Cholest-4,6-Dien-3-One Promote Epithelial-To-Mesenchymal Transition (EMT) in Biliary Tree Stem/Progenitor Cell Cultures In Vitro

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Tissue Sourcing
2.2. Cells Isolation
2.3. Kubota’s Medium
2.4. Cell Cultures
- Hedgehog pathway [25]: Purmorphamine (1.5 µM, Sigma, St. Louis, Missouri, USA #SML0868), DAPT (1.0 µM, Sigma, St. Louis, Missouri, USA #D5942) and dorsomorphin (2.0 µM, Sigma, St. Louis, Missouri, USA #P5499).
2.5. Proliferation and Viability Analysis
2.6. Cell Senescence
2.7. RNA Extraction and Quantitative Reverse-Transcription Polymerase Chain Reaction (RT-qPCR) Analysis
2.8. Protein Extraction and Western Blot
2.9. ELISA Assay
2.10. Telomerase Activity Quantification
2.11. Protein Pathway Analysis
2.12. Statistical Analysis
3. Results
3.1. Viability, Proliferation and Senescence after Chronic Cholest-4,6-Dien-3-One Exposure in hBTSC Cultures
3.1.1. Cell Number in hBTSC Cultures
3.1.2. Cell Viability
3.1.3. Cell Proliferation
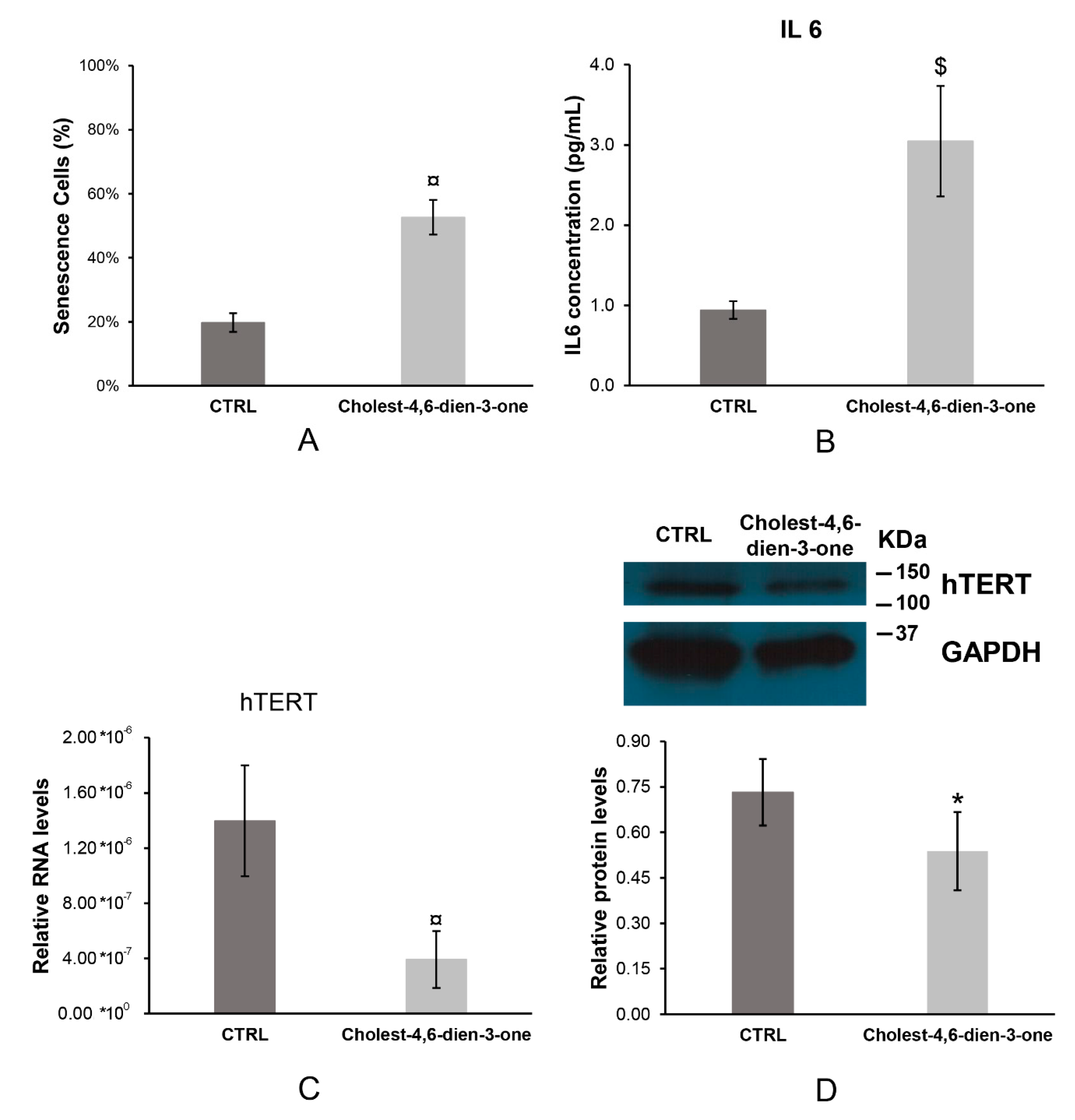
3.1.4. Cell Senescence
3.1.5. Interleukin 6
3.1.6. Human Telomerase (hTERT)
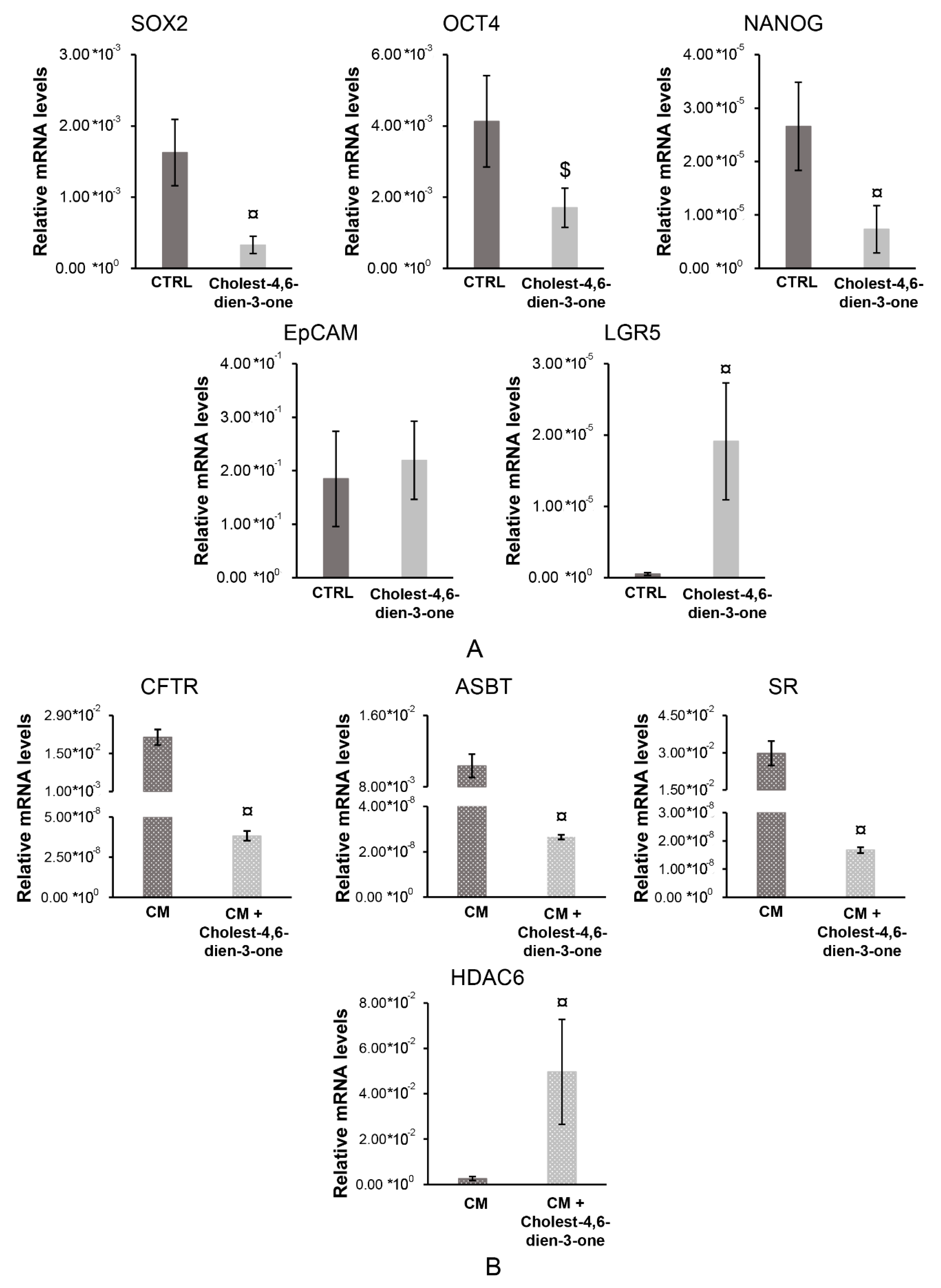
3.2. Expression of Stem Cell, Pluripotent Cell and Epithelial-To-Mesenchymal Transition Markers in hBTSCs Chronically Exposed to Cholest-4,6-Dien-3-One
3.2.1. Stem, Pluripotent and Mature cell Markers
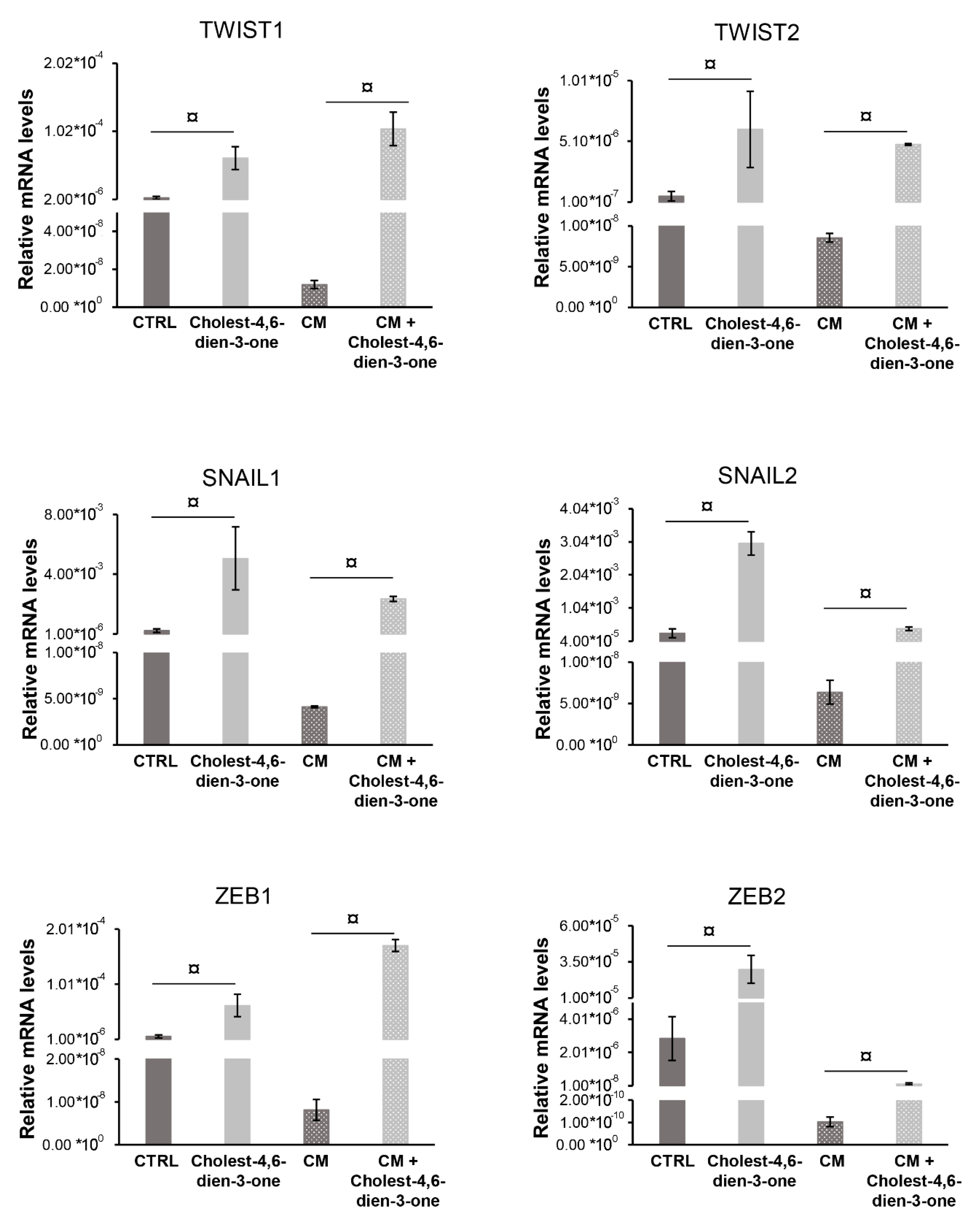
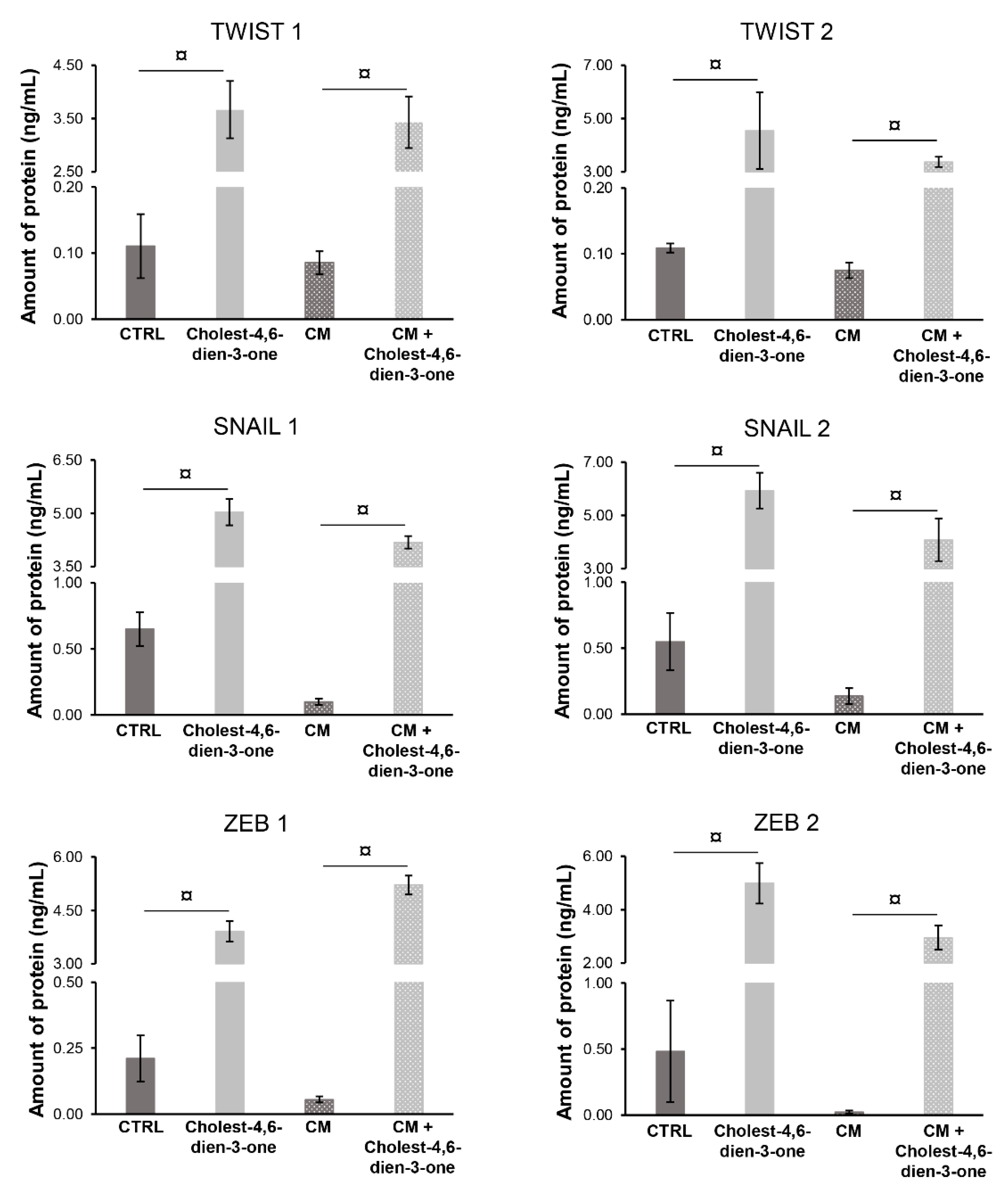
3.2.2. Markers of Epithelial-Mesenchymal-Transition
3.3. Cholest-4,6-Dien-3-One Enhanced the Bmp and Shh Pathway Expression in hBTSCs Treated for 10 Days
3.3.1. Cell Pathways Induction to Cholest-4,6-Dien-3-One
3.3.2. Bmp-4 and Shh Pathways Stimulated in hBTSCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carpino, G.; Cardinale, V.; Folseraas, T.; Overi, D.; Grzyb, K.; Costantini, D.; Berloco, P.B.; Di Matteo, S.; Karlsen, T.H.; Alvaro, D.; et al. Neoplastic Transformation of the Peribiliary Stem Cell Niche in Cholangiocarcinoma Arisen in Primary Sclerosing Cholangitis. Hepatology 2019, 69, 622–638. [Google Scholar] [CrossRef]
- Lazaridis, K.N.; LaRusso, N.F. Primary Sclerosing Cholangitis. N. Engl. J. Med. 2016, 375, 1161–1170. [Google Scholar] [CrossRef]
- Eaton, J.E.; Talwalkar, J.A.; Lazaridis, K.N.; Gores, G.J.; Lindor, K.D. Pathogenesis of primary sclerosing cholangitis and advances in diagnosis and management. Gastroenterology 2013, 145, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Goode, E.C.; Rushbrook, S.M. A review of the medical treatment of primary sclerosing cholangitis in the 21st century. Ther. Adv. Chronic Dis. 2016, 7, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.A.; Chapman, R.W. Review article: The management of primary sclerosing cholangitis. Aliment. Pharmacol. Ther. 1997, 11, 33–43. [Google Scholar] [CrossRef]
- Kummen, M.; Holm, K.; Anmarkrud, J.A.; Nygard, S.; Vesterhus, M.; Hoivik, M.L.; Troseid, M.; Marschall, H.U.; Schrumpf, E.; Moum, B.; et al. The gut microbial profile in patients with primary sclerosing cholangitis is distinct from patients with ulcerative colitis without biliary disease and healthy controls. Gut 2017, 66, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.C.; LaRusso, N.F.; Gores, G.J.; Lazaridis, K.N. Epigenetics in the Primary Biliary Cholangitis and Primary Sclerosing Cholangitis. Semin. Liver Dis. 2017, 37, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Rupp, C.; Bode, K.A.; Leopold, Y.; Sauer, P.; Gotthardt, D.N. Pathological features of primary sclerosing cholangitis identified by bile proteomic analysis. Biochim. Biophys. Acta (BBA) - Mol. Basis Dis. 2018, 1864, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.W.; Choi, H.S.; Lee, S.P.; Kuver, R. Oxysterols from human bile induce apoptosis of canine gallbladder epithelial cells in monolayer culture. Am. J. Physiol. Liver Physiol. 2004, 287, G1247–G1256. [Google Scholar] [CrossRef]
- Kendall, C.W.; Koo, M.; Sokoloff, E.; Rao, A.V. Effect of dietary oxidized cholesterol on azoxymethane-induced colonic preneoplasia in mice. Cancer Lett. 1992, 66, 241–248. [Google Scholar] [CrossRef]
- Moog, C.; Luu, B.; Altmeyer, A.; Bischoff, P. Studies on the immunosuppressive properties of 7,25 dihydroxycholesterol--II. Effects on early steps of T-cell activation. Int. J. Immunopharmacol. 1989, 11, 559–565. [Google Scholar] [CrossRef]
- Zhang, H.; Leung, P.S.C.; Gershwin, M.E.; Ma, X. How the biliary tree maintains immune tolerance? Biochim. Biophys Acta. Mol. Basis Dis. 2018, 1864, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Janowski, B.A.; Grogan, M.J.; Jones, S.A.; Wisely, G.B.; Kliewer, S.A.; Corey, E.J.; Mangelsdorf, D.J. Structural requirements of ligands for the oxysterol liver X receptors LXRalpha and LXRbeta. Proc. Natl. Acad. Sci. 1999, 96, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Haigh, W.G.; Lee, S.P. Identification of oxysterols in human bile and pigment gallstones. Gastroenterology 2001, 121, 118–123. [Google Scholar] [CrossRef]
- Staley, C.; Weingarden, A.R.; Khoruts, A.; Sadowsky, M.J. Interaction of gut microbiota with bile acid metabolism and its influence on disease states. Appl. Microbiol. Biotechnol. 2017, 101, 47–64. [Google Scholar] [CrossRef]
- De Assuncao, T.M.; Jalan-Sakrikar, N.; Huebert, R.C. Regenerative Medicine and the Biliary Tree. Semin. Liver Dis. 2017, 37, 17–27. [Google Scholar] [CrossRef]
- Lu, J.; Zhou, Y.; Hu, T.; Zhang, H.; Shen, M.; Cheng, P.; Dai, W.; Wang, F.; Chen, K.; Zhang, Y.; et al. Notch Signaling Coordinates Progenitor Cell-Mediated Biliary Regeneration Following Partial Hepatectomy. Sci. Rep. 2016, 6, 22754. [Google Scholar] [CrossRef]
- Carpino, G.; Nevi, L.; Overi, D.; Cardinale, V.; Lu, W.Y.; Di Matteo, S.; Safarikia, S.; Berloco, P.B.; Venere, R.; Onori, P.; et al. Peribiliary gland niche participates in biliary tree regeneration in mouse and in human primary sclerosing cholangitis. Hepatology 2019. [Google Scholar] [CrossRef]
- Nevi, L.; Cardinale, V.; Carpino, G.; Costantini, D.; Di Matteo, S.; Cantafora, A.; Melandro, F.; Brunelli, R.; Bastianelli, C.; Aliberti, C.; et al. Cryopreservation protocol for human biliary tree stem/progenitors, hepatic and pancreatic precursors. Sci. Rep. 2017, 7, 6080. [Google Scholar] [CrossRef]
- Nevi, L.; Carpino, G.; Costantini, D.; Cardinale, V.; Riccioni, O.; Di Matteo, S.; Melandro, F.; Berloco, P.B.; Reid, L.; Gaudio, E.; et al. Hyaluronan coating improves liver engraftment of transplanted human biliary tree stem/progenitor cells. Stem Cell Res. Ther. 2017, 8, 68. [Google Scholar] [CrossRef]
- Costantini, D.; Overi, D.; Casadei, L.; Cardinale, V.; Nevi, L.; Carpino, G.; Di Matteo, S.; Safarikia, S.; Valerio, M.; Melandro, F.; et al. Simulated microgravity promotes the formation of tridimensional cultures and stimulates pluripotency and a glycolytic metabolism in human hepatic and biliary tree stem/progenitor cells. Sci. Rep. 2019, 9, 5559. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, V.; Puca, R.; Carpino, G.; Scafetta, G.; Renzi, A.; De Canio, M.; Sicilia, F.; Nevi, L.; Casa, D.; Panetta, R.; et al. Adult Human Biliary Tree Stem Cells Differentiate to beta-Pancreatic Islet Cells by Treatment with a Recombinant Human Pdx1 Peptide. PLoS ONE 2015, 10, e0134677. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Reid, L.M. Clonogenic hepatoblasts, common precursors for hepatocytic and biliary lineages, are lacking classical major histocompatibility complex class I antigen. Proc. Natl. Acad. Sci. USA 2000, 97, 12132–12137. [Google Scholar] [CrossRef]
- Wang, Y.; Lanzoni, G.; Carpino, G.; Cui, C.B.; Dominguez-Bendala, J.; Wauthier, E.; Cardinale, V.; Oikawa, T.; Pileggi, A.; Gerber, D.; et al. Biliary tree stem cells, precursors to pancreatic committed progenitors: Evidence for possible life-long pancreatic organogenesis. Stem Cells 2013, 31, 1966–1979. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Z.B.; Gao, H.; Zhang, D.; Wang, W.L. Down-regulation of SHH/BMP4 signalling in human anorectal malformations. J. Int. Med Res. 2009, 37, 1842–1850. [Google Scholar] [CrossRef]
- Clemons, N.J.; Wang, D.H.; Croagh, D.; Tikoo, A.; Fennell, C.M.; Murone, C.; Scott, A.M.; Watkins, D.N.; Phillips, W.A. Sox9 drives columnar differentiation of esophageal squamous epithelium: A possible role in the pathogenesis of Barrett’s esophagus. Am. J. Physiol. Liver Physiol. 2012, 303, G1335–G1346. [Google Scholar] [CrossRef]
- Ribaudo, R.; Gilman, M.; Kingston, R.E.; Choczynski, P.; Sacchi, N. Preparation of RNA from tissues and cells. Curr. Protoc. Immunol. 2001, 3, 10–11. [Google Scholar] [CrossRef]
- Ortiz-Montero, P.; Londono-Vallejo, A.; Vernot, J.P. Senescence-associated IL-6 and IL-8 cytokines induce a self- and cross-reinforced senescence/inflammatory milieu strengthening tumorigenic capabilities in the MCF-7 breast cancer cell line. Cell Commun. Signal. 2017, 15, 17. [Google Scholar] [CrossRef]
- Salama, R.; Sadaie, M.; Hoare, M.; Narita, M. Cellular senescence and its effector programs. Genes Dev. 2014, 28, 99–114. [Google Scholar] [CrossRef]
- Sapochnik, M.; Fuertes, M.; Arzt, E. Programmed cell senescence: Role of IL-6 in the pituitary. J. Mol. Endocrinol. 2017, 58, R241–R253. [Google Scholar] [CrossRef]
- Carpino, G.; Cardinale, V.; Renzi, A.; Hov, J.R.; Berloco, P.B.; Rossi, M.; Karlsen, T.H.; Alvaro, D.; Gaudio, E. Activation of biliary tree stem cells within peribiliary glands in primary sclerosing cholangitis. J. Hepatol. 2015, 63, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Tabibian, J.H.; O’Hara, S.P.; Splinter, P.L.; Trussoni, C.E.; LaRusso, N.F. Cholangiocyte senescence by way of N-ras activation is a characteristic of primary sclerosing cholangitis. Hepatology 2014, 59, 2263–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuver, R. Mechanisms of oxysterol-induced disease: Insights from the biliary system. Clin. Lipidol. 2012, 7, 537–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradilone, S.A.; Radtke, B.N.; Bogert, P.S.; Huang, B.Q.; Gajdos, G.B.; LaRusso, N.F. HDAC6 inhibition restores ciliary expression and decreases tumor growth. Cancer Res. 2013, 73, 2259–2270. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, V.; Wang, Y.; Carpino, G.; Alvaro, D.; Reid, L.; Gaudio, E. Multipotent stem cells in the biliary tree. Ital. J. Anat. Embryol. 2010, 115, 85–90. [Google Scholar] [PubMed]
- Carpino, G.; Cardinale, V.; Onori, P.; Franchitto, A.; Berloco, P.B.; Rossi, M.; Wang, Y.; Semeraro, R.; Anceschi, M.; Brunelli, R.; et al. Biliary tree stem/progenitor cells in glands of extrahepatic and intraheptic bile ducts: An anatomical in situ study yielding evidence of maturational lineages. J. Anat. 2012, 220, 186–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpino, G.; Cardinale, V.; Gentile, R.; Onori, P.; Semeraro, R.; Franchitto, A.; Wang, Y.; Bosco, D.; Iossa, A.; Napoletano, C.; et al. Evidence for multipotent endodermal stem/progenitor cell populations in human gallbladder. J. Hepatol. 2014, 60, 1194–1202. [Google Scholar] [CrossRef]
- Yanes, O.; Clark, J.; Wong, D.M.; Patti, G.J.; Sanchez-Ruiz, A.; Benton, H.P.; Trauger, S.A.; Desponts, C.; Ding, S.; Siuzdak, G. Metabolic oxidation regulates embryonic stem cell differentiation. Nat. Chem. Biol. 2010, 6, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Bragazzi, M.C.; Ridola, L.; Safarikia, S.; Matteo, S.D.; Costantini, D.; Nevi, L.; Cardinale, V. New insights into cholangiocarcinoma: Multiple stems and related cell lineages of origin. Ann. Gastroenterol. 2018, 31, 42–55. [Google Scholar] [CrossRef]
- Eleazar, J.A.; Memeo, L.; Jhang, J.S.; Mansukhani, M.M.; Chin, S.; Park, S.M.; Lefkowitch, J.H.; Bhagat, G. Progenitor cell expansion: An important source of hepatocyte regeneration in chronic hepatitis. J. Hepatol. 2004, 41, 983–991. [Google Scholar] [CrossRef]
- Roskams, T.; Yang, S.Q.; Koteish, A.; Durnez, A.; DeVos, R.; Huang, X.; Achten, R.; Verslype, C.; Diehl, A.M. Oxidative stress and oval cell accumulation in mice and humans with alcoholic and nonalcoholic fatty liver disease. Am. J. Pathol. 2003, 163, 1301–1311. [Google Scholar] [CrossRef] [Green Version]
- Roskams, T. Progenitor cell involvement in cirrhotic human liver diseases: From controversy to consensus. J. Hepatol. 2003, 39, 431–434. [Google Scholar] [CrossRef] [Green Version]
- Paradis, V.; Youssef, N.; Dargere, D.; Ba, N.; Bonvoust, F.; Deschatrette, J.; Bedossa, P. Replicative senescence in normal liver, chronic hepatitis C, and hepatocellular carcinomas. Hum. Pathol. 2001, 32, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Crary, G.S.; Albrecht, J.H. Expression of cyclin-dependent kinase inhibitor p21 in human liver. Hepatology 1998, 28, 738–743. [Google Scholar] [CrossRef]
- Marshall, A.; Rushbrook, S.; Davies, S.E.; Morris, L.S.; Scott, I.S.; Vowler, S.L.; Coleman, N.; Alexander, G. Relation between hepatocyte G1 arrest, impaired hepatic regeneration, and fibrosis in chronic hepatitis C virus infection. Gastroenterology 2005, 128, 33–42. [Google Scholar] [CrossRef]
- Wiemann, S.U.; Satyanarayana, A.; Tsahuridu, M.; Tillmann, H.L.; Zender, L.; Klempnauer, J.; Flemming, P.; Franco, S.; Blasco, M.A.; Manns, M.P.; et al. Hepatocyte telomere shortening and senescence are general markers of human liver cirrhosis. FASEB J. 2002, 16, 935–942. [Google Scholar] [CrossRef]
- Falkowski, O.; An, H.J.; Ianus, I.A.; Chiriboga, L.; Yee, H.; West, A.B.; Theise, N.D. Regeneration of hepatocyte ‘buds’ in cirrhosis from intrabiliary stem cells. J. Hepatol. 2003, 39, 357–364. [Google Scholar] [CrossRef]
- Rudolph, K.L.; Chang, S.; Millard, M.; Schreiber-Agus, N.; DePinho, R.A. Inhibition of experimental liver cirrhosis in mice by telomerase gene delivery. Science 2000, 287, 1253–1258. [Google Scholar] [CrossRef]
- Shawi, M.; Autexier, C. Telomerase, senescence and ageing. Mech. Ageing Dev. 2008, 129, 3–10. [Google Scholar] [CrossRef]
- Schosserer, M.; Grillari, J.; Breitenbach, M. The Dual Role of Cellular Senescence in Developing Tumors and Their Response to Cancer Therapy. Front. Oncol. 2017, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Zeng, S.; Shen, W.H.; Liu, L. Senescence and Cancer. Cancer Transl. Med. 2018, 4, 70–74. [Google Scholar] [CrossRef]
- Kong, F.; Zheng, C.; Xu, D. Telomerase as a "stemness" enzyme. Sci. China Life Sci. 2014, 57, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Mace, T.A.; Shakya, R.; Pitarresi, J.R.; Swanson, B.; McQuinn, C.W.; Loftus, S.; Nordquist, E.; Cruz-Monserrate, Z.; Yu, L.; Young, G.; et al. IL-6 and PD-L1 antibody blockade combination therapy reduces tumour progression in murine models of pancreatic cancer. Gut 2018, 67, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Carnevale, G.; Carpino, G.; Cardinale, V.; Pisciotta, A.; Riccio, M.; Bertoni, L.; Gibellini, L.; De Biasi, S.; Nevi, L.; Costantini, D.; et al. Activation of Fas/FasL pathway and the role of c-FLIP in primary culture of human cholangiocarcinoma cells. Sci. Rep. 2017, 7, 14419. [Google Scholar] [CrossRef]
- Riccio, M.; Carnevale, G.; Cardinale, V.; Gibellini, L.; De Biasi, S.; Pisciotta, A.; Carpino, G.; Gentile, R.; Berloco, P.B.; Brunelli, R.; et al. The Fas/Fas ligand apoptosis pathway underlies immunomodulatory properties of human biliary tree stem/progenitor cells. J. Hepatol. 2014, 61, 1097–1105. [Google Scholar] [CrossRef]
- Gupta, S.; Takebe, N.; Lorusso, P. Targeting the Hedgehog pathway in cancer. Ther. Adv. Med Oncol. 2010, 2, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Martinez, V.G.; Rubio, C.; Martinez-Fernandez, M.; Segovia, C.; Lopez-Calderon, F.; Garin, M.I.; Teijeira, A.; Munera-Maravilla, E.; Varas, A.; Sacedon, R.; et al. BMP4 Induces M2 Macrophage Polarization and Favors Tumor Progression in Bladder Cancer. Clin. Cancer Res. 2017, 23, 7388–7399. [Google Scholar] [CrossRef]
- Sims-Mourtada, J.; Opdenaker, L.M.; Davis, J.; Arnold, K.M.; Flynn, D. Taxane-induced hedgehog signaling is linked to expansion of breast cancer stem-like populations after chemotherapy. Mol. Carcinog. 2015, 54, 1480–1493. [Google Scholar] [CrossRef]
- Grzelak, C.A.; Sigglekow, N.D.; Tirnitz-Parker, J.E.; Hamson, E.J.; Warren, A.; Maneck, B.; Chen, J.; Patkunanathan, B.; Boland, J.; Cheng, R.; et al. Widespread GLI expression but limited canonical hedgehog signaling restricted to the ductular reaction in human chronic liver disease. PLoS ONE 2017, 12, e0171480. [Google Scholar] [CrossRef]
- Dhanyamraju, P.K.; Holz, P.S.; Finkernagel, F.; Fendrich, V.; Lauth, M. Histone deacetylase 6 represents a novel drug target in the oncogenic Hedgehog signaling pathway. Mol. Cancer Ther. 2015, 14, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Peng, Y.; Li, H. The Injury-Related Activation of Hedgehog Signaling Pathway Modulates the Repair-Associated Inflammation in Liver Fibrosis. Front. Immunol. 2017, 8, 1450. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Yu, J.; Park, A.; Dubon, M.J.; Do, J.; Kim, Y.; Nam, D.; Noh, J.; Park, K.S. BMP-4 enhances epithelial mesenchymal transition and cancer stem cell properties of breast cancer cells via Notch signaling. Sci. Rep. 2019, 9, 11724. [Google Scholar] [CrossRef] [Green Version]
- Listopad, J.J.; Kammertoens, T.; Anders, K.; Silkenstedt, B.; Willimsky, G.; Schmidt, K.; Kuehl, A.A.; Loddenkemper, C.; Blankenstein, T. Fas expression by tumor stroma is required for cancer eradication. Proc. Natl. Acad. Sci. USA 2013, 110, 2276–2281. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, L.S.; Eliza, M.; Niven-Fairchild, T.; Naftolin, F.; Mor, G. The Fas/Fas-ligand system: A mechanism for immune evasion in human breast carcinomas. Breast Cancer Res. Treat. 1999, 54, 245–253. [Google Scholar] [CrossRef]
- Walker, P.R.; Saas, P.; Dietrich, P.Y. Role of Fas ligand (CD95L) in immune escape: The tumor cell strikes back. J. Immunol. 1997, 158, 4521–4524. [Google Scholar]
- Maecker, H.L.; Yun, Z.; Maecker, H.T.; Giaccia, A.J. Epigenetic changes in tumor Fas levels determine immune escape and response to therapy. Cancer Cell 2002, 2, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Hahne, M.; Rimoldi, D.; Schroter, M.; Romero, P.; Schreier, M.; French, L.E.; Schneider, P.; Bornand, T.; Fontana, A.; Lienard, D.; et al. Melanoma cell expression of Fas(Apo-1/CD95) ligand: Implications for tumor immune escape. Science 1996, 274, 1363–1366. [Google Scholar] [CrossRef]
- Lim, H.S.; Blann, A.D.; Chong, A.Y.; Freestone, B.; Lip, G.Y. Plasma vascular endothelial growth factor, angiopoietin-1, and angiopoietin-2 in diabetes: Implications for cardiovascular risk and effects of multifactorial intervention. Diabetes Care 2004, 27, 2918–2924. [Google Scholar] [CrossRef] [Green Version]
- Tsigkos, S.; Koutsilieris, M.; Papapetropoulos, A. Angiopoietins in angiogenesis and beyond. Expert Opin. Investig. Drugs 2003, 12, 933–941. [Google Scholar] [CrossRef]
- Iuliano, L.; Micheletta, F.; Natoli, S.; Ginanni Corradini, S.; Iappelli, M.; Elisei, W.; Giovannelli, L.; Violi, F.; Diczfalusy, U. Measurement of oxysterols and α-tocopherol in plasma and tissue samples as indices of oxidant stress status. Anal. Biochem. 2003, 312, 217–223. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
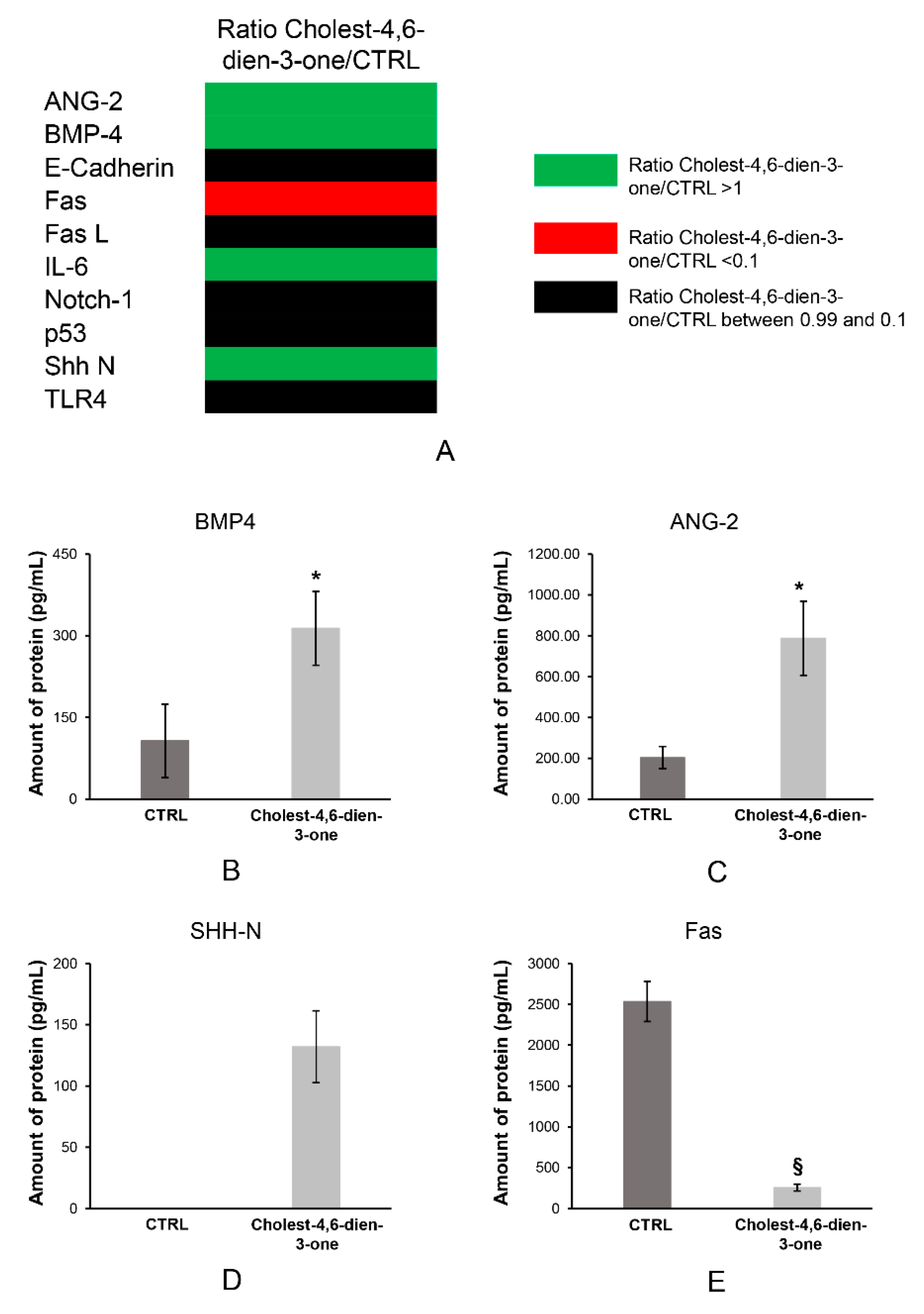{kind=link}
{kind=link}
| Sample | Ct | ΔCq | Activity hTERT | Mean & SD |
|---|---|---|---|---|
| KM 1 | 17.21 | -2.32 | 4.99 | 4.40 |
| KM 2 | 16.86 | -2.42 | 5.35 | 0.95 |
| KM 3 | 17.43 | -2.21 | 4.63 | |
| KM 4 | 17.94 | -1.57 | 2.97 | |
| KM 5 | 17.06 | -2.31 | 4.96 | |
| KM 6 | 17.63 | -1.81 | 3.51 | |
| Cholest-4,6-dien-3-one 1 | 19.53 | |||
| Cholest-4,6-dien-3-one 2 | 19.28 | |||
| Cholest-4,6-dien-3-one 3 | 19.64 | |||
| Cholest-4,6-dien-3-one 4 | 19.51 | |||
| Cholest-4,6-dien-3-one 5 | 19.37 | |||
| Cholest-4,6-dien-3-one 6 | 19.44 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nevi, L.; Costantini, D.; Safarikia, S.; Di Matteo, S.; Melandro, F.; Berloco, P.B.; Cardinale, V. Cholest-4,6-Dien-3-One Promote Epithelial-To-Mesenchymal Transition (EMT) in Biliary Tree Stem/Progenitor Cell Cultures In Vitro. Cells 2019, 8, 1443. https://doi.org/10.3390/cells8111443
Nevi L, Costantini D, Safarikia S, Di Matteo S, Melandro F, Berloco PB, Cardinale V. Cholest-4,6-Dien-3-One Promote Epithelial-To-Mesenchymal Transition (EMT) in Biliary Tree Stem/Progenitor Cell Cultures In Vitro. Cells. 2019; 8(11):1443. https://doi.org/10.3390/cells8111443
Chicago/Turabian StyleNevi, Lorenzo, Daniele Costantini, Samira Safarikia, Sabina Di Matteo, Fabio Melandro, Pasquale Bartolomeo Berloco, and Vincenzo Cardinale. 2019. "Cholest-4,6-Dien-3-One Promote Epithelial-To-Mesenchymal Transition (EMT) in Biliary Tree Stem/Progenitor Cell Cultures In Vitro" Cells 8, no. 11: 1443. https://doi.org/10.3390/cells8111443