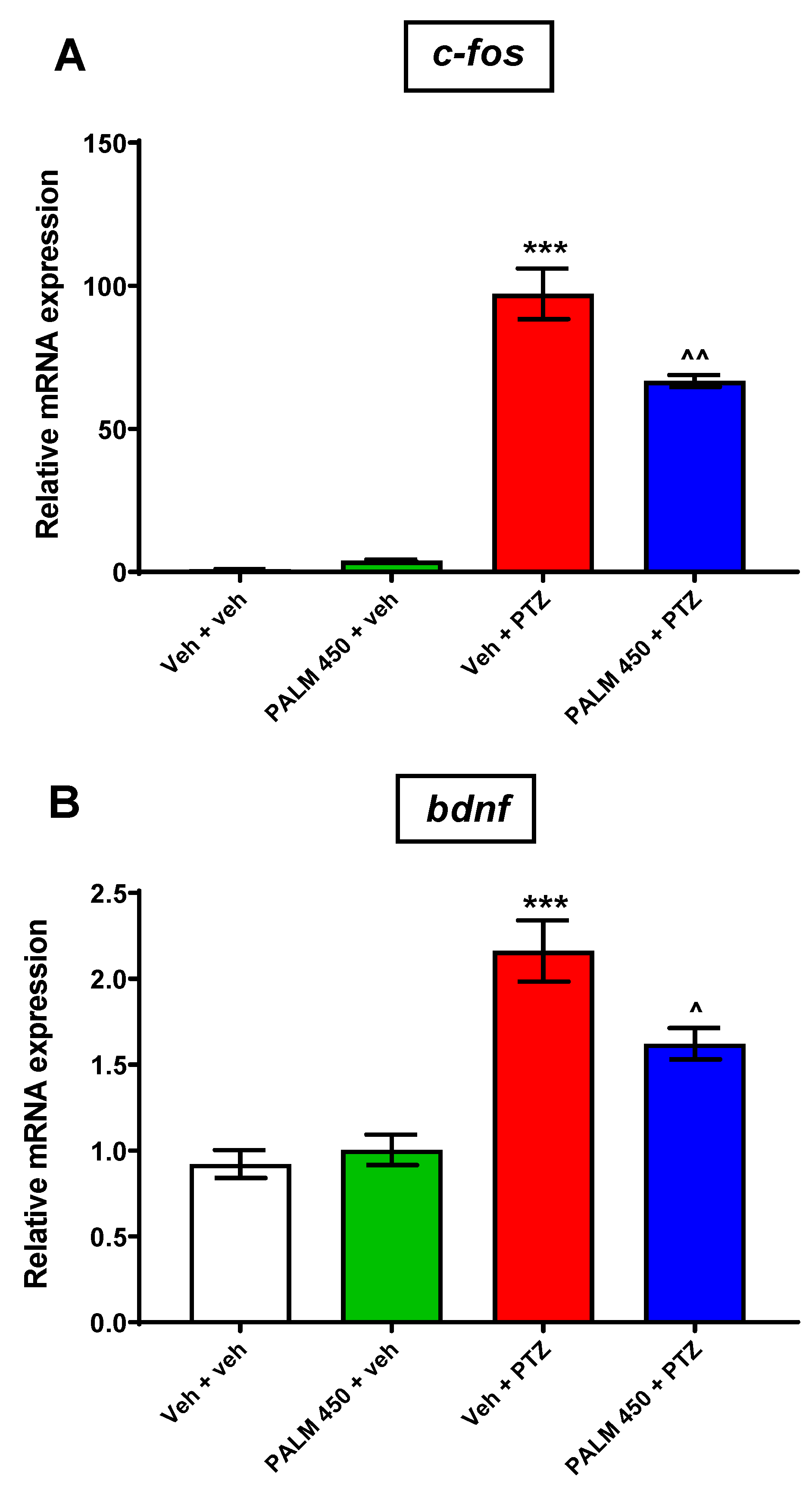
3.4. The Influence of PALM on c-fos and Bdnf Expression in PTZ-Treated Zebrafish Larvae
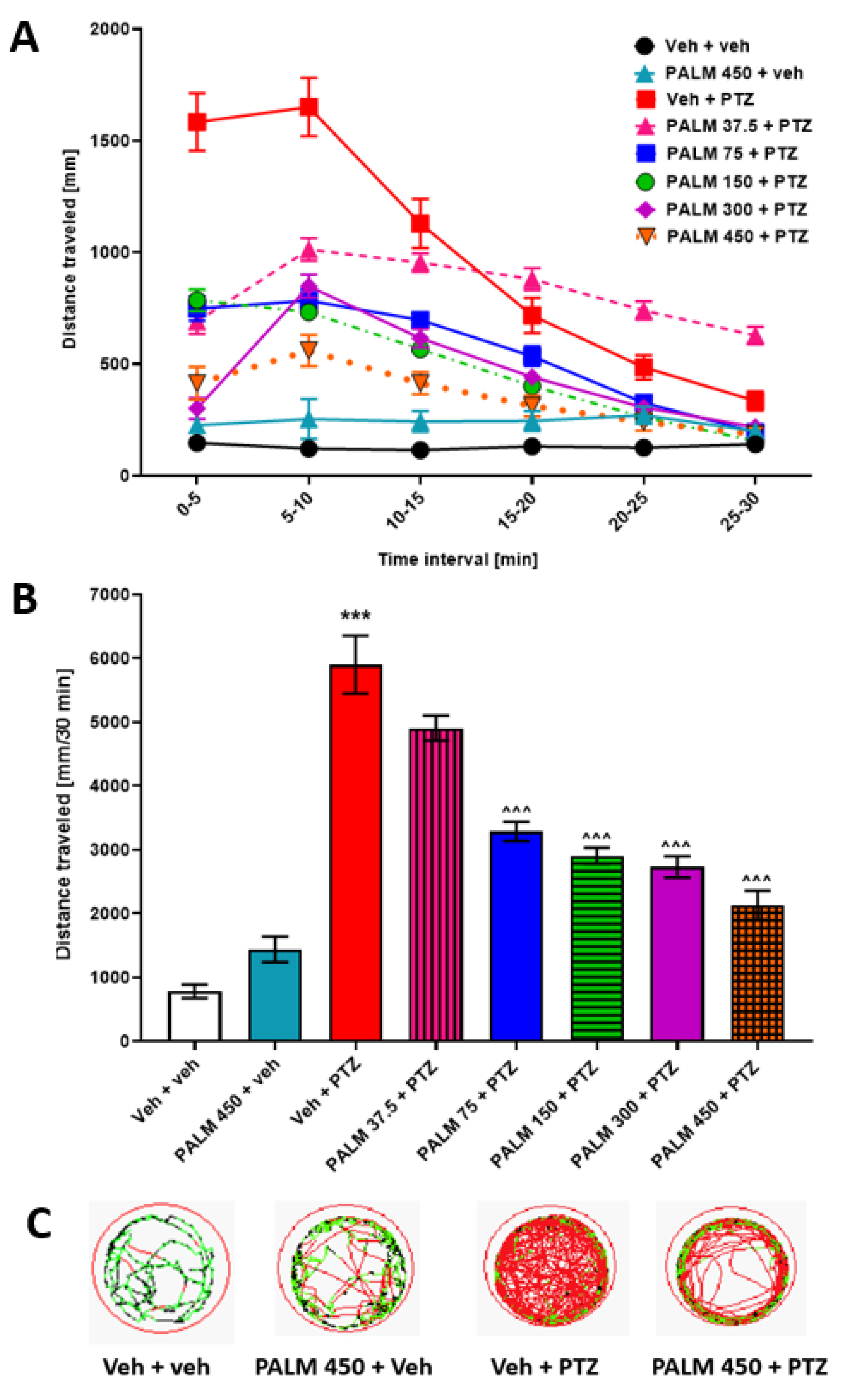
3.5. The Influence of PALM or BERB on PTZ-Induced Hyperlocomotion in Zebrafish
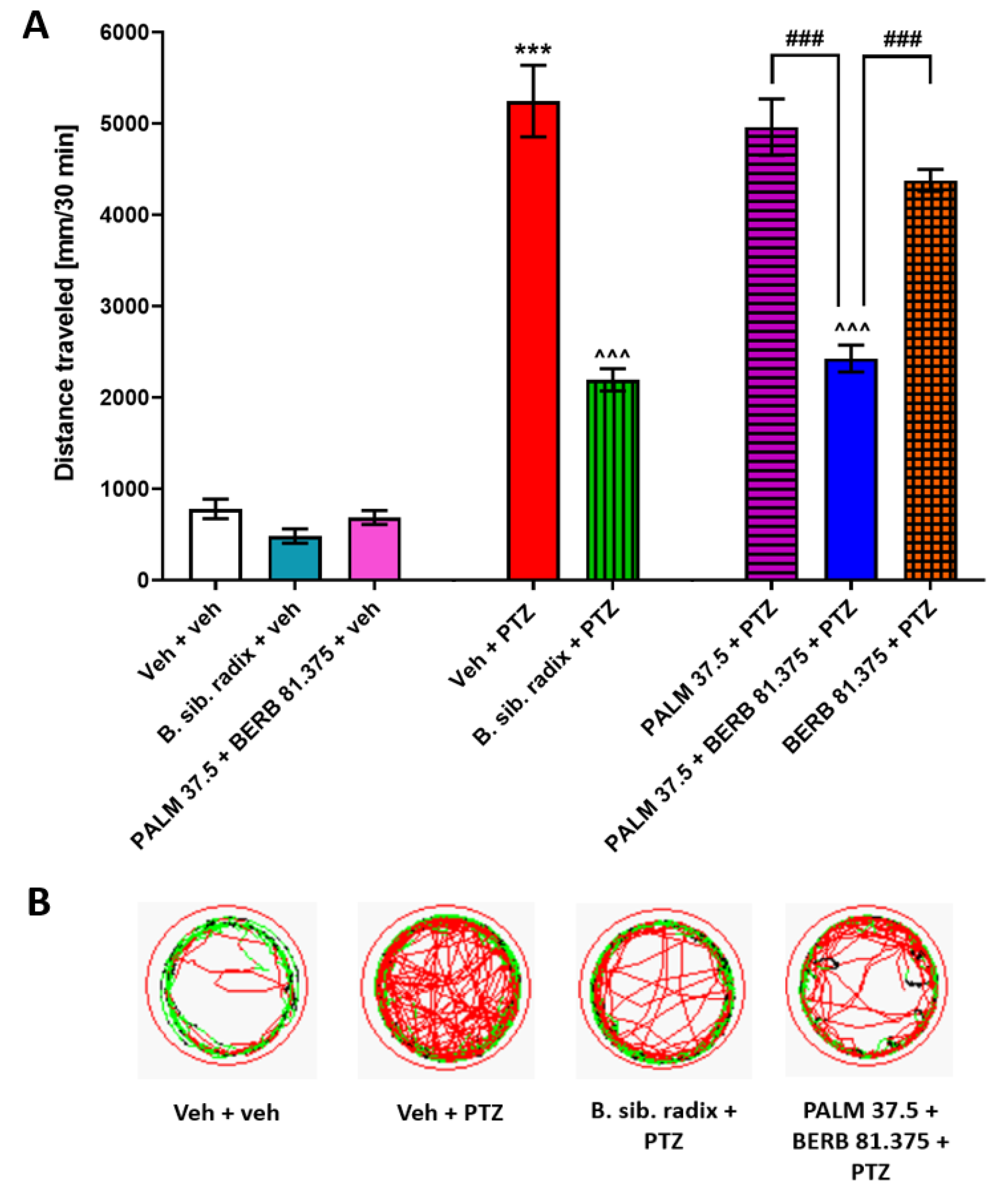
3.6. The Influence of Extract of Berberis Sibirica Radix and Combination of PALM and BERB on PTZ-Induced Hyperlocomotion in Zebrafish
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moshe, S.L.; Perucca, E.; Ryvlin, P.; Tomson, T. Epilepsy: New advances. Lancet 2015, 385, 884–898. [Google Scholar] [CrossRef]
- Engel, J., Jr. Approaches to refractory epilepsy. Ann. Indian Acad. Neurol. 2014, 17, S12–S17. [Google Scholar] [CrossRef] [PubMed]
- Perucca, P.; Gilliam, F.G. Adverse effects of antiepileptic drugs. Lancet Neurol. 2012, 11, 792–802. [Google Scholar] [CrossRef]
- Pena, I.A.; Roussel, Y.; Daniel, K.; Mongeon, K.; Johnstone, D.; Weinschutz Mendes, H.; Bosma, M.; Saxena, V.; Lepage, N.; Chakraborty, P.; et al. Pyridoxine-Dependent Epilepsy in Zebrafish Caused by Aldh7a1 Deficiency. Genetics 2017, 207, 1501–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kecskés, A.; Copmans, D.; Langlois, M.; Crawford, A.D.; Ceulemans, B.; Lagae, L.; de Witte, P.A.; Esguerra, C.V. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE 2015, 10, e0125898. [Google Scholar] [CrossRef] [Green Version]
- Siekierska, A.; Stamberger, H.; Deconinck, T.; Oprescu, S.N.; Partoens, M.; Zhang, Y.; Sourbron, J.; Adriaenssens, E.; Mullen, P.; Wiencek, P.; et al. Biallelic VARS variants cause developmental encephalopathy with microcephaly that is recapitulated in vars knockout zebrafish. Nat. Commun. 2019, 10, 708. [Google Scholar] [CrossRef] [Green Version]
- Samarut, É.; Swaminathan, A.; Riché, R.; Liao, M.; Hassan-Abdi, R.; Renault, S.; Allard, M.; Dufour, L.; Cossette, P.; Soussi-Yanicostas, N.; et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia 2018, 59, 2061–2074. [Google Scholar] [CrossRef] [Green Version]
- Swaminathan, A.; Hassan-Abdi, R.; Renault, S.; Siekierska, A.; Riché, R.; Liao, M.; de Witte, P.A.M.; Yanicostas, C.; Soussi-Yanicostas, N.; Drapeau, P.; et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Curr. Biol. 2018, 28, 1924–1937.e5. [Google Scholar] [CrossRef] [Green Version]
- Baraban, S.C.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef]
- Afrikanova, T.; Serruys, A.S.; Buenafe, O.E.; Clinckers, R.; Smolders, I.; de Witte, P.A.; Crawford, A.D.; Esguerra, C.V. Validation of the zebrafish pentylenetetrazol seizure model: Locomotor versus electrographic responses to antiepileptic drugs. PLoS ONE 2013, 8, e54166. [Google Scholar] [CrossRef] [Green Version]
- Nieoczym, D.; Socala, K.; Gawel, K.; Esguerra, C.V.; Wyska, E.; Wlaz, P. Anticonvulsant Activity of Pterostilbene in Zebrafish and Mouse Acute Seizure Tests. Neurochem. Res. 2019, 44, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminski, K.; Socala, K.; Zagaja, M.; Andres-Mach, M.; Abram, M.; Jakubiec, M.; Pierog, M.; Nieoczym, D.; Rapacz, A.; Gawel, K.; et al. N-Benzyl-(2,5-dioxopyrrolidin-1-yl)propanamide (AS-1) with Hybrid Structure as a Candidate for a Broad-Spectrum Antiepileptic Drug. Neurotherapeutics 2020, 17, 309–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brillatz, T.; Lauritano, C.; Jacmin, M.; Khamma, S.; Marcourt, L.; Righi, D.; Romano, G.; Esposito, F.; Ianora, A.; Queiroz, E.F.; et al. Zebrafish-based identification of the antiseizure nucleoside inosine from the marine diatom Skeletonema marinoi. PLoS ONE 2018, 13, e0196195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copmans, D.; Kildgaard, S.; Rasmussen, S.A.; Slezak, M.; Dirkx, N.; Partoens, M.; Esguerra, C.V.; Crawford, A.D.; Larsen, T.O.; de Witte, P.A.M. Zebrafish-Based Discovery of Antiseizure Compounds from the North Sea: Isoquinoline Alkaloids TMC-120A and TMC-120B. Mar. Drugs 2019, 17, 607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aourz, N.; Serruys, A.K.; Chabwine, J.N.; Balegamire, P.B.; Afrikanova, T.; Edrada-Ebel, R.; Grey, A.I.; Kamuhabwa, A.R.; Walrave, L.; Esguerra, C.V.; et al. Identification of GSK-3 as a Potential Therapeutic Entry Point for Epilepsy. ACS Chem. Neurosci. 2019, 10, 1992–2003. [Google Scholar] [CrossRef]
- Tarabasz, D.; Kukula-Koch, W. Palmatine: A review of pharmacological properties and pharmacokinetics. Phytother. Res. 2020, 34, 33–50. [Google Scholar] [CrossRef]
- Long, J.; Song, J.; Zhong, L.; Liao, Y.; Liu, L.; Li, X. Palmatine: A review of its pharmacology, toxicity and pharmacokinetics. Biochimie 2019, 162, 176–184. [Google Scholar] [CrossRef]
- Malhi, S.M.; Jawed, H.; Hanif, F.; Ashraf, N.; Zubair, F.; Siddiqui, B.S.; Begum, S.; Kabir, N.; Simjee, S.U. Modulation of c-Fos and BDNF protein expression in pentylenetetrazole-kindled mice following the treatment with novel antiepileptic compound HHL-6. Biomed. Res. Int. 2014, 2014, 876712. [Google Scholar] [CrossRef]
- Lim, B.V.; Shin, M.S.; Lee, J.M.; Seo, J.H. Treadmill exercise prevents GABAergic neuronal loss with suppression of neuronal activation in the pilocarpine-induced epileptic rats. J. Exerc. Rehabil. 2015, 11, 80–86. [Google Scholar] [CrossRef]
- Orellana-Paucar, A.M.; Afrikanova, T.; Thomas, J.; Aibuldinov, Y.K.; Dehaen, W.; de Witte, P.A.; Esguerra, C.V. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE 2013, 8, e81634. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Kumari, Y.; Abidin, S.A.Z.; Othman, I.; Shaikh, M.F. Pilocarpine Induced Behavioral and Biochemical Alterations in Chronic Seizure-Like Condition in Adult Zebrafish. Int. J. Mol. Sci. 2020, 21, 2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghnia, H.R.; Taji, A.R.; Forouzanfar, F.; Hosseinzadeh, H. Berberine attenuates convulsing behavior and extracellular glutamate and aspartate changes in 4-aminopyridine treated rats. Iran. J. Basic Med. Sci. 2017, 20, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.M.; Chen, B.; Jiang, J.D.; Zhang, J.P. Syntaxin 1B Mediates Berberine’s Roles in Epilepsy-Like Behavior in a Pentylenetetrazole-Induced Seizure Zebrafish Model. Front. Mol. Neurosci. 2018, 11, 378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, L.; Ji, X.; Zhang, S.; Sik, A.; Liu, K.; Jin, M. Anti-Inflammation Associated Protective Mechanism of Berberine and its Derivatives on Attenuating Pentylenetetrazole-Induced Seizures in Zebrafish. J. Neuroimmune Pharmacol. 2020. [Google Scholar] [CrossRef]
- Tang, Y.; Li, S.; Li, S.; Yang, X.; Qin, Y.; Liu, C.; Zhang, Y. Screening and isolating potential α-glucosidase inhibitors from Rhizoma Coptidis by ultrafiltration LC-PDA-ESI/MS combined with high-speed countercurrent chromatography and reverse-phase medium-pressure liquid chromatography. Med. Chem. Res. 2017, 26, 3384–3394. [Google Scholar] [CrossRef]
- Koch, W.; Angelis, A.; Halabalaki, M.; Aligiannis, N. Application of pH-zone refining hydrostatic countercurrent chromatography (hCCC) for the recovery of antioxidant phenolics and the isolation of alkaloids from Siberian barberry herb. Food Chem. 2016, 203, 394–401. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. San Diego Calif. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Litchfield, J.T., Jr.; Wilcoxon, F. A simplified method of evaluating dose-response experiments. J. Pharmacol. Exp. Ther. 1949, 96, 99–113. [Google Scholar]
- Bickel, U. How to measure drug transport across the blood-brain barrier. Neurotherapeutics 2005, 2, 15–26. [Google Scholar] [CrossRef]
- Jung, J.W.; Kwon, Y.S.; Jeong, J.S.; Son, M.; Kang, H.E. Pharmacokinetics and brain distribution of tetrahydropalmatine and tetrahydroberberine after oral administration of DA-9701, a new botanical gastroprokinetic agent, in rats. Biol. Pharm. Bull. 2015, 38, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Miao, Q.; Miao, P.; Zhao, Y.; Zhang, Y.; Chen, N.; Zhang, Y.; Ma, S. Pharmacokinetics and Brain Distribution and Metabolite Identification of Coptisine, a Protoberberine Alkaloid with Therapeutic Potential for CNS Disorders, in Rats. Biol. Pharm. Bull. 2015, 38, 1518–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wu, X.; Xian, Y.; Zhu, L.; Lin, G.; Lin, Z.X. Evidence on Integrating Pharmacokinetics to Find Truly Therapeutic Agent for Alzheimer’s Disease: Comparative Pharmacokinetics and Disposition Kinetics Profiles of Stereoisomers Isorhynchophylline and Rhynchophylline in Rats. Evid. Based Complement. Alternat. Med. 2019, 2019, 4016323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Li, J.; Cai, X.; Li, N.; Guo, Y.; Wang, D. Pharmacokinetics, Tissue Distribution, Plasma Protein Binding Studies of 10-Dehydroxyl-12-Demethoxy-Conophylline, a Novel Anti-Tumor Candidate, in Rats. Molecules 2019, 24, 283. [Google Scholar] [CrossRef] [Green Version]
- El-Gizawy, S.A.; Hedaya, M.A. Comparative brain tissue distribution of camptothecin and topotecan in the rat. Cancer Chemother. Pharmacol. 1999, 43, 364–370. [Google Scholar] [CrossRef]
- Lin-Hu, Y.; Xiao-Xi, H.; Chang, Y.; Xue, T.; Li-Sha, W.; Meng-Di, Z.; Yun-Feng, Z.; Qi, C. Pharmacokinetics of Nuciferine and N-Nornuciferine, Two Major Alkaloids From Nelumbo nucifera Leaves, in Rat Plasma and the Brain. Front. Pharmacol. 2018, 9, 902. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.S.; Ma, J.Y.; Feng, R.; Ma, C.; Chen, W.J.; Sun, Y.P.; Fu, J.; Huang, M.; He, C.Y.; Shou, J.W.; et al. Tissue distribution of berberine and its metabolites after oral administration in rats. PLoS ONE 2013, 8, e77969. [Google Scholar] [CrossRef]
- Qiao, W.; Guoshen, C. Pharmacokinetic behavior of huperzine A in plasma and cerebrospinal fluid after intranasal administration in rats. Biopharm. Drug Dispos. 2009, 30, 551–555. [Google Scholar] [CrossRef]
- Abraham, M.H.; Acree, W.E., Jr. Comparative analysis of solvation and selectivity in room temperature ionic liquids using the Abraham linear free energy relationship. Green. Chem. 2006, 8, 906–915. [Google Scholar] [CrossRef]
- Hansch, C. Quantitative approach to biochemical structure-activity relationships. Acc. Chem. Res. 1969, 2, 232–239. [Google Scholar] [CrossRef]
- Hansch, C. Quantitative structure—Activity relationships and the unnamed science. Acc. Chem. Res. 1993, 26, 147–153. [Google Scholar] [CrossRef]
- Qing, Z.X.; Cheng, P.; Liu, X.B.; Liu, Y.S.; Zeng, J.G. Systematic identification of alkaloids in Macleaya microcarpa fruits by liquid chromatography tandem mass spectrometry combined with the isoquinoline alkaloids biosynthetic pathway. J. Pharm. Biomed. Anal. 2015, 103, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Kukula-Koch, W. The Elevation of LC-ESI-Q-TOF-MS Response in the Analysis of Isoquinoline Alkaloids from Some Papaveraceae and Berberidaceae Representatives. J. Anal. Methods Chem. 2017, 2017, 8384107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, A.K.; Prathipathi, P. Comparison of MLR, PLS and GA-MLR in QSAR analysis. SAR QSAR Environ. Res. 2003, 14, 433–445. [Google Scholar] [CrossRef]
- Seiler, P. Interconversion of lipophilicities from hydrocarbon/water into the octanol/water system. Eur. J. Med. Chem. 1974, 9, 473–479. [Google Scholar]
- Goodwin, J.T.; Conradi, R.A.; Ho, N.F.; Burton, P.S. Physicochemical determinants of passive membrane permeability: Role of solute hydrogen-bonding potential and volume. J. Med. Chem. 2001, 44, 3721–3729. [Google Scholar] [CrossRef]
- Netzeva, T.I.; Worth, A.; Aldenberg, T.; Benigni, R.; Cronin, M.T.; Gramatica, P.; Jaworska, J.S.; Kahn, S.; Klopman, G.; Marchant, C.A.; et al. Current status of methods for defining the applicability domain of (quantitative) structure-activity relationships. The report and recommendations of ECVAM Workshop 52. Altern. Lab. Anim. 2005, 33, 155–173. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Kruk-Słomka, M.; Stępnik, K.; Szalak, R.; Biała, G. The Evaluation of Pro-Cognitive and Antiamnestic Properties of Berberine and Magnoflorine Isolated from Barberry Species by Centrifugal Partition Chromatography (CPC), in Relation to QSAR Modelling. Int. J. Mol. Sci. 2017, 18, 2511. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y. Golden rules and pitfalls in selecting optimum conditions for high-speed counter-current chromatography. J. Chromatogr. A. 2005, 1065, 145–168. [Google Scholar] [CrossRef]
- Bojczuk, M.; Żyżelewicz, D.; Hodurek, P. Centrifugal partition chromatography—A review of recent applications and some classic references. J. Sep. Sci. 2017, 40, 1597–1609. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Q.; Chen, M.; Huang, X.; Chen, X. Large-scale separation of acetylcholinesterase inhibitors from Zanthoxylum nitidum by pH-zone-refining counter-current chromatography target-guided by ultrafiltration high-performance liquid chromatography with ultraviolet and mass spectrometry screening. J. Sep. Sci. 2019, 42, 1194–1201. [Google Scholar] [CrossRef]
- Li, S.; Zhang, Y.; Shi, D.; Hou, W.; Xia, J.; Liu, C. Screening and isolation of cyclooxygenase-2 inhibitors from the stem bark of Phellodendron amurense Ruprecht by ultrafiltration with liquid chromatography and tandem mass spectrometry, and complex chromatography. J. Sep. Sci. 2019, 42, 1905–1914. [Google Scholar] [CrossRef]
- Li, Y.; Cai, F.; Zhang, M.; Zhang, H.; Wang, Y.; Hu, P. Two-stage fractionation of polar alkaloids from Rhizoma coptidis by countercurrent chromatography considering the strategy of reactive extraction. J. Chromatogr. A. 2015, 1378, 58–64. [Google Scholar] [CrossRef]
- Copmans, D.; Orellana-Paucar, A.M.; Steurs, G.; Zhang, Y.; Ny, A.; Foubert, K.; Exarchou, V.; Siekierska, A.; Kim, Y.; De Borggraeve, W.; et al. Methylated flavonoids as anti-seizure agents: Naringenin 4’,7-dimethyl ether attenuates epileptic seizures in zebrafish and mouse models. Neurochem. Int. 2018, 112, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Tiraboschi, E.; Martina, S.; van der Ent, W.; Grzyb, K.; Gawel, K.; Cordero-Maldonado, M.L.; Poovathingal, S.K.; Heintz, S.; Satheesh, S.V.; Brattespe, J.; et al. New insights into the early mechanisms of epileptogenesis in a zebrafish model of Dravet syndrome. Epilepsia 2020, 61, 549–560. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Ramezani, M.; Shafaei, H.; Taghiabadi, E. Anticonvulsant effect of Berberis integerrima L. root extracts in mice. J. Acupunct. Meridian Stud. 2013, 6, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, D.; Bhankher, A. Behavioral and biochemical evidences for antidepressant-like activity of palmatine in mice subjected to chronic unpredictable mild stress. Pharmacol. Rep. 2014, 66, 1–9. [Google Scholar] [CrossRef]
- Dhingra, D.; Kumar, V. Memory-enhancing activity of palmatine in mice using elevated plus maze and morris water maze. Adv. Pharmacol. Sci. 2012, 2012, 357368. [Google Scholar] [CrossRef]
- Hsieh, M.T.; Su, S.H.; Tsai, H.Y.; Peng, W.H.; Hsieh, C.C.; Chen, C.F. Effects of palmatine on motor activity and the concentration of central monoamines and its metabolites in rats. Jpn. J. Pharmacol. 1993, 61, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Yu, Y.; Qiu, L.; Qiu, M.; Zhu, Q. Palmatine plays a role in sedation and hypnosis by increasing 5-hydroxytryptamine. Cell. Mol. Biol. (Noisy-le-Grand) 2019, 65, 69–75. [Google Scholar] [CrossRef]
- Shen, Y.; Guan, S.; Ge, H.; Xiong, W.; He, L.; Liu, L.; Yin, C.; Liu, H.; Li, G.; Xu, C.; et al. Effects of palmatine on rats with comorbidity of diabetic neuropathic pain and depression. Brain. Res. Bull. 2018, 139, 56–66. [Google Scholar] [CrossRef]
- Choo, B.K.M.; Kundap, U.P.; Johan Arief, M.F.B.; Kumari, Y.; Yap, J.L.; Wong, C.P.; Othman, I.; Shaikh, M.F. Effect of newer anti-epileptic drugs (AEDs) on the cognitive status in pentylentetrazol induced seizures in a zebrafish model. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 92, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Bandara, S.B.; Carty, D.R.; Singh, V.; Harvey, D.J.; Vasylieva, N.; Pressly, B.; Wulff, H.; Lein, P.J. Susceptibility of larval zebrafish to the seizurogenic activity of GABA type A receptor antagonists. Neurotoxicology 2020, 76, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Moradi-Afrapoli, F.; Ebrahimi, S.N.; Smiesko, M.; Hamburger, M. HPLC-Based Activity Profiling for GABAA Receptor Modulators in Extracts: Validation of an Approach Utilizing a Larval Zebrafish Locomotor Assay. J. Nat. Prod. 2017, 80, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, K.M.; Herbrechter, R.; Ziemba, P.M.; Lepke, P.; Beltran, L.; Hatt, H.; Werner, M.; Gisselmann, G. Kampo Medicine: Evaluation of the Pharmacological Activity of 121 Herbal Drugs on GABAA and 5-HT3A Receptors. Front. Pharmacol. 2016, 7, 219. [Google Scholar] [CrossRef] [Green Version]
- Loscher, W.; Lehmann, H.; Teschendorf, H.J.; Traut, M.; Gross, G. Inhibiton of monoamine oxidase type A, but not type B, is and effective means of inducing anticonvulsant activity in the kindling model of epilepsy. J. Pharmacol. Exp. Ther. 1999, 288, 984–992. [Google Scholar]
- Teskey, G.C.; Radford, K.S.; Seif, I.; Dyck, R.H. MAO(A) knockout mice are more susceptible to seizures but show reduced epileptogenesis. Epilepsy Res. 2004, 59, 25–34. [Google Scholar] [CrossRef]
- Setini, A.; Pierucci, F.; Senatori, O.; Nicotra, A. Molecular characterization of monoamine oxidase in zebrafish (Danio rerio). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 140, 153–161. [Google Scholar] [CrossRef]
- Sallinen, V.; Sundvik, M.; Reenilä, I.; Peitsaro, N.; Khrustalyov, D.; Anichtchik, O.; Toleikyte, G.; Kaslin, J.; Panula, P. Hyperserotonergic phenotype after monoamine oxidase inhibition in larval zebrafish. J. Neurochem. 2009, 109, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, D.; McQuade, J.S.; Behbehani, M.; Tsien, J.Z.; Xu, M. c-fos regulates neuronal excitability and survival. Nat. Genet. 2002, 30, 416–420. [Google Scholar] [CrossRef]
- Cacialli, P.; Gueguen, M.M.; Coumailleau, P.; D’Angelo, L.; Kah, O.; Lucini, C.; Pellegrini, E. BDNF Expression in Larval and Adult Zebrafish Brain: Distribution and Cell Identification. PLoS ONE 2016, 11, e0158057. [Google Scholar] [CrossRef] [Green Version]
- Torres-Hernandez, B.A.; Colon, L.R.; Rosa-Falero, C.; Torrado, A.; Miscalichi, N.; Ortiz, J.G.; Gonzalez-Sepulveda, L.; Perez-Rios, N.; Suarez-Perez, E.; Bradsher, J.N.; et al. Reversal of pentylenetetrazole-altered swimming and neural activity-regulated gene expression in zebrafish larvae by valproic acid and valerian extract. Psychopharmacology (Berl.) 2016, 233, 2533–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dona, F.; Ulrich, H.; Persike, D.S.; Conceicao, I.M.; Blini, J.P.; Cavalheiro, E.A.; Fernandes, M.J. Alteration of purinergic P2X4 and P2X7 receptor expression in rats with temporal-lobe epilepsy induced by pilocarpine. Epilepsy Res. 2009, 83, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Pacheco, A.; Mesuret, G.; Sanz-Rodriguez, A.; Tanaka, K.; Mooney, C.; Conroy, R.; Miras-Portugal, M.T.; Diaz-Hernandez, M.; Henshall, D.C.; Engel, T. Increased neocortical expression of the P2X7 receptor after status epilepticus and anticonvulsant effect of P2X7 receptor antagonist A-438079. Epilepsia 2013, 54, 1551–1561. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Pacheco, A.; Diaz-Hernandez, M.; Arribas-Blazquez, M.; Sanz-Rodriguez, A.; Olivos-Ore, L.A.; Artalejo, A.R.; Alves, M.; Letavic, M.; Miras-Portugal, M.T.; Conroy, R.M.; et al. Transient P2X7 Receptor Antagonism Produces Lasting Reductions in Spontaneous Seizures and Gliosis in Experimental Temporal Lobe Epilepsy. J. Neurosci. 2016, 36, 5920–5932. [Google Scholar] [CrossRef]
- Song, P.; Hu, J.; Liu, X.; Deng, X. Increased expression of the P2X7 receptor in temporal lobe epilepsy: Animal models and clinical evidence. Mol. Med. Rep. 2019, 19, 5433–5439. [Google Scholar] [CrossRef]
- Barros-Barbosa, A.R.; Fonseca, A.L.; Guerra-Gomes, S.; Ferreirinha, F.; Santos, A.; Rangel, R.; Lobo, M.G.; Correia-de-Sa, P.; Cordeiro, J.M. Up-regulation of P2X7 receptor-mediated inhibition of GABA uptake by nerve terminals of the human epileptic neocortex. Epilepsia 2016, 57, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Fischer, W.; Franke, H.; Krugel, U.; Muller, H.; Dinkel, K.; Lord, B.; Letavic, M.A.; Henshall, D.C.; Engel, T. Critical Evaluation of P2X7 Receptor Antagonists in Selected Seizure Models. PLoS ONE 2016, 11, e0156468. [Google Scholar] [CrossRef]
- Nieoczym, D.; Socala, K.; Wlaz, P. Evaluation of the Anticonvulsant Effect of Brilliant Blue G, a Selective P2X7 Receptor Antagonist, in the iv PTZ-, Maximal Electroshock-, and 6 Hz-Induced Seizure Tests in Mice. Neurochem. Res. 2017, 42, 3114–3124. [Google Scholar] [CrossRef]
- Nateri, A.S.; Raivich, G.; Gebhardt, C.; Da Costa, C.; Naumann, H.; Vreugdenhil, M.; Makwana, M.; Brandner, S.; Adams, R.H.; Jefferys, J.G.; et al. ERK activation causes epilepsy by stimulating NMDA receptor activity. EMBO J. 2007, 26, 4891–4901. [Google Scholar] [CrossRef]
- Sun, J.; Nan, G. The extracellular signal-regulated kinase 1/2 pathway in neurological diseases: A potential therapeutic target (Review). Int. J. Mol. Med. 2017, 39, 1338–1346. [Google Scholar] [CrossRef] [Green Version]
- Merlo, D.; Cifelli, P.; Cicconi, S.; Tancredi, V.; Avoli, M. 4-Aminopyridine-induced epileptogenesis depends on activation of mitogen-activated protein kinase ERK. J. Neurochem. 2004, 89, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Otani, N.; Nawashiro, H.; Yano, A.; Katoh, H.; Ohnuki, A.; Miyazawa, T.; Shima, K. Characteristic phosphorylation of the extracellular signal-regulated kinase pathway after kainate-induced seizures in the rat hippocampus. Acta Neurochir. 2003, 86, 571–573. [Google Scholar] [CrossRef]
- Cheng, Y.; Mai, Q.; Zeng, X.; Wang, H.; Xiao, Y.; Tang, L.; Li, J.; Zhang, Y.; Ding, H. Propionate relieves pentylenetetrazol-induced seizures, consequent mitochondrial disruption, neuron necrosis and neurological deficits in mice. Biochem. Pharmacol. 2019, 169, 113607. [Google Scholar] [CrossRef] [PubMed]
- Berkeley, J.L.; Decker, M.J.; Levey, A.I. The role of muscarinic acetylcholine receptor-mediated activation of extracellular signal-regulated kinase 1/2 in pilocarpine-induced seizures. J. Neurochem. 2002, 82, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Houser, C.R.; Huang, C.S.; Peng, Z. Dynamic seizure-related changes in extracellular signal-regulated kinase activation in a mouse model of temporal lobe epilepsy. Neuroscience 2008, 156, 222–237. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.H.; Chiang, Y.W.; Liang, K.C.; Thompson, R.F.; Liu, I.Y. Extra-cellular signal-regulated kinase 1/2 (ERK1/2) activated in the hippocampal CA1 neurons is critical for retrieval of auditory trace fear memory. Brain Res. 2010, 1326, 143–151. [Google Scholar] [CrossRef]
- Glazova, M.V.; Nikitina, L.S.; Hudik, K.A.; Kirillova, O.D.; Dorofeeva, N.A.; Korotkov, A.A.; Chernigovskaya, E.V. Inhibition of ERK1/2 signaling prevents epileptiform behavior in rats prone to audiogenic seizures. J. Neurochem. 2015, 132, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.Q.; Wang, X.F.; He, R.Q.; Li, M.W.; Liu, X.Z.; Wang, L.Y.; Zhu, X.; Xiao, F.; Sun, J.J.; Li, J.M.; et al. Extracellular signal-regulated protein kinase in human intractable epilepsy. Eur. J. Neurol. 2007, 14, 865–872. [Google Scholar] [CrossRef]
- Mohammad Jafari, R.; Ghahremani, M.H.; Rahimi, N.; Shadboorestan, A.; Rashidian, A.; Esmaeili, J.; Ejtemaei Mehr, S.; Dehpour, A.R. The anticonvulsant activity and cerebral protection of chronic lithium chloride via NMDA receptor/nitric oxide and phospho-ERK. Brain Res. Bull. 2018, 137, 1–9. [Google Scholar] [CrossRef]
- Krens, S.F.; Spaink, H.P.; Snaar-Jagalska, B.E. Functions of the MAPK family in vertebrate-development. FEBS Lett. 2006, 580, 4984–4990. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basnet, R.M.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish Larvae as a Behavioral Model in Neuropharmacology. Biomedicines 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buenafe, O.E.; Orellana-Paucar, A.; Maes, J.; Huang, H.; Ying, X.; De Borggraeve, W.; Crawford, A.D.; Luyten, W.; Esguerra, C.V.; de Witte, P. Tanshinone IIA exhibits anticonvulsant activity in zebrafish and mouse seizure models. ACS Chem. Neurosci. 2013, 4, 1479–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | A | logPo/w | MW | logBBexp. |
|---|---|---|---|---|
| Tetrahydropalmatine | 0 | 3.373 | 355.43 | 0.43 |
| Tetrahydroberberine | 0 | 3.084 | 339.39 | 0.39 |
| Coptisine | 0 | 4.193 | 320.32 | 0.46 |
| Isorhynchophylline | 0.41 | 3.304 | 384.47 | −0.59 |
| Rhynchophylline | 0.41 | 3.304 | 384.47 | −0.59 |
| 10-dehydroxyl-12-demethoxy-conophylline | 0.5 | 7.675 | 794.89 | −0.92 |
| Camptothecin | 0.17 | 0.742 | 384.35 | −0.29 |
| Topotecan | 0.67 | 0.962 | 421.45 | −0.42 |
| Nuciferine | 0 | 4.107 | 295.38 | −0.52 |
| Nornuciferine | 0.13 | 3.752 | 295.38 | −0.18 |
| Berberine | 0 | 4.499 | 336.36 | −0.17 |
| Huperzine a | 0.47 | 1.483 | 242.32 | −0.70 |
| Dehydrocorydaline | 0 | 5.405 | 366.43 | −0.59 |
| Descriptors | SE Coefficient | p-Value | Variance Inflation Factors (VIF) | F-Value |
|---|---|---|---|---|
| A | 0.670 | 0.003 | 3.67 | 18.82 |
| logPo/w | 0.125 | 0.016 | 3.32 | 10.06 |
| MW | 0.00195 | 0.096 | 1.26 | 3.71 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gawel, K.; Kukula-Koch, W.; Nieoczym, D.; Stepnik, K.; van der Ent, W.; Banono, N.S.; Tarabasz, D.; Turski, W.A.; Esguerra, C.V. The Influence of Palmatine Isolated from Berberis sibirica Radix on Pentylenetetrazole-Induced Seizures in Zebrafish. Cells 2020, 9, 1233. https://doi.org/10.3390/cells9051233
Gawel K, Kukula-Koch W, Nieoczym D, Stepnik K, van der Ent W, Banono NS, Tarabasz D, Turski WA, Esguerra CV. The Influence of Palmatine Isolated from Berberis sibirica Radix on Pentylenetetrazole-Induced Seizures in Zebrafish. Cells. 2020; 9(5):1233. https://doi.org/10.3390/cells9051233
Chicago/Turabian StyleGawel, Kinga, Wirginia Kukula-Koch, Dorota Nieoczym, Katarzyna Stepnik, Wietske van der Ent, Nancy Saana Banono, Dominik Tarabasz, Waldemar A. Turski, and Camila V. Esguerra. 2020. "The Influence of Palmatine Isolated from Berberis sibirica Radix on Pentylenetetrazole-Induced Seizures in Zebrafish" Cells 9, no. 5: 1233. https://doi.org/10.3390/cells9051233