Stromal Cell-Derived Factor 1 Protects Brain Vascular Endothelial Cells from Radiation-Induced Brain Damage

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Animals
2.2. Transfection of CXCR4 siRNA
2.3. Ribonucleic Acid Extraction and Real-Time Polymerase Chain Reaction Analysis
2.4. Western Blotting
2.5. Enzyme-Linked Immunosorbent Assay
2.6. Cell Growth Assay
2.7. Senescence-Associated Beta-Galactosidase Staining
2.8. Tube Formation Assay
2.9. γH2AX Foci Staining
2.10. Radiation and Treatment with CXCR4 Agonist ATI2341
2.11. Transendothelial Electrical Resistance and Permeability Assay
2.12. TUNEL Assay in the Small Intestine
2.13. Brain Blood Vessel Isolation
2.14. Blood Flow Analysis
2.15. Novel Object Recognition Memory Test
2.16. Statistical Analysis
3. Results
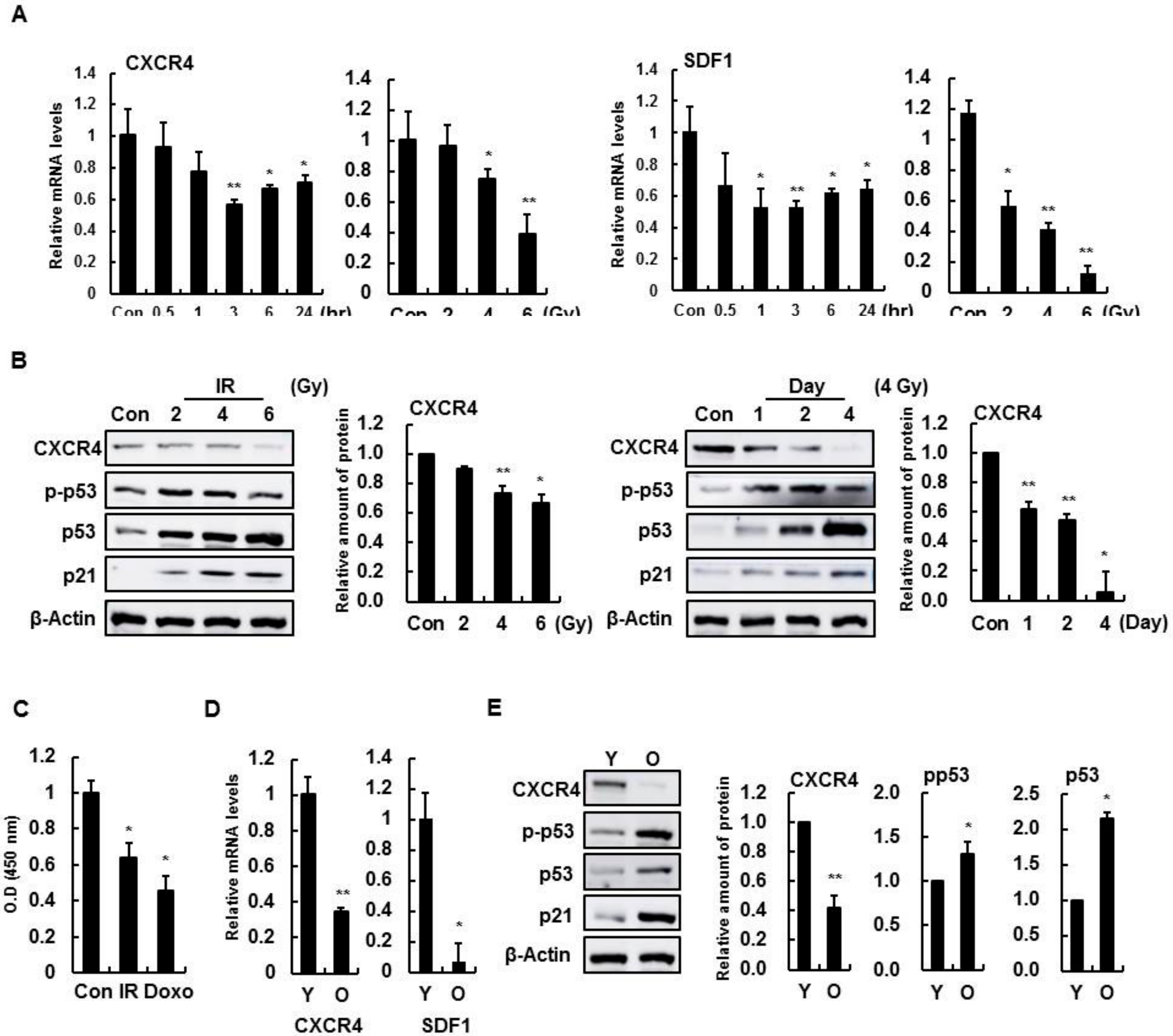
3.1. Decline of CXCR4 and SDF-1 Expression with IR Treatment and Aging in Brain Endothelial Cells
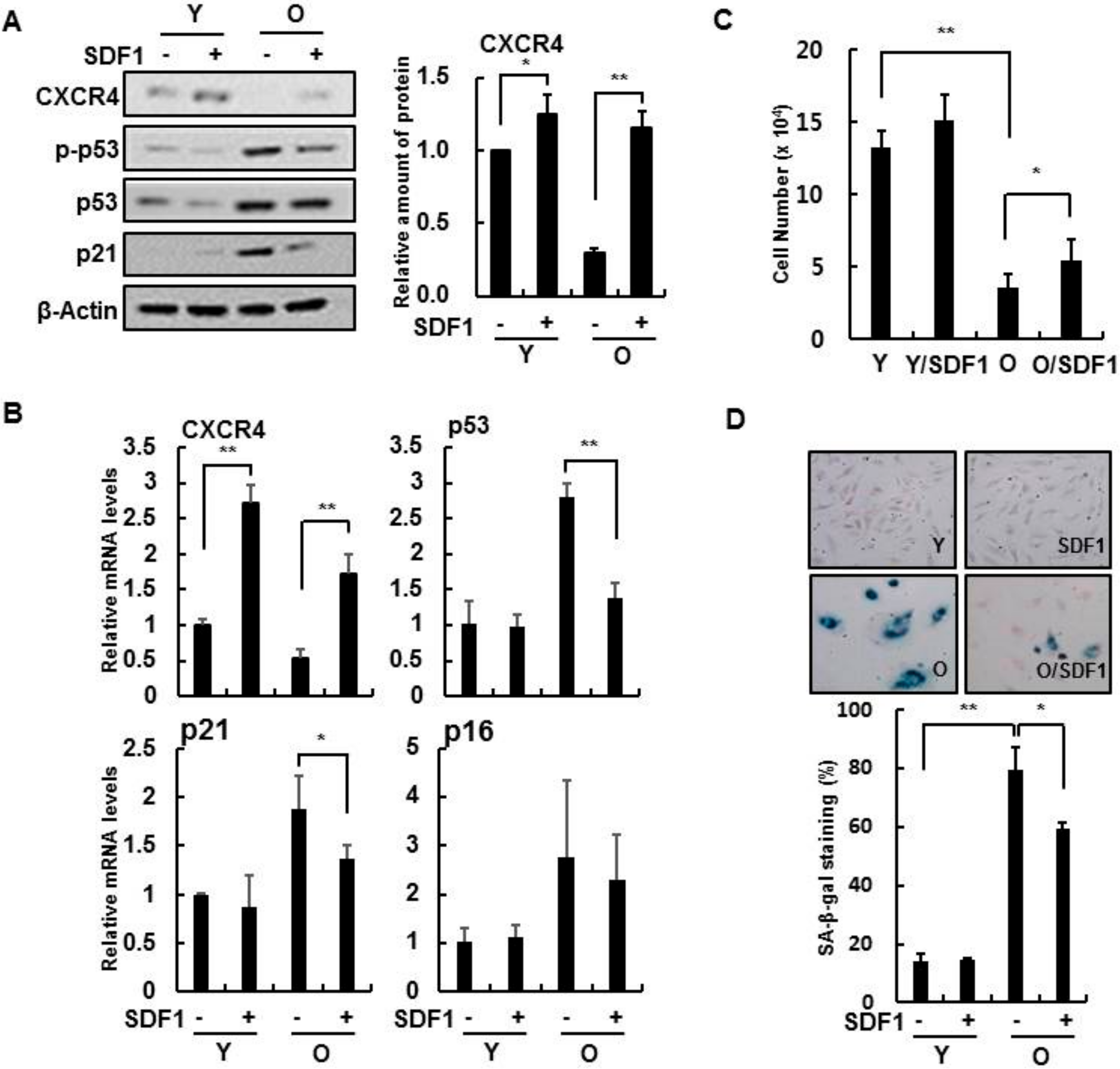
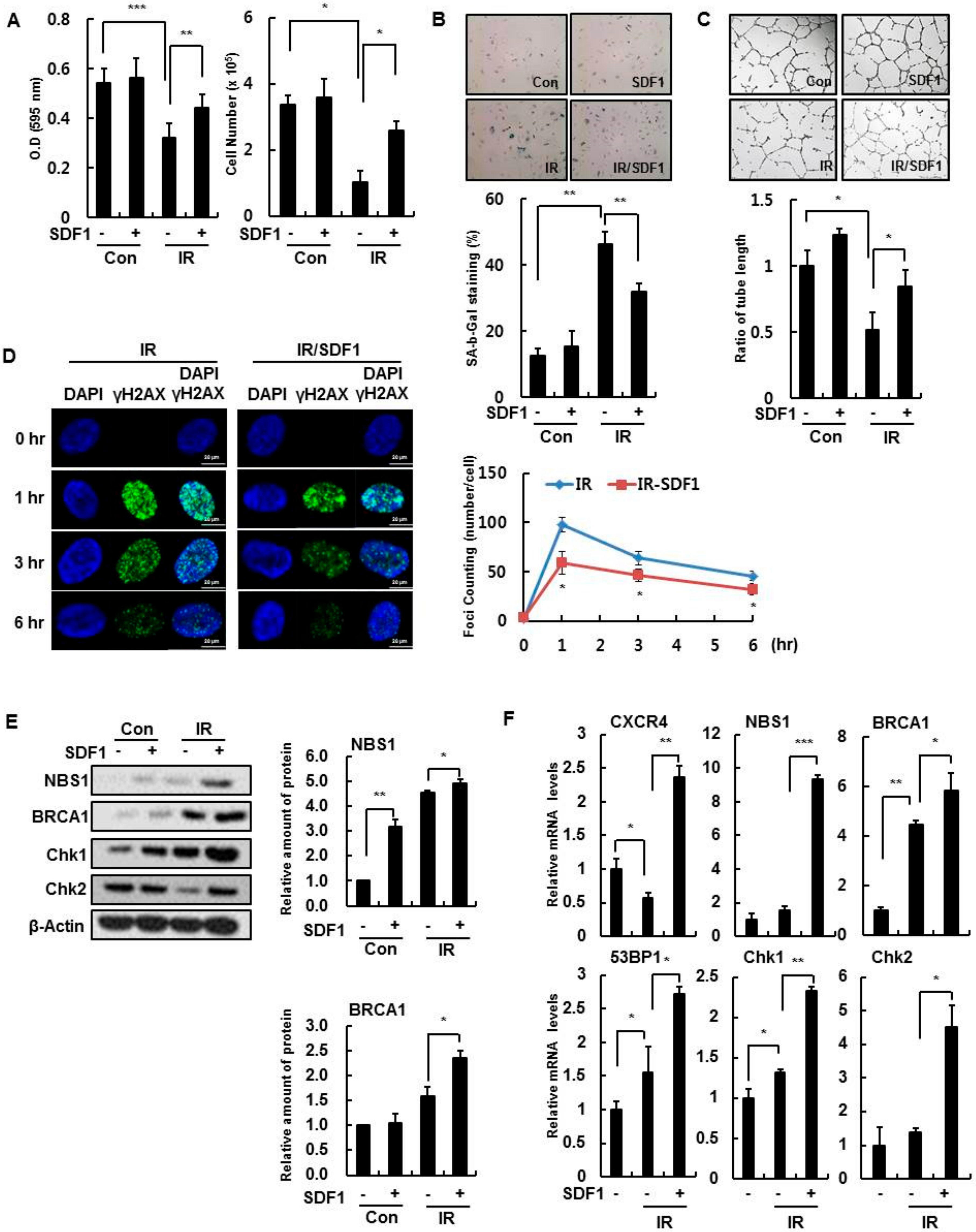
3.2. Effect of SDF-1 on Senescent HBMVECs and IR Exposure
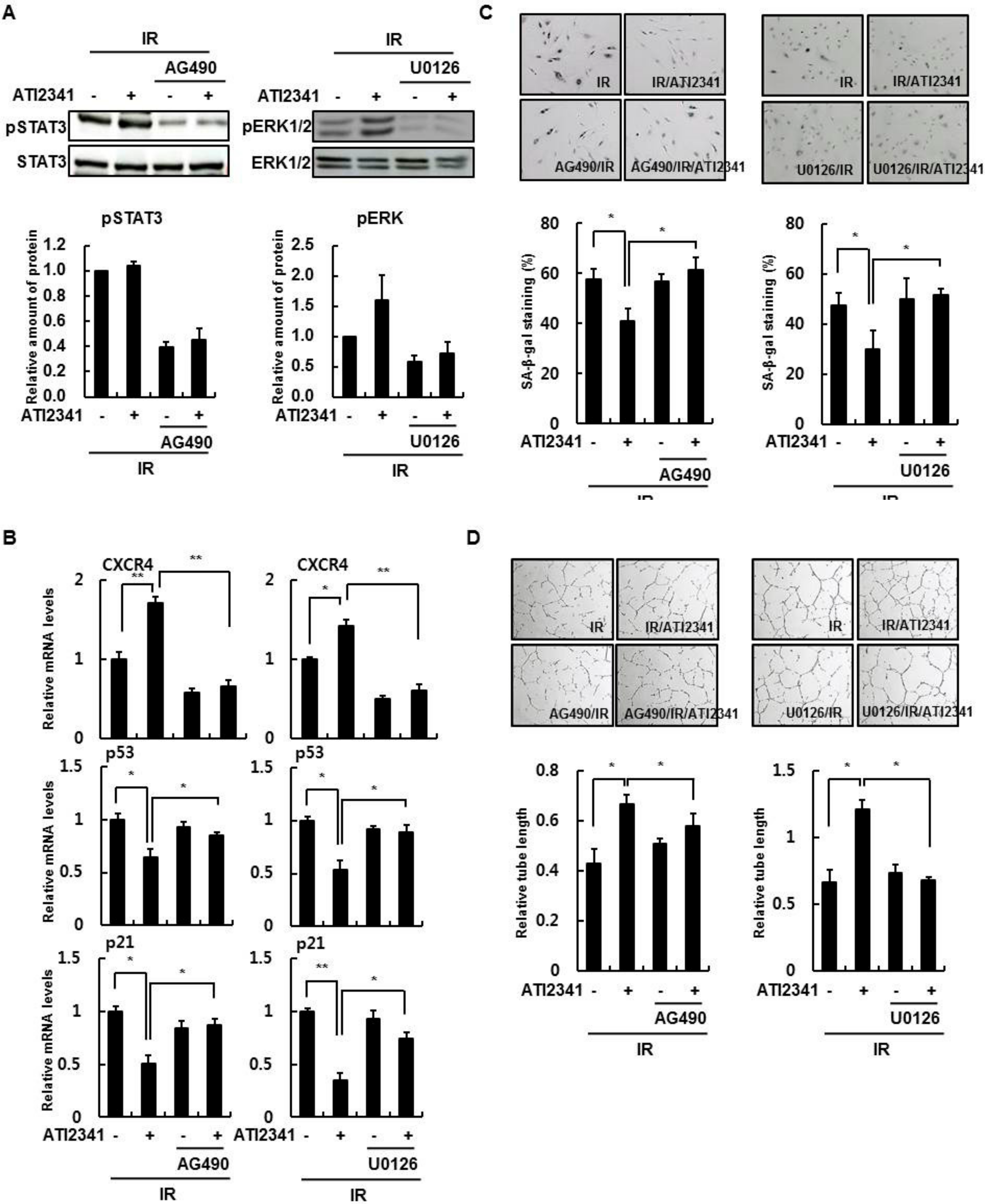
3.3. SDF-1 Plays a Role in a CXCR4-Dependent Mechanism
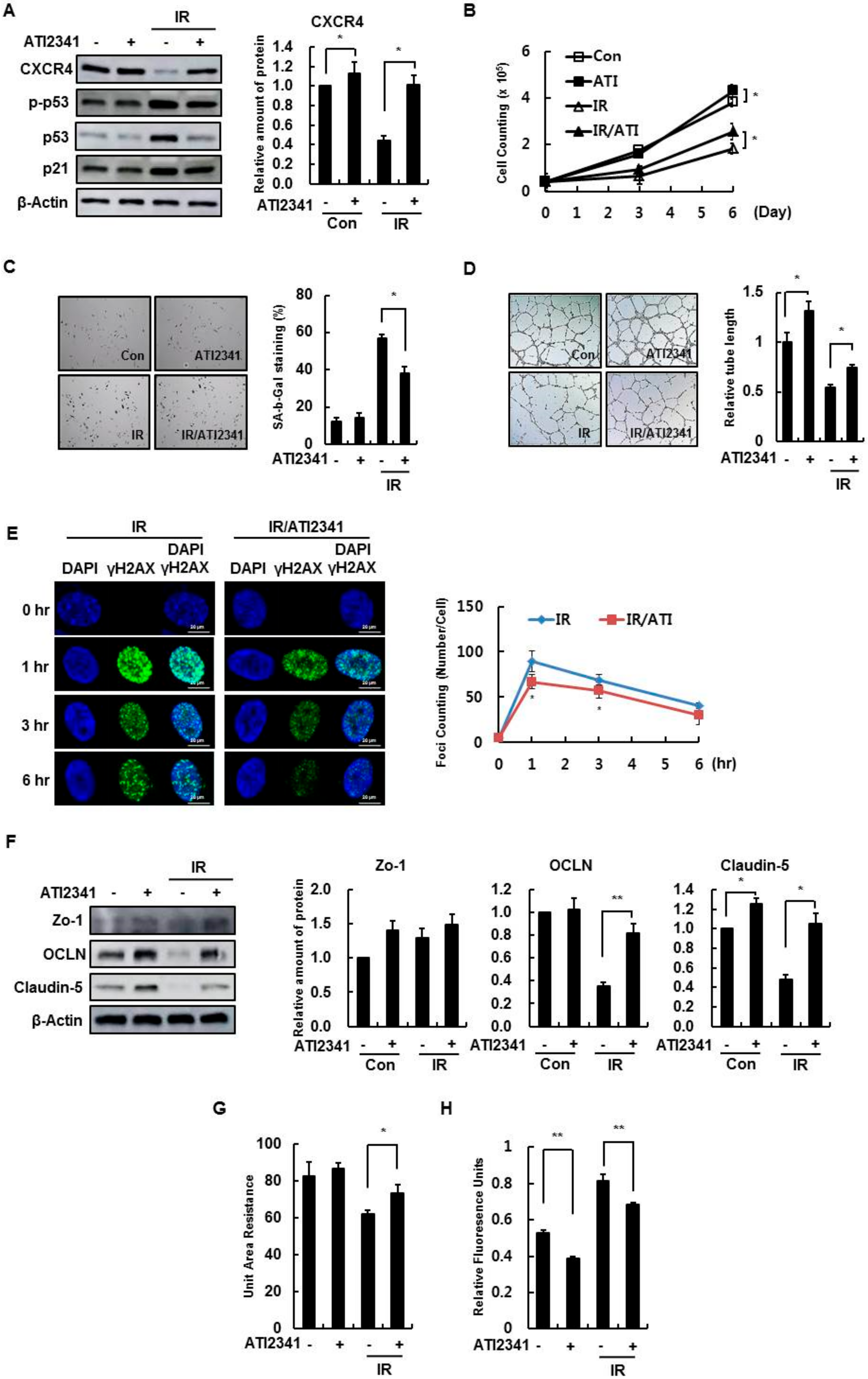
3.4. Effect of A CXCR4 Agonist on HBMVECs
3.5. Activation of STAT3 and ERK Is Important in the Function of CXCR4/SDF-1
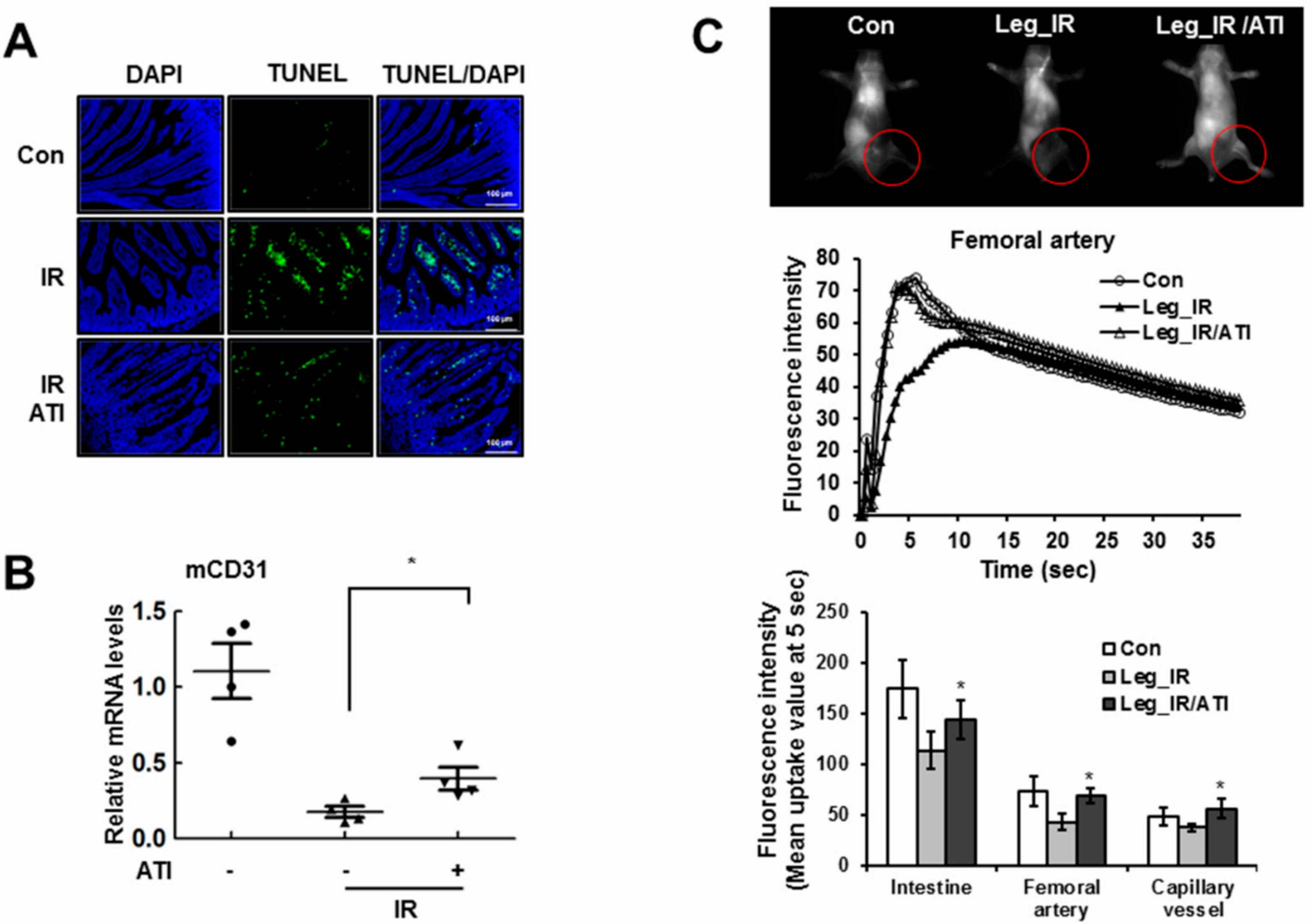
3.6. CXCR4 Agonist Mediates Radioprotection in Mice
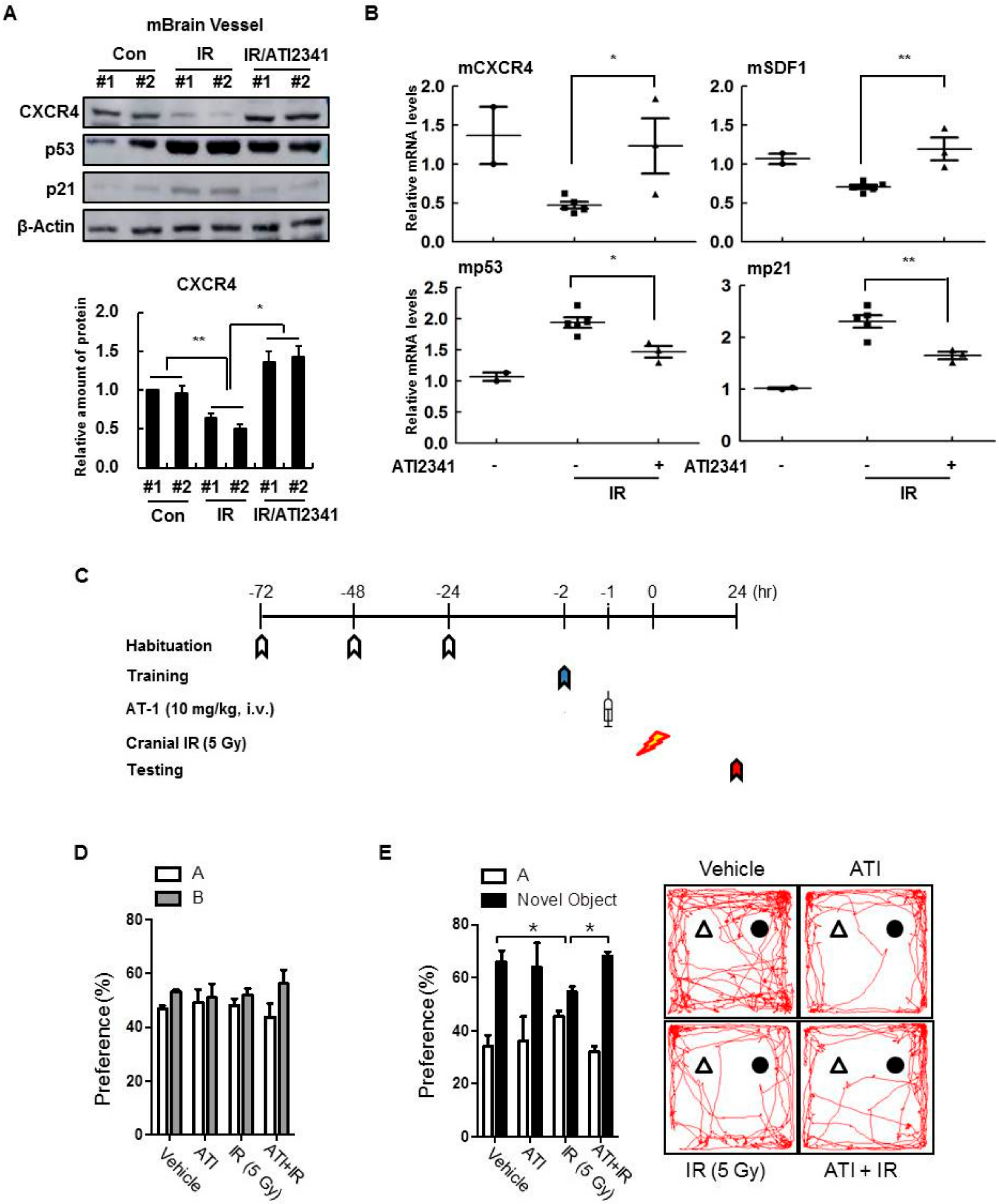
3.7. A CXCR4 Agonist Specifically Protects Mouse Brain Vessels and Contributes to the Improvement of Cognitive Disorders
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lowe, X.R.; Bhattacharya, S.; Marchetti, F.; Wyrobek, A.J. Early Brain Response to Low-Dose Radiation Exposure Involves Molecular Networks and Pathways Associated with Cognitive Functions, Advanced Aging and Alzheimer’s Disease. Radiat. Res. 2009, 171, 53–65. [Google Scholar] [CrossRef]
- Begum, N.; Wang, B.; Mori, M.; Vares, G. Does Ionizing Radiation Influence Alzheimer’s Disease Risk? J. Radiat. Res. 2012, 53, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Paris, F.; Fuks, Z.; Kang, A.; Capodieci, P.; Juan, G.; Ehleiter, D.; Haimovitz-Friedman, A.; Cordon-Cardo, C.; Kolesnick, R. Endothelial Apoptosis as the Primary Lesion Initiating Intestinal Radiation Damage in Mice. Science 2001, 293, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Prolla, T.A.; Mattson, M.P. Molecular Mechanisms of Brain Aging and Neurodegenerative Disorders: Lessons from Dietary Restriction. Trends Neurosci. 2001, 24, S21–S31. [Google Scholar] [CrossRef]
- Farkas, E.; Luiten, P.G. Cerebral Microvascular Pathology in Aging and Alzheimer’s Disease. Prog. Neurobiol. 2001, 64, 575–611. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, L.; Sun, W.L.; Wang, L.; Zhang, W.J.; You, H. Radiation-Induced Endothelial Cell loss and Reduction of the Relative Magnitude of the Blood Flow in the Rat Spinal Cord. Brain Res. 2014, 1583, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Chen, P.; Jain, V.; Reilly, R.M.; Wong, C.S. Early Radiation-Induced Endothelial Cell Loss and Blood-Spinal Cord Barrier Breakdown in the Rat Spinal Cord. Radiat. Res. 2004, 161, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Deak, F.; Freeman, W.M.; Ungvari, Z.; Csiszar, A.; Sonntag, W.E. Recent Developments in Understanding Brain Aging: Implications for Alzheimer’s Disease and Vascular Cognitive Impairment. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2016, 71, 13–20. [Google Scholar] [CrossRef]
- Erdő, F.; Denes, L.; de Lange, E. Age-Associated Physiological and Pathological Changes at the Blood–Brain Barrier: A Review. J. Cereb. Blood Flow Metab. 2017, 37, 4–24. [Google Scholar] [CrossRef]
- Bors, L.; Tóth, K.; Tóth, E.Z.; Bajza, Á.; Csorba, A.; Szigeti, K.; Máthé, D.; Perlaki, G.; Orsi, G.; Tóth, G.K.; et al. Structural and Functional Characterization of Age-Associated Changes at the Blood-Brain Barrier. A Comparative In Vivo Study in Young Adult and Middle Aged Wistar Rats. Brain Res. Bull. 2018, 139, 269–277. [Google Scholar] [CrossRef]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The Ageing Systemic Milieu Negatively Regulates Neurogenesis and Cognitive Function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young Blood Reverses Age-Related Impairments in Cognitive Function and Synaptic Plasticity in Mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Katsimpardi, L.; Litterman, N.K.; Schein, P.A.; Miller, C.M.; Loffredo, F.S.; Wojtkiewicz, G.R.; Chen, J.W.; Lee, R.T.; Wagers, A.J.; Rubin, L.L. Vascular and Neurogenic Rejuvenation of the Aging Mouse Brain by Young Systemic Factors. Science 2014, 344, 630–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozek, C.; Krolewski, R.C.; Buchanan, S.M.; Rubin, L.L. Growth Differentiation Factor 11 Treatment leads to Neuronal and Vascular Improvements in the Hippocampus of Aged Mice. Sci. Rep. 2018, 8, 17293. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, J.; Kusano, K.F.; Masuo, O.; Kawamoto, A.; Silver, M.; Murasawa, S.; Bosch-Marce, M.; Masuda, H.; Losordo, D.W.; Isner, J.M.; et al. Stromal Cell-Derived Factor-1 Effects on ex vivo Expanded Endothelial Progenitor Cell Recruitment for Ischemic Neovascularization. Circulation 2003, 107, 1322–1328. [Google Scholar] [CrossRef]
- Zernecke, A.; Schober, A.; Bot, I.; von Hundelshausen, P.; Liehn, E.A.; Mopps, B. SDF-1alpha/CXCR4 Axis is Instrumental in Neointimal Hyperplasia and Recruitment of Smoothe Muscle Progenitor Cells. Circ. Res. 2005, 96, 784–791. [Google Scholar] [CrossRef]
- Ganju, R.K.; Brubaker, S.A.; Meyer, J.; Dutt, P.; Yang, Y.; Qin, S.; Newman, W.; Groopman, J.E. The Alpha-Chemokine, Stromal Cell-Derived Factor-1Alpha, Binds to the Transmembrane G-Protein-Coupled CXCR-4 Receptor and Activates Multiple Signal Transduction Pathways. J. Biol. Chem. 1998, 273, 23169–23175. [Google Scholar] [CrossRef]
- Kucia, M.; Dawn, B.; Hunt, G.; Guo, Y.; Wysoczynski, M.; Majka, M. Cells Expressing Early Cardiac Markers Reside in the Bone Marrow and are Mobilized into the Peripheral Blood after Myocardial Infarction. Circ. Res. 2004, 95, 1191–1199. [Google Scholar] [CrossRef]
- Levoye, A.; Balabanian, K.; Baleux, F.; Bachelerie, F.; Lagane, B. CXCR7 Heterodimerizes with CXCR4 and Regulates CXCL12-Mediated G Protein Signaling. Blood 2009, 113, 6085–6093. [Google Scholar] [CrossRef]
- Decaillot, F.M.; Kazmi, M.A.; Lin, Y.; Ray-Saha, S.; Sakmar, T.P.; Sachdev, P. CXCR7/CXCR4 Heterodimer Constitutively Recruits Beta-Arrestin to Enhance Cell Migration. J. Biol. Chem. 2011, 286, 32188–32197. [Google Scholar] [CrossRef]
- Wang, F.; Yamauchi, M.; Muramatsu, M.; Osawa, T.; Tsuchida, R.; Shibuya, M. RACK1 Regulates VEGF/Flt1-Mediated Cell Migration via Activation of A PI3K/Akt Pathway. J. Biol. Chem. 2011, 286, 9097–9106. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, W.E.; Ramsey, M.; Carter, C.S. Growth Hormone and Insulin-Like Growth Factor-1 (IGF-1) and Their Influence on Cognitive Aging. Ageing Res. Rev. 2005, 4, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Middeldorp, J.; Lehallier, B.; Villeda, S.A.; Miedema, S.S.; Evans, E.; Czirr, E.; Zhang, H.; Luo, J.; Stan, T.; Mosher, K.I.; et al. Preclinical Assessment of Young Blood Plasma for Alzheimer Disease. JAMA Neurol. 2016, 73, 1325–1333. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, H.; Yung, A.; Villeda, S.A.; Jaeger, P.A.; Olayiwola, O.; Fainberg, N.; Wyss-Coray, T. ALK5-Dependent TGF-Beta Signaling is A Major Determinant of Late-Stage Adult Neurogenesis. Nat. Neurosci. 2014, 17, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.I.; Kim, W.; Choi, K.J.; Bae, S.; Jeong, J.H.; Kim, K.S. XIAP-Associating Factor 1, A Transcriptional Target of BRD7, Contributes to Endothelial Cell Senescence. Oncotarget 2016, 7, 5118–5130. [Google Scholar] [CrossRef] [PubMed]
- Boulay, A.C.; Saubamea, B.; Decleves, X.; Cohen-Salmon, M. Purification of Mouse Brain Vessels. J. Vis. Exp. 2015, 105, e53208. [Google Scholar] [CrossRef]
- Hagihara, H.; Toyama, K.; Yamasaki, N.; Miyakawa, T. Dissection of Hippocampal Dentate Gyrus from Adult Mouse. J. Vis. Exp. 2009, 33, 1543. [Google Scholar] [CrossRef]
- Ritz, M.F.; Grond-Ginsbach, C.; Kloss, M.; Tolnay, M.; Fluri, F.; Bonati, L.H.; Traenka, C.; Zeis, T.; Schaeren-Wiemers, N.; Peters, N.; et al. Identification of Inflammatory, Metabolic, and Cell Survival Pathways Contributing to Cerebral Small Vessel Disease by Postmortem Gene Expression Microarray. Curr. Neurovasc. Res. 2016, 13, 58–67. [Google Scholar] [CrossRef]
- Cuadrado-Godia, E.; Dwivedi, P.; Sharma, S.; Ois Santiago, A.; Roquer Gonzalez, J.; Balcells, M.; Laird, J.; Turk, M.; Suri, H.S.; Nicolaides, A.; et al. Cerebral Small Vessel Disease: A Review Focusing on Pathophysiology, Biomarkers, and Machine Learning Strategies. J. Stroke 2018, 20, 302–320. [Google Scholar] [CrossRef]
- Pantoni, L. Cerebral Small Vessel Disease: From Pathogenesis and Clinical Characteristics to Therapeutic Challenges. Lancet. Neurol. 2010, 9, 689–701. [Google Scholar] [CrossRef]
- Taylor, W.D.; Aizenstein, H.J.; Alexopoulos, G.S. The Vascular Depression Hypothesis: Mechanisms Linking Vascular Disease with Depression. Mol. Psychiatry 2013, 18, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Pinter, D.; Ritchie, S.J.; Doubal, F.; Gattringer, T.; Morris, Z.; Bastin, M.E.; Del, C.V.H.M.; Royle, N.A.; Corley, J.; Munoz Maniega, S.; et al. Impact of Small Vessel Disease in the Brain on Gait and Balance. Sci. Rep. 2017, 7, 41637. [Google Scholar] [CrossRef] [PubMed]
- Erdő, F.; Krajcsi, P. Age-Related Functional and Expressional Changes in Efflux Pathways at the Blood-Brain Barrier. Front. Aging Neurosci. 2019, 11, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesulu, B.P.; Mahadevan, L.S.; Aliru, M.L.; Yang, X.; Bodd, M.H.; Singh, P.K.; Yusuf, S.W.; Abe, J.I.; Krishnan, S. Radiation-Induced Endothelial Vascular Injury: A Review of Possible Mechanisms. JACC Basic Transl. Sci. 2018, 3, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Orengo, L.; Holman, D.W.; Dorsey, D.; Zhou, L.; Zhang, P.; Wright, M.; McCandless, E.E.; Patel, J.R.; Luker, G.D.; Littman, D.R.; et al. CXCR7 Influences Leukocyte Entry into the CNS Parenchyma by Controlling Abluminal CXCL12 Abundance during Autoimmunity. J. Exp. Med. 2011, 208, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Costello, C.M.; McCullagh, B.; Howell, K.; Sands, M.; Belperio, J.A.; Keane, M.P.; Gaine, S.; McLoughlin, P. A Role for the CXCL12 Receptor, CXCR7, in the Pathogenesis of Human Pulmonary Vascular Disease. Eur. Respir. J. 2012, 39, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Schutyser, E.; Su, Y.; Yu, Y.; Gouwy, M.; Zaja-Milatovic, S.; Van Damme, J.; Richmond, A. Hypoxia Enhances CXCR4 Expression in Human Microvascular Endothelial Cells and Human Melanoma Cells. Eur. Cytokine Netw. 2007, 18, 59–70. [Google Scholar] [CrossRef]
- Hattermann, K.; Held-Feindt, J.; Lucius, R.; Muerkoster, S.S.; Penfold, M.E.; Schall, T.J.; Mentlein, R. The Chemokine Receptor CXCR7 is Highly Expressed in Human Glioma Cells and Mediates Antiapoptotic Effects. Cancer Res. 2010, 70, 3299–3308. [Google Scholar] [CrossRef]
- Balabanian, K.; Lagane, B.; Infantino, S.; Chow, K.Y.; Harriague, J.; Moepps, B.; Arenzana-Seisdedos, F.; Thelen, M.; Bachelerie, F. The Chemokine SDF-1/CXCL12 Binds to and Signals through the Orphan Receptor RDC1 in T Lymphocytes. J. Biol. Chem. 2005, 280, 35760–35766. [Google Scholar] [CrossRef] [Green Version]
- Bauerle, K.T.; Schweppe, R.E.; Lund, G.; Kotnis, G.; Deep, G.; Agarwal, R.; Pozdeyev, N.; Wood, W.M.; Haugen, B.R. Nuclear Factor κB—Dependent Regulation of Angiogenesis, and Metastasis in an In Vivo Model of Thyroid Cancer Is Associated with Secreted Interleukin-8. J. Clin. Endocrinol. Metab. 2014, 99, E1436–E1444. [Google Scholar] [CrossRef]
- Kollmar, O.; Rupertus, K.; Scheuer, C.; Nickels, R.M.; Haberl, G.C.; Tilton, B.; Menger, M.D.; Schilling, M.K. CXCR4 and CXCR7 Regulate Angiogenesis and CT26. WT Tumor Growth Independent from SDF-1. Int. J. Cancer 2010, 126, 1302–1315. [Google Scholar] [CrossRef] [PubMed]
- Gerrits, H.; van Ingen Schenau, D.S.; Bakker, N.E.; van Disseldorp, A.J.; Strik, A.; Hermens, L.S.; Koenen, T.B.; Krajnc-Franken, M.A.; Gossen, J.A. Early Postnatal Lethality and Cardiovascular Defects in CXCR7-Deficient Mice. Genesis 2008, 46, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Perlin, J.R.; Talbot, W.S. Signals on the Move: Chemokine Receptors and Organogenesis in Zebrafish. Sci. STKE 2007, 2007, pe45. [Google Scholar] [CrossRef]
- Radenkovic, M.; Stojanovic, M.; Potpara, T.; Prostran, M. Therapeutic Approach in the Improvement of Endothelial Dysfunction: The Current State of the Art. Biomed Res. Int. 2013, 2013, 252158. [Google Scholar] [CrossRef] [PubMed]
- Deplanque, D.; Lavallee, P.C.; Labreuche, J.; Gongora-Rivera, F.; Jaramillo, A.; Brenner, D.; Abboud, H.; Klein, I.F.; Touboul, P.J.; Vicaut, E.; et al. Cerebral and Extracerebral Vasoreactivity in Symptomatic Lacunar Stroke Patients: A Case-Control Study. Int. J. Stroke 2013, 8, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, J.; Purkayastha, S.; Tang, Y.; Zhang, H.; Yin, Y.; Li, B.; Liu, G.; Cai, D. Hypothalamic Programming of Systemic Ageing Involving IKK-beta, NF-kappaB and GnRH. Nature 2013, 497, 211–216. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| hGAPDH | GACCACAGTCCATGCCATCA | GTCAAAGGTGGAGGAGTGGG |
| hCXCR4 | CACCGAGGCCCTAGCTTTCT | CACAGAGGTGAGTGCGTGCT |
| hSDF1 | CAGATGCCCATGCCGATT | AGTTTGGAGTGTTGAGAATT |
| hp53 | GAACAAGTTGGCCTGCACTG | GAAGTGGGCCCCTACCTAGA |
| hp21 | TATGGGGCTGGGAGTAGTTG | CGAGAGAAAACAGTCCAGGC |
| hp16 | AGTCCTCCTTCCTTGCCAAC | TCCGAGCACTTAGCGAATGT |
| hNBS1 | CCACCATTGTCCTAGCTACT | CTTGACTGGAACTCCCTTCT |
| hBRCA1 | GTGGTGCTTCTGTGGTGAAG | ACAGGTGCCTCACACATCTG |
| h53BP1 | AGGTTGGGTGTTCTTTGGCTT | TTGGTGTTGAGGCTTGTGGT |
| hChk1 | CTTTGGCTTGGCAACAGT | CCAGTCAGAATACTCCTG |
| hChk2 | GCGCCTGAAGTTCTTGTTTC | GCCTTTGGATCCACTACCAA |
| mGAPDH | TCAACGACCCCTTCATTGAC | ATGCAGGGATGATGTTCTGG |
| mCXCR4 | TCAGTGGCTGACCTCCTCTT | TTTCAGCCAGCAGTTTCCTT |
| mSDF1 | GGTTCTTCGAGAGCCACATC | GGGCAGCCTTTCTCTTCTTC |
| mp53 | CTCCGAAGACTGGATGACTGC | CAACAGATCGTCCATGCAGTG |
| mp21 | ACGGTGGAACTTTGACTTCGTC | CAGAGTGCAAGACAGCGACAAG |
| mCD31 | AGGCTTGCATAGAGCTCCAG | TTCTTGGTTTCCAGCTATGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heo, J.-I.; Kim, K.I.; Woo, S.-K.; Kim, J.S.; Choi, K.J.; Lee, H.-J.; Kim, K.S. Stromal Cell-Derived Factor 1 Protects Brain Vascular Endothelial Cells from Radiation-Induced Brain Damage. Cells 2019, 8, 1230. https://doi.org/10.3390/cells8101230
Heo J-I, Kim KI, Woo S-K, Kim JS, Choi KJ, Lee H-J, Kim KS. Stromal Cell-Derived Factor 1 Protects Brain Vascular Endothelial Cells from Radiation-Induced Brain Damage. Cells. 2019; 8(10):1230. https://doi.org/10.3390/cells8101230
Chicago/Turabian StyleHeo, Jong-Ik, Kwang Il Kim, Sang-Keun Woo, Joong Sun Kim, Kyu Jin Choi, Hae-June Lee, and Kwang Seok Kim. 2019. "Stromal Cell-Derived Factor 1 Protects Brain Vascular Endothelial Cells from Radiation-Induced Brain Damage" Cells 8, no. 10: 1230. https://doi.org/10.3390/cells8101230