Vascular Lipidomic Profiling of Potential Endogenous Fatty Acid PPAR Ligands Reveals the Coronary Artery as Major Producer of CYP450-Derived Epoxy Fatty Acids

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Vessel Organ Culture
2.3. Cell and Tissue Culture
2.4. Real-Time qRT-PCR
2.5. Oxylipin Measurements
2.6. Statistical Analyses
3. Results
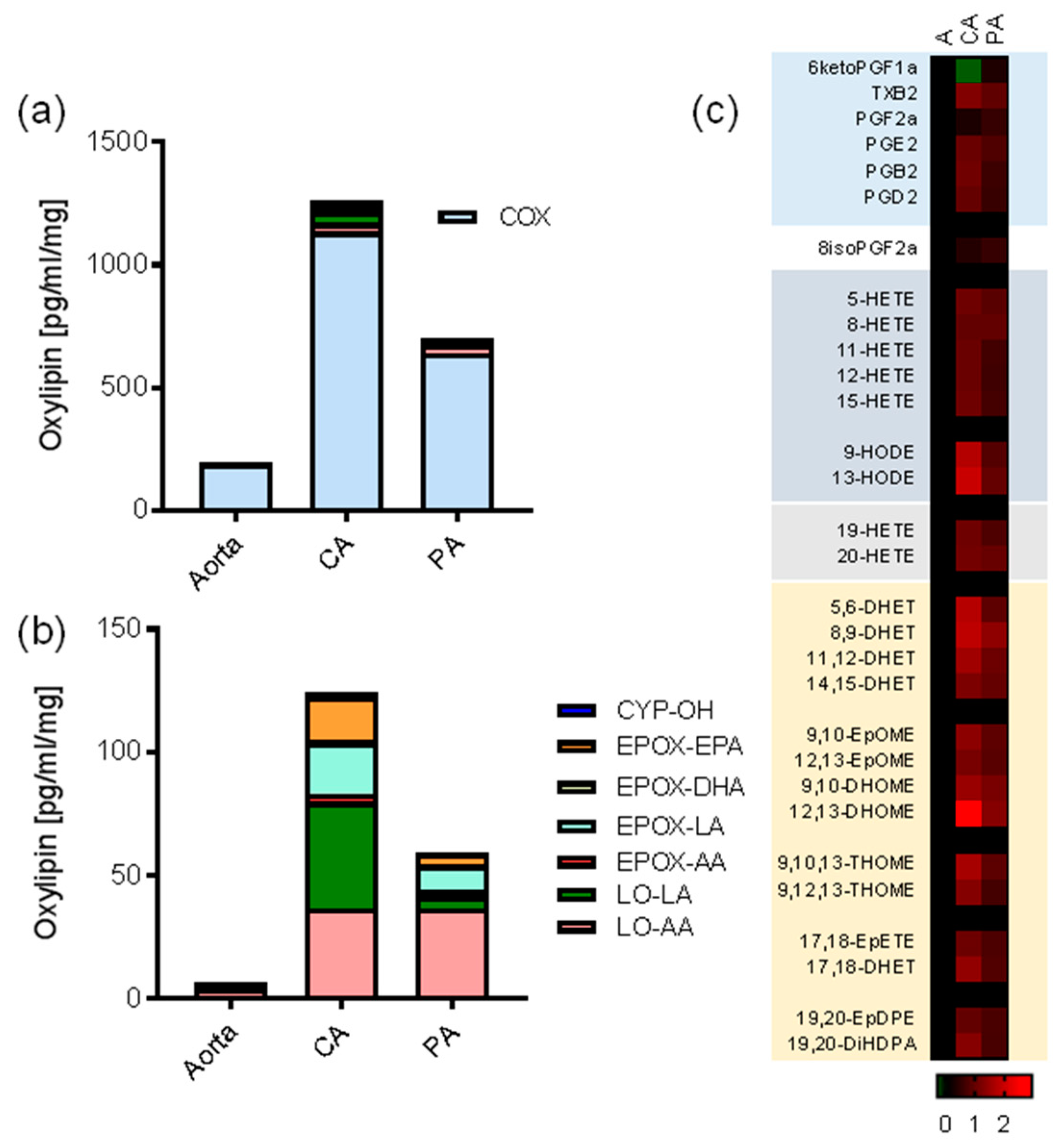
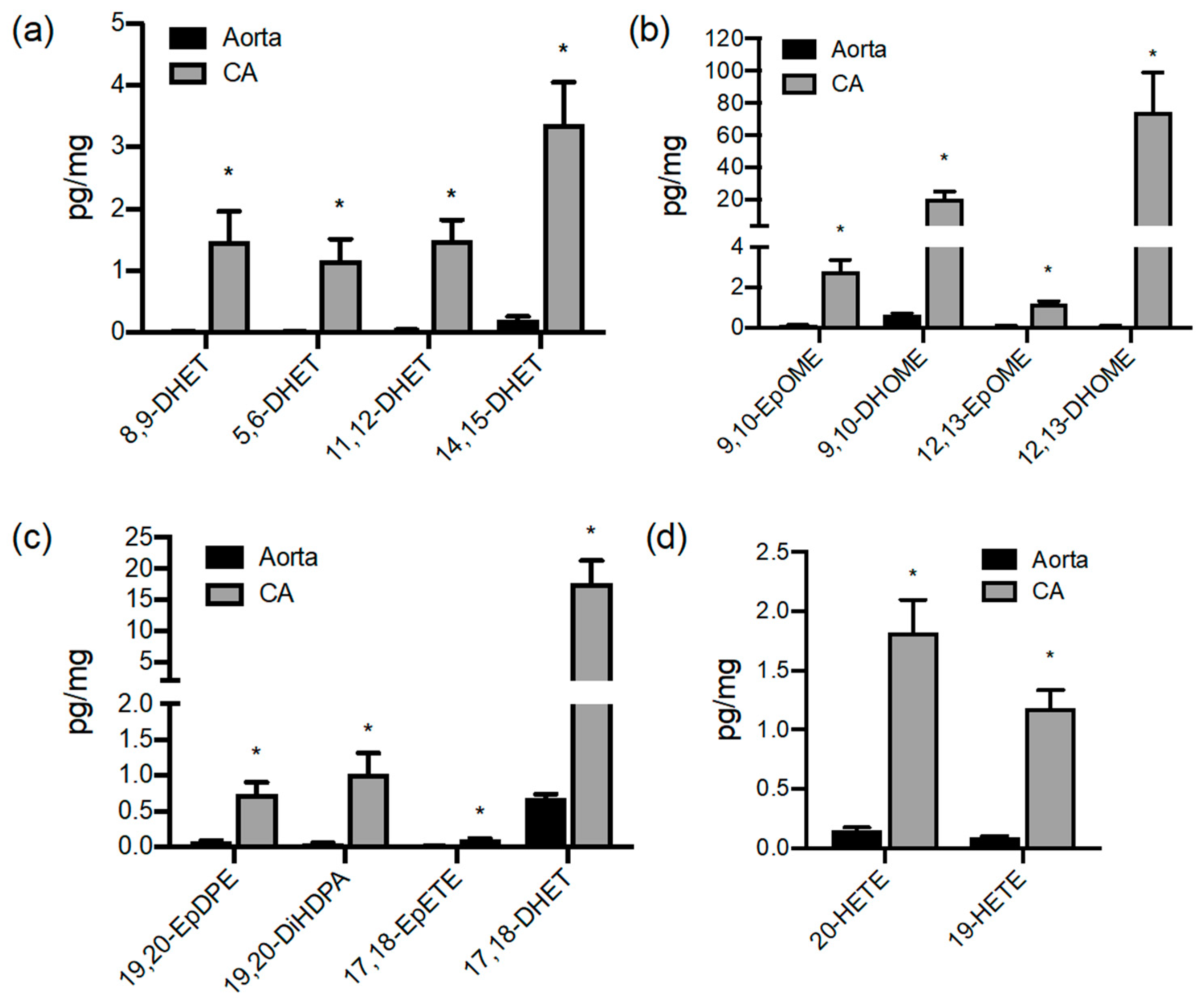
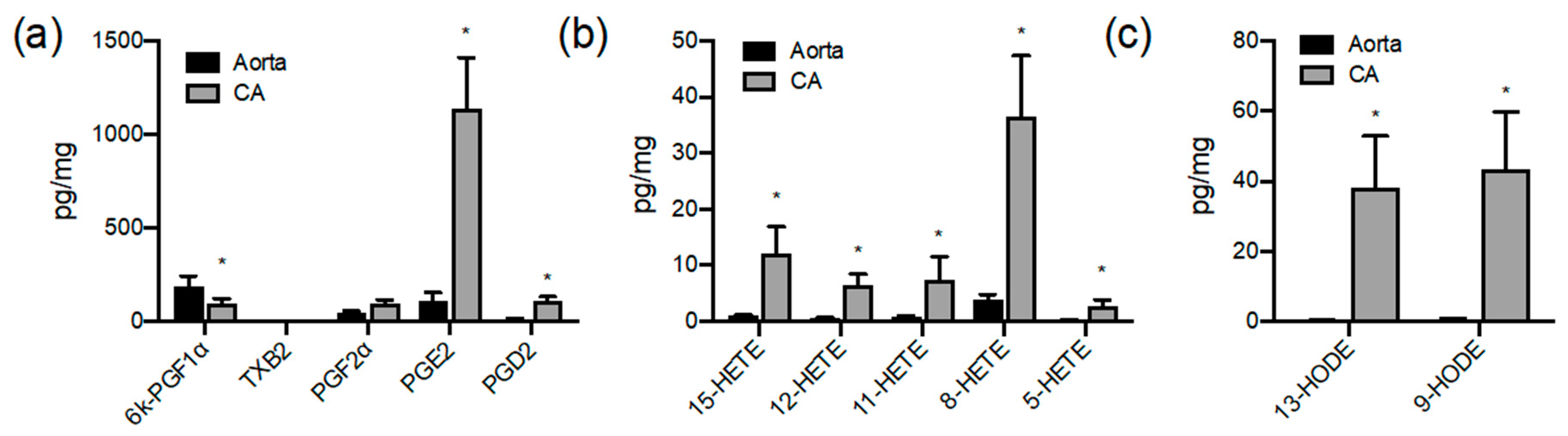
3.1. Oxylipin Lipidomic Profiling of the Large Vessels of the Pig
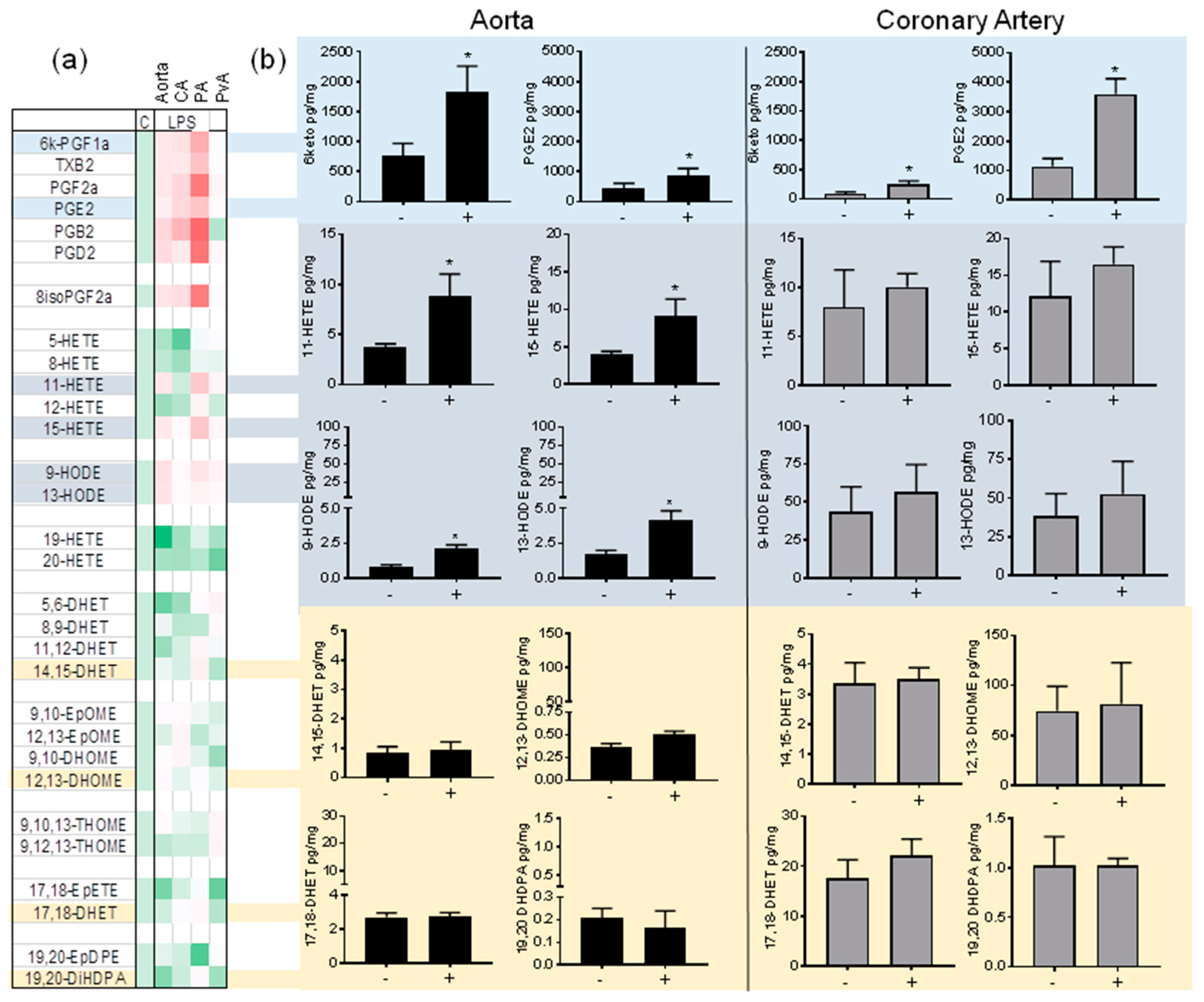
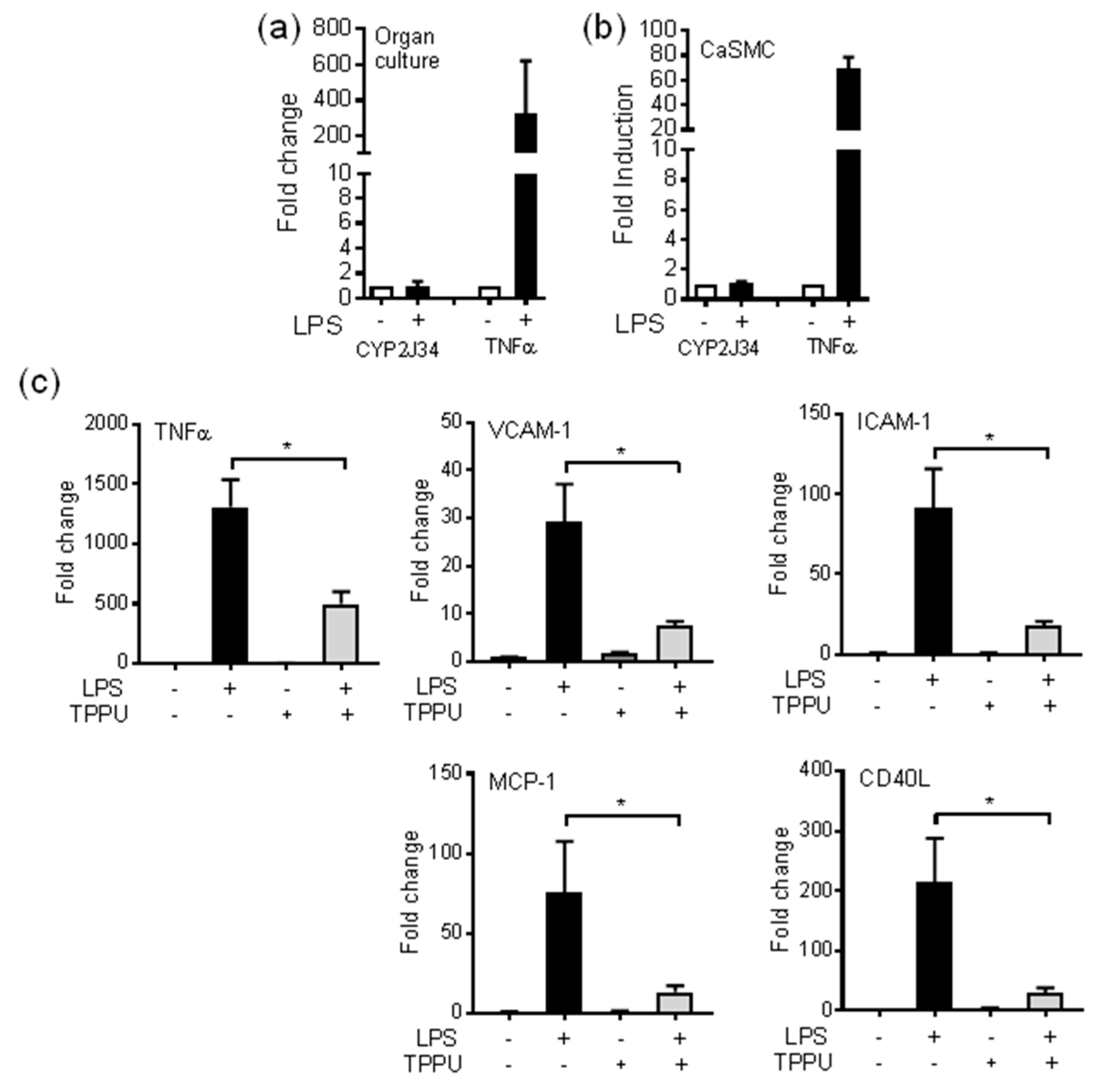
3.2. Regulation of Oxylipin Generation in the Large Vessels of the Pig by Inflammatory Stimuli: LPS/TLR4 Activation
3.3. The sEH Inhibitor TPPU Reduces TLR-4 Induced Inflammation in pCASMCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| CA | coronary artery |
| COX | cyclooxygenase |
| CYP | cytochrome P450 |
| DHA | docosahexaenoic acid |
| DHDPA | dihydroxydocosapentaenoic acid |
| DHET | dihydroxyeicosatrienoic acid |
| DHOME | dihydroxyoctadecamonoenoic acid |
| EET | epoxyeicosatrienoic acid |
| EPA | eicosapentaenoic acid |
| EpDPE | epoxydocosapentaenoic acid |
| EpETE | epoxyeicosatetraenoic acid |
| EpFA | epoxy fatty acid |
| EpOME | epoxyoctadecamonoenoic acid |
| HETE | hydroxyoctadecaenoic acid |
| HODE | hydroxyoctadecaenoic acid |
| LA | linoleic acid |
| LOX | lipoxygenase |
| PA | pulmonary artery |
| pCASMC | primary coronary artery smooth muscle cell |
| PPAR | peroxisome proliferator-activated receptor |
| PPRE | PPAR response element |
| PvA | perivascular adipose |
| sEH | soluble epoxide hydrolase |
| TPPU | (N-[1-(1-oxopropyl)-4-piperidinyl]-N’-[4-(trifluoromethoxy)phenyl)-urea) |
References
- Bishop-Bailey, D.; Wray, J. Peroxisome proliferator-activated receptors: A critical review on endogenous pathways for ligand generation. Prostaglandins Other Lipid Mediat. 2003, 71, 1–22. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; Zivkovic, A.M.; Wan, Y.J.; Watkins, S.M.; Nording, M.L.; Hammock, B.D.; German, J.B. Nutritional lipidomics: Molecular metabolism, analytics, and diagnostics. Mol. Nutr. Food Res. 2013, 57, 1319–1335. [Google Scholar] [CrossRef] [Green Version]
- Zeldin, D.C. Epoxygenase pathways of arachidonic acid metabolism. J. Biol. Chem. 2001, 276, 36059–36062. [Google Scholar] [CrossRef] [Green Version]
- Bishop-Bailey, D.; Thomson, S.; Askari, A.; Faulkner, A.; Wheeler-Jones, C. Lipid-metabolizing CYPs in the regulation and dysregulation of metabolism. Annu. Rev. Nutr. 2014, 34, 261–279. [Google Scholar] [CrossRef]
- Bailey, J.M.; Bryant, R.W.; Whiting, J.; Salata, K. Characterization of 11-HETE and 15-HETE, together with prostacyclin, as major products of the cyclooxygenase pathway in cultured rat aorta smooth muscle cells. J. Lipid Res. 1983, 24, 1419–1428. [Google Scholar]
- Thuresson, E.D.; Lakkides, K.M.; Smith, W.L. Different catalytically competent arrangements of arachidonic acid within the cyclooxygenase active site of prostaglandin endoperoxide H synthase-1 lead to the formation of different oxygenated products. J. Biol. Chem. 2000, 275, 8501–8507. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Moomaw, C.R.; Tomer, K.B.; Falck, J.R.; Zeldin, D.C. Molecular cloning and expression of CYP2J2, a human cytochrome P450 arachidonic acid epoxygenase highly expressed in heart. J. Biol. Chem. 1996, 271, 3460–3468. [Google Scholar] [CrossRef] [Green Version]
- Wray, J.A.; Sugden, M.C.; Zeldin, D.C.; Greenwood, G.K.; Samsuddin, S.; Miller-Degraff, L.; Bradbury, J.A.; Holness, M.J.; Warner, T.D.; Bishop-Bailey, D. The epoxygenases CYP2J2 activates the nuclear receptor PPARalpha in vitro and in vivo. PLoS ONE 2009, 4, e7421. [Google Scholar] [CrossRef]
- Edin, M.L.; Hamedani, B.G.; Gruzdev, A.; Graves, J.P.; Lih, F.B.; Arbes, S.J., 3rd; Singh, R.; Orjuela Leon, A.C.; Bradbury, J.A.; DeGraff, L.M.; et al. Epoxide hydrolase 1 (EPHX1) hydrolyzes epoxyeicosanoids and impairs cardiac recovery after ischemia. J. Biol. Chem. 2018, 293, 3281–3292. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.H.; Wecksler, A.T.; Wagner, K.; Hammock, B.D. Rationally designed multitarget agents against inflammation and pain. Curr. Med. Chem. 2013, 20, 1783–1799. [Google Scholar] [CrossRef]
- Imig, J.D.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases. Nat. Rev. Drug Discov. 2009, 8, 794–805. [Google Scholar] [CrossRef] [Green Version]
- Morisseau, C.; Hammock, B.D. Impact of soluble epoxide hydrolase and epoxyeicosanoids on human health. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 37–58. [Google Scholar] [CrossRef] [Green Version]
- Wray, J.; Bishop-Bailey, D. Epoxygenases and peroxisome proliferator-activated receptors in mammalian vascular biology. Exp. Physiol. 2008, 93, 148–154. [Google Scholar] [CrossRef]
- Bishop-Bailey, D.; Mitchell, J.A.; Warner, T.D. COX-2 in cardiovascular disease. Arter. Thromb. Vasc. Biol. 2006, 26, 956–958. [Google Scholar] [CrossRef] [Green Version]
- Baum, S.J.; Kris-Etherton, P.M.; Willett, W.C.; Lichtenstein, A.H.; Rudel, L.L.; Maki, K.C.; Whelan, J.; Ramsden, C.E.; Block, R.C. Fatty acids in cardiovascular health and disease: A comprehensive update. J. Clin. Lipidol. 2012, 6, 216–234. [Google Scholar] [CrossRef]
- Spiecker, M.; Liao, J.K. Vascular protective effects of cytochrome p450 epoxygenase-derived eicosanoids. Arch Biochem. Biophys. 2005, 433, 413–420. [Google Scholar] [CrossRef]
- Capra, V.; Back, M.; Barbieri, S.S.; Camera, M.; Tremoli, E.; Rovati, G.E. Eicosanoids and their drugs in cardiovascular diseases: Focus on atherosclerosis and stroke. Med. Res. Rev. 2013, 33, 364–438. [Google Scholar]
- Hersberger, M. Potential role of the lipoxygenase derived lipid mediators in atherosclerosis: Leukotrienes, lipoxins and resolvins. Clin. Chem. Lab. Med. 2010, 48, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Fleming, I. The factor in EDHF: Cytochrome P450 derived lipid mediators and vascular signaling. Vasc. Pharm. 2016, 86, 31–40. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Kirkby, N.S. Eicosanoids, prostacyclin and cyclooxygenase in the cardiovascular system. Br. J Pharm. 2019, 176, 1038–1050. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Warner, T.D. COX isoforms in the cardiovascular system: Understanding the activities of non-steroidal anti-inflammatory drugs. Nat. Rev. Drug Discov. 2006, 5, 75–86. [Google Scholar] [CrossRef]
- Chaudhary, K.R.; Zordoky, B.N.; Edin, M.L.; Alsaleh, N.; El-Kadi, A.O.; Zeldin, D.C.; Seubert, J.M. Differential effects of soluble epoxide hydrolase inhibition and CYP2J2 overexpression on postischemic cardiac function in aged mice. Prostaglandins Other Lipid Mediat. 2013, 104–105, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Edin, M.L.; Theken, K.N.; Schuck, R.N.; Flake, G.P.; Kannon, M.A.; Degraff, L.M.; Lih, F.B.; Foley, J.; Bradbury, J.A.; et al. Endothelial CYP epoxygenase overexpression and soluble epoxide hydrolase disruption attenuate acute vascular inflammatory responses in mice. Faseb. J. 2011, 25, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, Y.; Schmelzer, K.; Lee, T.S.; Fang, X.; Zhu, Y.; Spector, A.A.; Gill, S.; Morisseau, C.; Hammock, B.D.; et al. The antiinflammatory effect of laminar flow: The role of PPARgamma, epoxyeicosatrienoic acids, and soluble epoxide hydrolase. Proc. Natl. Acad. Sci. USA 2005, 102, 16747–16752. [Google Scholar] [CrossRef] [Green Version]
- Revermann, M.; Schloss, M.; Barbosa-Sicard, E.; Mieth, A.; Liebner, S.; Morisseau, C.; Geisslinger, G.; Schermuly, R.T.; Fleming, I.; Hammock, B.D.; et al. Soluble epoxide hydrolase deficiency attenuates neointima formation in the femoral cuff model of hyperlipidemic mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 909–914. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.N.; Vincelette, J.; Cheng, Y.; Mehra, U.; Chen, D.; Anandan, S.K.; Gless, R.; Webb, H.K.; Wang, Y.X. Inhibition of soluble epoxide hydrolase attenuated atherosclerosis, abdominal aortic aneurysm formation, and dyslipidemia. Arter. Thromb. Vasc. Biol. 2009, 29, 1265–1270. [Google Scholar] [CrossRef] [Green Version]
- Askari, A.; Thomson, S.J.; Edin, M.L.; Zeldin, D.C.; Bishop-Bailey, D. Roles of the epoxygenase CYP2J2 in the endothelium. Prostaglandins Other Lipid Mediat 2013, 107, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Bystrom, J.; Thomson, S.J.; Johansson, J.; Edin, M.L.; Zeldin, D.C.; Gilroy, D.W.; Smith, A.M.; Bishop-Bailey, D. Inducible CYP2J2 and its product 11,12-EET promotes bacterial phagocytosis: A role for CYP2J2 deficiency in the pathogenesis of Crohn’s disease? PLoS ONE 2013, 8, e75107. [Google Scholar] [CrossRef] [Green Version]
- Bystrom, J.; Wray, J.A.; Sugden, M.C.; Holness, M.J.; Swales, K.E.; Warner, T.D.; Edin, M.L.; Zeldin, D.C.; Gilroy, D.W.; Bishop-Bailey, D. Endogenous epoxygenases are modulators of monocyte/macrophage activity. PLoS ONE 2011, 6, e26591. [Google Scholar] [CrossRef] [Green Version]
- Gilroy, D.W.; Edin, M.L.; De Maeyer, R.P.; Bystrom, J.; Newson, J.; Lih, F.B.; Stables, M.; Zeldin, D.C.; Bishop-Bailey, D. CYP450-derived oxylipins mediate inflammatory resolution. Proc. Natl. Acad. Sci. USA 2016, 113, E3240–E3249. [Google Scholar] [CrossRef] [Green Version]
- Edin, M.L.; Wang, Z.; Bradbury, J.A.; Graves, J.P.; Lih, F.B.; DeGraff, L.M.; Foley, J.F.; Torphy, R.; Ronnekleiv, O.K.; Tomer, K.B.; et al. Endothelial expression of human cytochrome P450 epoxygenase CYP2C8 increases susceptibility to ischemia-reperfusion injury in isolated mouse heart. Faseb. J. 2011, 25, 3436–3447. [Google Scholar] [CrossRef] [Green Version]
- Fromel, T.; Jungblut, B.; Hu, J.; Trouvain, C.; Barbosa-Sicard, E.; Popp, R.; Liebner, S.; Dimmeler, S.; Hammock, B.D.; Fleming, I. Soluble epoxide hydrolase regulates hematopoietic progenitor cell function via generation of fatty acid diols. Proc. Natl. Acad. Sci. USA 2012, 109, 9995–10000. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, B.; Angioni, C.; Osthues, T.; Toewe, A.; Thomas, D.; Pierre, S.C.; Geisslinger, G.; Scholich, K.; Sisignano, M. The oxidized linoleic acid metabolite 12,13-DiHOME mediates thermal hyperalgesia during inflammatory pain. Biochim. Et. Biophys. Acta 2018, 1863, 669–678. [Google Scholar] [CrossRef]
- Moghaddam, M.F.; Grant, D.F.; Cheek, J.M.; Greene, J.F.; Williamson, K.C.; Hammock, B.D. Bioactivation of leukotoxins to their toxic diols by epoxide hydrolase. Nat. Med. 1997, 3, 562–566. [Google Scholar] [CrossRef]
- Sinal, C.J.; Miyata, M.; Tohkin, M.; Nagata, K.; Bend, J.R.; Gonzalez, F.J. Targeted disruption of soluble epoxide hydrolase reveals a role in blood pressure regulation. J. Biol. Chem. 2000, 275, 40504–40510. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D.; Zhao, X.; Capdevila, J.H.; Morisseau, C.; Hammock, B.D. Soluble epoxide hydrolase inhibition lowers arterial blood pressure in angiotensin II hypertension. Hypertension 2002, 39, 690–694. [Google Scholar] [CrossRef] [Green Version]
- Luria, A.; Bettaieb, A.; Xi, Y.; Shieh, G.J.; Liu, H.C.; Inoue, H.; Tsai, H.J.; Imig, J.D.; Haj, F.G.; Hammock, B.D. Soluble epoxide hydrolase deficiency alters pancreatic islet size and improves glucose homeostasis in a model of insulin resistance. Proc. Natl. Acad. Sci. USA 2011, 108, 9038–9043. [Google Scholar] [CrossRef] [Green Version]
- Revermann, M.; Barbosa-Sicard, E.; Dony, E.; Schermuly, R.T.; Morisseau, C.; Geisslinger, G.; Fleming, I.; Hammock, B.D.; Brandes, R.P. Inhibition of the soluble epoxide hydrolase attenuates monocrotaline-induced pulmonary hypertension in rats. J. Hypertens. 2009, 27, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, J.; Guo, L.; Uyeminami, D.; Dong, H.; Hammock, B.D.; Pinkerton, K.E. Use of a soluble epoxide hydrolase inhibitor in smoke-induced chronic obstructive pulmonary disease. Am. J. Respir. Cell Mol. Biol. 2012, 46, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Podolin, P.L.; Bolognese, B.J.; Foley, J.F.; Long, E., 3rd; Peck, B.; Umbrecht, S.; Zhang, X.; Zhu, P.; Schwartz, B.; Xie, W.; et al. In vitro and in vivo characterization of a novel soluble epoxide hydrolase inhibitor. Prostaglandins Other Lipid Mediat. 2013, 104–105, 25–31. [Google Scholar] [CrossRef]
- Hanif, A.; Edin, M.L.; Zeldin, D.C.; Morisseau, C.; Falck, J.R.; Nayeem, M.A. Vascular endothelial overexpression of human CYP2J2 (Tie2-CYP2J2 Tr) modulates cardiac oxylipin profiles and enhances coronary reactive hyperemia in mice. PLoS ONE 2017, 12, e0174137. [Google Scholar] [CrossRef] [Green Version]
- Hanif, A.; Edin, M.L.; Zeldin, D.C.; Morisseau, C.; Falck, J.R.; Nayeem, M.A. Vascular Endothelial Over-Expression of Human Soluble Epoxide Hydrolase (Tie2-sEH Tr) Attenuates Coronary Reactive Hyperemia in Mice: Role of Oxylipins and omega-Hydroxylases. PLoS ONE 2017, 12, e0169584. [Google Scholar] [CrossRef]
- Hanif, A.; Edin, M.L.; Zeldin, D.C.; Morisseau, C.; Nayeem, M.A. Effect of Soluble Epoxide Hydrolase on the Modulation of Coronary Reactive Hyperemia: Role of Oxylipins and PPARgamma. PLoS ONE 2016, 11, e0162147. [Google Scholar] [CrossRef]
- Hanif, A.; Edin, M.L.; Zeldin, D.C.; Morisseau, C.; Nayeem, M.A. Deletion of soluble epoxide hydrolase enhances coronary reactive hyperemia in isolated mouse heart: Role of oxylipins and PPARgamma. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R676–R688. [Google Scholar] [CrossRef] [Green Version]
- Nayeem, M.A.; Pradhan, I.; Mustafa, S.J.; Morisseau, C.; Falck, J.R.; Zeldin, D.C. Adenosine A2A receptor modulates vascular response in soluble epoxide hydrolase-null mice through CYP-epoxygenases and PPARgamma. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 304, R23–R32. [Google Scholar] [CrossRef] [Green Version]
- Samokhvalov, V.; Vriend, J.; Jamieson, K.L.; Akhnokh, M.K.; Manne, R.; Falck, J.R.; Seubert, J.M. PPAR gamma signaling is required for mediating EETs protective effects in neonatal cardiomyocytes exposed to LPS. Front Pharmacol. 2014, 5, 242. [Google Scholar] [CrossRef]
- He, Z.; Zhang, X.; Chen, C.; Wen, Z.; Hoopes, S.L.; Zeldin, D.C.; Wang, D.W. Cardiomyocyte-specific expression of CYP2J2 prevents development of cardiac remodelling induced by angiotensin II. Cardiovasc. Res. 2015, 105, 304–317. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Zhao, G.; Yan, J.; Liu, W.; Feng, W.; Ma, B.; Yang, L.; Wang, J.A.; Tu, L.; Wang, D.W. CYP2J2 overexpression increases EETs and protects against angiotensin II-induced abdominal aortic aneurysm in mice. J. Lipid Res. 2013, 54, 1448–1456. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Imig, J.D.; Yang, J.; Hammock, B.D.; Padanilam, B.J. Inhibition of soluble epoxide hydrolase prevents renal interstitial fibrosis and inflammation. Am. J. Physiol. Renal Physiol. 2014, 307, F971–F980. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D.; Walsh, K.A.; Hye Khan, M.A.; Nagasawa, T.; Cherian-Shaw, M.; Shaw, S.M.; Hammock, B.D. Soluble epoxide hydrolase inhibition and peroxisome proliferator activated receptor gamma agonist improve vascular function and decrease renal injury in hypertensive obese rats. Exp. Biol. Med. (Maywood) 2012, 237, 1402–1412. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.Y.; Davis, B.B.; Wang, Z.H.; Zhao, S.P.; Wasti, B.; Liu, Z.L.; Li, N.; Morisseau, C.; Chiamvimonvat, N.; Hammock, B.D. A potent soluble epoxide hydrolase inhibitor, t-AUCB, acts through PPARgamma to modulate the function of endothelial progenitor cells from patients with acute myocardial infarction. Int. J. Cardiol. 2012, 167, 1298–1304. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.; Al-Mashhadi, R.H.; Sorensen, C.B.; Bentzon, J.F. Large animal models of atherosclerosis--new tools for persistent problems in cardiovascular medicine. J. Pathol. 2016, 238, 257–266. [Google Scholar] [CrossRef]
- Tanaka, K.; Sata, M. Roles of Perivascular Adipose Tissue in the Pathogenesis of Atherosclerosis. Front. Physiol. 2018, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Huang Cao, Z.F.; Stoffel, E.; Cohen, P. Role of Perivascular Adipose Tissue in Vascular Physiology and Pathology. Hypertension 2017, 69, 770–777. [Google Scholar] [CrossRef]
- Liu, J.Y.; Lin, Y.P.; Qiu, H.; Morisseau, C.; Rose, T.E.; Hwang, S.H.; Chiamvimonvat, N.; Hammock, B.D. Substituted phenyl groups improve the pharmacokinetic profile and anti-inflammatory effect of urea-based soluble epoxide hydrolase inhibitors in murine models. Eur. J. Pharm. Sci. 2013, 48, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Bishop-Bailey, D.; Larkin, S.W.; Warner, T.D.; Chen, G.; Mitchell, J.A. Characterization of the induction of nitric oxide synthase and cyclo-oxygenase in rat aorta in organ culture. Br. J. Pharm. 1997, 121, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Bishop-Bailey, D.; Pepper, J.R.; Haddad, E.B.; Newton, R.; Larkin, S.W.; Mitchell, J.A. Induction of cyclooxygenase-2 in human saphenous vein and internal mammary artery. Arter. Thromb. Vasc. Biol. 1997, 17, 1644–1648. [Google Scholar] [CrossRef]
- Bishop-Bailey, D.; Pepper, J.R.; Larkin, S.W.; Mitchell, J.A. Differential induction of cyclooxygenase-2 in human arterial and venous smooth muscle: Role of endogenous prostanoids. Arter. Thromb. Vasc. Biol. 1998, 18, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.R.; Imig, J.D.; Edin, M.L.; Foley, J.; DeGraff, L.M.; Bradbury, J.A.; Graves, J.P.; Lih, F.B.; Clark, J.; Myers, P.; et al. Endothelial expression of human cytochrome P450 epoxygenases lowers blood pressure and attenuates hypertension-induced renal injury in mice. Faseb. J. 2010, 24, 3770–3781. [Google Scholar] [CrossRef] [Green Version]
- Hecker, M.; Bara, A.T.; Bauersachs, J.; Busse, R. Characterization of endothelium-derived hyperpolarizing factor as a cytochrome P450-derived arachidonic acid metabolite in mammals. J. Physiol. 1994, 481, 407–414. [Google Scholar] [CrossRef]
- Fang, X.; Kaduce, T.L.; Weintraub, N.L.; Harmon, S.; Teesch, L.M.; Morisseau, C.; Thompson, D.A.; Hammock, B.D.; Spector, A.A. Pathways of epoxyeicosatrienoic acid metabolism in endothelial cells. Implications for the vascular effects of soluble epoxide hydrolase inhibition. J. Biol. Chem. 2001, 276, 14867–14874. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Weintraub, N.L.; Oltman, C.L.; Stoll, L.L.; Kaduce, T.L.; Harmon, S.; Dellsperger, K.C.; Morisseau, C.; Hammock, B.D.; Spector, A.A. Human coronary endothelial cells convert 14,15-EET to a biologically active chain-shortened epoxide. Am. J. Physiol. 2002, 283, H2306–H2314. [Google Scholar] [CrossRef] [Green Version]
- Cowart, L.A.; Wei, S.; Hsu, M.H.; Johnson, E.F.; Krishna, M.U.; Falck, J.R.; Capdevila, J.H. The CYP4A isoforms hydroxylate epoxyeicosatrienoic acids to form high affinity peroxisome proliferator-activated receptor ligands. J. Biol. Chem. 2002, 277, 35105–35112. [Google Scholar]
- Zarbock, A.; Distasi, M.R.; Smith, E.; Sanders, J.M.; Kronke, G.; Harry, B.L.; Von Vietinghoff, S.; Buscher, K.; Nadler, J.L.; Ley, K. Improved survival and reduced vascular permeability by eliminating or blocking 12/15-lipoxygenase in mouse models of acute lung injury (ALI). J. Immunol. 2009, 183, 4715–4722. [Google Scholar]
- Serio, K.J.; Reddy, K.V.; Bigby, T.D. Lipopolysaccharide induces 5-lipoxygenase-activating protein gene expression in THP-1 cells via a NF-kappaB and C/EBP-mediated mechanism. Am. J. Physiol. Cell Physiol. 2005, 288, C1125–C1133. [Google Scholar] [CrossRef] [Green Version]
- Kirkby, N.S.; Reed, D.M.; Edin, M.L.; Rauzi, F.; Mataragka, S.; Vojnovic, I.; Bishop-Bailey, D.; Milne, G.L.; Longhurst, H.; Zeldin, D.C.; et al. Inherited human group IVA cytosolic phospholipase A2 deficiency abolishes platelet, endothelial, and leucocyte eicosanoid generation. Faseb. J. 2015, 29, 4568–4578. [Google Scholar] [CrossRef] [Green Version]
- Apaya, M.K.; Lin, C.Y.; Chiou, C.Y.; Yang, C.C.; Ting, C.Y.; Shyur, L.F. Simvastatin and a plant galactolipid protect animals from septic shock by regulating oxylipin mediator dynamics through the MAPK-cPLA2 signaling pathway. Mol. Med. 2015, 21, 988–1001. [Google Scholar]
- Arnold, C.; Markovic, M.; Blossey, K.; Wallukat, G.; Fischer, R.; Dechend, R.; Konkel, A.; Von Schacky, C.; Luft, F.C.; Muller, D.N.; et al. Arachidonic acid-metabolizing cytochrome P450 enzymes are targets of 1-3 fatty acids. J. Biol. Chem. 2010, 285, 32720–32733. [Google Scholar]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; Von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profile in man primarily via the CYP-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar]
- Han, L.; Shen, W.J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of metabolism and as therapeutic targets in cardiovascular disease. Part I: PPAR-α. Future Cardiol. 2017, 13, 259–278. [Google Scholar]
- Han, L.; Shen, W.J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of metabolism and as therapeutic targets in cardiovascular disease. Part II: PPAR-β/δ and PPAR-γ. Future Cardiol. 2017, 13, 279–296. [Google Scholar]
- Luo, P.; Chang, H.H.; Zhou, Y.; Zhang, S.; Hwang, S.H.; Morisseau, C.; Wang, C.Y.; Inscho, E.W.; Hammock, B.D.; Wang, M.H. Inhibition or deletion of soluble epoxide hydrolase prevents hyperglycemia, promotes insulin secretion, and reduces islet apoptosis. J. Phar. Exp. Ther. 2010, 334, 430–438. [Google Scholar]
- Xiao, B.; Li, X.; Yan, J.; Yu, X.; Yang, G.; Xiao, X.; Voltz, J.W.; Zeldin, D.C.; Wang, D.W. Overexpression of cytochrome P450 epoxygenases prevents development of hypertension in spontaneously hypertensive rats by enhancing atrial natriuretic peptide. J. Phar. Exp. Ther. 2010, 334, 784–794. [Google Scholar]
- Oni-Orisan, A.; Edin, M.L.; Lee, J.A.; Wells, M.A.; Christensen, E.S.; Vendrov, K.C.; Lih, F.B.; Tomer, K.B.; Bai, X.; Taylor, J.M.; et al. Cytochrome P450-derived epoxyeicosatrienoic acids and coronary artery disease in humans: A targeted metabolomics study. J. Lipid Res. 2016, 57, 109–119. [Google Scholar]
- Schuck, R.N.; Theken, K.N.; Edin, M.L.; Caughey, M.; Bass, A.; Ellis, K.; Tran, B.; Steele, S.; Simmons, B.P.; Lih, F.B.; et al. Cytochrome P450-derived eicosanoids and vascular dysfunction in coronary artery disease patients. Atherosclerosis 2013, 227, 442–448. [Google Scholar]
- Ng, V.Y.; Huang, Y.; Reddy, L.M.; Falck, J.R.; Lin, E.T.; Kroetz, D.L. Cytochrome P450 eicosanoids are activators of peroxisome proliferator-activated receptor alpha. Drug Metab. Dispos. 2007, 35, 1126–1134. [Google Scholar]
- Messina, A.; Siniscalco, A.; Puccinelli, E.; Gervasi, P.G.; Longo, V. Cloning and tissues expression of the pig CYP1B1 and CYP2J34. Res. Vet. Sci. 2012, 92, 438–443. [Google Scholar]
- Askari, A.A.; Thomson, S.; Edin, M.L.; Lih, F.B.; Zeldin, D.C.; Bishop-Bailey, D. Basal and inducible anti-inflammatory epoxygenase activity in endothelial cells. Biochem. Biophys. Res. Commun. 2014, 446, 633–637. [Google Scholar]
- Bishop-Bailey, D.; Hla, T.; Warner, T.D. Intimal smooth muscle cells as a target for peroxisome proliferator-activated receptor-gamma ligand therapy. Circ. Res. 2002, 91, 210–217. [Google Scholar]
- Thomson, S.; Edin, M.L.; Lih, F.B.; Davies, M.; Yaqoob, M.M.; Hammock, B.D.; Gilroy, D.; Zeldin, D.C.; Bishop-Bailey, D. Intimal smooth muscle cells are a source but not a sensor of anti-inflammatory CYP450 derived oxylipins. Biochem. Biophys. Res. Commun. 2015, 463, 774–780. [Google Scholar]
- Del Pozo, R.; Hernandez Gonzalez, I.; Escribano-Subias, P. The prostacyclin pathway in pulmonary arterial hypertension: A clinical review. Expert. Rev. Respir. Med. 2017, 11, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.Y.; Egan, K.; FitzGerald, G.A.; Desvergne, B.; Wahli, W.; Bishop-Bailey, D.; Warner, T.D.; Mitchell, J.A. Role of prostacyclin versus peroxisome proliferator-activated receptor beta receptors in prostacyclin sensing by lung fibroblasts. Am. J. Respir. Cell Mol. Biol. 2006, 34, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Pokreisz, P.; Fleming, I.; Kiss, L.; Barbosa-Sicard, E.; Fisslthaler, B.; Falck, J.R.; Hammock, B.D.; Kim, I.H.; Szelid, Z.; Vermeersch, P.; et al. Cytochrome P450 epoxygenase gene function in hypoxic pulmonary vasoconstriction and pulmonary vascular remodeling. Hypertension 2006, 47, 762–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Wang, L.; Li, R.; Ma, B.; Tu, L.; Xu, X.; Dackor, R.T.; Zeldin, D.C.; Wang, D.W. Gene delivery of cytochrome p450 epoxygenase ameliorates monocrotaline-induced pulmonary artery hypertension in rats. Am. J. Respir. Cell Mol. Biol. 2010, 43, 740–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edin, M.L.; Lih, F.B.; Hammock, B.D.; Thomson, S.; Zeldin, D.C.; Bishop-Bailey, D. Vascular Lipidomic Profiling of Potential Endogenous Fatty Acid PPAR Ligands Reveals the Coronary Artery as Major Producer of CYP450-Derived Epoxy Fatty Acids. Cells 2020, 9, 1096. https://doi.org/10.3390/cells9051096
Edin ML, Lih FB, Hammock BD, Thomson S, Zeldin DC, Bishop-Bailey D. Vascular Lipidomic Profiling of Potential Endogenous Fatty Acid PPAR Ligands Reveals the Coronary Artery as Major Producer of CYP450-Derived Epoxy Fatty Acids. Cells. 2020; 9(5):1096. https://doi.org/10.3390/cells9051096
Chicago/Turabian StyleEdin, Matthew L., Fred B. Lih, Bruce D. Hammock, Scott Thomson, Darryl C. Zeldin, and David Bishop-Bailey. 2020. "Vascular Lipidomic Profiling of Potential Endogenous Fatty Acid PPAR Ligands Reveals the Coronary Artery as Major Producer of CYP450-Derived Epoxy Fatty Acids" Cells 9, no. 5: 1096. https://doi.org/10.3390/cells9051096