Characterization of Sialic Acid-Binding Immunoglobulin-Type Lectins in Fish Reveals Teleost-Specific Structures and Expression Patterns

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Husbandry and Experimental Treatment
2.2. Isolation and Microscopic Characterization of Head Kidney Cells from Maraena Whitefish
2.3. Flow Cytometry
2.4. RNA Isolation
2.5. Primer Design and Quantitative PCR
2.6. Cloning
2.7. In Silico Analyses
3. Results and Discussion
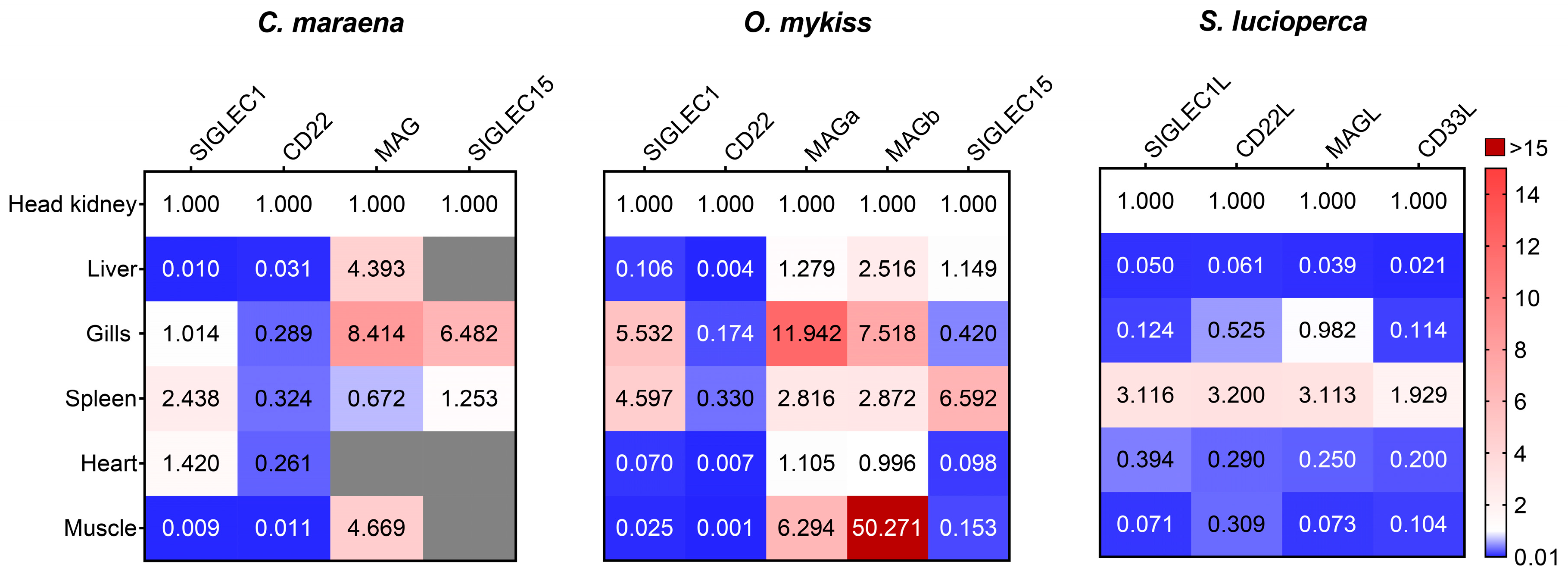
3.1. The Expression of Siglec-Encoding Genes in Different Tissues of Salmonid and Percid Fishes
3.2. The Expression Patterns of Siglec-Encoding Genes in Cell Populations of Maraena Whitefish
3.3. Sequence Comparison of Siglecs Expressed by Salmonid and Percid Fishes
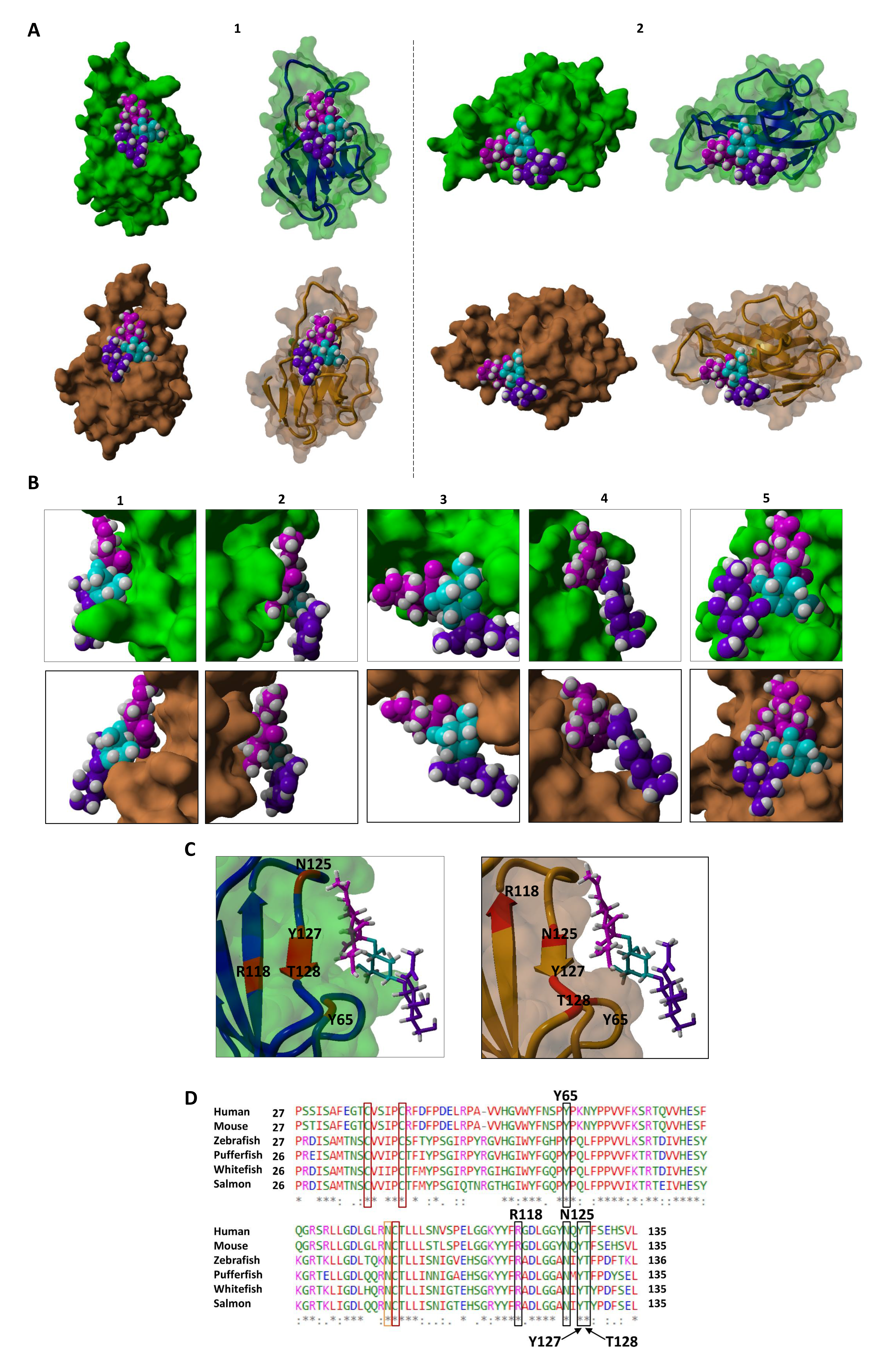
3.3.1. Sequence Comparison of Siglec2 (CD22)
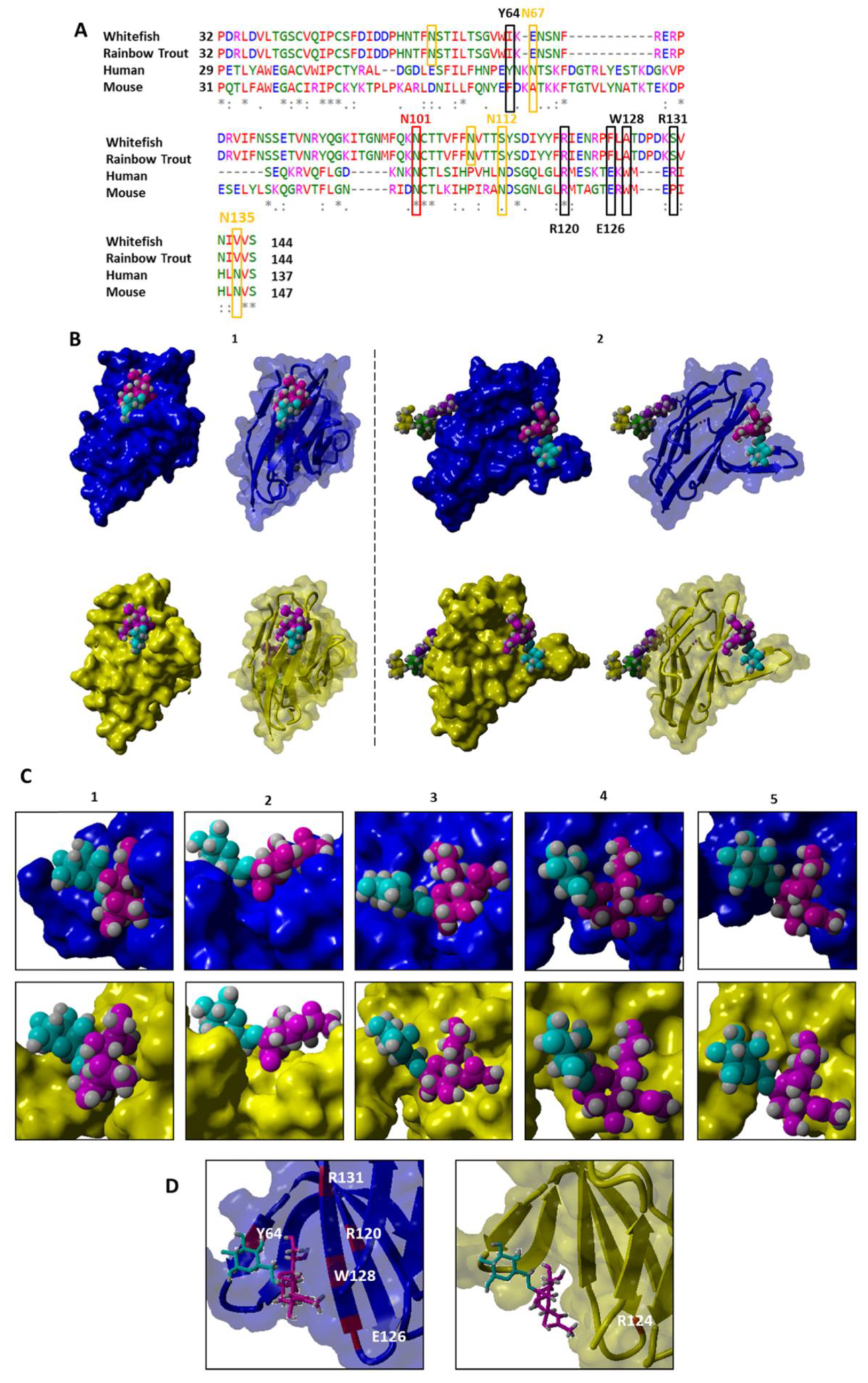
3.3.2. Sequence Comparison of Siglec15
3.3.3. Sequence Comparison of Siglec4 (MAG)
3.4. The Influence of Handling Stress on the Expression of Siglecs in Maraena Whitefish
4. Conclusions
- Our qPCR analyses suggested that the basal gene-expression patterns of Siglec1, CD22, MAG, and Siglec15 are largely conserved across salmonid and percid fishes. In contrast to mammals, CD22 is highly expressed in several blood-cell populations. Similarly, the expression of MAG in fish is not restricted to the cells of the nervous system but is detectable in a range of blood cells.
- Stress modulates the expression of Siglecs (but not of the associated enzymes) in a tissue-dependent fashion and most likely influences the cellular reactivity against PAMPs and DAMPs.
- The genomes of fish lack CD33-related Siglecs, which exert inhibitory functions. Our structural analyses indicated that CD22 and MAG contain inhibitory motifs (ITIM) in salmonid fish. We speculate that these ITIM-containing Siglecs may compensate the deficiency of the canonical inhibitory Siglecs. This first assumption might be the starting point for subsequent studies to clarify whether CD22 and MAG have an immunosuppressive effect in fish.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Uribe, C.; Folch, H.; Enríquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486–503. [Google Scholar] [CrossRef] [Green Version]
- Rebl, A.; Goldammer, T. Under control: The innate immunity of fish from the inhibitors’ perspective. Fish Shellfish Immunol. 2018, 77, 328–349. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Are humans prone to autoimmunity? Implications from evolutionary changes in hominin sialic acid biology. J. Autoimmun. 2017, 83, 134–142. [Google Scholar] [CrossRef]
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varki, A. Letter to the glyco-forum: Since there are pamps and damps, there must be samps? Glycan “self-associated molecular patterns” dampen innate immunity, but pathogens can mimic them. Glycobiology 2011, 21, 1121–1124. [Google Scholar] [CrossRef]
- Bloemendal, V.R.L.J.; Moons, S.J.; Heming, J.J.A.; Chayoua, M.; Niesink, O.; van Hest, J.C.M.; Boltje, T.J.; Rutjes, F.P.J.T. Chemoenzymatic synthesis of sialic acid derivatives using immobilized n-acetylneuraminate lyase in a continuous flow reactor. Adv. Synth. Catal. 2019, 361, 2443–2447. [Google Scholar] [CrossRef] [Green Version]
- Coady, A.; Nizet, V. Samp-ending down sepsis. Ann. Transl. Med. 2016, 4, 509. [Google Scholar] [CrossRef]
- Varki, A.; Angata, T. Siglecs—The major subfamily of i-type lectins. Glycobiology 2006, 16, 1R–27R. [Google Scholar] [CrossRef] [Green Version]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef]
- Angata, T.; Margulies, E.H.; Green, E.D.; Varki, A. Large-scale sequencing of the cd33-related siglec gene cluster in five mammalian species reveals rapid evolution by multiple mechanisms. Proc. Natl. Acad. Sci. USA 2004, 101, 13251–13256. [Google Scholar] [CrossRef] [Green Version]
- Ravetch, J.V.; Lanier, L.L. Immune inhibitory receptors. Science 2000, 290, 84–89. [Google Scholar] [CrossRef]
- Pillai, S.; Netravali, I.A.; Cariappa, A.; Mattoo, H. Siglecs and immune regulation. Annu. Rev. Immunol. 2012, 30, 357–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, M.K.; Paulson, J.C. Siglecs as targets for therapy in immune-cell-mediated disease. Trends Pharmacol. Sci. 2009, 30, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crocker, P.R.; Varki, A. Siglecs in the immune system. Immunology 2001, 103, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Bornhöfft, K.F.; Goldammer, T.; Rebl, A.; Galuska, S.P. Siglecs: A journey through the evolution of sialic acid-binding immunoglobulin-type lectins. Dev. Comp. Immunol. 2018, 86, 219–231. [Google Scholar] [CrossRef]
- Lehmann, F.; Gathje, H.; Kelm, S.; Dietz, F. Evolution of sialic acid-binding proteins: Molecular cloning and expression of fish siglec-4. Glycobiology 2004, 14, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Angata, T.; Tabuchi, Y.; Nakamura, K.; Nakamura, M. Siglec-15: An immune system siglec conserved throughout vertebrate evolution. Glycobiology 2007, 17, 838–846. [Google Scholar] [CrossRef]
- Robertson, F.M.; Gundappa, M.K.; Grammes, F.; Hvidsten, T.R.; Redmond, A.K.; Lien, S.; Martin, S.A.M.; Holland, P.W.H.; Sandve, S.R.; Macqueen, D.J. Lineage-specific rediploidization is a mechanism to explain time-lags between genome duplication and evolutionary diversification. Genome Biol. 2017, 18, 111. [Google Scholar] [CrossRef] [Green Version]
- Korytar, T.; Nipkow, M.; Altmann, S.; Goldammer, T.; Kollner, B.; Rebl, A. Adverse husbandry of maraena whitefish directs the immune system to increase mobilization of myeloid cells and proinflammatory responses. Front. Immunol. 2016, 7, 631. [Google Scholar] [CrossRef] [Green Version]
- Rebl, A.; Verleih, M.; Nipkow, M.; Altmann, S.; Bochert, R.; Goldammer, T. Gradual and acute temperature rise induces crossing endocrine, metabolic, and immunological pathways in maraena whitefish (coregonus maraena). Front. Genet. 2018, 9, 241. [Google Scholar] [CrossRef]
- Swirplies, F.; Wuertz, S.; Baßmann, B.; Orban, A.; Schäfer, N.; Brunner, R.M.; Hadlich, F.; Goldammer, T.; Rebl, A. Identification of molecular stress indicators in pikeperch sander lucioperca correlating with rising water temperatures. Aquaculture 2019, 501, 260–271. [Google Scholar] [CrossRef]
- Rebl, A.; Rebl, H.; Verleih, M.; Haupt, S.; Kobis, J.M.; Goldammer, T.; Seyfert, H.M. At least two genes encode many variants of irak3 in rainbow trout, but neither the full-length factor nor its variants interfere directly with the tlr-mediated stimulation of inflammation. Front. Immunol. 2019, 10, 2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorell Ribera, J.; Nipkow, M.; Viergutz, T.; Brunner, R.M.; Bochert, R.; Koll, R.; Goldammer, T.; Gimsa, U.; Rebl, A. Early response of salmonid head-kidney cells to stress hormones and toll-like receptor ligands. Fish Shellfish Immunol. 2020, 98, 950–961. [Google Scholar] [CrossRef] [PubMed]
- Nguinkal, J.A.; Brunner, R.M.; Verleih, M.; Rebl, A.; de Los Rios-Perez, L.; Schafer, N.; Hadlich, F.; Stueken, M.; Wittenburg, D.; Goldammer, T. The first highly contiguous genome assembly of pikeperch (sander lucioperca), an emerging aquaculture species in europe. Genes (Basel) 2019, 10, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmann, S.; Rebl, A.; Kuhn, C.; Goldammer, T. Identification and de novo sequencing of housekeeping genes appropriate for gene expression analyses in farmed maraena whitefish (coregonus maraena) during crowding stress. Fish Physiol. Biochem. 2015, 41, 397–412. [Google Scholar] [CrossRef]
- Rebl, A.; Rebl, H.; Korytar, T.; Goldammer, T.; Seyfert, H.M. The proximal promoter of a novel interleukin-8-encoding gene in rainbow trout (oncorhynchus mykiss) is strongly induced by cebpa, but not nf-kappab p65. Dev. Comp. Immunol. 2014, 46, 155–164. [Google Scholar] [CrossRef]
- Rebl, A.; Anders, E.; Wimmers, K.; Goldammer, T. Characterization of dehydrodolichyl diphosphate synthase gene in rainbow trout (oncorhynchus mykiss). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 260–265. [Google Scholar] [CrossRef]
- Brietzke, A.; Borchel, A.; Altmann, S.; Nipkow, M.; Rebl, A.; Brunner, R.M.; Goldammer, T. Transcriptome sequencing of maraena whitefish (coregonus maraena). Mar. Genomics 2016, 29, 27–29. [Google Scholar] [CrossRef]
- Ereño-Orbea, J.; Sicard, T.; Cui, H.; Mazhab-Jafari, M.T.; Benlekbir, S.; Guarné, A.; Rubinstein, J.L.; Julien, J.P. Molecular basis of human cd22 function and therapeutic targeting. Nat. Commun. 2017, 8, 764. [Google Scholar] [CrossRef]
- Pronker, M.F.; Lemstra, S.; Snijder, J.; Heck, A.J.R.; Thies-Weesie, D.M.E.; Pasterkamp, R.J.; Janssen, B.J.C. Structural basis of myelin-associated glycoprotein adhesion and signalling. Nat. Commun. 2016, 7, 13584. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Olson, J.; Louie, A.; Crocker, P.R.; Varki, A.; Nizet, V. Role of macrophage sialoadhesin in host defense against the sialylated pathogen group b streptococcus. J. Mol. Med. (Berl). 2014, 92, 951–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, A.; Diez, B.; Cejalvo, T.; Gutierrez-de Frias, C.; Cortes, A. Ontogeny of the immune system of fish. Fish Shellfish Immunol. 2006, 20, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Macauley, M.S.; Crocker, P.R.; Paulson, J.C. Siglec regulation of immune cell function in disease. Nat. Rev. Immunol. 2014, 14, 653–666. [Google Scholar] [CrossRef] [Green Version]
- Quarles, R.H. Myelin-associated glycoprotein (mag): Past, present and beyond. J. Neurochem. 2007, 100, 1431–1448. [Google Scholar] [CrossRef]
- Kopatz, J.; Beutner, C.; Welle, K.; Bodea, L.G.; Reinhardt, J.; Claude, J.; Linnartz-Gerlach, B.; Neumann, H. Siglec-h on activated microglia for recognition and engulfment of glioma cells. Glia 2013, 61, 1122–1133. [Google Scholar] [CrossRef]
- Blasius, A.L.; Colonna, M. Sampling and signaling in plasmacytoid dendritic cells: The potential roles of siglec-h. Trends Immunol. 2006, 27, 255–260. [Google Scholar] [CrossRef]
- Mott, R.T.; Ait-Ghezala, G.; Town, T.; Mori, T.; Vendrame, M.; Zeng, J.; Ehrhart, J.; Mullan, M.; Tan, J. Neuronal expression of cd22: Novel mechanism for inhibiting microglial proinflammatory cytokine production. Glia 2004, 46, 369–379. [Google Scholar] [CrossRef]
- Duan, S.; Paulson, J.C. Siglecs as immune cell checkpoints in disease. Annu. Rev. Immunol. 2020, 38. [Google Scholar] [CrossRef] [Green Version]
- Puente-Marin, S.; Nombela, I.; Ciordia, S.; Mena, M.C.; Chico, V.; Coll, J.; Ortega-Villaizan, M.D.M. In silico functional networks identified in fish nucleated red blood cells by means of transcriptomic and proteomic profiling. Genes (Basel) 2018, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Passantino, L.; Massaro, M.A.; Jirillo, F.; Di Modugno, D.; Ribaud, M.R.; Modugno, G.D.; Passantino, G.F.; Jirillo, E. Antigenically activated avian erythrocytes release cytokine-like factors: A conserved phylogenetic function discovered in fish. Immunopharmacol. Immunotoxicol. 2007, 29, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Morera, D.; Roher, N.; Ribas, L.; Balasch, J.C.; Donate, C.; Callol, A.; Boltana, S.; Roberts, S.; Goetz, G.; Goetz, F.W.; et al. Rna-seq reveals an integrated immune response in nucleated erythrocytes. PLoS ONE 2011, 6, e26998. [Google Scholar] [CrossRef] [Green Version]
- Workenhe, S.T.; Kibenge, M.J.; Wright, G.M.; Wadowska, D.W.; Groman, D.B.; Kibenge, F.S. Infectious salmon anaemia virus replication and induction of alpha interferon in atlantic salmon erythrocytes. Virol. J. 2008, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puente-Marin, S.; Thwaite, R.; Mercado, L.; Coll, J.; Roher, N.; Ortega-Villaizan, M.D.M. Fish red blood cells modulate immune genes in response to bacterial inclusion bodies made of tnfα and a g-vhsv fragment. Front. Immunol. 2019, 10, 1055. [Google Scholar] [CrossRef] [PubMed]
- Raja-Sabudin, R.Z.; Othman, A.; Ahmed-Mohamed, K.A.; Ithnin, A.; Alauddin, H.; Alias, H.; Abdul-Latif, Z.; Das, S.; Abdul-Wahid, F.S.; Hussin, N.H. Immature reticulocyte fraction is an early predictor of bone marrow recovery post chemotherapy in patients with acute leukemia. Saudi Med. J. 2014, 35, 346–349. [Google Scholar]
- Ney, P.A. Normal and disordered reticulocyte maturation. Curr. Opin. Hematol. 2011, 18, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.A.; Giltiay, N.V. Cd22: A regulator of innate and adaptive b cell responses and autoimmunity. Front. Immunol. 2018, 9, 2235. [Google Scholar] [CrossRef]
- Nitschke, L. The role of cd22 and other inhibitory co-receptors in b-cell activation. Curr. Opin. Immunol. 2005, 17, 290–297. [Google Scholar] [CrossRef]
- Tedder, T.F.; Poe, J.C.; Haas, K.M. Cd22: A multifunctional receptor that regulates B lymphocyte survival and signal transduction. Adv. Immunol. 2005, 88, 1–50. [Google Scholar]
- Parra, D.; Korytář, T.; Takizawa, F.; Sunyer, J.O. B cells and their role in the teleost gut. Dev. Comp. Immunol. 2016, 64, 150–166. [Google Scholar] [CrossRef] [Green Version]
- Parra, D.; Takizawa, F.; Sunyer, J.O. Evolution of b cell immunity. Annu. Rev. Anim. Biosci. 2013, 1, 65–97. [Google Scholar] [CrossRef] [Green Version]
- Wasim, L.; Buhari, F.H.M.; Yoganathan, M.; Sicard, T.; Ereño-Orbea, J.; Julien, J.-P.; Treanor, B. N-linked glycosylation regulates cd22 organization and function. Front. Immunol. 2019, 10, 699. [Google Scholar] [CrossRef] [Green Version]
- Muller, J.; Obermeier, I.; Wohner, M.; Brandl, C.; Mrotzek, S.; Angermuller, S.; Maity, P.C.; Reth, M.; Nitschke, L. Cd22 ligand-binding and signaling domains reciprocally regulate B-cell Ca2+ signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 12402–12407. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Shaper, N.L.; Itonori, S.; Heffer-Lauc, M.; Sheikh, K.A.; Schnaar, R.L. Myelin-associated glycoprotein (siglec-4) expression is progressively and selectively decreased in the brains of mice lacking complex gangliosides. Glycobiology 2004, 14, 851–857. [Google Scholar] [CrossRef]
- Cagnoni, A.J.; Pérez Sáez, J.M.; Rabinovich, G.A.; Mariño, K.V. Turning-off signaling by siglecs, selectins, and galectins: Chemical inhibition of glycan-dependent interactions in cancer. Front. Oncol. 2016, 6, 109. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bornhöfft, K.F.; Martorell Ribera, J.; Viergutz, T.; Venuto, M.T.; Gimsa, U.; Galuska, S.P.; Rebl, A. Characterization of Sialic Acid-Binding Immunoglobulin-Type Lectins in Fish Reveals Teleost-Specific Structures and Expression Patterns. Cells 2020, 9, 836. https://doi.org/10.3390/cells9040836
Bornhöfft KF, Martorell Ribera J, Viergutz T, Venuto MT, Gimsa U, Galuska SP, Rebl A. Characterization of Sialic Acid-Binding Immunoglobulin-Type Lectins in Fish Reveals Teleost-Specific Structures and Expression Patterns. Cells. 2020; 9(4):836. https://doi.org/10.3390/cells9040836
Chicago/Turabian StyleBornhöfft, Kim F., Joan Martorell Ribera, Torsten Viergutz, Marzia T. Venuto, Ulrike Gimsa, Sebastian P. Galuska, and Alexander Rebl. 2020. "Characterization of Sialic Acid-Binding Immunoglobulin-Type Lectins in Fish Reveals Teleost-Specific Structures and Expression Patterns" Cells 9, no. 4: 836. https://doi.org/10.3390/cells9040836