1. Introduction
The liver has the exceptional ability to regenerate after injury or surgical resection of tissue. It is widely recognized that this regenerating process is governed by an intricate interplay of hundreds of genes, which together form dozens of interwoven networks of signaling pathways. These pathways regulate biological processes, such as cell cycle arrest, cell cycle progression, apoptosis, inflammation, angiogenesis, etc. The entire process of liver regeneration is broadly defined by three distinct phases: an initiation phase, a proliferation phase, and a termination phase. Traditionally, it is thought that during the initiation phase, key signaling pathways induced by TNF-α and IL-6 drive cell proliferation in several hepatic cell types [
1]. Then, growth factors and cytokines guide the progression of liver regeneration through expression of several cell-cycle-related proteins mainly by PI3K/AKT, wnt/β-catenin, Ras/MAPK, and JAK/STAT signaling pathways [
2]. Equally important are pathways that control the speed of proliferation and determine the terminal point of liver regeneration [
3]. Previously, we demonstrated that expression and activation of p21, an inhibitor of a subset of cyclin dependent kinases including cdc2, is a pivotal factor in arrested liver regeneration [
4]. P21 is a fundamental cell cycle control checkpoint regulator, in particular, as it is inhibiting the G2-M phase progression. As such, it is downstream of many signaling networks, including those induced by p53, injury, growth factors (TGF-β), and inflammatory cytokines [
3].
Limits to physiological liver regeneration are experienced in the clinic. Following extended hepatectomy that leaves behind a marginal liver remnant, liver failure may develop, a syndrome known as the small-for-size syndrome (SFSS). The SFSS is typified by metabolic liver dysfunction and represents the most frequent cause of postoperative death due to liver surgery [
1]. Extended resection however is the most frequent intervention against highly prevalent liver tumors, which can be cured through complete removal only. The SFSS is currently untreatable and hence creates a medical conflict in that it limits the surgical cure of liver cancer [
1,
4]. Despite numerous studies addressing normal liver regeneration, we still lack comprehensive understanding of the biological processes underlying normal and failed liver regeneration. Such knowledge is required to categorize the biological processes of failed liver regeneration as observed in the SFSS.
With the development of reliable surgical mouse models, which reflect normal and extended liver hepatectomy in humans, and the advent of appropriate bioinformatics tools to analyze genome wide gene expression profiling, comprehensive categorization has now become feasible. To identify the intracellular signaling pathways (ISP) involved in normal and extended liver regeneration, we applied the OncoFinder algorithm for the functional annotation of the primary next generation sequencing genome-wide expression data. The advantage of OncoFinder over alternative tools, such as Metacore and Ingenuity Pathway Analysis (IPA), is that it calculates the pathway activation strength (PAS), so that differences between the two surgical procedures can be quantified [
5,
6].
Here, we analyzed the expression profiles of >20,000 genes and determined how they interact together in 271 IPA-defined intracellular signaling pathways. Our analyses of more than 12 million data points demonstrate that liver regeneration is a highly orchestrated flowing process, in which many signaling pathways operate simultaneously, either additively or inversely. We also show that the distinct stages of normal and failed regeneration are clearly discernable by a discrete activation and deactivation of a small number of signaling pathways. The five time points we studies in our nHx and eHx models were clearly demarcated by only a limited number of pathways, which may reflect normal and abnormal regeneration processes—the latter leading to liver failure and/or delayed regeneration.
4. Discussion
Failed liver regeneration after major resection has been a subject of intense scrutiny, as it posits a severe restraint to liver surgery. Resection beyond 70 percent of the total liver mass is critical, since it often leads to the small for size syndrome (SFSS), a pathology characterized by hyper-bilirubinemia, diminished (or failed) liver function, and, as such, is the most frequent cause of death due to liver surgery. Earlier, we demonstrated that p21 activation mediates a transient barrier to the progression and completion of the cell cycle, thereby inhibiting liver regeneration and deteriorating survival [
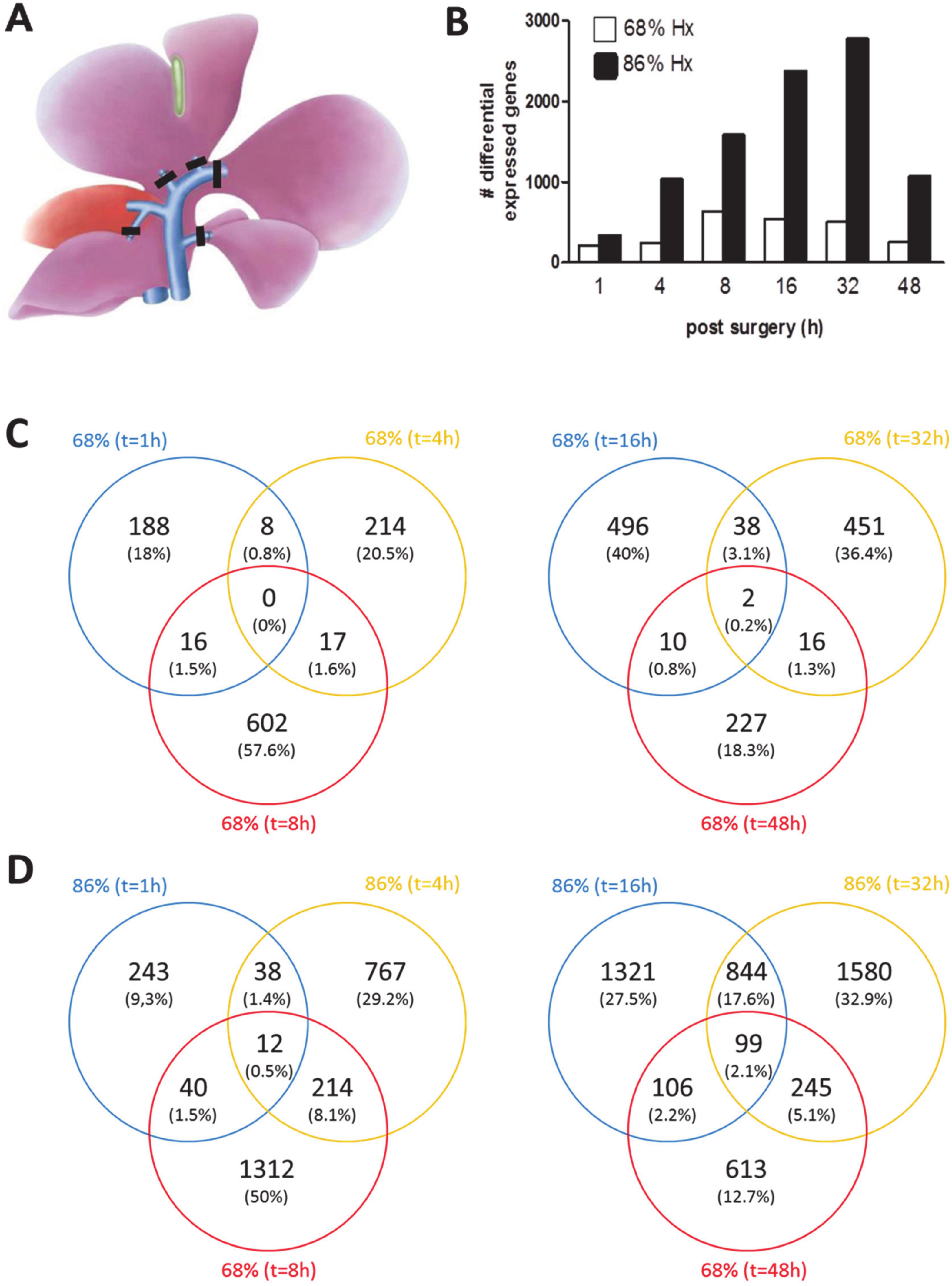
4]. To comprehend the molecular background of SFSS, however, the total of regulatory processes operating in the liver during normal and failed/delayed regeneration should be integrated, a challenge, which can only be overcome using bioinformatics approaches. This study is the first systematic and comprehensive analysis of liver-expressed genes to profile intracellular signaling pathways (ISPs) after nHx and eHx. Our study submits that relative to eHx, nHx requires a lower level of gene activation with an optimal expression of candidate genes around 8 h post-surgery. Thereafter, gene activation rapidly returns to control levels (
Figure 1B). In contrast, eHx is associated with a 3- to 4-fold overshoot of differentially regulated candidate genes, which is most pronounced 32 h post-surgery. Forty-eight hours after the extended procedure, the liver seems to have mastered the affliction and gene expression levels start to go down. Relative to the normal procedure, however, there is still an enormous overshoot of genetic activity. These facts indicate that normal liver regeneration requires a stringent control over gene expression, which seems to be transiently, but severely, disturbed after extended hepatectomy.
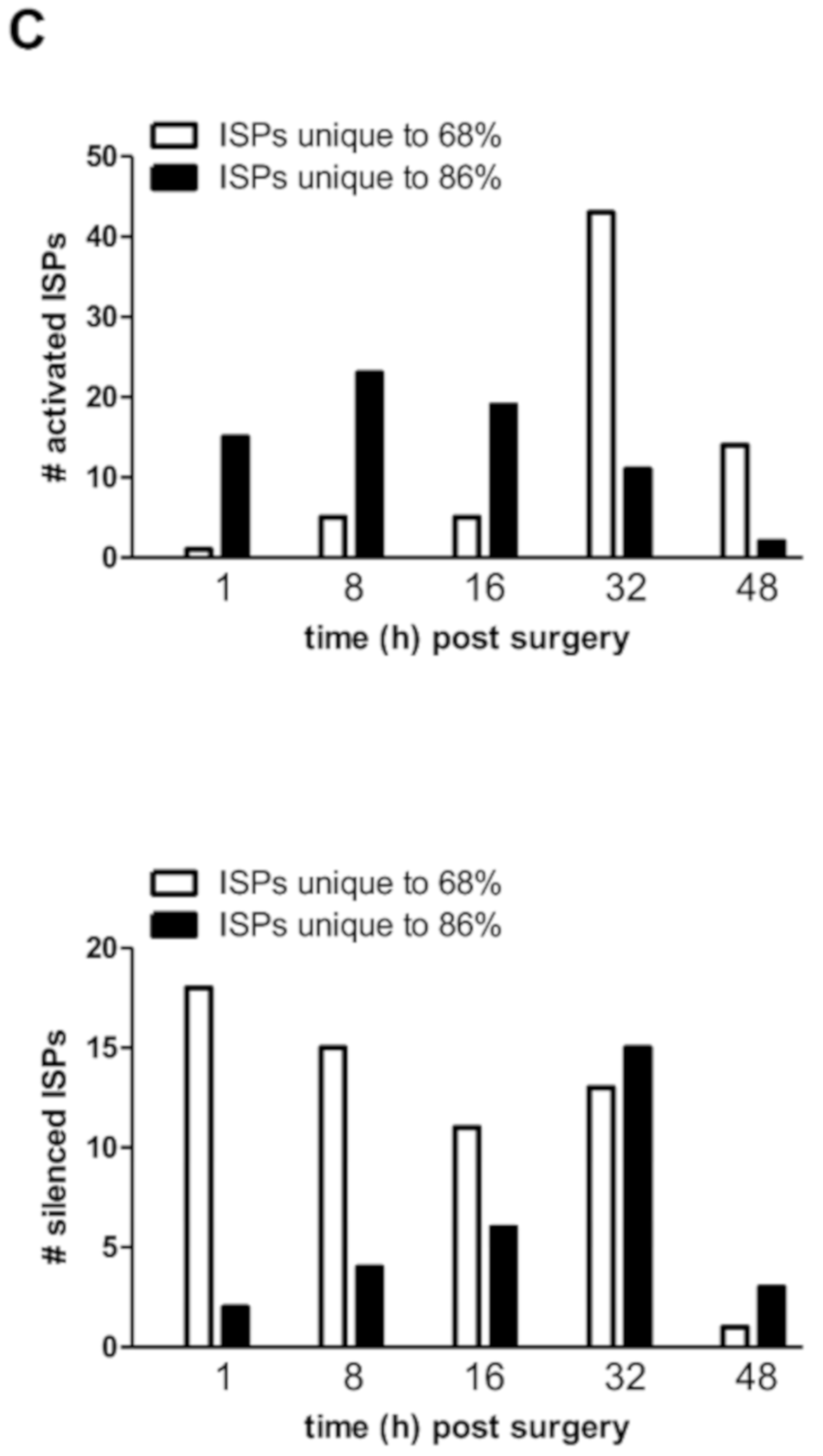
How all the liver-expressed genes interact together and affect biological relevant processes was studied using OncoFinder, software designed to quantify pathway activation strengths according to gene expression levels [
5,
8]. We observed that normal liver regeneration is an orchestration of the sequential silencing and activation of several dozen cooperating ISPs (
Figure 2C). Importantly, the early phase of normal liver regeneration is defined by a predominant silencing, rather than an activation of ISPs. Then, during the late phase of normal liver regeneration (32–48 h), whole arrays of ISPs are being activated simultaneously but transiently. The opposite was observed after extended hepatectomy, which resulted in an immediate activation of a plethora of signaling cascades. Therefore, delayed (or failed) liver regeneration may result from a hyper-activation, rather than hypo-activation (or silencing) of ISPs. The earlier described activation of p21 by Lehmann et al. [
4] may represent the ensuing biological response, since it would transiently stall the cell cycle, so that (epi)genetic salvage programs can be initiated.
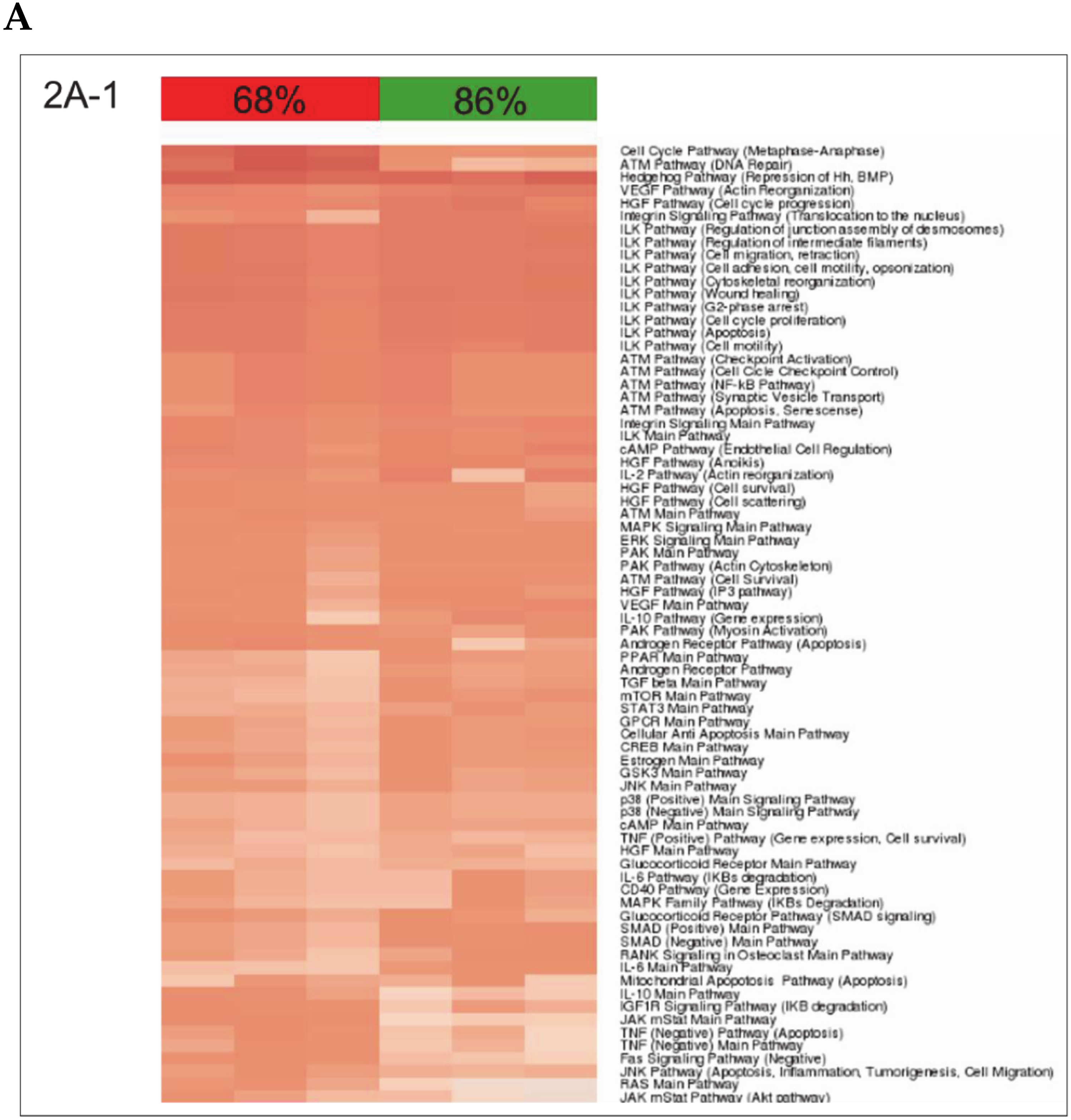
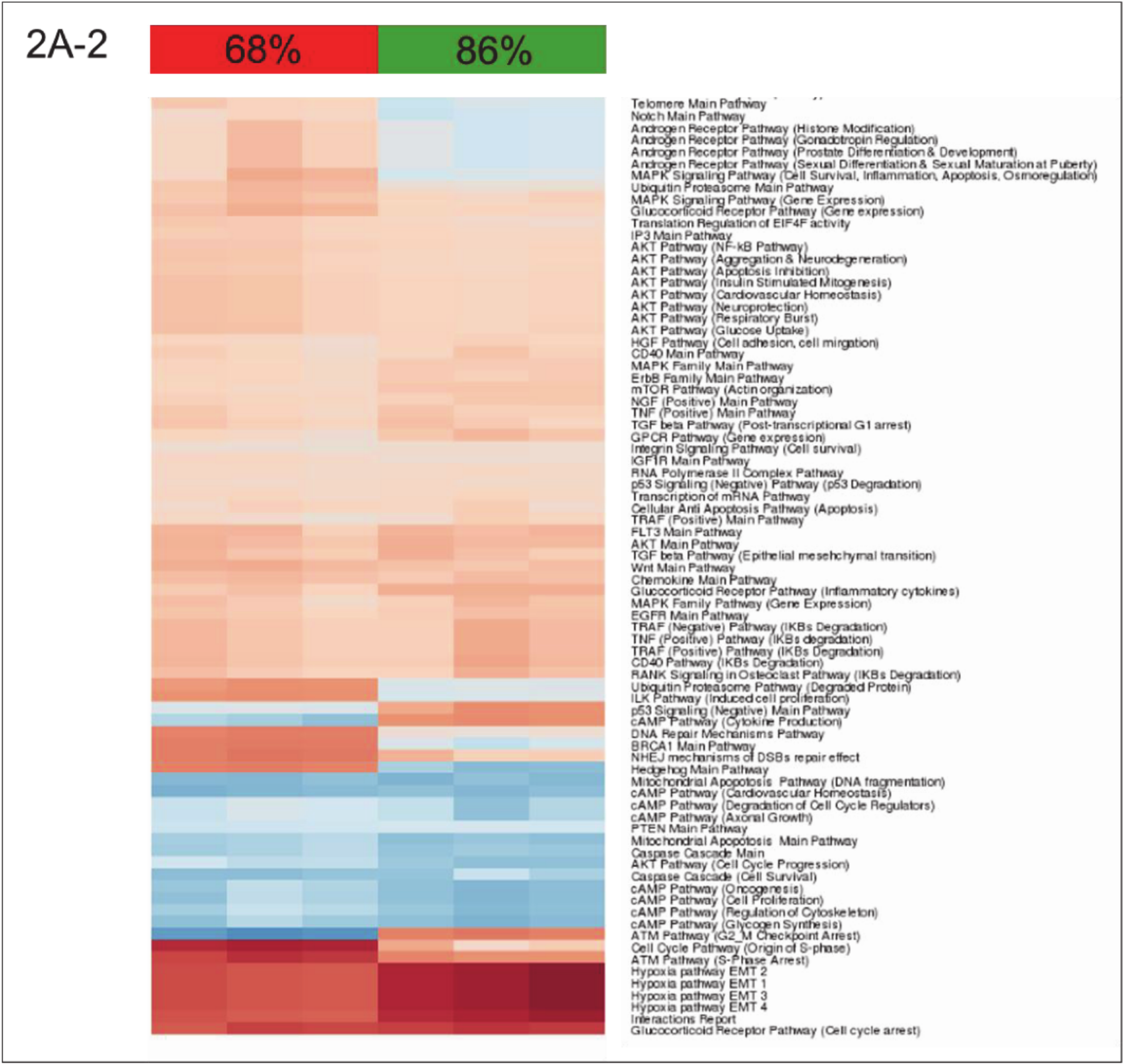
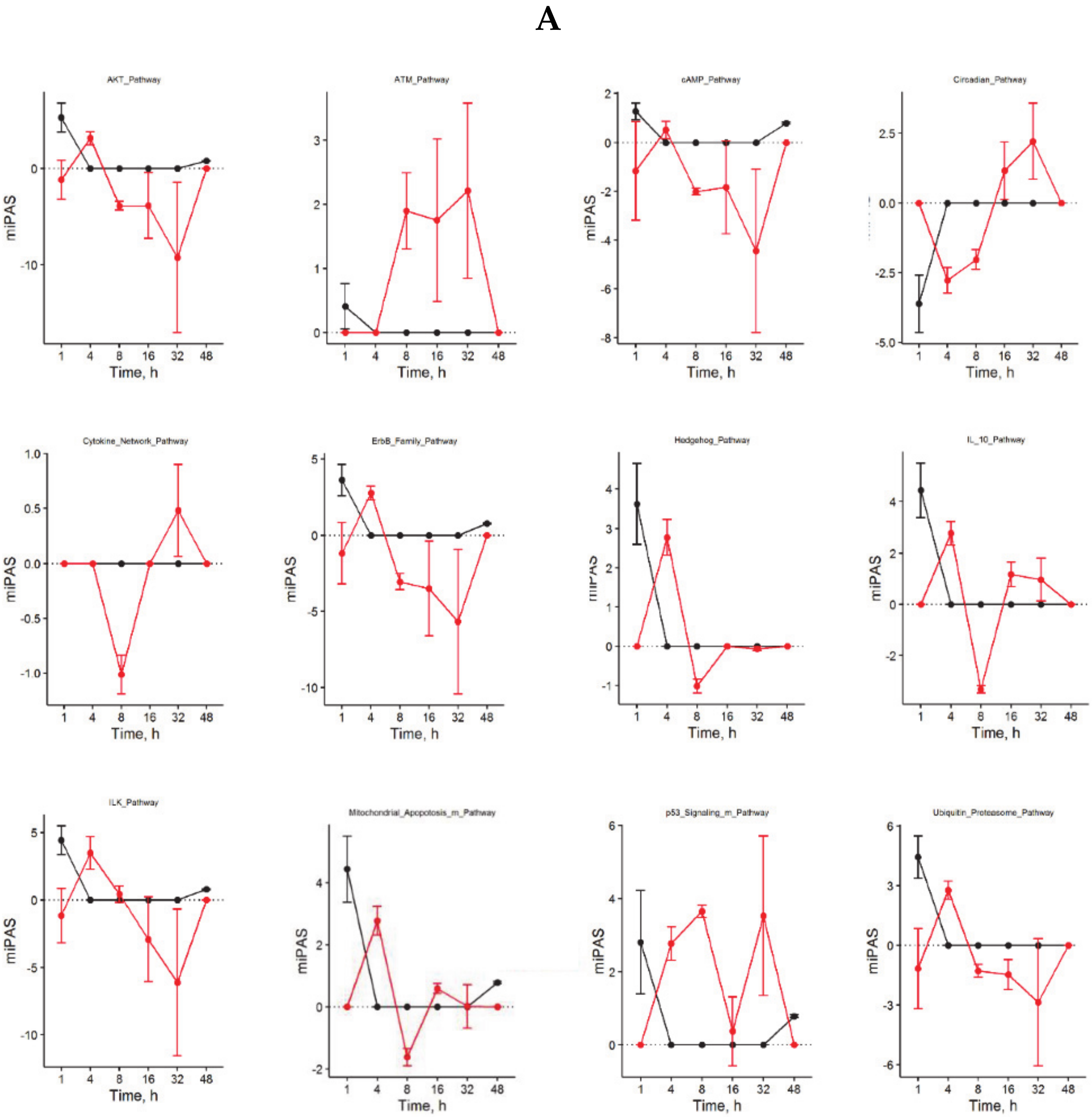
A closer look at the identified ISPs revealed that both procedures recruit pathways that traditionally have been linked to liver regeneration. The most pronounced common pathways activated after both nHx and eHx include the
Hedgehog Pathway (
Figure 2A-1) and four branches of the
Hypoxia Pathway involved in epithelial-mesenchymal transition (
Figure 2A-2). In addition, we identified the
IL10 Pathway as an important ISP after both normal and extended hepatectomies. A pleiotropic cytokine with important immuno-regulatory functions, IL-10 may act to silence the initial inflammatory response through counteracting the expression of inflammatory cytokines such as TNF-α, interleukin (IL)-6, and IL-1 by activated macrophages [
14]. The most distinct common pathways silenced after both nHx and eHx are the seven branches of the
cAMP Pathway. In the liver, cAMP-responsive signaling plays an important role, particularly in regulating hepatocyte proliferation as observed after nHx [
15]. In addition, silencing of the cAMP-induced pathway may prompt hepatic cells to become less committed and prepare them for cell cycle entry [
16]. Most PAS values presented in
Figure 2A-1 demonstrate only minor differences between the two surgical procedures. Still, small but significant differences in PAS values may lead to significant biological effects, as they represent the additive activation strengths of the components of the pathway as a whole [
5,
8]. More pronounced differences also reflect stronger involvement in the pathological processes observed after delayed liver regeneration.
Among the unique ISPs activated after
nHx, the
Akt Pathway represented with 13 branches, the
Chromatin Pathway and the
DDR Pathway—with 3 branches each—dominated (
Table 1A). The
Akt Pathways, which are triggered essentially by growth factors, induce the progression of G1→M and G2→M transitions via silencing the activity of p53 and p21, two proteins crucial to cell cycle entry. The
chromatin- and
DDR Pathways signify the importance of epigenetic configurations after nHx. Crucial for a normal regeneration process seems to be the sequential silencing of a score of ISPs, including most branches of the
ILK Pathway during the late phase, which are not (yet) detected after extended hepatectomy (
Table 2). Indeed, the integrin-linked kinase (ILK) is a protein involved in transmitting extracellular matrix signals and associated with the termination of liver regeneration [
17].
Among the unique ISPs activated after extended hepatectomy, the
ATM Main Pathway and two of its branches (
Cell Survival; G2_M Checkpoint Arrest) dominated. Both pathways may be induced by severe (DNA) damage inflicted by extended hepatectomy, dramatically increased blood pressure, or both, which retain the cells in the G2 phase. In addition, five branches of the
HIF1-Alpha Main Pathway (
Gene Expression Pathway; NOS Pathway; Pyruvate Pathway; VEGF Pathway) prevailed. These pathways, which are closely associated with the
VEGF Main Pathway, may reflect the immediate early hypoxic conditions after extended liver hepatectomy. Both nHx and eHx rapidly induced hypoxia-associated ISPs, an observation, which may reflect the overshoot of oxygen-poor blood delivered from the portal vein, and probably required for the early phase of liver regeneration through Hif2a [
18]. Interestingly, the hypoxia-induced pathways are swiftly down-regulated after 32 h, but only following nHx. Hypoxia may thus be an important fine-tuning mechanism to induce liver regeneration, functioning within a very narrow window. Prolonged hypoxia-associated signaling may stall the normal regeneration process after eHx, however, as it may inhibit G1/S transitions through regulation of p27 expression [
19]. Indeed, loss of p27 leads to accelerated DNA synthesis in hepatocytes DNA replication after partial hepatectomy in mice [
20].
Fourteen ISPs demonstrated a strong inverse regulation after nHx and eHx (
Table 3; PAS ≥ 0.1, highlighted). Together with the surgery-specific pathways discussed above, they may represent the most relevant pathways determining a healthy/normal or diseased/delayed regeneration process. Here, the
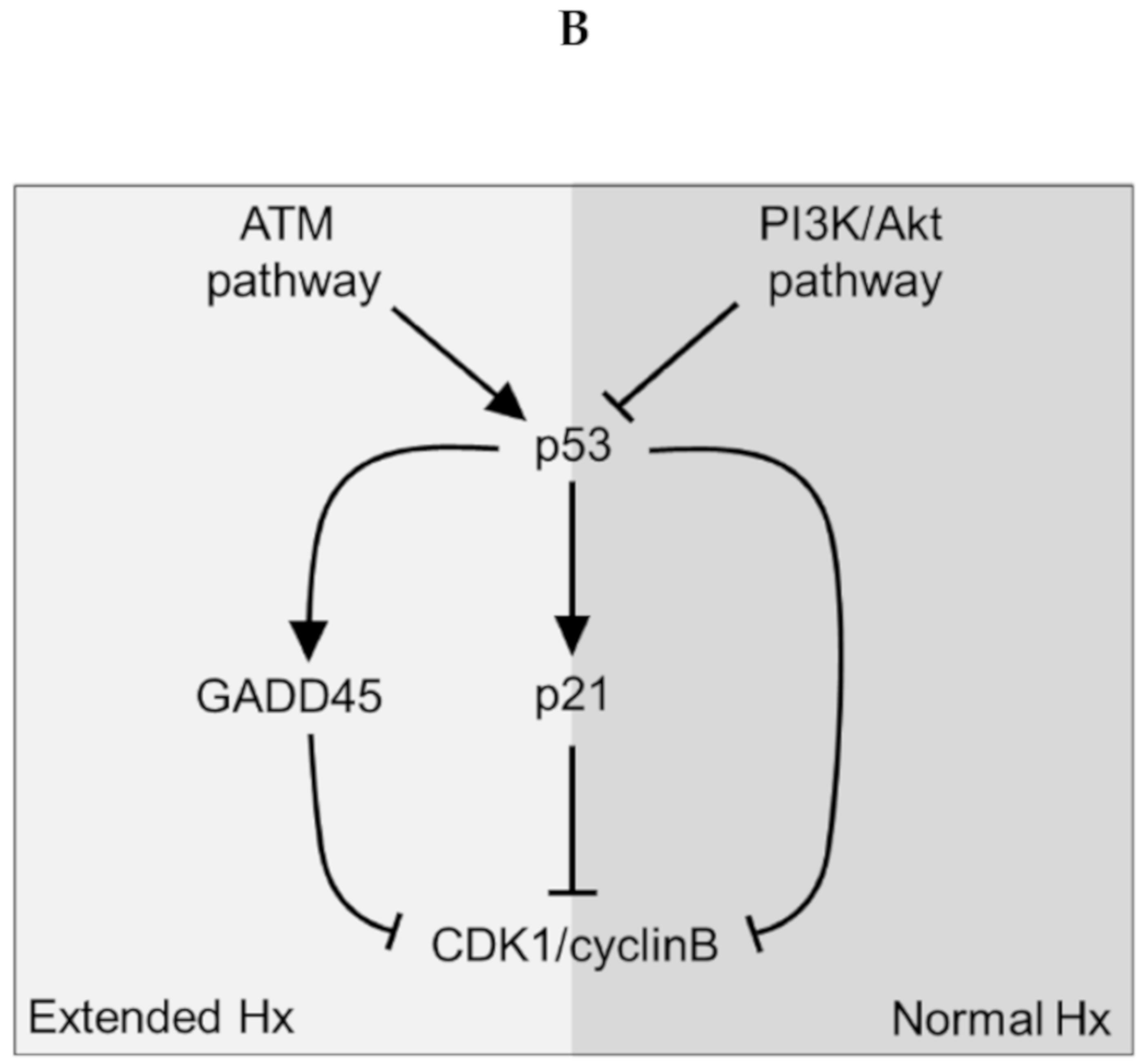
ATM Pathway (
G2_M checkpoint arrest) is of particular interest. Earlier, we observed that liver dysfunction after eHx resulted from a deficiency in cell cycle progression caused by a transient activation of p21 just before hepatocyte division, rather than from parenchymal injury [
4]. Oncofinder identified the
ATM Pathway (
G2_M checkpoint arrest) with p53 and p21 as dominant cell cycle checkpoints, to be the most differentially activated pathway—thus confirming our earlier data. Therefore, a deficiency in cell-cycle progression after extended hepatectomy and concomitant liver failure might be overcome by counteracting this pathway. Our data suggest this can be achieved by stimulation of the Akt-associated pathways, which are explicitly activated early after nHx but not after eHx, and interfere with the ATM pathway at the level of p53, the activator of p21. The pre-miRNA identified in the eHx samples also indicated a strong reciprocal regulation of these pathways. Our comprehensive ISP analyses thus reveals potential intervention points to treat failed liver regeneration.
Taken together, a timely—but transiently—activation and silencing of approximately two dozen of ISPs warrant a normal regenerative process, culminating in full recovery of small liver remnants. The most affected ISPs are revolving around Akt-mediated signaling—which is tightly associated with several pathways, including the
JNK Pathway, the
NF-κB, and the
ERK Pathway—and the
ATM Pathway via the p53-p21 axis (
Figure 4B). Together, they govern the G1/S Checkpoint and the G2/M DNA Damage Checkpoint, in which p21 and p53 are major executors. The miRNAs found exclusively after hepatectomy may interfere with Akt-signaling, thereby silencing the downstream cell cycle programs. Indeed, 20 of the identified miRNAs specific for extended hepatectomy target the transcripts of 18 genes operating directly or indirectly in the Akt pathway. One miRNA, mmu-miR692, is expressed immediately after normal regeneration and 48 h after extended hepatectomy, suggesting that it may prepare for a smooth regeneration process—as suggested by the predominant ISP silencing during these stages. Alternatively, the observed overload of pathway activities (
Figure 1) after 86% may reflect a metabolic overload as a major cause of liver failure. Metabolic changes after hepatectomy are thought to provide regenerative triggers, but might also serve to satisfy energy demands. Liver is the major glucose provider, and hypoglycaemia inevitably develops when liver mass is lost. Indeed, hypoglycaemia is an essential regenerative signal [
21]. Moreover, hypoglycemia is thought to trigger a systemic response leading to a redistribution of lipids from the periphery into the regenerating liver [
22]. This could mean that delay or inhibition of proliferation is the consequence of hepatocyte functional impairment and not that of the cell cycle, persee. Indeed, mesenchymal stem cells have been reported to ameliorate hepatic dysfunction and improve liver regeneration after extended resection by paracrine, metabolic mechanisms [
23]. Additional analyses into the metabolic pathways underlying liver failure are currently being addressed in our laboratory. In summary, our comprehensive analyses not only recapitulated current knowledge on normal liver regeneration processes by retrieving Hedgehog-, Hypoxia-, and p21-associated pathways from thousands of data points, they also disclosed novel genes and surprising connections not earlier reported. In particular, the involvement of the
CD40-, the
BRCA1, and the
Ubiquitin-Proteasome Pathways are unexpectedly involved in the normal regeneration process, whereas the
IL10-, the
cAMP-, and the
ATM Pathways are defining the ceased regeneration process. The importance of miRNAs governing the molecular principles underlying liver regeneration should also be envisaged. Liver-specific signaling networks are tightly controlled by miRNAs that regulate key hepatic functions during liver injury and disease. Several miRNA affect hepatocyte proliferation, including miR-21, miR-29a, and miR-382, which target genes at important signaling crossroads, such as the crucial PI3K/AKT-signaling mediator PTEN [
24]. Still, miRNA data should be interpreted cautiously, because their expression depends on the genetic background, as well as on inter- and intra-species variability. The involvement of one—or a few—specific miRNA is not only very hard to establish, but also rather unlikely to be driving biological processes in which thousands of genes interact. In reality, extended arrays of differentially expressed miRNA determine the ultimate biological response.

 and
and 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


