The Deubiquitinating Enzyme Inhibitor PR-619 Enhances the Cytotoxicity of Cisplatin via the Suppression of Anti-Apoptotic Bcl-2 Protein: In Vitro and In Vivo Study

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Reagents and Antibodies
2.3. Measurement of Cell Viability
2.4. Western Blot Analysis
2.5. Apoptosis Assay
2.6. Cell Cycle Analysis by Flow Cytometry
2.7. Combination Effect of PR-619 and Cisplatin
2.8. Immunohistochemistry (IHC) in Human UC Specimens
2.9. In Vivo Xenograft Experiments
2.10. Statistical Analysis
3. Results
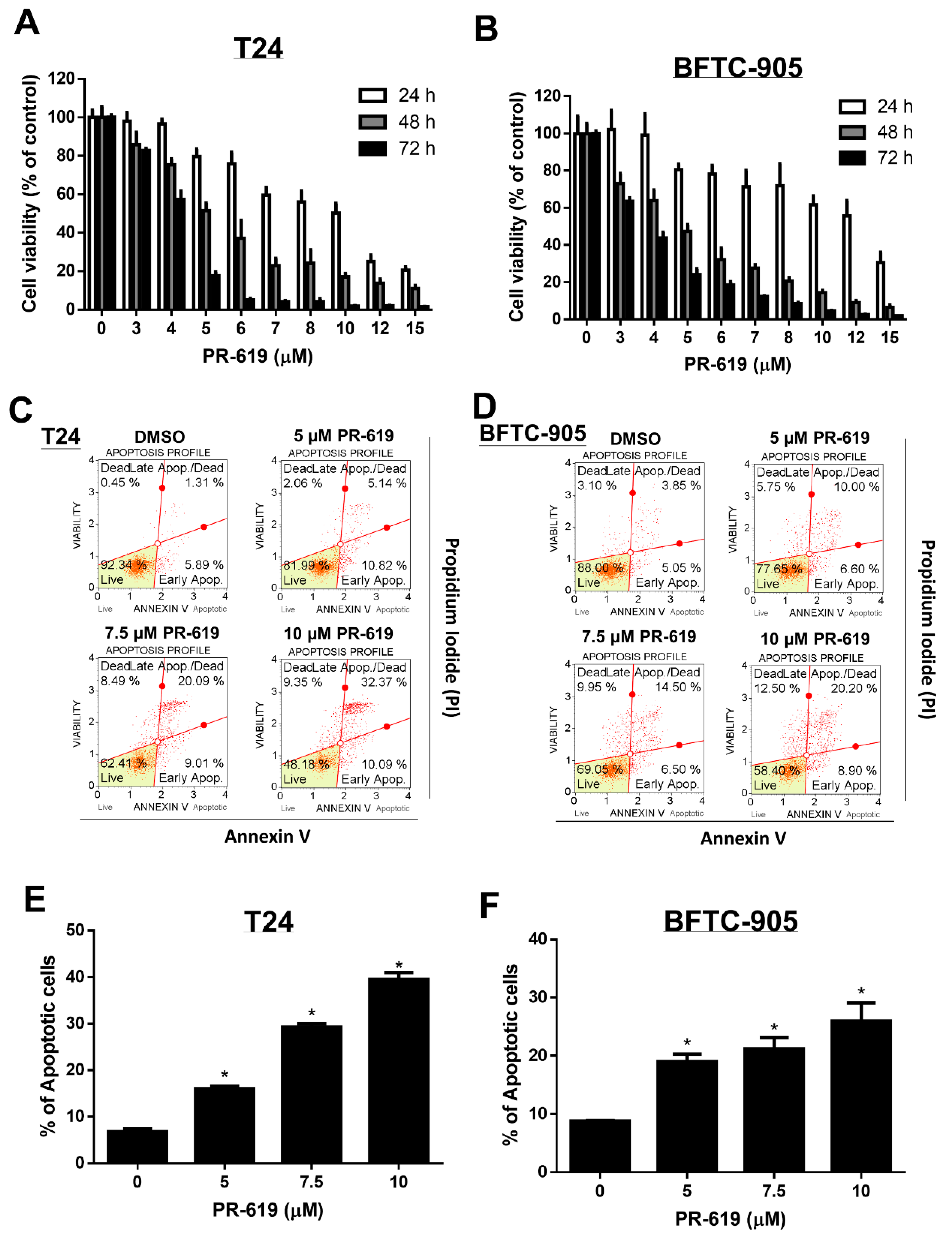
3.1. PR-619 Induced Cytotoxicity and Apoptosis in Human UC Cells in a Dose-dependent and Time-Dependent Manner
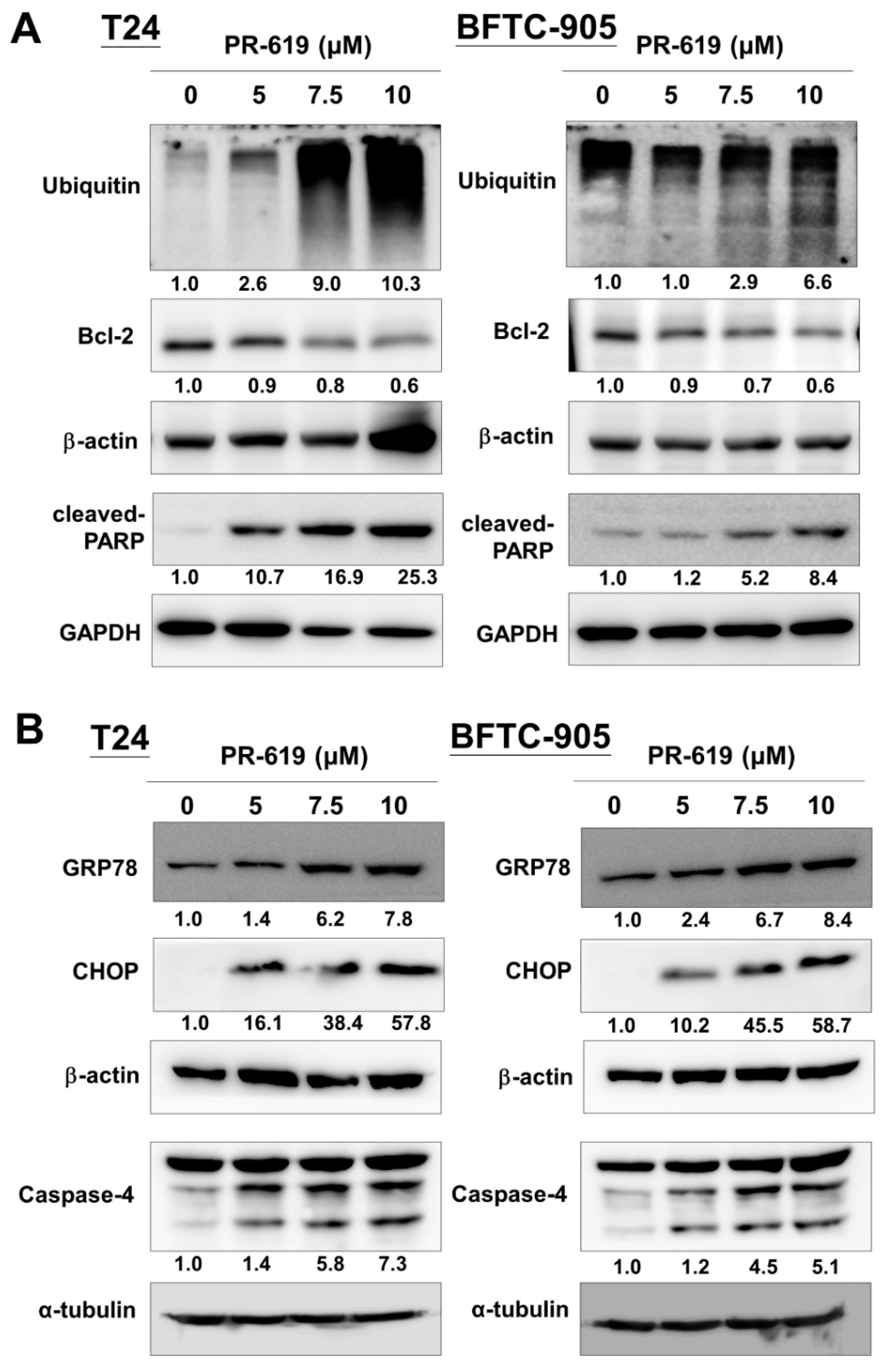
3.2. PR-619 Induced ER Stress and ER-Stress Related Apoptosis in Human UC Cells
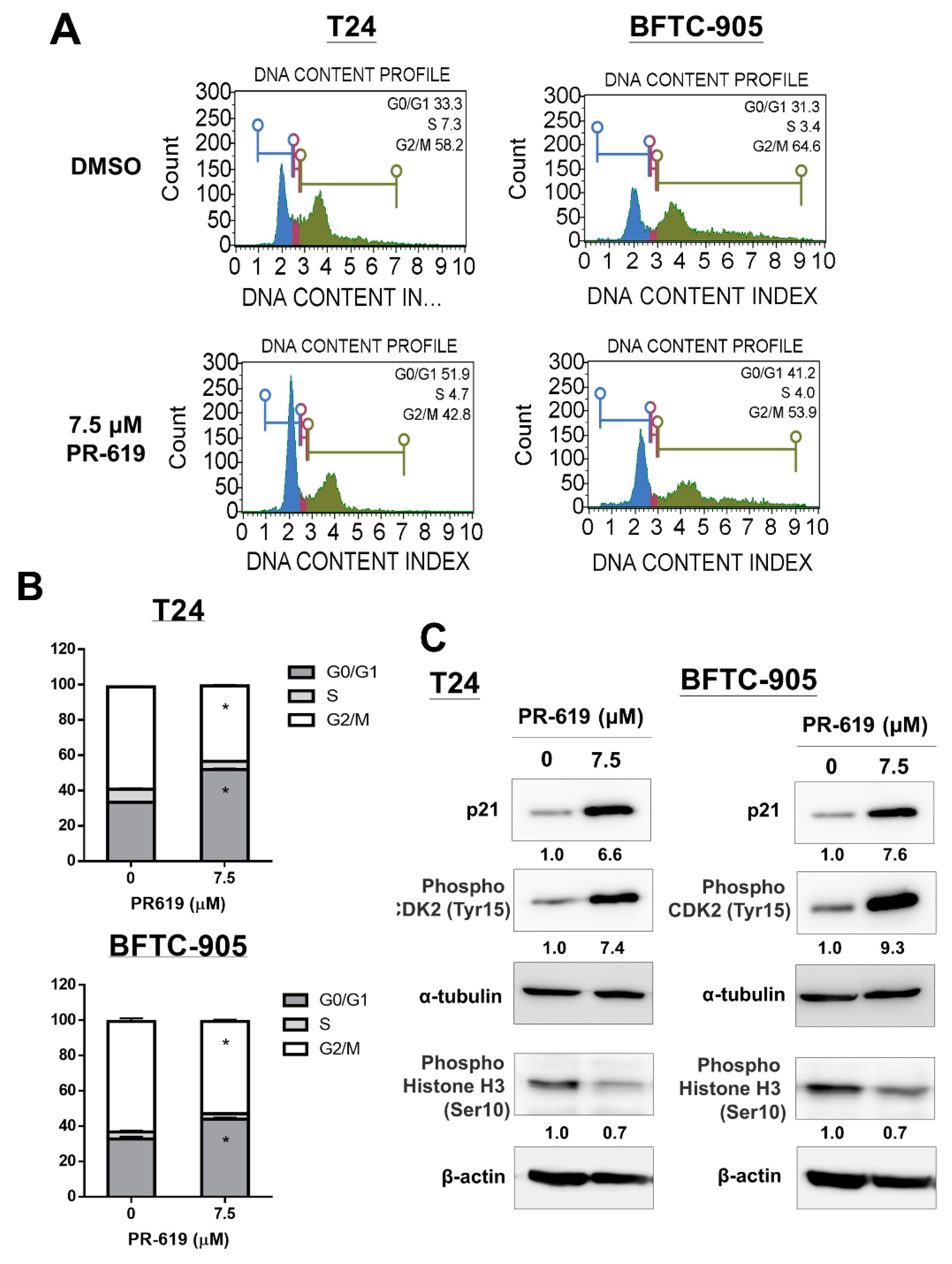
3.3. PR-619 Induced G0/G1 Arrest in UC Cells
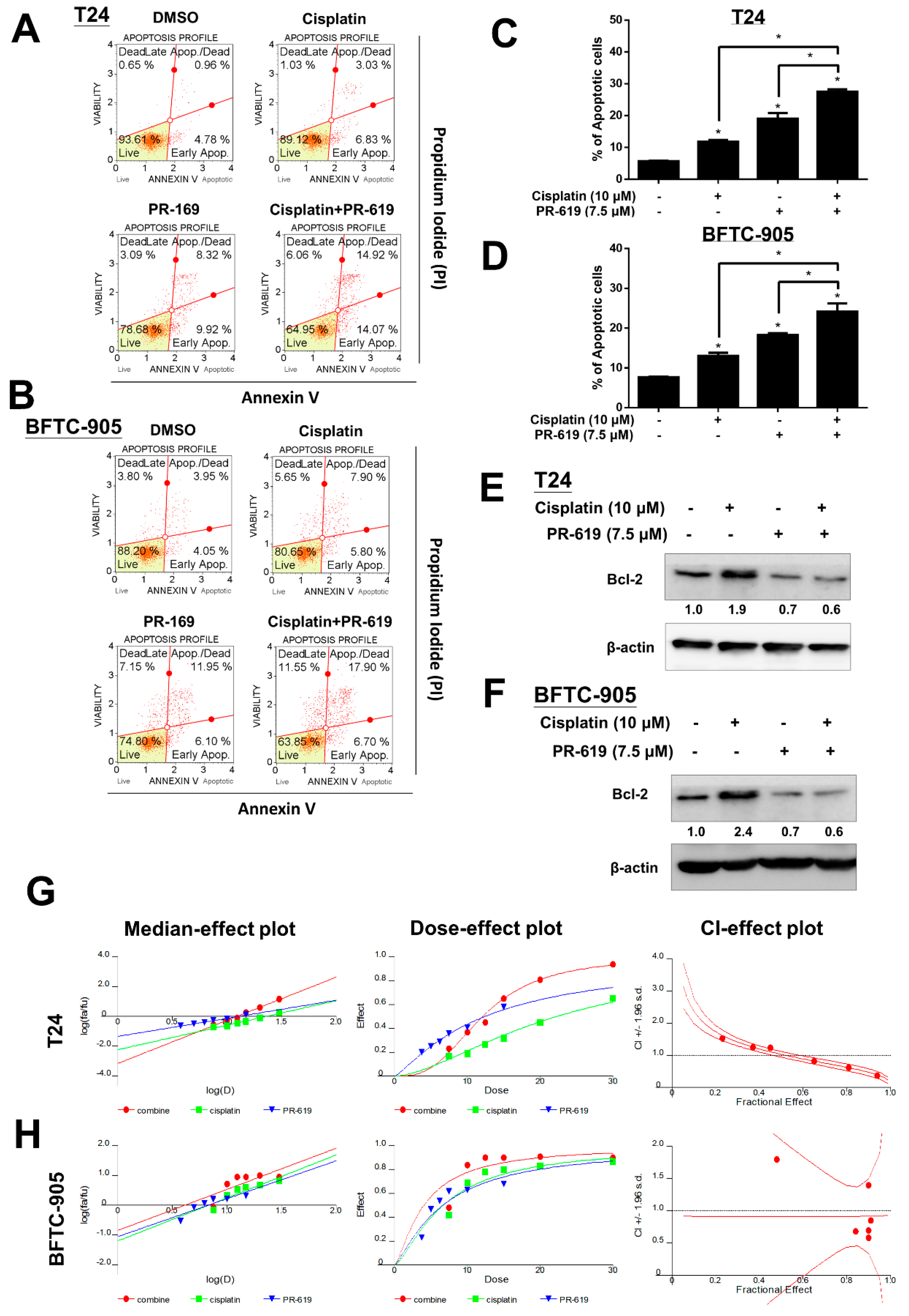
3.4. PR-619 Enhanced Cisplatin-Induced Cytotoxicity in Human UC Cells
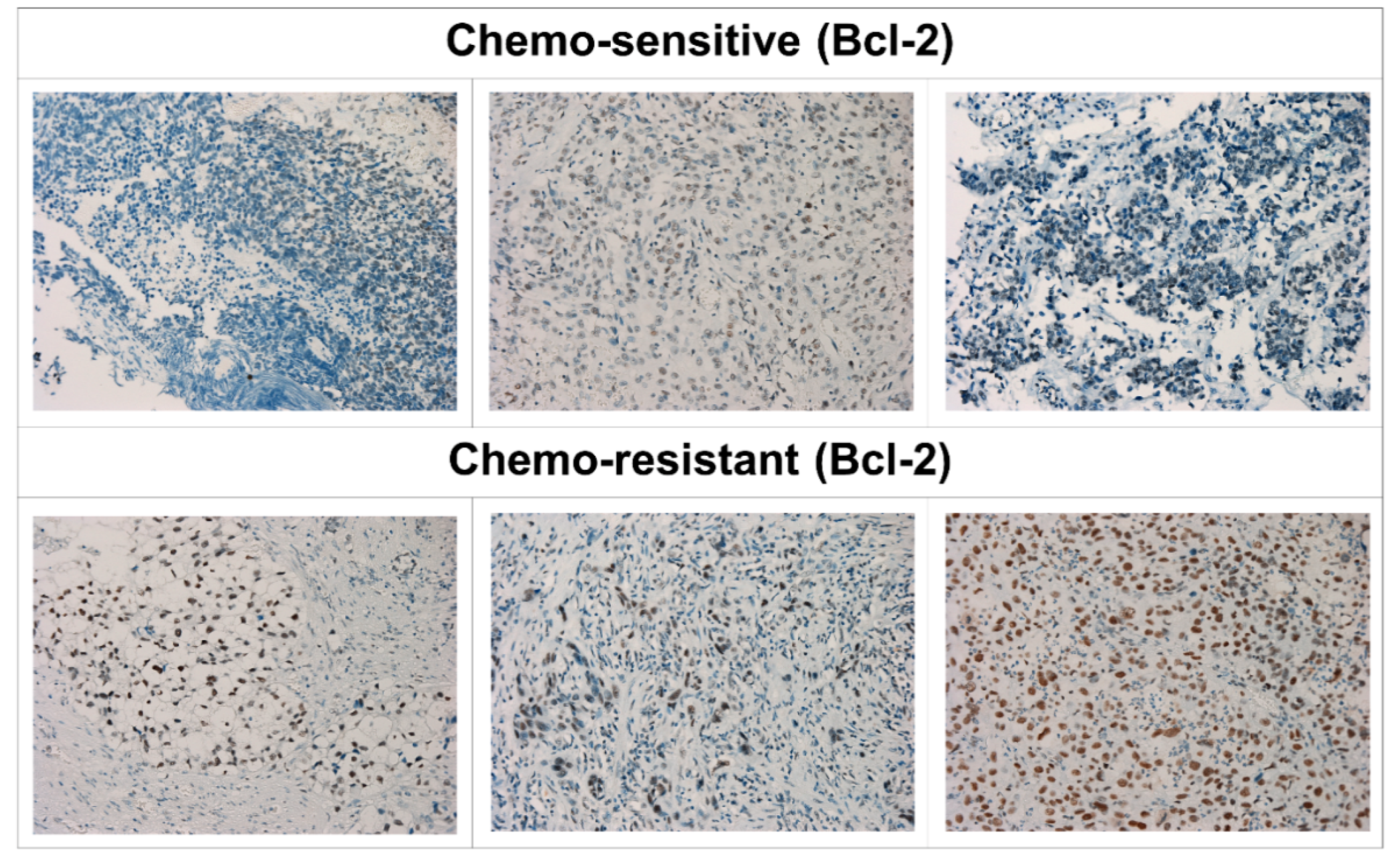
3.5. Overexpression of Bcl-2 Was Associated with Chemotherapy Resistance in Patients with Metastatic UC
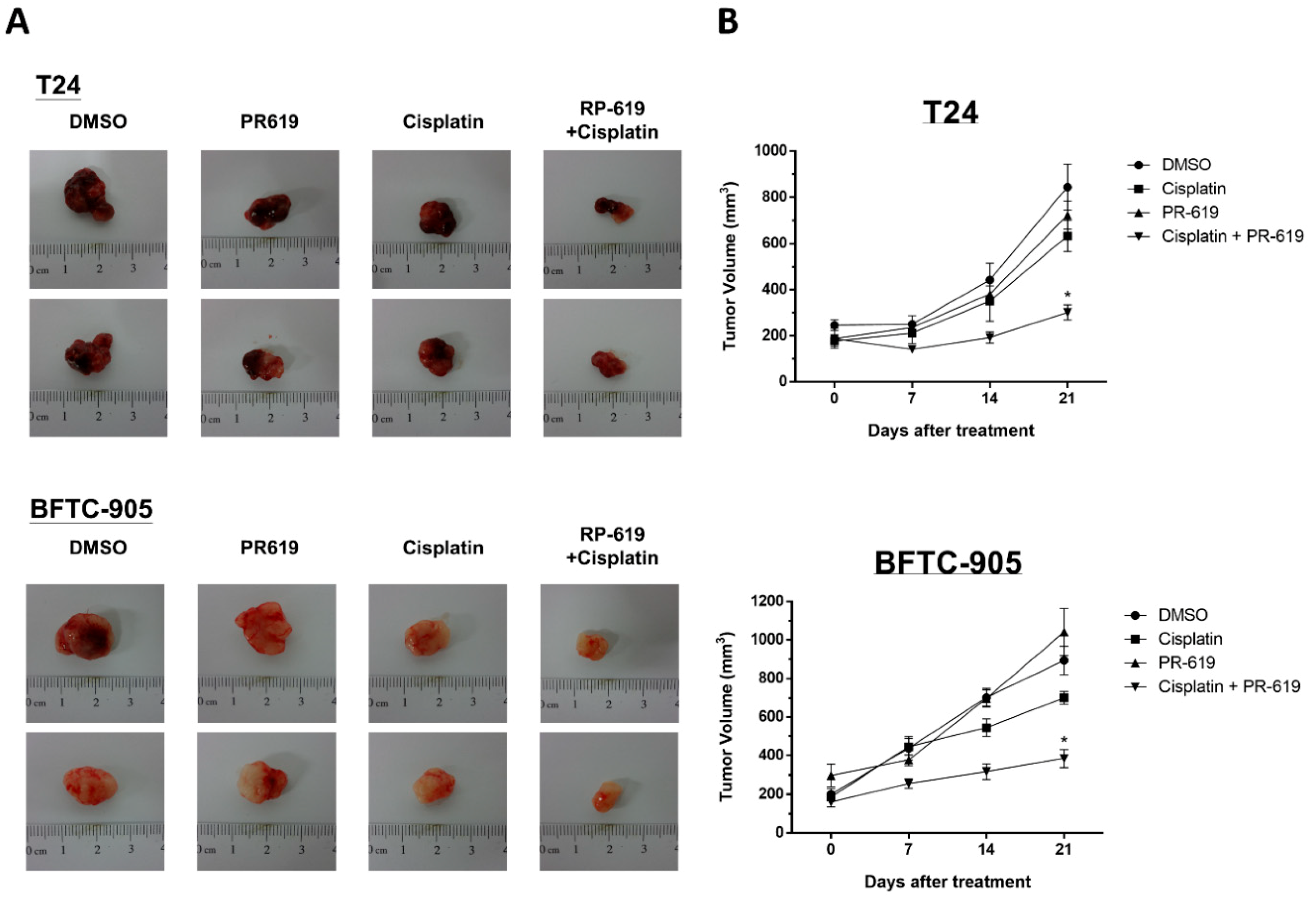
3.6. PR-619 Enhanced the Antitumor Effect of Cisplatin on a Cisplatin-Naïve and Cisplatin-Resistant UC Xenograft of Nude Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Bcl-2 | B-cell lymphoma-2 |
| UC | urothelial carcinoma |
| DUB | deubiquitinating enzyme |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef]
- Cohen, M.H.; Rothmann, M.; Von Der Maase, H.; Hayden, A.M. Gemcitabine and Cisplatin for Advanced, Metastatic Bladder Cancer. J. Clin. Oncol. 2001, 19, 1229–1231. [Google Scholar] [CrossRef]
- Von Der Maase, H.; Hansen, S.; Roberts, J.; Dogliotti, L.; Oliver, T.; Moore, M.; Bodrogi, I.; Albers, P.; Knuth, A.; Lippert, C.; et al. Gemcitabine and Cisplatin Versus Methotrexate, Vinblastine, Doxorubicin, and Cisplatin in Advanced or Metastatic Bladder Cancer: Results of a Large, Randomized, Multinational, Multicenter, Phase III Study. J. Clin. Oncol. 2000, 18, 3068–3077. [Google Scholar] [CrossRef]
- Von Der Maase, H.; Sengelov, L.; Roberts, J.T.; Ricci, S.; Dogliotti, L.; Oliver, T.; Moore, M.J.; Zimmermann, A.; Arning, M. Long-Term Survival Results of a Randomized Trial Comparing Gemcitabine Plus Cisplatin, With Methotrexate, Vinblastine, Doxorubicin, Plus Cisplatin in Patients with Bladder Cancer. J. Clin. Oncol. 2005, 23, 4602–4608. [Google Scholar] [CrossRef]
- Schwartz, A.L.; Ciechanover, A. Targeting Proteins for Destruction by the Ubiquitin System: Implications for Human Pathobiology. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 73–96. [Google Scholar] [CrossRef] [Green Version]
- Amerik, A.Y.; Hochstrasser, M. Mechanism and function of deubiquitinating enzymes. Biochim. Biophys. Acta (BBA)—Bioenerg. 2004, 1695, 189–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijman, S.M.; Luna-Vargas, M.P.; Velds, A.; Brummelkamp, T.R.; Dirac, A.M.; Sixma, T.K.; Bernards, R. A Genomic and Functional Inventory of Deubiquitinating Enzymes. Cell 2005, 123, 773–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yang, Q.; Chen, J.; Zhang, P.; Huang, Q.; Zhang, X.; Yang, L.; Xu, D.; Zhao, C.; Wang, X.; et al. Inhibition of Proteasomal Deubiquitinase by Silver Complex Induces Apoptosis in Non-Small Cell Lung Cancer Cells. Cell. Physiol. Biochem. 2018, 49, 780–797. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, Q.; Xiao, L.; Tang, D.; Dou, Q.P.; Liu, J. Metal-based proteasomal deubiquitinase inhibitors as potential anticancer agents. Cancer Metastasis Rev. 2017, 36, 655–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Chen, X.; Zang, D.; Lan, X.; Liao, S.; Yang, C.; Zhang, P.; Wu, J.; Li, X.; Liu, N.; et al. A novel nickel complex works as a proteasomal deubiquitinase inhibitor for cancer therapy. Oncogene 2016, 35, 5916–5927. [Google Scholar] [CrossRef] [PubMed]
- Suresh, B.; Lee, J.; Kim, H.; Ramakrishna, S. Regulation of pluripotency and differentiation by deubiquitinating enzymes. Cell Death Differ. 2016, 23, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Isamiddinova, N.S.; Peroutka, R.J.; Goldenberg, S.J.; Mattern, M.R.; Nicholson, B.; Leach, C. Characterization of Selective Ubiquitin and Ubiquitin-Like Protease Inhibitors Using a Fluorescence-Based Multiplex Assay Format. ASSAY Drug Dev. Technol. 2011, 9, 165–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullally, J.E.; Moos, P.J.; Edes, K.; Fitzpatrick, F.A. Cyclopentenone Prostaglandins of the J Series Inhibit the Ubiquitin Isopeptidase Activity of the Proteasome Pathway. J. Boil. Chem. 2001, 276, 30366–30373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, K.-L.; Liu, S.-H.; Lin, W.-C.; Hsu, F.-S.; Chow, P.-M.; Chang, Y.-W.; Yang, S.-P.; Shi, C.-S.; Hsu, C.-H.; Liao, S.-M.; et al. Trifluoperazine, an Antipsychotic Drug, Effectively Reduces Drug Resistance in Cisplatin-Resistant Urothelial Carcinoma Cells via Suppressing Bcl-xL: An In Vitro and In Vivo Study. Int. J. Mol. Sci. 2019, 20, 3218. [Google Scholar] [CrossRef]
- Kuo, K.-L.; Ho, I.-L.; Shi, C.-S.; Wu, J.-T.; Lin, W.-C.; Tsai, Y.-C.; Chang, H.-C.; Chou, C.-T.; Hsu, C.-H.; Hsieh, J.-T.; et al. MLN4924, a novel protein neddylation inhibitor, suppresses proliferation and migration of human urothelial carcinoma: In vitro and in vivo studies. Cancer Lett. 2015, 363, 127–136. [Google Scholar] [CrossRef]
- Hsu, F.-S.; Wu, J.-T.; Lin, J.-Y.; Yang, S.-P.; Kuo, K.-L.; Lin, W.-C.; Shi, C.-S.; Chow, P.-M.; Liao, S.-M.; Pan, C.-I.; et al. Histone Deacetylase Inhibitor, Trichostatin A, Synergistically Enhances Paclitaxel-Induced Cytotoxicity in Urothelial Carcinoma Cells by Suppressing the ERK Pathway. Int. J. Mol. Sci. 2019, 20, 1162. [Google Scholar] [CrossRef]
- Ho, I.-L.; Kuo, K.-L.; Liu, S.-H.-; Chang, H.-C.; Hsieh, J.-T.; Wu, J.-T.; Chiang, C.-K.; Lin, W.-C.; Tsai, Y.-C.; Chou, C.-T.; et al. MLN4924 Synergistically Enhances Cisplatin-induced Cytotoxicity via JNK and Bcl-xL Pathways in Human Urothelial Carcinoma. Sci. Rep. 2015, 5, 16948. [Google Scholar] [CrossRef]
- Chou, T.-C.; Talalay, P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Sartorius, U.A.; Krammer, P.H. Upregulation of Bcl-2 is involved in the mediation of chemotherapy resistance in human small cell lung cancer cell lines. Int. J. Cancer 2002, 97, 584–592. [Google Scholar] [CrossRef]
- Todi, S.V.; Paulson, H.L. Balancing act: deubiquitinating enzymes in the nervous system. Trends Neurosci. 2011, 34, 370–382. [Google Scholar] [CrossRef] [Green Version]
- Altun, M.; Kramer, H.B.; Willems, L.I.; McDermott, J.L.; Leach, C.A.; Goldenberg, S.J.; Kumar, K.S.; Konietzny, R.; Fischer, R.; Kogan, E.; et al. Activity-Based Chemical Proteomics Accelerates Inhibitor Development for Deubiquitylating Enzymes. Chem. Boil. 2011, 18, 1401–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, T.J. The Cancer Genome Atlas Research Network: A Sight to Behold. Endocr. Pathol. 2014, 25, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-H.; Kuo, K.-L.; Chen, S.-C.; Weng, T.-I.; Chuang, Y.-T.; Tsai, Y.-C.; Pu, Y.-S.; Chiang, C.-K.; Liu, S.-H. Down-Regulation of Glucose-Regulated Protein (GRP) 78 Potentiates Cytotoxic Effect of Celecoxib in Human Urothelial Carcinoma Cells. PLoS ONE 2012, 7, e33615. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, J.M.; Soane, L. Multiple Functions of BCL-2 Family Proteins. Cold Spring Harb. Perspect. Boil. 2013, 5, a008722. [Google Scholar] [CrossRef]
- Trisciuoglio, D.; Tupone, M.G.; Desideri, M.; Di Martile, M.; Gabellini, C.; Buglioni, S.; Pallocca, M.; Alessandrini, G.; D’Aguanno, S.; Del Bufalo, D. BCL-XL overexpression promotes tumor progression-associated properties. Cell Death Dis. 2017, 8, 3216. [Google Scholar] [CrossRef]
- Thomas, S.; Quinn, B.A.; Das, S.K.; Dash, R.; Emdad, L.; Dasgupta, S.; Wang, X.Y.; Dent, P.; Reed, J.C.; Pellecchia, M.; et al. Targeting the Bcl-2 family for cancer therapy. Expert Opin. Targets 2013, 17, 61–75. [Google Scholar] [CrossRef]
- Reed, J.C. Bcl-2 on the brink of breakthroughs in cancer treatment. Cell Death Differ. 2018, 25, 3–6. [Google Scholar] [CrossRef]
- García-Aranda, M.; Pérez-Ruiz, E.; Redondo, M. Bcl-2 Inhibition to Overcome Resistance to Chemo- and Immunotherapy. Int. J. Mol. Sci. 2018, 19, 3950. [Google Scholar] [CrossRef]
- Schenk, R.L.; Strasser, A.; Dewson, G. BCL-2: Long and winding path from discovery to therapeutic target. Biochem. Biophys. Res. Commun. 2017, 482, 459–469. [Google Scholar] [CrossRef]
- Bodur, C.; Basaga, H. Bcl-2 inhibitors: emerging drugs in cancer therapy. Curr. Med. Chem. 2012, 19, 1804–1820. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, K.-L.; Liu, S.-H.; Lin, W.-C.; Chow, P.-M.; Chang, Y.-W.; Yang, S.-P.; Shi, C.-S.; Hsu, C.-H.; Liao, S.-M.; Chang, H.-C.; et al. The Deubiquitinating Enzyme Inhibitor PR-619 Enhances the Cytotoxicity of Cisplatin via the Suppression of Anti-Apoptotic Bcl-2 Protein: In Vitro and In Vivo Study. Cells 2019, 8, 1268. https://doi.org/10.3390/cells8101268
Kuo K-L, Liu S-H, Lin W-C, Chow P-M, Chang Y-W, Yang S-P, Shi C-S, Hsu C-H, Liao S-M, Chang H-C, et al. The Deubiquitinating Enzyme Inhibitor PR-619 Enhances the Cytotoxicity of Cisplatin via the Suppression of Anti-Apoptotic Bcl-2 Protein: In Vitro and In Vivo Study. Cells. 2019; 8(10):1268. https://doi.org/10.3390/cells8101268
Chicago/Turabian StyleKuo, Kuan-Lin, Shing-Hwa Liu, Wei-Chou Lin, Po-Ming Chow, Yu-Wei Chang, Shao-Ping Yang, Chung-Sheng Shi, Chen-Hsun Hsu, Shih-Ming Liao, Hong-Chiang Chang, and et al. 2019. "The Deubiquitinating Enzyme Inhibitor PR-619 Enhances the Cytotoxicity of Cisplatin via the Suppression of Anti-Apoptotic Bcl-2 Protein: In Vitro and In Vivo Study" Cells 8, no. 10: 1268. https://doi.org/10.3390/cells8101268