Recent Research Advances in Mitosis during Mammalian Gametogenesis


1
Beijing Key Laboratory for Animal Genetic Improvement, National Engineering Laboratory for Animal Breeding, Key Laboratory of Animal Genetics and Breeding of the Ministry of Agriculture, College of Animal Science and Technology, China Agricultural University, Beijing 100193, China
2
CAS Key Laboratory of Genome Sciences and Information, Beijing Institute of Genomics, Chinese Academy of Sciences, Beijing 100101, China
3
State Key Laboratory of Stem Cell and Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cells 2019, 8(6), 567; https://doi.org/10.3390/cells8060567
Submission received: 22 April 2019
/
Revised: 2 June 2019
/
Accepted: 6 June 2019
/
Published: 10 June 2019
(This article belongs to the Special Issue Molecular Regulation of Mitosis and Its Role in Disease)
Abstract
:Mitosis is a highly sophisticated and well-regulated process during the development and differentiation of mammalian gametogenesis. The regulation of mitosis plays an essential role in keeping the formulation in oogenesis and gametogenesis. In the past few years, substantial research progress has been made by showing that cyclins/cyclin-dependent kinase (CDK) have roles in the regulation of meiosis. In addition, more functional signaling molecules have been discovered in mitosis. Growing evidence has also indicated that miRNAs influence cell cycling. In this review, we focus on specific genes, cyclins/Cdk, signaling pathways/molecules, and miRNAs to discuss the latest achievements in understanding their roles in mitosis during gametogenesis. Further elucidation of mitosis during gametogenesis may facilitate delineating all processes of mammalian reproduction and the development of disease treatments.
1. Introduction
Gametogenesis is an essential biological process to produce heritable haploid gametes in mammalian gonads, which includes oogenesis and spermatogenesis [1,2,3]. Between eight and nine days post-coitum (dpc), the majority of primordial germ cells (PGCs) are arrested at the G2 phase of the cell cycle [4]. In the germ cell proliferation stage, mouse PGCs enter into the genital ridge by migrating along the endoderm and mesentery of the hindgut, during which PGCs proliferate to form a certain number of germ cells via mitosis [5,6,7]. After arrival at the genital ridge, germ cells not only differentiate into spermatogonia or oocytes to enter gametogenesis but also form the syncytium in which multiple cells share one cytoplasm because of incomplete mitosis [8]. The syncytium contains a mass of cytoplasm that has many nuclei but no internal cell boundaries due to a series of incomplete cell division cycles. Before entering the genital ridge, there is no difference between XX (female) and XY (male) PGCs. However, starting at about 12.5 dpc, female and male PGCs begin to diverge dramatically [9]. In male mice before spermatogenesis, the mitosis of germ cells stops at the G0/G1 phase, which resume division when germ cells become spermatogonia after birth [8]. After mice are born, their germ cells begin to differentiate into spermatogonia via a series of mitotic cell divisions [10]. Conversely, in the female ovary, the mitosis of germ cells continues. Germ cells enter their first meiosis at 13.5 dpc and are then arrested in the diploid phase at 17.5 dpc [11]. During meiosis, homologous chromosomes align and pair with the synaptonemal complex formation to undergo homologous recombination. Then, oocytes and spermatocytes also generate haploid germ cells [12,13].
It is clear that mitosis is an essential process for PGC migration and auxocyte proliferation. Mitosis, a phase of the cell cycle, involves the following processes by which chromosomes migrate to the middle of the cell simultaneously and segregate into two daughter cells equally through the mitotic spindle before cytokinesis [14]. There are four phases in the cell cycle: G1, S, G2, and M. The G1 phase is alternatively called the pre-replicative phase, during which some mRNAs and proteins required for other phases are synthesized [15]. The S phase involves DNA replication, histone synthesis, and nucleosome replication [16,17,18]. In the G2 phase, active cell growth and protein synthesis ensures that cells enter mitosis. The M phase includes both mitosis and cytokinesis to separate the genome and ensures that the two daughter cells inherit an equal and identical complement of chromosomes [14,19]. It is exceedingly complex and highly regulated. Therefore, the M phase is usually divided into five phases: prophase, prometaphase, metaphase, anaphase, and telophase. Prophase, the first phase of mitosis in mammalian cells, includes the condensation of chromosomes, movement of centrosomes, formation of the mitotic spindle, and nucleoli breakdown. Afterwards, the nuclear envelope breaks down, and the chromosomes inside form protein structures named kinetochores during prometaphase [20]. Kinetochores are protein structures that form at the centromere during cell division and attach the chromosomes to the spindle fibers. In metaphase, chromosomes align along the metaphase plate and attach to microtubules anchored to centrosomes which duplicate in the S phase but separate in mitosis [21]. During anaphase, the separated sister chromosomes move from the center of the spindle in the metaphase plate toward opposite poles of the cell (anaphase A), and the mitotic spindle fibers elongate (anaphase B) [22]. Finally, as chromosomes reach the cell poles, a nuclear envelope is reassembled around each set of chromatids, nucleoli reappear, and chromosomes begin to decondense back into the expanded chromatin that is present during interphase.
2. Mitosis of Male Gametogenesis
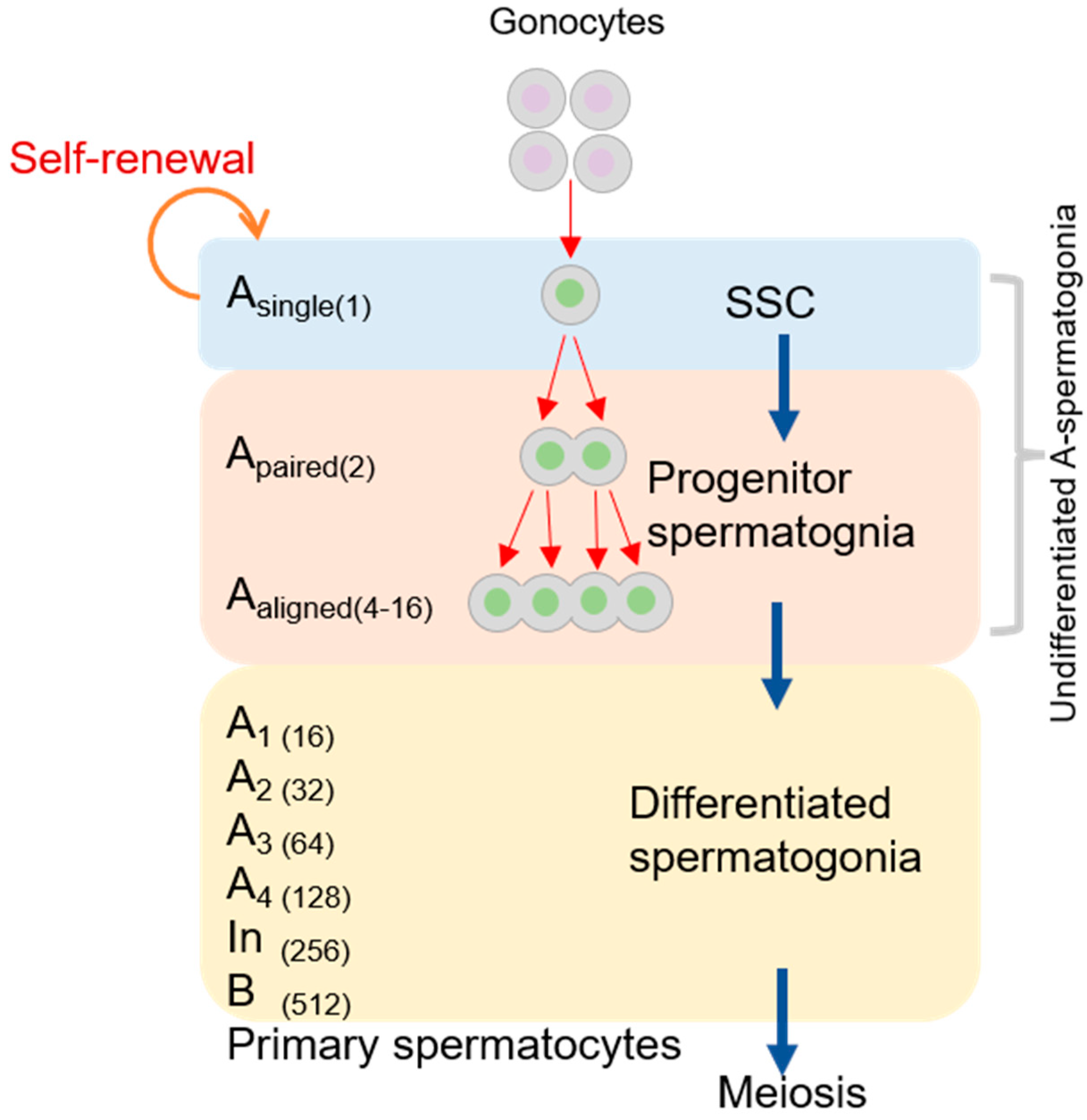
Spermatogenesis is the process through which diploid spermatogenic stem cells grow into haploid sperms in the male testis. There are some indispensable stages, including the mitosis of the spermatogonial stem cell (SSC), meiosis of the spermatocyte, and spermiogenesis. The mitotic division of SSCs is located adjacent to the basement membrane, which produces type A or B spermatogonia (see Figure 1) [23]. Type A spermatogonia replenish the stem cells, and type B spermatogonia develop into spermatocytes [24,25]. A single type A spermatogonia (Asingle or As) undergoes a self-renewing division to produce two new As cells. As spermatogonia, often considered SSCs, must undergo division to produce a pair of spermatogonial cells (Apaired or Apr). They then differentiate into 4–16 and even 32 spermatogonial cells (Aaligned or Aal) via a series of mitotic cell divisions [26,27]. Due to rapid and incomplete cytokinesis, Apr and Aal connect to one another using intercellular bridges (see Figure 2) [27,28]. Studies of the mouse male germline have shown that the cytoplasmic bridges may permit “cytoplasmic sharing” of essential signals for synchronous cell divisions and facilitate the sharing of gene products between post-meiotic haploid spermatids so that genetically distinct spermatids remain phenotypically diploid [29,30]. During the mitosis of type A1 spermatogonia, which differentiate from Aal cells, these germ cells located in the basement membrane migrate to the seminiferous tubules [31]. A1 spermatogonia subsequently undergo six mitoses each, forming A2, A3, A4, In, and B spermatogonia. Sertoli cells (SCs) also play a central role in spermatogenesis, providing structural support and nutrition to developing germ cells and producing proteins that influence the mitotic activity of spermatogonia [32,33]. The small kinetochore-associated protein (SKAP) is a component of the mitotic spindle, which is essential for faithful chromosome segregation during anaphase [34]. SKAP−/− mice grow normally without any obvious developmental defects. Therefore, SKAP is dispensable for somatic cell divisions in mice. However, SKAP affects mitosis in spermatogenesis because Skap−/− mice have smaller testes and a strong decrease in sperm production before meiosis compared with wildtype mice [35].
3. Mitosis of Female Gametogenesis
Oogenesis is the process of female gamete development which takes place in ovaries. It is complex and regulated by a vast number of intra- and extra-ovarian factors [36]. Oogonia, which are generated from PGCs, proliferate by mitosis and form primary oocytes. However, unlike spermatogenesis, oogonia are formed in large numbers from PGCs by mitosis during early fetal development, which then arrest at prophase stage of the first meiotic division around the time of birth [37,38].
4. Gene Regulation of Mitosis during Mammalian Gametogenesis
PGCs divide into eggs or spermatids and emerge as clusters of multiple cells that share one cytoplasm in early embryos [39,40]. Then, PGCs propagate rapidly and grow in number but stop propagation during the late pregnancy period in mammals [41]. In this period, female germ cells enter the meiotic prophase instantly, whereas male germ cells subsequently arrest in the G1 phase until puberty. The process of mitosis in gametes is regulated by several genes. Studies have demonstrated that the specific deletion of Mastl in mouse PGCs leads to the failure of cells to proceed beyond the metaphase-like stage of mitosis. This mitotic defect results in the activation of the DNA damage response pathway. Thus, the majority of Mastl−/− PGCs undergo apoptosis [42]. Pin1, a peptidyl-prolyl isomerase, is involved in the regulation of mammalian PGC proliferation. PGCs have a prolonged cell cycle in the absence of Pin1, leading to fewer cell divisions and strikingly fewer Pin1−/− PGCs by the end of the proliferative phase. Therefore, male and female Pin1−/− mice have profound fertility defects [43]. The PTEN gene can inhibit cell proliferation via restraint of the PI3K/AKT pathway, as revealed by Pten−/− PGCs both in vivo and in vitro. These cells show significantly increased mitosis [44,45,46]. Similarly, Kit (Kit oncogene) and Kitl (Kit ligand or stem cell factor) are also required for the migration and proliferation of PGCs throughout embryogenesis [47,48].
Some DNA repair-associated genes are also involved in the proliferation of PGCs. Rev7 and Mcm9 are related to cell cycle regulation and homologous recombination repair by recruiting RAD51 to sites of DNA damage in mammals [49,50,51]. Germ cell depletion is the result of reduced PGC numbers both before and after they arrive in the primitive gonads of Mcn9 mutant mice [52]. Rev7−/− mice display the loss of germ cells by apoptotic cell death during migration and germ cell aplasia in both testes and ovaries after birth [53]. According to these studies, DNA repair-associated genes, such as Rev, also regulate gamete mitosis.
In many mammalian species, the Nanos2 gene encoding RNA-binding proteins was identified as functional in controlling the proliferation of PGCs and maintaining the stemness of undifferentiating SSCs [54]. In male nanos2/nanos3-null mice, the size and weight of their testes are reduced and no gamete cells can be detected compared to nanos2/nanos3+/+ mice [55,56]. These findings suggest that Nanos genes are involved in the maintenance of mitosis in gametes by supporting their proliferation and/or suppressing apoptosis. The Sox4 gene is expressed in gonadal supporting cells, the organizing center of gonad organogenesis. However, Nanos2 in male Sox4-null mice is expressed at levels substantially lower than in wildtype testes, indicating that the mitosis of male germ cells is severely impaired in Sox4–/– testes [57]. DND1 expressed in mouse germ cells regulates the mitotic arrest of male germ cells through the translational regulation of cell cycle-related genes [58].
4.1. Cyclin/CDK Regulation of Mitosis during Mammalian Gametogenesis
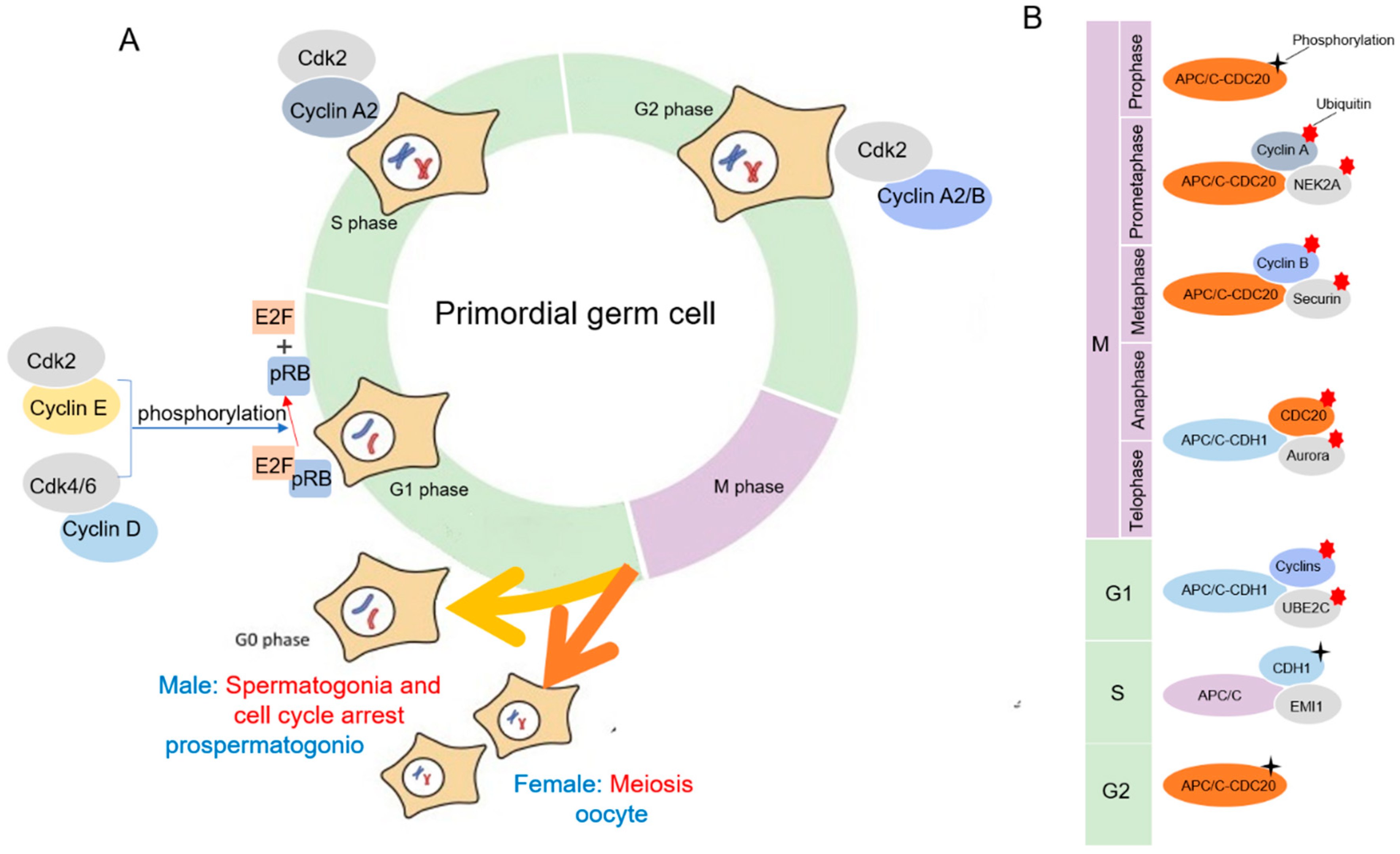
Cdks are serine/threonine kinases, and their catalytic activities are modulated by interactions with cyclins and Cdk inhibitors. They have roles in regulating the cell cycle, transcription, and mRNA processing [59]. In mammalian somatic cells, there are at least four different Cdks to regulate the interphase of mitosis: Cdk2, Cdk3, Cdk4, and Cdk6. After interphase, Cdk1 drive cells through mitosis (Figure 2A) [60,61]. To perform kinase activity, CDK must bind to a cyclin as a regulatory protein. Only the cyclin–CDK complex is an active kinase. Using a cell cycle array revealed that PGCs were comparable with somatic cells in the G1 phase, and it included expression of cyclin D3 (ccnd3), the retinoblastoma protein (pRB) family, and CDK inhibitors [62]. According to the mammalian PGC cycle, cyclin E and cyclin D were found to be the predominant cyclins. Cyclin D activates Cdk4/6, and the pRB family is phosphorylated in the early G1 phase. On the contrary, the reason why prospermatogonia undergo mitotic arrest at the G1/S phase checkpoint is that cyclin E1/2 and cyclin B1/2 are downregulated and cyclinB3 is upregulated, which result in the hypophosphorylation of pRB. Then, E2F transcription factors are released, leading to the activation and transcription of E2F-responsive genes for cell cycle progression [63]. During the late G1 phase, Cdk2 is stimulated via binding to cyclin E [64]. A study shows that the presence of lower levels of cyclin E1/E2 and cyclin B1/B2 in the 13.5–14.5 dpc cell cycle arrested prospermatogonia, as compared with mitotic 11.5–12.5 dpc PGCs [62]. Subsequently, prospermatogonia pass the restriction point of the G1/S phase and enter the S phase. Similar to E-type cyclin binding, cyclin A synthesized at the onset of S phase also activates Cdk2 that phosphorylates proteins involved in DNA replication [65]. At the transition phase of G2/M in germinal cells, cyclin A2 binds to Cdk1, which is required for the initiation of prophase. Finally, the cyclin B complex with Cdk1 drives entry into the M phase [66,67].
Ccna2, a cyclin A gene, is expressed in the spermatogonia of the adult mouse testis, which may have an S phase function in the mitotic cell cycle of spermatogonial germ cells. During embryogenesis, D-type cyclin expression is primarily restricted to CCND3 in male germ cells [68]. Cyclins D1 and D3 are expressed in spermatogonia at all cell cycle phases of the seminiferous tubule epithelium in the adult testis. However, cyclin D2 is detected at the stage when type Aal spermatogonia differentiate into type A1 spermatogonia, indicating that cyclin D2 is involved in SSC proliferation [69]. The Cdkn1b-encoded protein binds to and prevents the activation of cyclin E–CDK2 and cyclin D–CDK4 complexes. Cdkn1b−/− SSCs show suppressed proliferation and diminished expression of CDK4 and pRB1, resulting in the poor phosphorylation of pRB1. In cultured SSCs, pRB1 deficiency leads to cell cycle arrest and apoptosis [70]. In mitosis-arrested male germ cells, p27Kip1 and p15INK4b encoded by Cdkn1b and Cdkn2b genes are upregulated, which inhibit CyclinE–cdk2 and CyclinD–cdk4/6, respectively. P27Kip1 and p15INK4b ensure that hypophosphorylated pRB can inhibit the G1/S phase transition and suppress cyclin E expression [63]. pRB1 is required for germ cell entry into G1/0 arrest in the normal gonad. However, in pRB−/− mice, upregulation of other cell cycle suppressors, including Cdkn1b and Cdkn2b, can induce delayed germ cell arrest [71].
4.2. APC/C Regulation of Mitosis during Mammalian Gametogenesis
The anaphase-promoting complex or cyclosome (APC/C), the E3 ubiquitin ligase, plays an important role in the regulation of the mitotic cell cycle [72]. In the mitotic cell, the spindle assembly checkpoint (SAC) ensures that each daughter cell inherits an identical set of chromosomes. With the activated APC/C, it coordinates the accurate attachment of sister chromatid kinetochores to the spindle [73].
During mitosis in mammalian gametogenesis, the germ cell cycle is controlled by the oscillation in activity of CDKs. Like the somatic cells, this precisely regulated degradation process is accomplished by APC/C catalyzed ubiquitination in germ cells [74]. Cell division cycle 20 (CDC20) and CDC20 homologue 1 (CDH1) are the activating subunits of APC/C (Figure 2B) [74]. The APC/C is inactive from the late G1 phase to early prophase to ensure its main substrates accumulate. The APC/C–CDH1 complex mainly regulates the process of anaphase and the early G1 phase. At the G1/S transition, APC/C–CDH1 is inactivated by a combination of binding to the APC/C inhibitor early mitotic inhibitor 1 (EMI1), degradation of ubiquitin-conjugating enzyme E2C (UBE2C), and CDH1 phosphorylation. APC/C–CDC20 complexes are primarily regulated by the degradation of related substrates in the prometaphase and metaphase phases [75]. In the G2 phase, CDC20 is phosphorylated by CDK1, partially activating the phase using APC/C interaction. In the prometaphase, the activity of the APC/C–CDC20 complex is inhibited during the G2/M phase. The APC/C–CDC20 complex ubiquitinates cell cyclin A and NIMA-related expressed kinase 2A (NEK2A). Cyclin B1–CDK1 complexes and securin keep the cell in the M phase. Thus, in the metaphase, the APC/C–CDC20 complex degrades cyclin B1 and CDK1 to end cell division. In anaphase, CDH1 is dephosphorylated and the APC/C–CDH1 complex is activated. The APC/C–CDH1 complex ubiquitinates CDC20, Aurora A/B, and other kinases to promote the end of mitosis [72].
5. Signaling Pathways/Molecules Regulating Mitosis during Mammalian Gametogenesis
SSCs are located in the basal membrane of the convoluted spermatogonia, which can not only undergo self-renewal to maintain a stable SSC number but also generate spermatocytes by directional differentiation [76]. The processes of SSC self-renewal and differentiation are also regulated by some signaling pathways or signaling molecules [77].
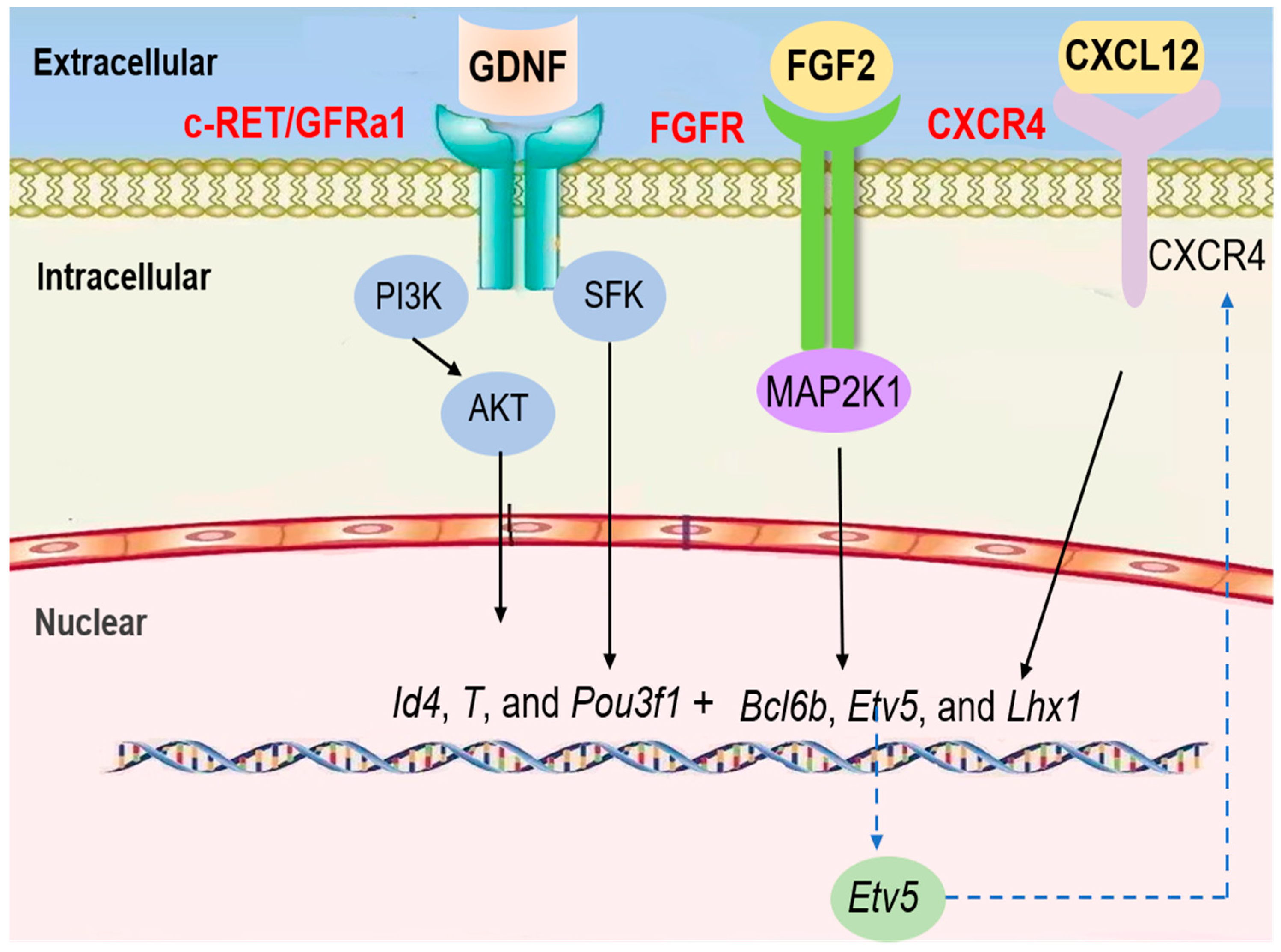
The steady state of mitosis in SSCs is driven by a complex paracrine and endocrine system within structurally well-organized tissue. Central to this system are SCs that provide nutritional and structural support for the self-renewal and differentiation of germ cells (Figure 3) [78,79]. GDNF, a paracrine factor, is secreted by SCs. It promotes SSC self-renewal and maintenance via mitosis. In mice, GDNF plays a role in promoting spermatogonial self-renewal using RET tyrosine kinase and a ligand-specific co-receptor, GFRα1, which is expressed on undifferentiated type A spermatogonia [80,81,82]. Because GFRαA1 and RET are concomitantly expressed in Apaired spermatogonia and some Aaligned spermatogonia, GDNF induces and maintains the proliferation of these cells as SSCs begin differentiation [83]. According to single cell RNA-seq data from mice and humans, GFRα1 is mostly expressed in undifferentiated spermatogonia and co-expressed with Pou3f1, Bcl6b, and Etv5 [84,85]. Etv5 is a transcription factor secreted by SCs which can promote the RET synthesis of GDNF receptor and ensure that the GDNF signal pathway is transmitted from the extracellular to the intracellular space [86]. Pou3f1 is a member of the transcriptor OCT combined with the family, which also has an important modulatory function for SSC proliferation [87]. Bcl6b, a member of the poxvirus and zinc finger (POZ) family of transcriptional repressors, is another target of GDNF that was recently identified in SSCs by microarray analysis [88]. When GDNF is applied to a germline stem cell culture, Akt is phosphorylated rapidly, and the addition of a chemical inhibitor of PI3K prevents GS cell self-renewal, indicating that the PI3K/Akt pathway is essential for the self-renewal of spermatogonial stem cells [89]. GDNF binds to GFRα1 on the SSC membranes to form the GDNF–GFRα1 complex, which binds and activates RET. The MAPK, SFK, and PI3K/AKT signaling pathways will be further activated [90]. In SSCs, the phosphorylation of AKT is mainly activated by PI3K, which results in AKT self-activation. The activation of AKT can switch up the expression of transcription factors, such as Etv5, Pou3f1, and Bcl6b, to promote SSC self-renewal [91].
Similarly, the SFK (SRC family kinase) pathway also mediates GDNF functions in SSCs in vitro [92]. Experiments using SSCs treated with GDNF have shown that GDNF increases the number of SSCs and that downstream targets of the GDNF/RET signaling pathway, such as ID4, BCL6, ETV5, and LHX1, are critical for SSC self-renewal [93,94]. GFRα-1 and Ret are expressed in SSCs, and the Src family co-precipitates with Ret after GDNF stimulation. SFK promotes SSC proliferation through Ret activation. Src and Yes play an especially predominant role in the immediate response of primary SSCs to GDNF. Further, Src activates a PI3K/Akt signaling pathway and switches up the expression of transcription factors, such as Etv5, Pou3f1, and Bcl6b [95].
FGF is also a bona fide mitosis factor for the self-renewal of spermatogonial stem cells. In vitro studies have shown that FGF2 promotes mitogenic effects, and the differentiation of SSCs can be blocked by the addition of FGF2 to culture medium, although it’s in vivo effects are unclear [96,97]. FGF2 relies on MAP2K1 activation to drive SSC self-renewal via upregulation of Etv5, Bcl6b, and Lhx1 genes, according to a mouse germline stem cell culture system that allows the in vitro expansion of SSCs [98]. Similar to FGF2, FGF5 promotes the proliferation of cultured GFRα1+ spermatogonia and mouse SSC line C18-4 in a time- and dose-dependent manner via ERK and AKT activation [99,100]. FGF5 also upregulates genes associated with self-renewal, such as Etv5, Id4, and Shisa6 [101,102,103]. CXCL12, a chemokine protein, specifically binds to the CXCR4 receptor, which is expressed in Sertoli cells to regulate SSC self-renewal and maintenance. The signaling response to CXCL12–CXCR4 activation is involved in the prevention of the transition to a progenitor state, the regulation of SSC proliferation, and the guidance of SSC homing to cognate niches [104].
Taken together, GDNF and FGF2 signaling work in concert to activate PI3K/AKT, SFK, and MAP2K1, which are critical regulators to switch up the expression of transcription factors. Furthermore, the transcription factor Etv5 regulates CXCR4 expression in SSCs, thereby controlling the signaling response of CXCL12 to influence self-renewal, proliferation, and homing [105].
6. mRNA Regulation in Mitosis during Mammalian Gametogenesis
During the mitosis of mammalian gametes, microRNAs (miRNAs), small non-coding RNAs 18–23 nt in length, are viewed as active regulators in the post-transcriptional regulatory processes of germ cells [106,107]. Dicer, as an RNase III endonuclease, plays a critical role in the biogenesis of miRNAs [108]. In the Dicer-deleted testis, PGCs and spermatogonia exhibit poor proliferation [109]. Here, we discuss some specific roles of a few miRNA molecules in mitosis during gametogenesis (Table 1). miR-17-2 is highly expressed at the stage of PGC development, which regulates the mitosis of PGCs. Moreover, miRNA-17-5p, -18, -19a, and -19b, which are only expressed in females, regulate PGC exit from mitotic proliferation [110]. MiR-19a and -19b may regulate PTEN dosage, which negatively controls PGC proliferation [111]. In a recent study, miRNA-31-5p mimics decreased the level of cyclin A2 rather than cyclin D1 or cyclin E1, which regulates the proliferation and DNA synthesis of human SSCs via the PAK1-JAZF1-cyclin A2 pathway [112]. The miR-290-295 cluster is only present in placental mammals. It consists of seven miRNA precursors: miR-290, miR-291a, miR-292, miR-291b, miR-293, miR-294, and miR-295. The miR-290-295 cluster affects the cell cycle of PGCs at multiple points. Under certain conditions, it might assist G1/S progression and regulate the G2–M transition of PGCs and ES cells [110,113]. MiR-302 family members were specifically expressed in PGCs, and the validated target gene is the cyclin-dependent kinase inhibitor 1A (Cdkn1a). MiR-302 downregulated Cdkn1a to ensure that PGCs enter the G1/S transition of mitosis [114]. MiR-202 family members, including miR-202-3p and miR-202-5p, are highly expressed in mouse spermatogonial stem cells (SSCs) and are oppositely regulated by GDNF, a key factor for SSC self-renewal. By using CRISPR/Cas9-mediated knockout of miR-202 in cultured SSCs, a study found that miR-202−/− SSCs initiate premature differentiation, accompanied by reduced stem cell activity and increased mitosis [115]. Dmrt1 determines whether male germ cells undergo mitosis and spermatogonial differentiation or meiosis by controlling cyclical gene expression in Sertoli cells [116]. MiR-224 targets the DMRT1 3′-UTR, and the overexpression of miR-224 influences SSC differentiation by suppressing DMRT1 expression [117].
7. Concluding Remarks
We have systematically summarized the regulation of genes, cyclins/CDK, signaling molecules, and miRNAs in mitosis during gametogenesis. PGCs undergo rapid mitosis during migration to the reproductive ridge, and spermatogonial stem cells undergo mitosis during self-renewal and differentiation [118]. In recent years, with the deepening of research and technological progress, we are gradually expanding our knowledge of mitosis during gametogenesis, although specific mitosis mechanisms of gametes are still unclear. For example, it remains to be revealed how different intracellular pathways activated by external signals are integrated and functionally associated with cell cycle cyclins, CDK controllers, and miRNAs in mitosis of gametes [64]. In addition, it is unknown how male, but not female, mouse PGCs enter mitotic arrest at 13.5–14.5 dpc. To further explore these aspects, in vitro reconstitution of mouse germ cell development from mPSCs may provide a robust foundation to understand the mitosis mechanism of germ cells, including signaling pathways, transcriptional networks, and epigenetic regulation. In the near future, omics analysis and structural biology may also play indispensable roles in resolving basic issues in gamete mitosis. Many molecules involved in the regulation of mitosis during gametogenesis have been identified; however, further investigation will be essential to elucidate the signaling pathways assigned for maintenance of undifferentiated state, self-renewal, and differentiation [82]. In the future, the functions of putative mitosis during gametogenesis molecules and signaling pathways need to be verified via a functional transplantation assay.
Author Contributions
S.-L.D., K.Y., J.-H.W., Y.-X.L., Z.-X.L., and Y.L. wrote the paper.
Funding
This work was supported by grants from the National Transgenic Creature Breeding Grand Project (2016zx08008-003).
Acknowledgments
We thank Martyn Rittman, English Editing Manager ([email protected]), for editing the English text of a draft of this manuscript. We apologize to some authors for not citing their interesting work. Our choice was not intended to be exclusive.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| APC/C | anaphase-promoting complex or cyclosome |
| Akt | thymoma viral proto-oncogene 1 |
| ATF | activating transcription factor |
| Bcl6b | B-cell CLL/lymphoma 6, member B |
| Ccnd | cyclin D |
| CDC20 | cell division cycle 20 |
| CDH1 | CDC20 homologue 1 |
| Cdk | cyclin-dependent kinase |
| CXCL12 | chemokine (C-X-C motif) ligand 12 |
| CXCR4 | C-X-C chemokine receptor type 4 |
| EMI1 | early mitotic inhibitor 1 |
| Etv5 | ets variant gene 5 |
| Fgf2 | fibroblast growth factor 2 |
| Gdnf | glial cell line-derived neurotrophic factor |
| Gfrα-1 | glial cell line-derived neurotrophic factor family receptor alpha 1 |
| Kitl | KIT ligand or stem cell factor |
| NEK2A | NIMA-related expressed kinase 2A |
| PI3K | phosphatidylinositol 3-kinase |
| POZ | poxvirus and zinc finger |
| pRB | retinoblastoma protein |
| SCs | Sertoli cells |
| SFK | src family kinase |
| SKAP | Small kinetochore-associated protein |
| SSC | spermatogonial stem cell |
| Taf4B | TATA box-binding protein (TBP)-associated factor, subunit 4B |
| UBE2C | ubiquitin-conjugating enzyme E2C |
References
- Carreau, S.; Hess, R.A. Oestrogens and spermatogenesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1517–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesch, B.J.; Page, D.C. Genetics of germ cell development. Nat. Rev. Genet. 2012, 13, 781–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, Z.; Szczepny, A.; McLaughlin, E.A.; Meistrich, M.L.; Zhou, W.; Ustunel, I.; Loveland, K.L. Dynamic Hedgehog signalling pathway activity in germline stem cells. Andrology 2014, 2, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, Y.; Yamaji, M.; Yabuta, Y.; Sano, M.; Shigeta, M.; Matsui, Y.; Saga, Y.; Tachibana, M.; Shinkai, Y.; Saitou, M. Cellular dynamics associated with the genome-wide epigenetic reprogramming in migrating primordial germ cells in mice. Development 2007, 134, 2627–2638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Shirayoshi, Y.; Koshimizu, U.; Hashimoto, S.; Yonehara, S.; Eguchi, Y.; Tsujimoto, Y.; Nakatsuji, N. Gene transfection of mouse primordial germ cells in vitro and analysis of their survival and growth control. Exp. Cell Res. 1997, 230, 76–83. [Google Scholar] [CrossRef]
- Oatley, J.A.; Brinster, R.L. Regulation of Spermatogonial Stem Cell Self-Renewal in Mammals. Annu. Rev. Cell Dev. Bi. 2008, 24, 263–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, E.; Tumbar, T.; Guasch, G. Socializing with the neighbors: Stem cells and their niche. Cell 2004, 116, 769–778. [Google Scholar] [CrossRef]
- Cooke, H.J.; Saunders, P.T. Mouse models of male infertility. Nat. Rev. Genet. 2002, 3, 790–801. [Google Scholar] [CrossRef]
- Edson, M.A.; Nagaraja, A.K.; Matzuk, M.M. The mammalian ovary from genesis to revelation. Endocr. Rev. 2009, 30, 624–712. [Google Scholar] [CrossRef]
- De Rooij, D.G. Proliferation and differentiation of spermatogonial stem cells. Reproduction 2001, 121, 347–354. [Google Scholar] [CrossRef]
- Pepling, M.E. From primordial germ cell to primordial follicle: Mammalian female germ cell development. Genesis 2006, 44, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Hunter, N. Meiotic Recombination: The Essence of Heredity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudat, F.; Imai, Y.; de Massy, B. Meiotic recombination in mammals: Localization and regulation. Nat. Rev. Genet. 2013, 14, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, M. The Role of Model Organisms in the History of Mitosis Research. Csh. Perspect. Biol. 2014, 6, a15768. [Google Scholar] [CrossRef] [PubMed]
- Vallier, L. Cell Cycle Rules Pluripotency. Cell Stem Cell 2015, 17, 131–132. [Google Scholar] [CrossRef] [Green Version]
- Takeda, D.Y.; Dutta, A. DNA replication and progression through S phase. Oncogene 2005, 24, 2827–2843. [Google Scholar] [CrossRef] [Green Version]
- DeRan, M.; Pulvino, M.; Greene, E.; Su, C.; Zhao, J. Transcriptional activation of histone genes requires NPAT-dependent recruitment of TRRAP-Tip60 complex to histone promoters during the G1/S phase transition. Mol. Cell. Biol. 2008, 28, 435–447. [Google Scholar] [CrossRef]
- Ramachandran, S.; Henikoff, S. Replicating Nucleosomes. Sci. Adv. 2015, 1, e1500587. [Google Scholar] [CrossRef]
- De Souza, C.P.; Osmani, S.A. Mitosis, not just open or closed. Eukaryot. Cell 2007, 6, 1521–1527. [Google Scholar] [CrossRef]
- Guttinger, S.; Laurell, E.; Kutay, U. Orchestrating nuclear envelope disassembly and reassembly during mitosis. Nat. Rev. Mol. Cell Biol. 2009, 10, 178–191. [Google Scholar] [CrossRef]
- Jongsma, M.L.; Berlin, I.; Neefjes, J. On the move: Organelle dynamics during mitosis. Trends Cell Biol. 2015, 25, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.J.; Wordeman, L. Mitosis, microtubule dynamics and the evolution of kinesins. Exp. Cell Res. 2015, 334, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Hanukoglu, A.; Hanukoglu, I. Localization of epithelial sodium channel (ENaC) and CFTR in the germinal epithelium of the testis, Sertoli cells, and spermatozoa. J. Mol. Histol. 2018, 49, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Rato, L.; Alves, M.G.; Socorro, S.; Duarte, A.I.; Cavaco, J.E.; Oliveira, P.F. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Zarzycka, M.; Mruk, D.D. Biology of the Sertoli Cell in the Fetal, Pubertal, and Adult Mammalian Testis. Results Probl Cell Differ. 2016, 58, 225–251. [Google Scholar] [PubMed]
- De Rooij, D.G.; Russell, L.D. All you wanted to know about spermatogonia but were afraid to ask. J. Androl. 2000, 21, 776–798. [Google Scholar] [PubMed]
- Licatalosi, D.D. Roles of RNA-binding Proteins and Post-transcriptional Regulation in Driving Male Germ Cell Development in the Mouse. Adv. Exp. Med. Biol. 2016, 907, 123–151. [Google Scholar] [PubMed]
- Griswold, M.D. Spermatogenesis: The Commitment to Meiosis. Physiol. Rev. 2016, 96, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.P.; Russell, L.D. Clonal development of interconnected germ cells in the rat and its relationship to the segmental and subsegmental organization of spermatogenesis. Am. J. Anat. 1991, 192, 121–128. [Google Scholar] [CrossRef]
- Haglund, K.; Nezis, I.P.; Stenmark, H. Structure and functions of stable intercellular bridges formed by incomplete cytokinesis during development. Commun. Integr. Biol. 2011, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, H.; L’Hernault, S.W. Spermatogenesis. Curr. Biol. 2017, 27, R988–R994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Thompson, L.A.; Dufour, J.M. Sertoli cells—Immunological sentinels of spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Thompson, D.J.; Varner, D.D. Role of Sertoli cell number and function on regulation of spermatogenesis. Anim. Reprod. Sci. 2008, 105, 23–51. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.C.; Kiyomitsu, T.; Hori, T.; Backer, C.B.; Fukagawa, T.; Cheeseman, I.M. Aurora B kinase controls the targeting of the Astrin-SKAP complex to bioriented kinetochores. J. Cell Biol. 2010, 191, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Grey, C.; Espeut, J.; Ametsitsi, R.; Kumar, R.; Luksza, M.; Brun, C.; Verlhac, M.H.; Suja, J.A.; de Massy, B. SKAP, an outer kinetochore protein, is required for mouse germ cell development. Reproduction 2016, 151, 239–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porras-Gomez, T.J.; Moreno-Mendoza, N. Neo-oogenesis in mammals. Zygote 2017, 25, 404–422. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, F.; Smitz, J. Molecular control of oogenesis. Biochim. Biophys. Acta 2012, 1822, 1896–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Liu, H.; Gu, X.; Boots, C.; Moley, K.H.; Wang, Q. Metabolic control of oocyte development: Linking maternal nutrition and reproductive outcomes. Cell. Mol. Life Sci. 2015, 72, 251–271. [Google Scholar] [CrossRef] [PubMed]
- McLaren, A. Primordial germ cells in the mouse. Dev. Biol. 2003, 262, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kurimoto, K.; Saitou, M. Mechanism and Reconstitution In Vitro of Germ Cell Development in Mammals. Cold Spring Harb. Symp. Quant. Biol. 2015, 80, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Western, P. Foetal germ cells: Striking the balance between pluripotency and differentiation. Int. J. Dev. Biol. 2009, 53, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Risal, S.; Zhang, J.; Adhikari, D.; Liu, X.; Shao, J.; Hu, M.; Busayavalasa, K.; Tu, Z.; Chen, Z.; Kaldis, P.; et al. MASTL is essential for anaphase entry of proliferating primordial germ cells and establishment of female germ cells in mice. Cell Discov. 2017, 3, 16052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atchison, F.W.; Capel, B.; Means, A.R. Pin1 regulates the timing of mammalian primordial germ cell proliferation. Development 2003, 130, 3579–3586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaswamy, S.; Nakamura, N.; Vazquez, F.; Batt, D.B.; Perera, S.; Roberts, T.M.; Sellers, W.R. Regulation of G1 progression by the PTEN tumor suppressor protein is linked to inhibition of the phosphatidylinositol 3-kinase/Akt pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 2110–2115. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lesche, R.; Li, D.M.; Liliental, J.; Zhang, H.; Gao, J.; Gavrilova, N.; Mueller, B.; Liu, X.; Wu, H. PTEN modulates cell cycle progression and cell survival by regulating phosphatidylinositol 3,4,5,-trisphosphate and Akt/protein kinase B signaling pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6199–6204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neirijnck, Y.; Kuhne, F.; Mayere, C.; Pavlova, E.; Sararols, P.; Foti, M.; Atanassova, N.; Nef, S. Tumor Suppressor PTEN Regulates Negatively Sertoli Cell Proliferation, Testis Size, and Sperm Production In Vivo. Endocrinology 2019, 160, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Tohda, A.; Nishimune, Y. Proliferation and differentiation of spermatogonial stem cells in the w/wv mutant mouse testis. Biol. Reprod. 2003, 69, 1815–1821. [Google Scholar] [CrossRef]
- Driancourt, M.A.; Reynaud, K.; Cortvrindt, R.; Smitz, J. Roles of KIT and KIT LIGAND in ovarian function. Rev. Reprod. 2000, 5, 143–152. [Google Scholar] [CrossRef]
- Lutzmann, M.; Grey, C.; Traver, S.; Ganier, O.; Maya-Mendoza, A.; Ranisavljevic, N.; Bernex, F.; Nishiyama, A.; Montel, N.; Gavois, E.; et al. MCM8- and MCM9-deficient mice reveal gametogenesis defects and genome instability due to impaired homologous recombination. Mol. Cell 2012, 47, 523–534. [Google Scholar] [CrossRef]
- Park, J.; Long, D.T.; Lee, K.Y.; Abbas, T.; Shibata, E.; Negishi, M.; Luo, Y.; Schimenti, J.C.; Gambus, A.; Walter, J.C.; et al. The MCM8-MCM9 complex promotes RAD51 recruitment at DNA damage sites to facilitate homologous recombination. Mol. Cell. Biol. 2013, 33, 1632–1644. [Google Scholar] [CrossRef]
- Murakumo, Y.; Roth, T.; Ishii, H.; Rasio, D.; Numata, S.; Croce, C.M.; Fishel, R. A human REV7 homolog that interacts with the polymerase zeta catalytic subunit hREV3 and the spindle assembly checkpoint protein hMAD2. J. Biol. Chem. 2000, 275, 4391–4397. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Schimenti, J.C. MCM9 deficiency delays primordial germ cell proliferation independent of the ATM pathway. Genesis 2015, 53, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Mii, S.; Asai, N.; Asai, M.; Niimi, K.; Ushida, K.; Kato, T.; Enomoto, A.; Ishii, H.; Takahashi, M.; et al. The REV7 subunit of DNA polymerase zeta is essential for primordial germ cell maintenance in the mouse. J. Biol. Chem. 2013, 288, 10459–10471. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Tripathi, A.; Dubey, P.K. Expression and intracellular localization of Nanos2-homologue protein in primordial germ cells and spermatogonial stem cells. Zygote 2019, 27, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Sasaoka, Y.; Kiso, M.; Abe, K.; Haraguchi, S.; Kobayashi, S.; Saga, Y. Conserved role of nanos proteins in germ cell development. Science 2003, 301, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Saga, Y. Nanos2 suppresses meiosis and promotes male germ cell differentiation. Genes Dev. 2008, 22, 430–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Arsenault, M.; Ng, E.T.; Longmuss, E.; Chau, T.C.; Hartwig, S.; Koopman, P. SOX4 regulates gonad morphogenesis and promotes male germ cell differentiation in mice. Dev. Biol. 2017, 423, 46–56. [Google Scholar] [CrossRef]
- Cook, M.S.; Munger, S.C.; Nadeau, J.H.; Capel, B. Regulation of male germ cell cycle arrest and differentiation by DND1 is modulated by genetic background. Development 2011, 138, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Kaldis, P. Cdks, cyclins and CKIs: Roles beyond cell cycle regulation. Development 2013, 140, 3079–3093. [Google Scholar] [CrossRef]
- Malumbres, M.; Barbacid, M. Mammalian cyclin-dependent kinases. Trends Biochem. Sci. 2005, 30, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Sisakhtnezhad, S.; Heshmati, P. Comparative analysis of single-cell RNA sequencing data from mouse spermatogonial and mesenchymal stem cells to identify differentially expressed genes and transcriptional regulators of germline cells. J. Cell. Physiol. 2018, 233, 5231–5242. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, E.; Nazzicone, V.; Farini, D.; Campagnolo, L.; De Felici, M. Comparative transcript profiles of cell cycle-related genes in mouse primordial germ cells, embryonic stem cells and embryonic germ cells. Gene Expr. Patterns 2007, 7, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Western, P.S.; Miles, D.C.; van den Bergen, J.A.; Burton, M.; Sinclair, A.H. Dynamic regulation of mitotic arrest in fetal male germ cells. Stem Cells 2008, 26, 339–347. [Google Scholar] [CrossRef] [PubMed]
- De Felici, M.; Farini, D. The control of cell cycle in mouse primordial germ cells: Old and new players. Curr. Pharm. Des. 2012, 18, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Ravnik, S.E.; Wolgemuth, D.J. The developmentally restricted pattern of expression in the male germ line of a murine cyclin A, cyclin A2, suggests roles in both mitotic and meiotic cell cycles. Dev. Biol. 1996, 173, 69–78. [Google Scholar] [CrossRef]
- Satyanarayana, A.; Kaldis, P. Mammalian cell-cycle regulation: Several Cdks, numerous cyclins and diverse compensatory mechanisms. Oncogene 2009, 28, 2925–2939. [Google Scholar] [CrossRef]
- Wolgemuth, D.J.; Manterola, M.; Vasileva, A. Role of cyclins in controlling progression of mammalian spermatogenesis. Int. J. Dev. Biol. 2013, 57, 159–168. [Google Scholar] [CrossRef]
- Lanza, D.G.; Dawson, E.P.; Rao, P.; Heaney, J.D. Misexpression of cyclin D1 in embryonic germ cells promotes testicular teratoma initiation. Cell Cycle 2016, 15, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Beumer, T.L.; Roepers-Gajadien, H.L.; Gademan, I.S.; Kal, H.B.; de Rooij, D.G. Involvement of the D-type cyclins in germ cell proliferation and differentiation in the mouse. Biol. Reprod. 2000, 63, 1893–1898. [Google Scholar] [CrossRef]
- Tanaka, T.; Kanatsu-Shinohara, M.; Shinohara, T. The CDKN1B-RB1-E2F1 pathway protects mouse spermatogonial stem cells from genomic damage. J. Reprod. Dev. 2015, 61, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Spiller, C.M.; Wilhelm, D.; Koopman, P. Retinoblastoma 1 protein modulates XY germ cell entry into G1/G0 arrest during fetal development in mice. Biol. Reprod. 2010, 82, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, S.; Gorbsky, G.J. Spatiotemporal regulation of the anaphase-promoting complex in mitosis. Nat. Rev. Mol. Cell Biol. 2015, 16, 82–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfieri, C.; Chang, L.; Zhang, Z.; Yang, J.; Maslen, S.; Skehel, M.; Barford, D. Molecular basis of APC/C regulation by the spindle assembly checkpoint. Nature 2016, 536, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeganathan, K.B.; van Deursen, J.M. Differential mitotic checkpoint protein requirements in somatic and germ cells. Biochem. Soc. Trans. 2006, 34, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Hsiao, J.Y.; Davey, N.E.; Van Voorhis, V.A.; Foster, S.A.; Tang, C.; Morgan, D.O. Multiple mechanisms determine the order of APC/C substrate degradation in mitosis. J. Cell Biol. 2014, 207, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Kanatsu-Shinohara, M.; Onoyama, I.; Nakayama, K.I.; Shinohara, T. Skp1-Cullin-F-box (SCF)-type ubiquitin ligase FBXW7 negatively regulates spermatogonial stem cell self-renewal. Proc. Natl. Acad. Sci. USA 2014, 111, 8826–8831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.W.; Wilkinson, M.F. Transcriptional control of spermatogonial maintenance and differentiation. Semin. Cell Dev. Biol. 2014, 30, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Absalan, F.; Movahedin, M.; Mowla, S.J. Spermatogonial stem cell transplantation and subsequent orchidopexy in the bilateral cryptorchid mouse model. Cell J. 2011, 13, 143–148. [Google Scholar]
- Hermann, B.P.; Cheng, K.; Singh, A.; Roa-De, L.C.L.; Mutoji, K.N.; Chen, I.C.; Gildersleeve, H.; Lehle, J.D.; Mayo, M.; Westernstroer, B.; et al. The Mammalian Spermatogenesis Single-Cell Transcriptome, from Spermatogonial Stem Cells to Spermatids. Cell Rep. 2018, 25, 1650–1667. [Google Scholar] [CrossRef]
- Naughton, C.K.; Jain, S.; Strickland, A.M.; Gupta, A.; Milbrandt, J. Glial cell-line derived neurotrophic factor-mediated RET signaling regulates spermatogonial stem cell fate. Biol. Reprod. 2006, 74, 314–321. [Google Scholar] [CrossRef]
- Jijiwa, M.; Kawai, K.; Fukihara, J.; Nakamura, A.; Hasegawa, M.; Suzuki, C.; Sato, T.; Enomoto, A.; Asai, N.; Murakumo, Y.; et al. GDNF-mediated signaling via RET tyrosine 1062 is essential for maintenance of spermatogonial stem cells. Genes Cells 2008, 13, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Kanatsu-Shinohara, M.; Shinohara, T. Spermatogonial stem cell self-renewal and development. Annu. Rev. Cell Dev. Biol. 2013, 29, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Braun, R.E. Cyclical expression of GDNF is required for spermatogonial stem cell homeostasis. Development 2018, 145, dev151555. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Green, C.D.; Ma, Q.; Manske, G.L.; Shami, A.N.; Zheng, X.; Marini, S.; Moritz, L.; Sultan, C.; Gurczynski, S.J.; Moore, B.B.; et al. A Comprehensive Roadmap of Murine Spermatogenesis Defined by Single-Cell RNA-Seq. Dev. Cell 2018, 46, 651–667. [Google Scholar] [CrossRef] [PubMed]
- Eo, J.; Song, H.; Lim, H.J. Etv5, a transcription factor with versatile functions in male reproduction. Clin. Exp. Reprod. Med. 2012, 39, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Oatley, J.M.; Oatley, M.J.; Kaucher, A.V.; Avarbock, M.R.; Brinster, R.L. The POU domain transcription factor POU3F1 is an important intrinsic regulator of GDNF-induced survival and self-renewal of mouse spermatogonial stem cells. Biol. Reprod. 2010, 82, 1103–1111. [Google Scholar] [CrossRef]
- Oatley, J.M.; Avarbock, M.R.; Telaranta, A.I.; Fearon, D.T.; Brinster, R.L. Identifying genes important for spermatogonial stem cell self-renewal and survival. Proc. Natl. Acad. Sci. USA 2006, 103, 9524–9529. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kanatsu-Shinohara, M.; Inoue, K.; Ogonuki, N.; Miki, H.; Toyokuni, S.; Kimura, T.; Nakano, T.; Ogura, A.; Shinohara, T. Akt mediates self-renewal division of mouse spermatogonial stem cells. Development 2007, 134, 1853–1859. [Google Scholar] [CrossRef] [Green Version]
- Ocon-Grove, O.M.; Oatley, J.M. Molecular Mechanisms Regulating Spermatogonial Stem Cell Fate Decisions. In Stem Cell Biology and Regenerative Medicine; Orwig, K.E., Hermann, B.P., Eds.; Humana Press Inc.: Totowa, NJ, USA, 2011; pp. 135–157. [Google Scholar]
- Oatley, J.M.; Brinster, R.L. The germline stem cell niche unit in mammalian testes. Physiol. Rev. 2012, 92, 577–595. [Google Scholar] [CrossRef]
- Oatley, J.M.; Avarbock, M.R.; Brinster, R.L. Glial cell line-derived neurotrophic factor regulation of genes essential for self-renewal of mouse spermatogonial stem cells is dependent on Src family kinase signaling. J. Biol. Chem. 2007, 282, 25842–25851. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Goodyear, S.M.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. Spermatogonial stem cell self-renewal requires ETV5-mediated downstream activation of Brachyury in mice. Biol. Reprod. 2011, 85, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.; Garcia, T.X.; Hofmann, M.C. Regulation of GDNF expression in Sertoli cells. Reproduction 2019, 157, R95–R107. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, M.C. Gdnf signaling pathways within the mammalian spermatogonial stem cell niche. Mol. Cell. Endocrinol. 2008, 288, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Matoba, S.; Morimoto, H.; Ogura, A.; Shinohara, T. Improved serum- and feeder-free culture of mouse germline stem cells. Biol. Reprod. 2014, 91, 88. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Kanatsu-Shinohara, M.; Tanaka, T.; Morimoto, H.; Inoue, K.; Ogonuki, N.; Jijiwa, M.; Takahashi, M.; Ogura, A.; Shinohara, T. Functional differences between GDNF-dependent and FGF2-dependent mouse spermatogonial stem cell self-renewal. Stem Cell Rep. 2015, 4, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Kanatsu-Shinohara, M.; Toyokuni, S.; Shinohara, T. FGF2 mediates mouse spermatogonial stem cell self-renewal via upregulation of Etv5 and Bcl6b through MAP2K1 activation. Development 2012, 139, 1734–1743. [Google Scholar] [CrossRef] [PubMed]
- Kitadate, Y.; Jorg, D.J.; Tokue, M.; Maruyama, A.; Ichikawa, R.; Tsuchiya, S.; Segi-Nishida, E.; Nakagawa, T.; Uchida, A.; Kimura-Yoshida, C.; et al. Competition for Mitogens Regulates Spermatogenic Stem Cell Homeostasis in an Open Niche. Cell Stem Cell 2019, 24, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Yao, C.; Yang, C.; Zhu, Z.; Li, C.; Zhi, E.; Wang, J.; Li, P.; Chen, H.; Yuan, Q.; et al. Fibroblast growth factor-5 promotes spermatogonial stem cell proliferation via ERK and AKT activation. Stem Cell Res. Ther. 2019, 10, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, F.; Oatley, M.J.; Kaucher, A.V.; Yang, Q.E.; Bieberich, C.J.; Shashikant, C.S.; Oatley, J.M. Functional and molecular features of the Id4+ germline stem cell population in mouse testes. Genes Dev. 2014, 28, 1351–1362. [Google Scholar] [CrossRef]
- Garbuzov, A.; Pech, M.F.; Hasegawa, K.; Sukhwani, M.; Zhang, R.J.; Orwig, K.E.; Artandi, S.E. Purification of GFRalpha1+ and GFRalpha1-Spermatogonial Stem Cells Reveals a Niche-Dependent Mechanism for Fate Determination. Stem Cell Rep. 2018, 10, 553–567. [Google Scholar] [CrossRef]
- Tokue, M.; Ikami, K.; Mizuno, S.; Takagi, C.; Miyagi, A.; Takada, R.; Noda, C.; Kitadate, Y.; Hara, K.; Mizuguchi, H.; et al. SHISA6 Confers Resistance to Differentiation-Promoting Wnt/beta-Catenin Signaling in Mouse Spermatogenic Stem Cells. Stem Cell Rep. 2017, 8, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Kanatsu-Shinohara, M.; Inoue, K.; Takashima, S.; Takehashi, M.; Ogonuki, N.; Morimoto, H.; Nagasawa, T.; Ogura, A.; Shinohara, T. Reconstitution of mouse spermatogonial stem cell niches in culture. Cell Stem Cell 2012, 11, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.E.; Kim, D.; Kaucher, A.; Oatley, M.J.; Oatley, J.M. CXCL12-CXCR4 signaling is required for the maintenance of mouse spermatogonial stem cells. J. Cell Sci. 2013, 126, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- Wienholds, E.; Plasterk, R.H. MicroRNA function in animal development. FEBS Lett. 2005, 579, 5911–5922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Kanellopoulou, C.; Muljo, S.A.; Kung, A.L.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.M.; Rajewsky, K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005, 19, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Li, L.; Fu, H.; Li, S.; Li, J. Inactivation of Dicer1 has a severe cumulative impact on the formation of mature germ cells in mouse testes. Biochem. Biophys. Res. Commun. 2012, 422, 114–120. [Google Scholar] [CrossRef]
- Hayashi, K.; Chuva, D.S.L.S.; Kaneda, M.; Tang, F.; Hajkova, P.; Lao, K.; O’Carroll, D.; Das, P.P.; Tarakhovsky, A.; Miska, E.A.; et al. MicroRNA biogenesis is required for mouse primordial germ cell development and spermatogenesis. PLoS ONE 2008, 3, e1738. [Google Scholar] [CrossRef]
- Kimura, T.; Suzuki, A.; Fujita, Y.; Yomogida, K.; Lomeli, H.; Asada, N.; Ikeuchi, M.; Nagy, A.; Mak, T.W.; Nakano, T. Conditional loss of PTEN leads to testicular teratoma and enhances embryonic germ cell production. Development 2003, 130, 1691–1700. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Zhou, F.; Yuan, Q.; Zhang, W.; Qiu, Q.; Yu, X.; He, Z. miRNA-31-5p Mediates the Proliferation and Apoptosis of Human Spermatogonial Stem Cells via Targeting JAZF1 and Cyclin A2. Mol. Ther.-Nucleic Acids 2019, 14, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Lichner, Z.; Pall, E.; Kerekes, A.; Pallinger, E.; Maraghechi, P.; Bosze, Z.; Gocza, E. The miR-290-295 cluster promotes pluripotency maintenance by regulating cell cycle phase distribution in mouse embryonic stem cells. Differentiation 2011, 81, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Subramanyam, D.; Lamouille, S.; Judson, R.L.; Liu, J.Y.; Bucay, N.; Derynck, R.; Blelloch, R. Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Cai, T.; Zheng, C.; Lin, X.; Wang, G.; Liao, S.; Wang, X.; Gan, H.; Zhang, D.; Hu, X.; et al. MicroRNA-202 maintains spermatogonial stem cells by inhibiting cell cycle regulators and RNA binding proteins. Nucleic Acids Res. 2017, 45, 4142–4157. [Google Scholar] [CrossRef] [PubMed]
- Matson, C.K.; Murphy, M.W.; Griswold, M.D.; Yoshida, S.; Bardwell, V.J.; Zarkower, D. The mammalian doublesex homolog DMRT1 is a transcriptional gatekeeper that controls the mitosis versus meiosis decision in male germ cells. Dev. Cell 2010, 19, 612–624. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Hao, G.; Zhao, Z.; Wang, F.; Cao, J.; Yang, A. MicroRNA-224 regulates self-renewal of mouse spermatogonial stem cells via targeting DMRT1. J. Cell. Mol. Med. 2016, 20, 1503–1512. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Lin, X.; Du, T.; Xu, K.; Shen, H.; Wei, F.; Hao, W.; Lin, T.; Lin, X.; Qin, Y.; et al. Targeted Disruption of miR-17-92 Impairs Mouse Spermatogenesis by Activating mTOR Signaling Pathway. Medicine (Baltimore) 2016, 95, e2713. [Google Scholar] [CrossRef] [PubMed]
- Dyce, P.W.; Toms, D.; Li, J. Stem cells and germ cells: MicroRNA and gene expression signatures. Histol. Histopathol. 2010, 25, 505–513. [Google Scholar] [PubMed]
- Qi, J.; Yu, J.Y.; Shcherbata, H.R.; Mathieu, J.; Wang, A.J.; Seal, S.; Zhou, W.; Stadler, B.M.; Bourgin, D.; Wang, L.; et al. microRNAs regulate human embryonic stem cell division. Cell Cycle 2009, 8, 3729–3741. [Google Scholar] [CrossRef] [Green Version]
- Radisky, D.C. miR-200c at the nexus of epithelial-mesenchymal transition, resistance to apoptosis, and the breast cancer stem cell phenotype. Breast Cancer Res. 2011, 13, 110. [Google Scholar] [CrossRef]
- Niu, Z.; Goodyear, S.M.; Rao, S.; Wu, X.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. MicroRNA-21 regulates the self-renewal of mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 2011, 108, 12740–12745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kedde, M.; Strasser, M.J.; Boldajipour, B.; Oude, V.J.; Slanchev, K.; le Sage, C.; Nagel, R.; Voorhoeve, P.M.; van Duijse, J.; Orom, U.A.; et al. RNA-binding protein Dnd1 inhibits microRNA access to target mRNA. Cell 2007, 131, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Lu, Y.; Sun, H.; Tao, D.; Zhang, S.; Liu, W.; Ma, Y. A microarray for microRNA profiling in mouse testis tissues. Reproduction 2007, 134, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Lu, Y.; Sun, H.; Qiu, W.; Tao, D.; Liu, Y.; Chen, H.; Yang, Y.; Zhang, S.; Li, X.; et al. Microarray profiling of microRNAs expressed in testis tissues of developing primates. J. Assist. Reprod. Genet. 2009, 26, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Characteristics of mammalian spermatogonial stem cell (SSC) development. Gray areas correspond to the cytoplasm, dark gray areas correspond to the cytomembrane, lavender and green areas correspond to the nucleus.
Figure 1.
Characteristics of mammalian spermatogonial stem cell (SSC) development. Gray areas correspond to the cytoplasm, dark gray areas correspond to the cytomembrane, lavender and green areas correspond to the nucleus.

Figure 2.
(A) Illustration of the main cell cycle genes expressed and likely controlling the cell cycle in proliferating mouse PGCs. (B) The role of APC/C in the cell mitosis cycle.
Figure 2.
(A) Illustration of the main cell cycle genes expressed and likely controlling the cell cycle in proliferating mouse PGCs. (B) The role of APC/C in the cell mitosis cycle.

Figure 3.
Current understanding of the signaling pathways regulating the mitosis of SSCs in mouse testes.
Figure 3.
Current understanding of the signaling pathways regulating the mitosis of SSCs in mouse testes.


{kind=link}
{kind=link}
{kind=link}
Table 1.
miRNA molecules implicated in germ cell mitosis.
| Name | Expression | Proposed Function | Targets Involved in Mammalian Gametogenesis | Reference |
|---|---|---|---|---|
| miR-17-92 cluster | PGCs, ES cells | Regulator of differentiation, proliferation and apoptosis | STAT3, E2F1, PTEN | [110,118,119] |
| miRNA-31-5p | SSCs | Regulator of SSCs proliferation | JAZF1 and Cyclin A2 | [112] |
| miR-290-295 cluster | PGCs | G1 to S phase cell cycle control | WEE1, FBXL5 | [110,113] |
| miR-202 | SSCs | Regulator of cell cycle and apoptosis of mitosis | Rbfox, Cpeb1 | [115] |
| miR-224 | SSCs | Control SSCs self-renewal and cyclical gene expression | DMRT1 | [117] |
| MiR-302-67 cluster | PGCs | targeting inhibitors of the G1/S transition | Cdkn1a | [114,120] |
| miR-125a | Later male PGCs | Control of differentiation | LIN28 | [110] |
| miR-200c | Early PGCs | Control of apoptosis | ZEB1, TRKB | [110,121] |
| miR-21 | SSCs | SSCs self-renewal, anti-apoptosis | ZEB1, TRKB | [122] |
| miR-221 | PGCs | Regulate mitotic arrest in male germ cells | DND1 | [123] |
| miR-34c | PGCs | Cell cycle regulator | CCND3, CCNG1, CCNB1 NOTCH2 | [124,125] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, J.-H.; Li, Y.; Deng, S.-L.; Liu, Y.-X.; Lian, Z.-X.; Yu, K. Recent Research Advances in Mitosis during Mammalian Gametogenesis. Cells 2019, 8, 567. https://doi.org/10.3390/cells8060567
AMA Style
Wang J-H, Li Y, Deng S-L, Liu Y-X, Lian Z-X, Yu K. Recent Research Advances in Mitosis during Mammalian Gametogenesis. Cells. 2019; 8(6):567. https://doi.org/10.3390/cells8060567
Chicago/Turabian StyleWang, Jia-Hao, Yan Li, Shou-Long Deng, Yi-Xun Liu, Zheng-Xing Lian, and Kun Yu. 2019. "Recent Research Advances in Mitosis during Mammalian Gametogenesis" Cells 8, no. 6: 567. https://doi.org/10.3390/cells8060567
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.