
Biotechnological Potential of the Stress Response and Plant Cell Death Regulators Proteins in the Biofuel Industry


 ,
,  , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Arabidopsis thaliana Plant Material
2.2. Populus tremula × tremuloides Plant Material and Genetic Transformation
2.3. Aspen Growth Condition
2.4. Inducing of Lysigenous Aerenchyma Formation by Root Hypoxia Stress
2.5. Determining to Reduce Sugar Content in Arabidopsis thaliana Hypocotyls
2.6. Cellulase and Xylanase Activity Assays
2.7. Native-PAGE Separation and Zymographic Activity Assay
2.8. Maldi-TOF Analysis of Isolated Proteins
2.9. RNA Isolation and qPCR Analysis
2.10. RNAseq Analysis
2.11. Biometric Parameters and Gas Exchanging Measurement
2.12. Determination of Photosynthetic Pigments Content
2.13. Gene Ontology
2.14. Cellulose, Hemicellulose and Lignin Content
2.15. The Alkaline Pre-Treatment and Enzymatic Hydrolysis of Aspen Wood
2.16. The Ethanol Fermentation Process
2.17. The HPLC Analysis
2.18. The Analysis of Cellulose Polymerization Degree
2.19. Statistic
3. Results
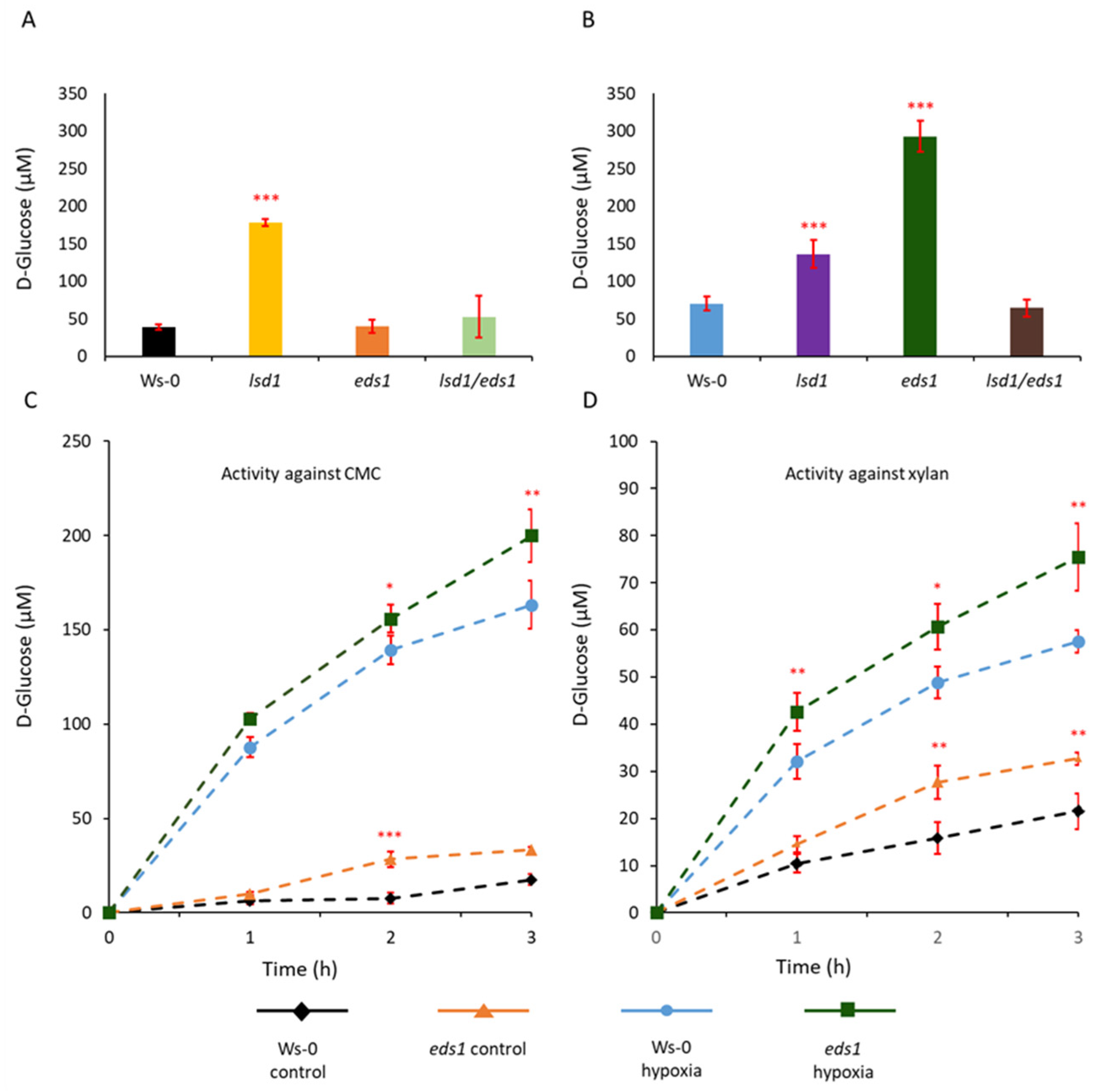
3.1. EDS1 and LSD1 Are Involved in Lysigenous Aerenchyma Formation
3.2. In Vitro Cellulolytic and Xylanolytic Activities of Enzymes Isolated from eds1 Hypocotyls
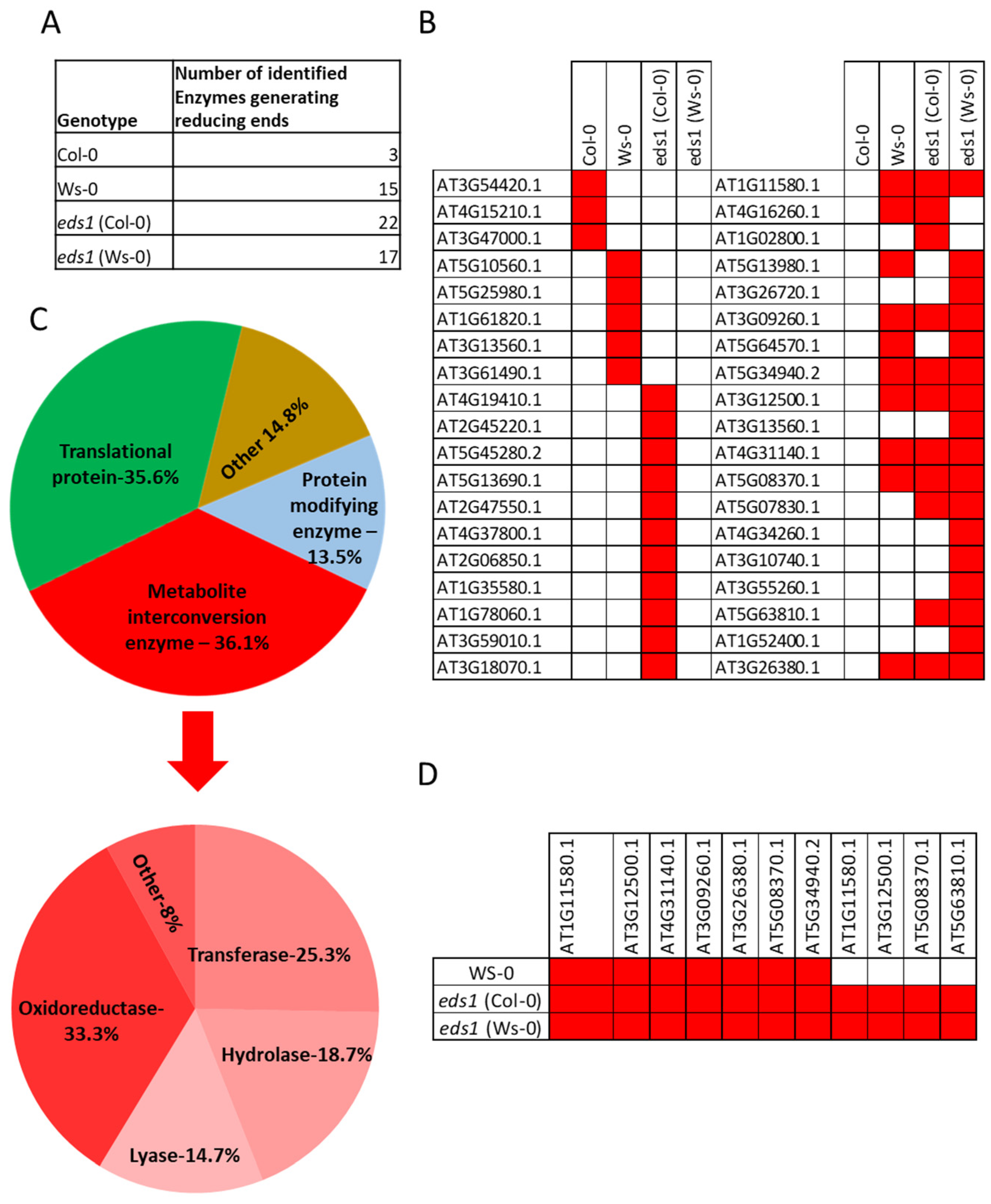
3.3. Enzymes Generating Reducing Ends Were Found in Hypocotyls and Roots Undergoing Lysigenous Aerenchyma Formation
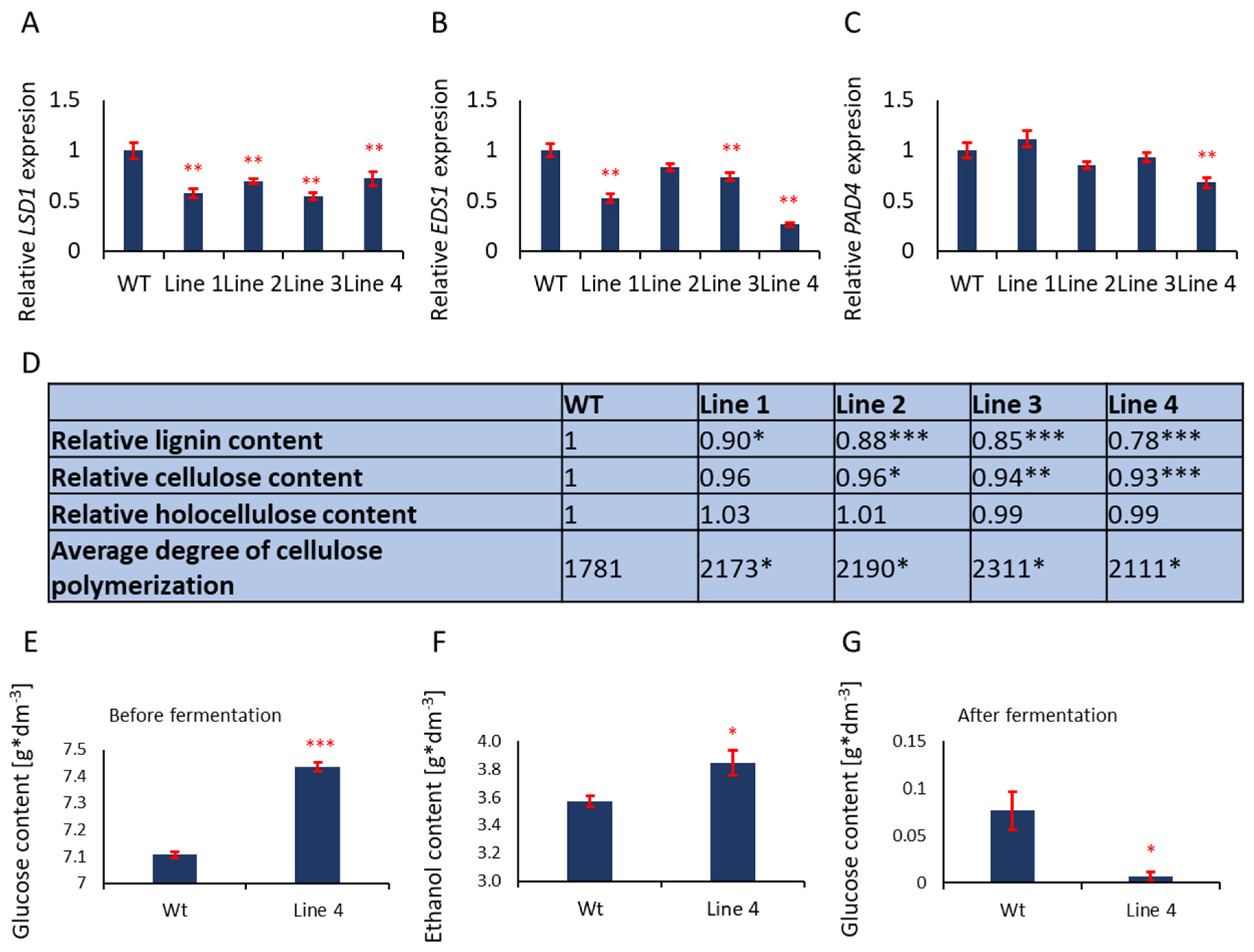
3.4. Cell Walls of Aspen with Deregulated PCD Growing in the Field Contain Less Lignin and Longer Cellulose Fibers
3.5. Strong Reduction in Lignin and Higher Cellulose Polymerization Degree in the Wood of Line 4 Allows High Fermentation Efficiency
3.6. PtLSD1, PtEDS1, and PtPAD4 Strongly Influence Cambium and Xylem Transcriptome in Line 4
3.7. Transgenic Lines Exhibited Similar Phenotype and Development Level as Wild Type Trees
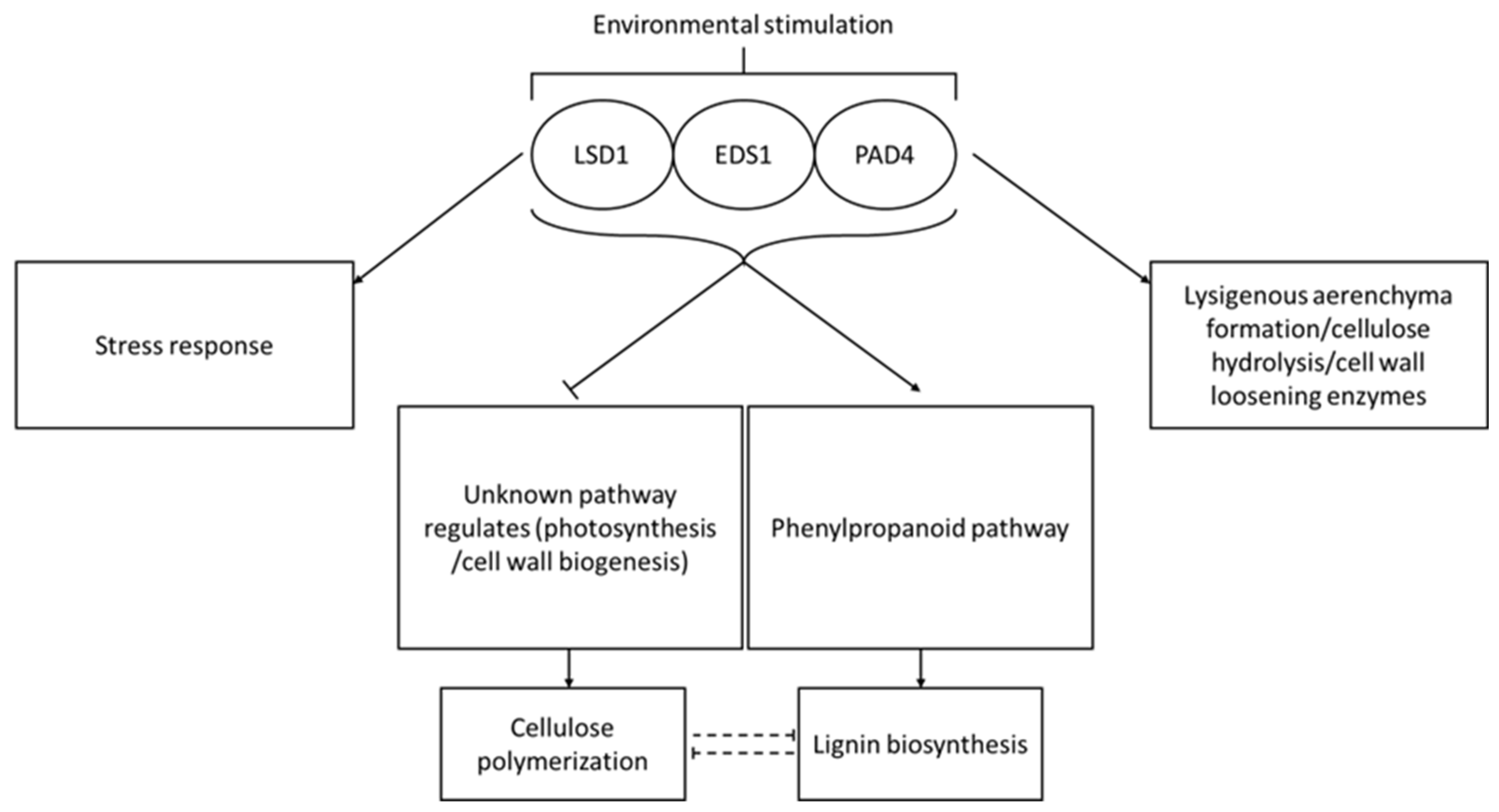
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LSD1 | LESION SIMULATING DISEASE 1 |
| PAD4 | PHYTOALEXIN DEFICIENT 4 |
| EDS1 | ENHANCED DISEASE SUSCEPTIBILITY |
| PCD | Programed Cell Death |
| CMC | Carboxymethyl cellulose |
| DEG | Differential expressed genes |
| 4CL | 4-coumarate:coenzyme A ligase |
| CWL | cell wall loosening |
| DEG | differentially expressed genes |
| MS | Murashige and Skoog medium |
References
- Hoengenaert, L.; Wouters, M.; Kim, H.; De Meester, B.; Morreel, K.; Vandersyppe, S.; Pollier, J.; Desmet, S.; Goeminne, G.; Ralph, J.; et al. Overexpression of the Scopoletin Biosynthetic Pathway Enhances Lignocellulosic Biomass Processing. Sci. Adv. 2022, 8, eabo5738. [Google Scholar] [CrossRef]
- Zoghlami, A.; Paës, G. Lignocellulosic Biomass: Understanding Recalcitrance and Predicting Hydrolysis. Front. Chem. 2019, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, A.; Kumar, D.; Girdhar, M.; Kumar, A.; Goyal, A.; Malik, T.; Mohan, A. Strategies of Pretreatment of Feedstocks for Optimized Bioethanol Production: Distinct and Integrated Approaches. Biotechnol. Biofuels Bioprod. 2023, 16, 44. [Google Scholar] [CrossRef]
- Czekała, W.; Pulka, J.; Jasiński, T.; Szewczyk, P.; Bojarski, W.; Jasiński, J. Waste as Substrates for Agricultural Biogas Plants: A Case Study from Poland. J. Water Land Dev. 2023, 45–50. [Google Scholar] [CrossRef]
- Turumtay, H. Cell Wall Engineering by Heterologous Expression of Cell Wall-Degrading Enzymes for Better Conversion of Lignocellulosic Biomass into Biofuels. Bioenerg. Res. 2015, 8, 1574–1588. [Google Scholar] [CrossRef]
- Khan, I.; Akhtar, M.W. BioenergyProduction from Plant Biomass: Bioethanol from Concept to Reality. Nat. Prec. 2011, 1. [Google Scholar] [CrossRef] [Green Version]
- Alonso, D.M.; Hakim, S.H.; Zhou, S.; Won, W.; Hosseinaei, O.; Tao, J.; Garcia-Negron, V.; Motagamwala, A.H.; Mellmer, M.A.; Huang, K.; et al. Increasing the Revenue from Lignocellulosic Biomass: Maximizing Feedstock Utilization. Sci. Adv. 2017, 3, e1603301. [Google Scholar] [CrossRef] [Green Version]
- Sidana, A.; Yadav, S.K. Recent Developments in Lignocellulosic Biomass Pretreatment with a Focus on Eco-Friendly, Non-Conventional Methods. J. Clean. Prod. 2022, 335, 130286. [Google Scholar] [CrossRef]
- Antczak, A.; Szadkowski, J.; Szadkowska, D.; Zawadzki, J. Assessment of the Effectiveness of Liquid Hot Water and Steam Explosion Pretreatments of Fast-Growing Poplar (Populus trichocarpa) Wood. Wood Sci. Technol. 2022, 56, 87–109. [Google Scholar] [CrossRef]
- Lu, X.; Zheng, X.; Li, X.; Zhao, J. Adsorption and Mechanism of Cellulase Enzymes onto Lignin Isolated from Corn Stover Pretreated with Liquid Hot Water. Biotechnol. Biofuels 2016, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, E.; Baetz, K. Multi-Faceted Systems Biology Approaches Present a Cellular Landscape of Phenolic Compound Inhibition in Saccharomyces Cerevisiae. Front. Bioeng. Biotechnol. 2020, 8, 539902. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, Q.; Rojas, R.; Yan, M.; Lawoko, M.; Berglund, L. Lignin-Retaining Transparent Wood. ChemSusChem 2017, 10, 3445–3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escamez, S.; Tuominen, H. Contribution of Cellular Autolysis to Tissular Functions during Plant Development. Curr. Opin. Plant Biol. 2017, 35, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Mühlenbock, P.; Plaszczyca, M.; Plaszczyca, M.; Mellerowicz, E.; Karpinski, S. Lysigenous Aerenchyma Formation in Arabidopsis Is Controlled by LESION SIMULATING DISEASE1. Plant Cell 2007, 19, 3819–3830. [Google Scholar] [CrossRef] [Green Version]
- Taylor, G. Populus: Arabidopsis for Forestry. Do We Need a Model Tree? Ann. Bot. 2002, 90, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Chaffey, N.; Cholewa, E.; Regan, S.; Sundberg, B. Secondary Xylem Development in Arabidopsis: A Model for Wood Formation. Physiol. Plant 2002, 114, 594–600. [Google Scholar] [CrossRef]
- Wituszyńska, W.; Ślesak, I.; Vanderauwera, S.; Szechyńska-Hebda, M.; Kornaś, A.; Kelen, K.V.D.; Mühlenbock, P.; Karpińska, B.; Maćkowski, S.; Breusegem, F.V.; et al. LESION SIMULATING DISEASE1, ENHANCED DISEASE SUSCEPTIBILITY1, and PHYTOALEXIN DEFICIENT4 Conditionally Regulate Cellular Signaling Homeostasis, Photosynthesis, Water Use Efficiency, and Seed Yield in Arabidopsis. Plant Physiol. 2013, 161, 1795–1805. [Google Scholar] [CrossRef] [Green Version]
- Bernacki, M.J.; Czarnocka, W.; Rusaczonek, A.; Witoń, D.; Kęska, S.; Czyż, J.; Szechyńska-Hebda, M.; Karpiński, S. LSD1, EDS1 and PAD4-Dependent Conditional Correlation among Salicylic Acid, Hydrogen Peroxide, Water Use Efficiency, and Seed Yield in Arabidopsis Thaliana. Physiol. Plant. 2018, 165, 369–382. [Google Scholar] [CrossRef]
- Czarnocka, W.; Van Der Kelen, K.; Willems, P.; Szechyńska-Hebda, M.; Shahnejat-Bushehri, S.; Balazadeh, S.; Rusaczonek, A.; Mueller-Roeber, B.; Van Breusegem, F.; Karpiński, S. The Dual Role of LESION SIMULATING DISEASE 1 as a Condition-Dependent Scaffold Protein and Transcription Regulator. Plant Cell Environ. 2017, 40, 2644–2662. [Google Scholar] [CrossRef] [Green Version]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAYTM Vectors for Agrobacterium-Mediated Plant Transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Nilsson, O.; Aldén, T.; Sitbon, F.; Little, C.H.A.; Chalupa, V.; Sandberg, G.; Olsson, O. Spatial Pattern of Cauliflower Mosaic Virus 35S Promoter-Luciferase Expression in Transgenic Hybrid Aspen Trees Monitored by Enzymatic Assay and Non-Destructive Imaging. Transgenic Res. 1992, 1, 209–220. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Czarnocka, W.; Witoń, D.; Rusaczonek, A.; Szechyńska-Hebda, M.; Ślesak, I.; Dąbrowska-Bronk, J.; Karpiński, S. ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1) Affects Development, Photosynthesis, and Hormonal Homeostasis in Hybrid Aspen (Populus tremula L. × P. tremuloides). J. Plant Physiol. 2018, 226, 91–102. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The Genome of Black Cottonwood, Populus Trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusaczonek, A.; Czarnocka, W.; Kacprzak, S.; Witoń, D.; Ślesak, I.; Szechyńska-Hebda, M.; Gawroński, P.; Karpiński, S. Role of Phytochromes A and B in the Regulation of Cell Death and Acclimatory Responses to UV Stress in Arabidopsis Thaliana. J. Exp. Bot. 2015, 66, erv375. [Google Scholar] [CrossRef] [Green Version]
- Wise, L.E.; Murphy, M.; Addieco, A.A.d’. Chlorite Holocellulose, Its Fractionation and Bearing on Summative Wood Analysis and Studies on the Hemicelluloses. Pap. Trade J. 1946, 122, 35–43. [Google Scholar]
- Sluiter, A. Determination of Extractives in Biomass: Laboratory Analytical Procedure (LAP); Issue Date 7/17/2005. Tech. Rep. 2008, 12. [Google Scholar]
- Antczak, A.; Marchwicka, M.; Szadkowski, J.; Drożdżek, M.; Gawron, J.; Radomski, A.; Zawadzki, J. Sugars Yield Obtained after Acid and Enzymatic Hydrolysis of Fast-Growing Poplar Wood Species. BioResources 2018, 13, 8629–8645. [Google Scholar] [CrossRef]
- Adney, B.; Baker, J. Measurement of Cellulase Activities: Laboratory Analytical Procedure (LAP); Issue Date: 08/12/1996. Tech. Rep. 2008, 11. [Google Scholar]
- Mateo, A.; Mühlenbock, P.; Rustérucci, C.; Chang, C.C.-C.; Miszalski, Z.; Karpinska, B.; Parker, J.E.; Mullineaux, P.M.; Karpinski, S. LESION SIMULATING DISEASE 1 Is Required for Acclimation to Conditions That Promote Excess Excitation Energy. Plant Physiol. 2004, 136, 2818–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feys, B.J.; Moisan, L.J.; Newman, M.-A.; Parker, J.E. Direct Interaction between the Arabidopsis Disease Resistance Signaling Proteins, EDS1 and PAD4. EMBO J. 2001, 20, 5400–5411. [Google Scholar] [CrossRef] [PubMed]
- Ślesak, I.; Szechyńska-Hebda, M.; Fedak, H.; Sidoruk, N.; Dąbrowska-Bronk, J.; Witoń, D.; Rusaczonek, A.; Antczak, A.; Drożdżek, M.; Karpińska, B.; et al. PHYTOALEXIN DEFICIENT 4 Affects Reactive Oxygen Species Metabolism, Cell Wall and Wood Properties in Hybrid Aspen (Populus tremula L. × Tremuloides). Plant Cell Environ. 2015, 38, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Szechyńska-Hebda, M.; Czarnocka, W.; Hebda, M.; Karpiński, S. PAD4, LSD1 and EDS1 Regulate Drought Tolerance, Plant Biomass Production, and Cell Wall Properties. Plant Cell Rep. 2016, 35, 527–539. [Google Scholar] [CrossRef]
- Pinto, A.S.S.; Brondi, M.G.; de Freitas, J.V.; Furlan, F.F.; Ribeiro, M.P.A.; Giordano, R.C.; Farinas, C.S. Mitigating the Negative Impact of Soluble and Insoluble Lignin in Biorefineries. Renew. Energy 2021, 173, 1017–1026. [Google Scholar] [CrossRef]
- Douglas, C.J. Phenylpropanoid Metabolism and Lignin Biosynthesis: From Weeds to Trees. Trends Plant Sci. 1996, 1, 171–178. [Google Scholar] [CrossRef]
- Kumar, V.; Hainaut, M.; Delhomme, N.; Mannapperuma, C.; Immerzeel, P.; Street, N.R.; Henrissat, B.; Mellerowicz, E.J. Poplar Carbohydrate-Active Enzymes: Whole-Genome Annotation and Functional Analyses Based on RNA Expression Data. Plant J. 2019, 99, 589–609. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Donev, E.N.; Barbut, F.R.; Kushwah, S.; Mannapperuma, C.; Urbancsok, J.; Mellerowicz, E.J. Genome-Wide Identification of Populus Malectin/Malectin-Like Domain-Containing Proteins and Expression Analyses Reveal Novel Candidates for Signaling and Regulation of Wood Development. Front. Plant Sci. 2020, 11, 588846. [Google Scholar] [CrossRef]
- Taherzadeh, M.J.; Karimi, K. Pretreatment of Lignocellulosic Wastes to Improve Ethanol and Biogas Production: A Review. Int. J. Mol. Sci. 2008, 9, 1621–1651. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, Y.; Li, C.; Yin, B.; Liu, X.; Guo, X.; Zhang, C.; Liu, D.; Hwang, I.; Li, H.; et al. PtomtAPX Is an Autonomous Lignification Peroxidase during the Earliest Stage of Secondary Wall Formation in Populus Tomentosa Carr. Nat. Plants 2022, 8, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Teeples, M.; Lin, L.; de Lucas, M.; Turco, G.; Toal, T.W.; Gaudinier, A.; Young, N.F.; Trabucco, G.M.; Veling, M.T.; Lamothe, R.; et al. An Arabidopsis Gene Regulatory Network for Secondary Cell Wall Synthesis. Nature 2015, 517, 571–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuetz, M.; Smith, R.; Ellis, B. Xylem Tissue Specification, Patterning, and Differentiation Mechanisms. J. Exp. Bot. 2013, 64, 11–31. [Google Scholar] [CrossRef]
- Karpiński, S.; Szechyńska-Hebda, M.; Wituszyńska, W.; Burdiak, P. Light Acclimation, Retrograde Signalling, Cell Death and Immune Defences in Plants. Plant Cell Environ. 2013, 36, 736–744. [Google Scholar] [CrossRef]
- Mühlenbock, P.; Szechynska-Hebda, M.; Plaszczyca, M.; Baudo, M.; Mateo, A.; Mullineaux, P.M.; Parker, J.E.; Karpinska, B.; Karpinski, S. Chloroplast Signaling and LESION SIMULATING DISEASE1 Regulate Crosstalk between Light Acclimation and Immunity in Arabidopsis. Plant Cell 2008, 20, 2339–2356. [Google Scholar] [CrossRef] [Green Version]
- Bjurhager, I.; Olsson, A.-M.; Zhang, B.; Gerber, L.; Kumar, M.; Berglund, L.A.; Burgert, I.; Sundberg, B.; Salmén, L. Ultrastructure and Mechanical Properties of Populus Wood with Reduced Lignin Content Caused by Transgenic Down-Regulation of Cinnamate 4-Hydroxylase. Biomacromolecules 2010, 11, 2359–2365. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-J.; Harding, S.A.; Lung, J.; Popko, J.L.; Ralph, J.; Stokke, D.D.; Tsai, C.-J.; Chiang, V.L. Repression of Lignin Biosynthesis Promotes Cellulose Accumulation and Growth in Transgenic Trees. Nat. Biotechnol. 1999, 17, 808–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, C.; Zhao, H.; Wang, H.; Xing, Z.; Du, K.; Song, Y.; Wei, J. Obtaining the Transgenic Poplars with Low Lignin Content through Down-Regulation Of4CL. Chin. Sci. Bull. 2004, 49, 905–909. [Google Scholar] [CrossRef]
- Li, L.; Zhou, Y.; Cheng, X.; Sun, J.; Marita, J.M.; Ralph, J.; Chiang, V.L. Combinatorial Modification of Multiple Lignin Traits in Trees through Multigene Cotransformation. Proc. Natl. Acad. Sci. USA 2003, 100, 4939–4944. [Google Scholar] [CrossRef]
- Sen, S.K.; Min, D.; Savithri, D.; Lu, F.; Jameel, H.; Chiang, V.; Chang, H. Field-Grown Transgenic Hybrid Poplar with Modified Lignin Biosynthesis to Improve Enzymatic Saccharification Efficiency. ACS Sustain. Chem. Eng. 2017, 5, 2407–2414. [Google Scholar] [CrossRef]
- Stout, A.T.; Davis, A.A.; Domec, J.-C.; Yang, C.; Shi, R.; King, J.S. Growth under Field Conditions Affects Lignin Content and Productivity in Transgenic Populus Trichocarpa with Altered Lignin Biosynthesis. Biomass Bioenergy 2014, 68, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Leplé, J.-C.; Dauwe, R.; Morreel, K.; Storme, V.; Lapierre, C.; Pollet, B.; Naumann, A.; Kang, K.-Y.; Kim, H.; Ruel, K.; et al. Downregulation of Cinnamoyl-Coenzyme A Reductase in Poplar: Multiple-Level Phenotyping Reveals Effects on Cell Wall Polymer Metabolism and Structure. Plant Cell 2007, 19, 3669–3691. [Google Scholar] [CrossRef] [Green Version]
- Van Acker, R.; Leplé, J.-C.; Aerts, D.; Storme, V.; Goeminne, G.; Ivens, B.; Légée, F.; Lapierre, C.; Piens, K.; Van Montagu, M.C.E.; et al. Improved Saccharification and Ethanol Yield from Field-Grown Transgenic Poplar Deficient in Cinnamoyl-CoA Reductase. Proc. Natl. Acad. Sci. USA 2014, 111, 845–850. [Google Scholar] [CrossRef]
- Tian, X.; Xie, J.; Zhao, Y.; Lu, H.; Liu, S.; Qu, L.; Li, J.; Gai, Y.; Jiang, X. Sense-, Antisense- and RNAi-4CL1 Regulate Soluble Phenolic Acids, Cell Wall Components and Growth in Transgenic Populus Tomentosa Carr. Plant Physiol. Biochem. 2013, 65, 111–119. [Google Scholar] [CrossRef]
- Min, D.; Li, Q.; Jameel, H.; Chiang, V.; Chang, H. The Cellulase-Mediated Saccharification on Wood Derived from Transgenic Low-Lignin Lines of Black Cottonwood (Populus trichocarpa). Appl Biochem Biotechnol 2012, 168, 947–955. [Google Scholar] [CrossRef]
- Min, D.; Yang, C.; Chiang, V.; Jameel, H.; Chang, H. The Influence of Lignin–Carbohydrate Complexes on the Cellulase-Mediated Saccharification II: Transgenic Hybrid Poplars (Populus nigra L. and Populus maximowiczii A.). Fuel 2014, 116, 56–62. [Google Scholar] [CrossRef]
- Coleman, H.D.; Park, J.-Y.; Nair, R.; Chapple, C.; Mansfield, S.D. RNAi-Mediated Suppression of p-Coumaroyl-CoA 3’-Hydroxylase in Hybrid Poplar Impacts Lignin Deposition and Soluble Secondary Metabolism. Proc. Natl. Acad. Sci. USA 2008, 105, 4501–4506. [Google Scholar] [CrossRef]
- Wituszyńska, W.; Gałązka, K.; Rusaczonek, A.; Vanderauwera, S.; Van Breusegem, F.; Karpiński, S. Multivariable Environmental Conditions Promote Photosynthetic Adaptation Potential in Arabidopsis Thaliana. J. Plant Physiol. 2013, 170, 548–559. [Google Scholar] [CrossRef]
- Gao, F.; Shu, X.; Ali, M.B.; Howard, S.; Li, N.; Winterhagen, P.; Qiu, W.; Gassmann, W. A Functional EDS1 Ortholog Is Differentially Regulated in Powdery Mildew Resistant and Susceptible Grapevines and Complements an Arabidopsis Eds1 Mutant. Planta 2010, 231, 1037–1047. [Google Scholar] [CrossRef]
- Guo, J.; Bai, P.; Yang, Q.; Liu, F.; Wang, X.; Huang, L.; Kang, Z. Wheat Zinc Finger Protein TaLSD1, a Negative Regulator of Programmed Cell Death, Is Involved in Wheat Resistance against Stripe Rust Fungus. Plant Physiol. Biochem. 2013, 71, 164–172. [Google Scholar] [CrossRef]
- Guan, Q.; Ma, H.; Wang, Z.; Wang, Z.; Bu, Q.; Liu, S. A Rice LSD1-like-Type ZFP Gene OsLOL5 Enhances Saline-Alkaline Tolerance in Transgenic Arabidopsis Thaliana, Yeast and Rice. BMC Genom. 2016, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Li, B.; Liu, Y.; Zhu, J.; Li, G.; Hou, G.; Zhou, J.; Qiu, X. Critical Role of Degree of Polymerization of Cellulose in Super-Strong Nanocellulose Films. Matter 2020, 2, 1000–1014. [Google Scholar] [CrossRef]
- Dietrich, R.A.; Richberg, M.H.; Schmidt, R.; Dean, C.; Dangl, J.L. A Novel Zinc Finger Protein Is Encoded by the Arabidopsis LSD1 Gene and Functions as a Negative Regulator of Plant Cell Death. Cell 1997, 88, 685–694. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernacki, M.J.; Mielecki, J.; Antczak, A.; Drożdżek, M.; Witoń, D.; Dąbrowska-Bronk, J.; Gawroński, P.; Burdiak, P.; Marchwicka, M.; Rusaczonek, A.; et al. Biotechnological Potential of the Stress Response and Plant Cell Death Regulators Proteins in the Biofuel Industry. Cells 2023, 12, 2018. https://doi.org/10.3390/cells12162018
Bernacki MJ, Mielecki J, Antczak A, Drożdżek M, Witoń D, Dąbrowska-Bronk J, Gawroński P, Burdiak P, Marchwicka M, Rusaczonek A, et al. Biotechnological Potential of the Stress Response and Plant Cell Death Regulators Proteins in the Biofuel Industry. Cells. 2023; 12(16):2018. https://doi.org/10.3390/cells12162018
Chicago/Turabian StyleBernacki, Maciej Jerzy, Jakub Mielecki, Andrzej Antczak, Michał Drożdżek, Damian Witoń, Joanna Dąbrowska-Bronk, Piotr Gawroński, Paweł Burdiak, Monika Marchwicka, Anna Rusaczonek, and et al. 2023. "Biotechnological Potential of the Stress Response and Plant Cell Death Regulators Proteins in the Biofuel Industry" Cells 12, no. 16: 2018. https://doi.org/10.3390/cells12162018