CRISPR/Cas9-Mediated Genome Editing Reveals Oosp Family Genes are Dispensable for Female Fertility in Mice

 , , and
, , and 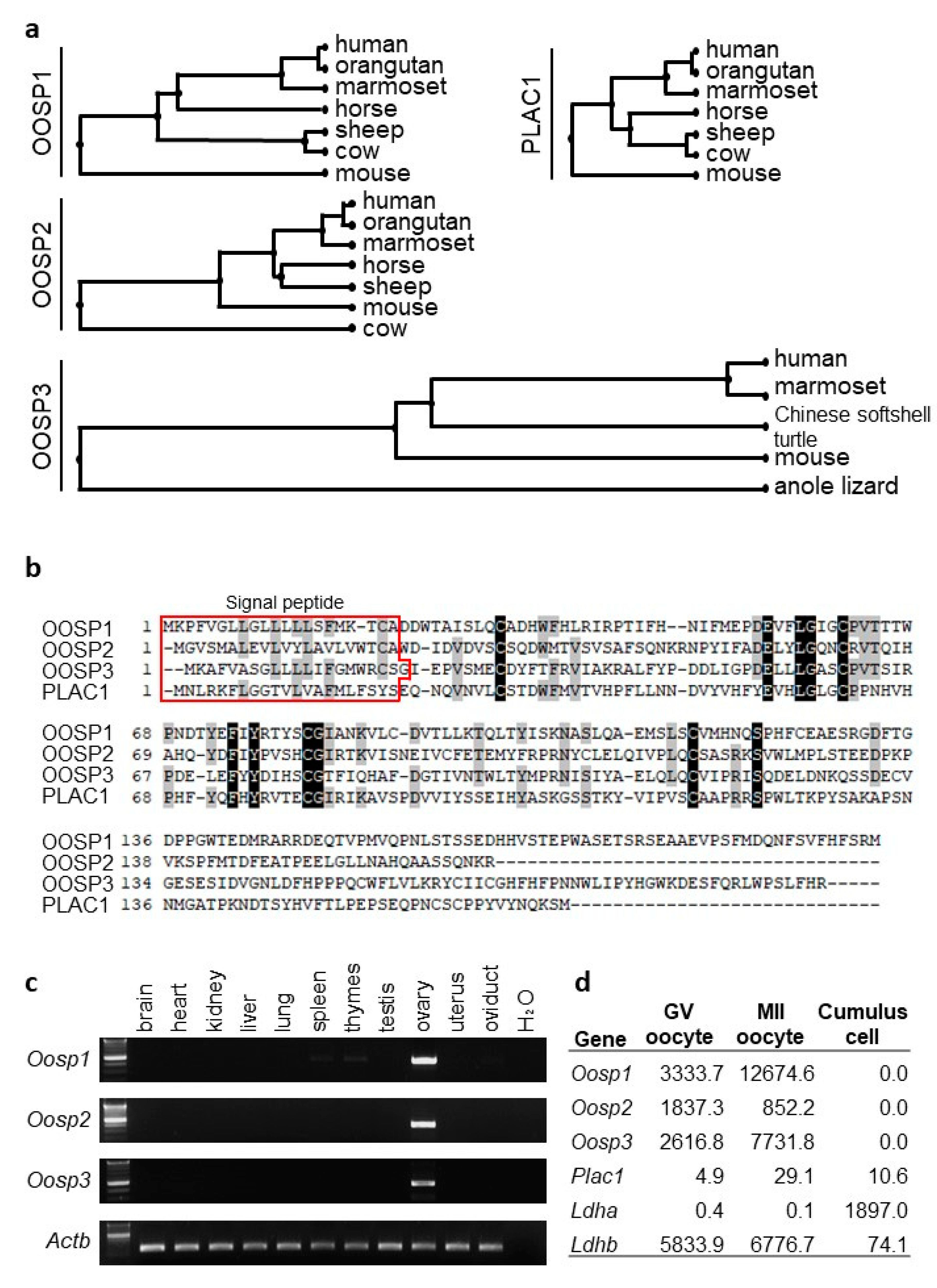{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Phylogenetic Tree and Protein Multiple Sequence Alignment
2.3. RT-PCR and RNAseq Analysis
2.4. Generation of KO Mice with Conventional Approach
2.5. Generation of KO Mice with CRISPR/Cas9 System
2.6. Fertility Test for KO Female Mice
2.7. Ovary Collection and HE Staining
2.8. Sperm-ZP Binding Assay and In Vitro Fertilization (IVF)
2.9. Statistics
3. Results
3.1. In Silico Experimental Strategies to Discover Ovary-Enriched Genes for Functional Analysis
3.2. RT-PCR and FPKM Analysis Confirms Ovary-Enriched Expression in Mice
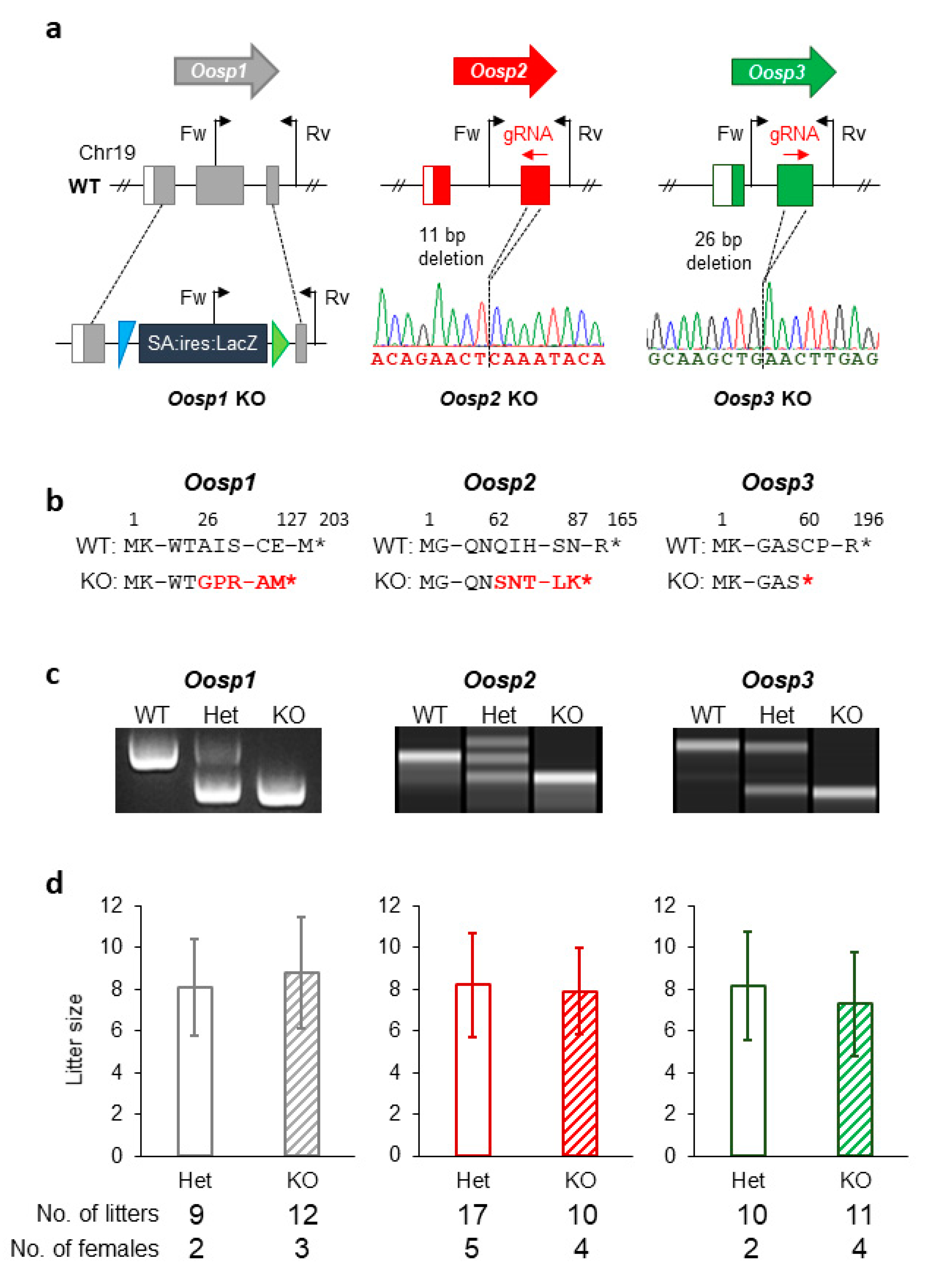
3.3. Generation and Phenotypic Analysis of Oosp1 Mutant Mice Using Traditional KO Methods
3.4. Generation and Phenotypic Analysis of Oosp2 and Oosp3 Mutant Mice Using the CRISPR/Cas9 System
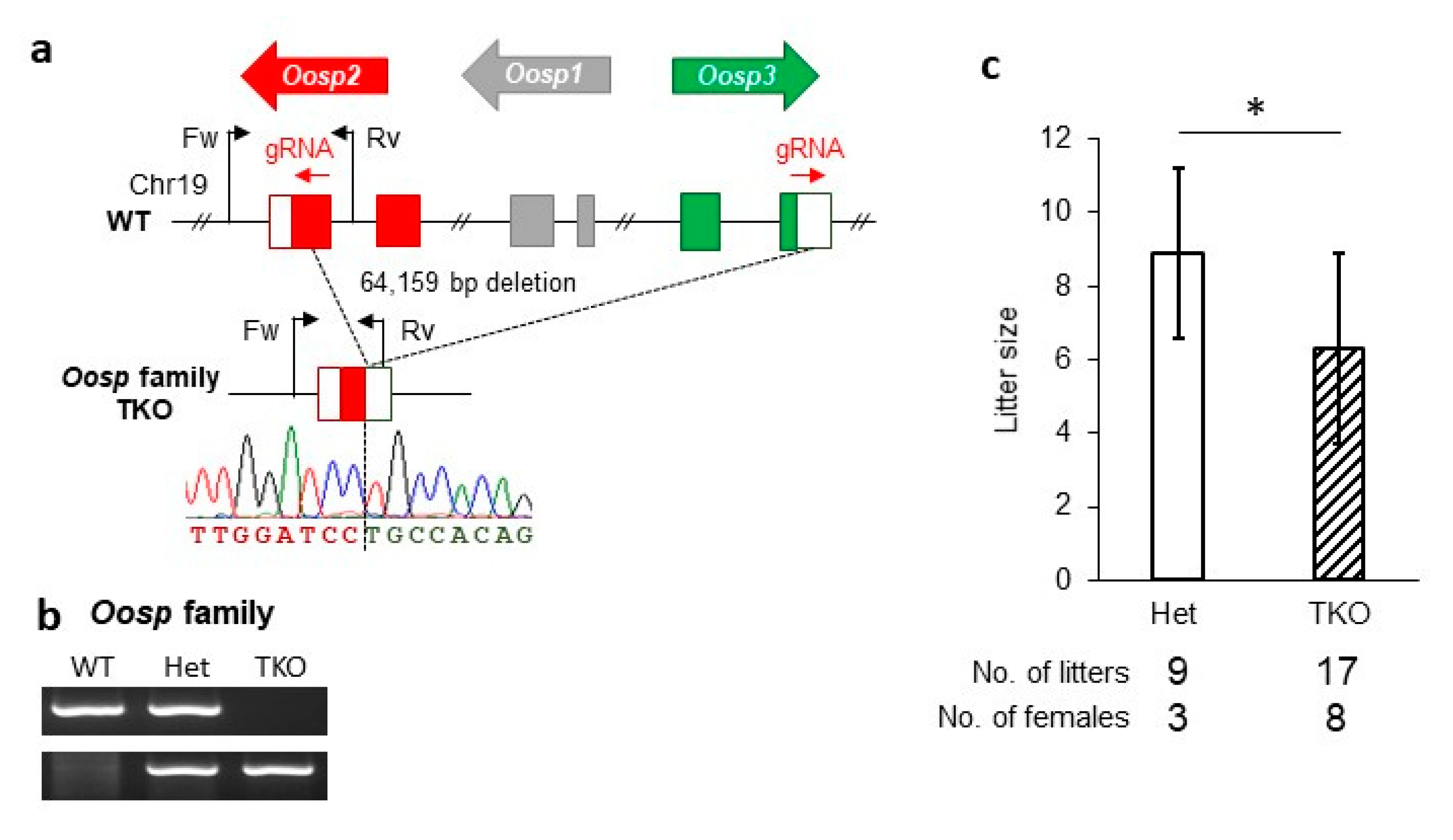
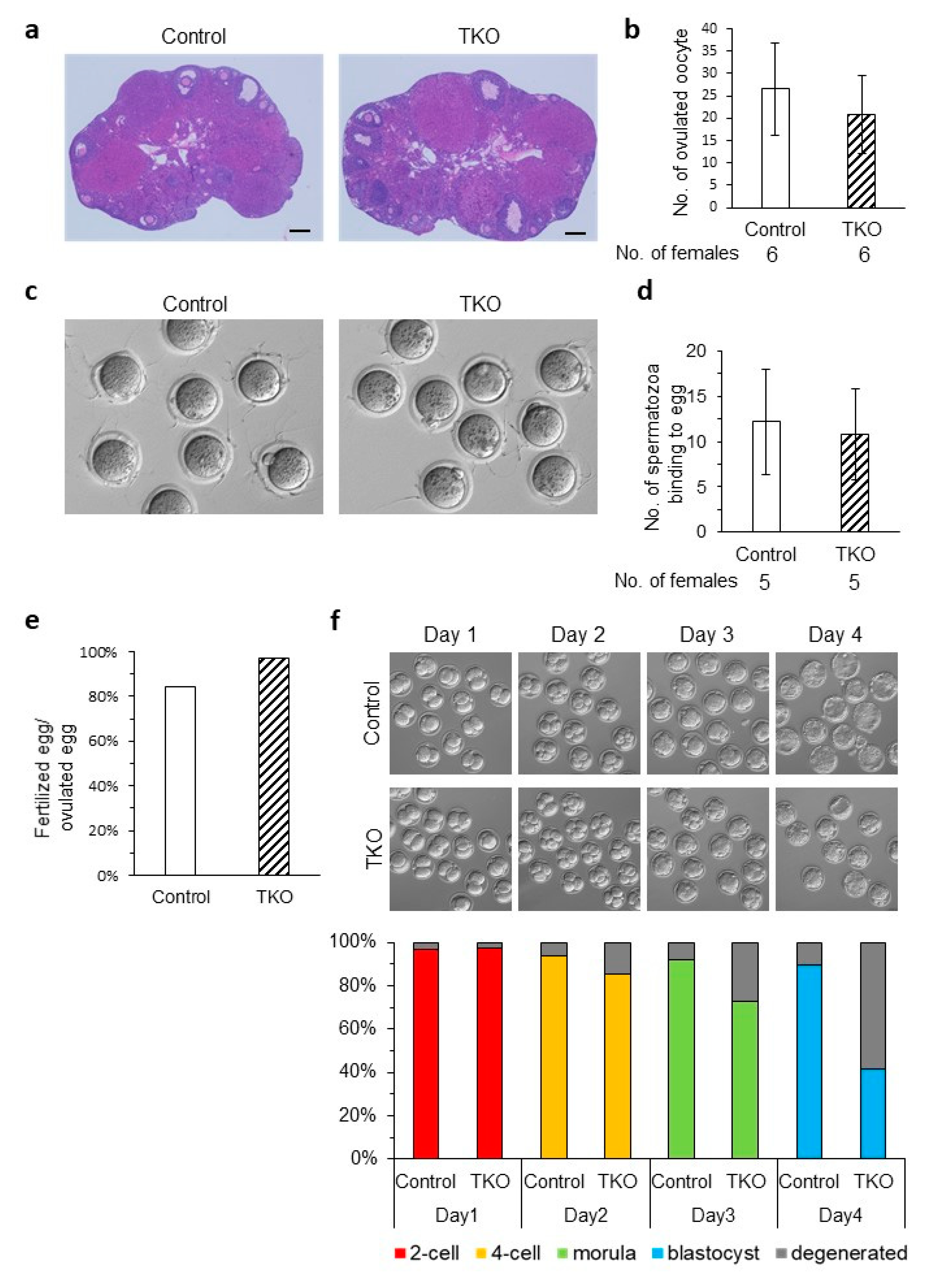
3.5. Generation and Phenotypic Analysis of the Oosp Family Deletion Mice
3.6. Generation and Phenotypic Analysis of Cd160 and Egfl6 Mutant Mice Using the CRISPR/Cas9 System
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, Regional, and Global Trends in Infertility Prevalence Since 1990: A Systematic Analysis of 277 Health Surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- McLeod, C.; Ponesse, J. Infertility and Moral Luck: The Politics of Women Blaming Themselves for Infertility. IJFAB 2008, 1, 126–144. [Google Scholar] [CrossRef]
- Lotti, F.; Maggi, M. Sexual Dysfunction and Male Infertility. Nat. Rev. Urol. 2018, 15, 287–307. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.; Liu, J.H. Primary Ovarian Insufficiency: An Update. Int. J. Women’s Health. 2014, 6, 235–243. [Google Scholar]
- Lindsay, T.J.; Vitrikas, K.R. Evaluation and treatment of infertility. Am. Fam. Physician 2015, 91, 308–314. [Google Scholar] [PubMed]
- Tomassetti, C.; D’Hooghe, T. Endometriosis and infertility: Insights into the causal link and management strategies. Best Pr. Res. Clin. Obstet. Gynaecol. 2018, 51, 25–33. [Google Scholar] [CrossRef]
- Mojarrad, M.; Hassanzadeh-Nazarabadi, M.; Tafazoli, N. Polymorphism of Genes and Implantation Failure. Int. J. Mol. Cell. Med. 2013, 2, 1–8. [Google Scholar]
- Zorrilla, M.; Yatsenko, A.N. The Genetics of Infertility: Current Status of the Field. Curr. Genet. Med. Rep. 2013, 1, 247–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, K.M.; Boulet, S.L.; Jamieson, D.J.; Kissin, D.M.; National Assisted Reproductive Technology Surveillance System (NASS) Group. Trends and Outcomes of Gestational Surrogacy in the United States. Fertil. Steril. 2016, 106, 435–442.e2. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Wang, N.; Jin, F. Long-Term Follow-up of Children Conceived through Assisted Reproductive Technology. J. Zhejiang Univ. Sci. B 2013, 14, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Gunter, C.; Dhand, R. Human Biology by Proxy. Nature 2002, 420, 509. [Google Scholar] [CrossRef]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G.J. Juno Is the Egg Izumo Receptor and Is Essential for Mammalian Fertilisation. Nature 2014, 508, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Wassarman, P.M. Zona Pellucida Glycoproteins. J. Biol. Chem. 2008, 283, 24285–24289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rankin, T.; Talbot, P.; Lee, E.; Dean, J. Abnormal Zonae Pellucidae in Mice Lacking ZP1 Result in Early Embryonic Loss. Development 1999, 126, 3847–3855. [Google Scholar]
- Abbasi, F.; Miyata, H.; Ikawa, M. Revolutionizing Male Fertility Factor Research in Mice by Using the Genome Editing Tool CRISPR/Cas9. Reprod. Med. Biol. 2017, 17, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Mashiko, D.; Fujihara, Y.; Satouh, Y.; Miyata, H.; Isotani, A.; Ikawa, M. Generation of Mutant Mice by Pronuclear Injection of Circular Plasmid Expressing Cas9 and Single Guided RNA. Sci. Rep. 2013, 3, 3355. [Google Scholar] [CrossRef]
- Abbasi, F.; Miyata, H.; Shimada, K.; Morohoshi, A.; Nozawa, K.; Matsumura, T.; Xu, Z.; Pratiwi, P.; Ikawa, M. RSPH6A Is Required for Sperm Flagellum Formation and Male Fertility in Mice. J. Cell. Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Pendola, F.L.; Jacob, R.; Lau, A.L.; Eppig, J.J.; Matzuk, M.M. Oosp1 Encodes a Novel Mouse Oocyte-Secreted Protein. Genesis 2001, 31, 105–110. [Google Scholar] [CrossRef]
- Paillisson, A.; Dadé, S.; Callebaut, I.; Bontoux, M.; Dalbiès-Tran, R.; Vaiman, D.; Monget, P. Identification, Characterization and Metagenome Analysis of Oocyte-Specific Genes Organized in Clusters in the Mouse Genome. BMC Genomics 2005, 6, 76. [Google Scholar] [CrossRef]
- Cunningham, F.; Achuthan, P.; Akanni, W.; Allen, J.; Amode, M.R.; Armean, I.M.; Bennett, R.; Bhai, J.; Billis, K.; Boddu, S.; et al. Ensembl 2019. Nucleic Acids Res 2019, 47, D745–D751. [Google Scholar] [CrossRef] [Green Version]
- Skarnes, W.C.; Rosen, B.; West, A.P.; Koutsourakis, M.; Bushell, W.; Iyer, V.; Mujica, A.O.; Thomas, M.; Harrow, J.; Cox, T.; et al. A Conditional Knockout Resource for the Genome-Wide Study of Mouse Gene Function. Nature 2011, 474, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujihara, Y.; Kaseda, K.; Inoue, N.; Ikawa, M.; Okabe, M. Production of Mouse Pups from Germline Transmission-Failed Knockout Chimeras. Transgenic Res. 2013, 22, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for Designing CRISPR/Cas Guide RNA with Reduced off-Target Sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Mashimo, T. Simple Genome Editing of Rodent Intact Embryos by Electroporation. PLoS ONE 2015, 10, e0142755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muro, Y.; Hasuwa, H.; Isotani, A.; Miyata, H.; Yamagata, K.; Ikawa, M.; Yanagimachi, R.; Okabe, M. Behavior of Mouse Spermatozoa in the Female Reproductive Tract from Soon after Mating to the Beginning of Fertilization. Biol. Reprod. 2016, 94, 80. [Google Scholar] [CrossRef] [PubMed]
- Yates, B.; Braschi, B.; Gray, K.; Seal, R.L.; Tweedie, S.; Bruford, E. Genenames.org: The HGNC and VGNC resources in 2017. Nucleic Acids Res. 2016, 45, D619–D625. [Google Scholar] [CrossRef]
- Muto, M.; Fujihara, Y.; Tobita, T.; Kiyozumi, D.; Ikawa, M. Lentiviral Vector-Mediated Complementation Restored Fetal Viability but Not Placental Hyperplasia in Plac1-Deficient Mice. Biol. Reprod. 2016, 94, 6. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Miyata, H.; Castaneda, J.M.; Fujihara, Y.; Yu, Z.; Archambeault, D.R.; Isotani, A.; Kiyozumi, D.; Kriseman, M.L.; Mashiko, D.; Matsumura, T.; et al. Genome Engineering Uncovers 54 Evolutionarily Conserved and Testis-Enriched Genes That Are Not Required for Male Fertility in Mice. Proc. Natl. Acad. Sci. USA 2016, 113, 7704–7710. [Google Scholar] [CrossRef] [Green Version]
- Noda, T.; Sakurai, N.; Nozawa, K.; Kobayashi, S.; Devlin, D.J.; Matzuk, M.M.; Ikawa, M. Nine Genes Abundantly Expressed in the Epididymis Are Not Essential for Male Fecundity in Mice. Andrology 2019, 7, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Oura, S.; Matsumura, T.; Oji, A.; Sakurai, N.; Fujihara, Y.; Shimada, K.; Miyata, H.; Tobita, T.; Noda, T.; et al. CRISPR/Cas9-Mediated Genome Editing Reveals 30 Testis-Enriched Genes Dispensable for Male Fertility in Mice†. Biol. Reprod. 2019, 101, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, K.; Satouh, Y.; Fujimoto, T.; Oji, A.; Ikawa, M. Sperm-Borne Phospholipase C Zeta-1 Ensures Monospermic Fertilization in Mice. Sci. Rep. 2018, 8, 1315. [Google Scholar] [CrossRef]
- Giustiniani, J.; Marie-Cardine, A.; Bensussan, A. A Soluble Form of the MHC Class I-Specific CD160 Receptor Is Released from Human Activated NK Lymphocytes and Inhibits Cell-Mediated Cytotoxicity. J. Immunol. 2007, 178, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Meggyes, M.; Szereday, L.; Jakso, P.; Bogar, B.; Bogdan, A.; Nörenberg, J.; Miko, E.; Barakonyi, A. Expansion of CD4 Phenotype among CD160 Receptor-Expressing Lymphocytes in Murine Pregnancy. Am. J. Reprod. Immunol. 2017, 78, e12745. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Ingram, P.; Chen, Y.-C.; Deng, N.; Pearson, A.; Niknafs, Y.; O’Hayer, P.; Wang, Y.; Zhang, Z.-Y.; Boscolo, E.; et al. EGFL6 Regulates the Asymmetric Division, Maintenance and Metastasis of ALDH+ Ovarian Cancer Cells. Cancer Res. 2016, 76, 6396–6409. [Google Scholar] [CrossRef] [Green Version]
- Amoushahi, M.; Sunde, L.; Lykke-Hartmann, K. The Pivotal Roles of the NOD-like Receptors with a PYD Domain, NLRPs, in Oocytes and Early Embryo Development. Biol. Reprod. 2019, 101, 284–296. [Google Scholar] [CrossRef]
- Kuchmiy, A.A.; D’Hont, J.; Hochepied, T.; Lamkanfi, M. NLRP2 Controls Age-Associated Maternal Fertility. J. Exp. Med. 2016, 213, 2851–2860. [Google Scholar] [CrossRef] [Green Version]
- Qin, D.; Gao, Z.; Xiao, Y.; Zhang, X.; Ma, H.; Yu, X.; Nie, X.; Fan, N.; Wang, X.; Ouyang, Y.; et al. The Subcortical Maternal Complex Protein Nlrp4f Is Involved in Cytoplasmic Lattice Formation and Organelle Distribution. Development 2019, 146. [Google Scholar] [CrossRef] [Green Version]
- Tong, Z.-B.; Gold, L.; Pfeifer, K.E.; Dorward, H.; Lee, E.; Bondy, C.A.; Dean, J.; Nelson, L.M. Mater, a Maternal Effect Gene Required for Early Embryonic Development in Mice. Nat. Genet. 2000, 26, 267–268. [Google Scholar] [CrossRef]
- Fujihara, Y.; Noda, T.; Kobayashi, K.; Oji, A.; Kobayashi, S.; Matsumura, T.; Larasati, T.; Oura, S.; Kojima-Kita, K.; Yu, Z.; et al. Identification of Multiple Male Reproductive Tract-Specific Proteins That Regulate Sperm Migration through the Oviduct in Mice. Proc. Natl. Acad. Sci. USA 2019, 116, 18498–18506. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbasi, F.; Kodani, M.; Emori, C.; Kiyozumi, D.; Mori, M.; Fujihara, Y.; Ikawa, M. CRISPR/Cas9-Mediated Genome Editing Reveals Oosp Family Genes are Dispensable for Female Fertility in Mice. Cells 2020, 9, 821. https://doi.org/10.3390/cells9040821
Abbasi F, Kodani M, Emori C, Kiyozumi D, Mori M, Fujihara Y, Ikawa M. CRISPR/Cas9-Mediated Genome Editing Reveals Oosp Family Genes are Dispensable for Female Fertility in Mice. Cells. 2020; 9(4):821. https://doi.org/10.3390/cells9040821
Chicago/Turabian StyleAbbasi, Ferheen, Mayo Kodani, Chihiro Emori, Daiji Kiyozumi, Masashi Mori, Yoshitaka Fujihara, and Masahito Ikawa. 2020. "CRISPR/Cas9-Mediated Genome Editing Reveals Oosp Family Genes are Dispensable for Female Fertility in Mice" Cells 9, no. 4: 821. https://doi.org/10.3390/cells9040821