

Astrocytic CD44 Deficiency Reduces the Severity of Kainate-Induced Epilepsy
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Generation of CD44 Knockout Mice
2.3. Western Blot
2.4. Induction of Status Epilepticus
2.5. Electrode Implantation and EEG-Video Monitoring
2.6. Analysis of Video EEG Recordings
2.7. Immunofluorescence
2.8. Fluorescence Microscopy Analysis
2.9. SBEM Sample Preparation
2.10. SBEM Imaging
2.11. SBEM Analysis
2.12. Statistical Analysis and Figures
3. Results
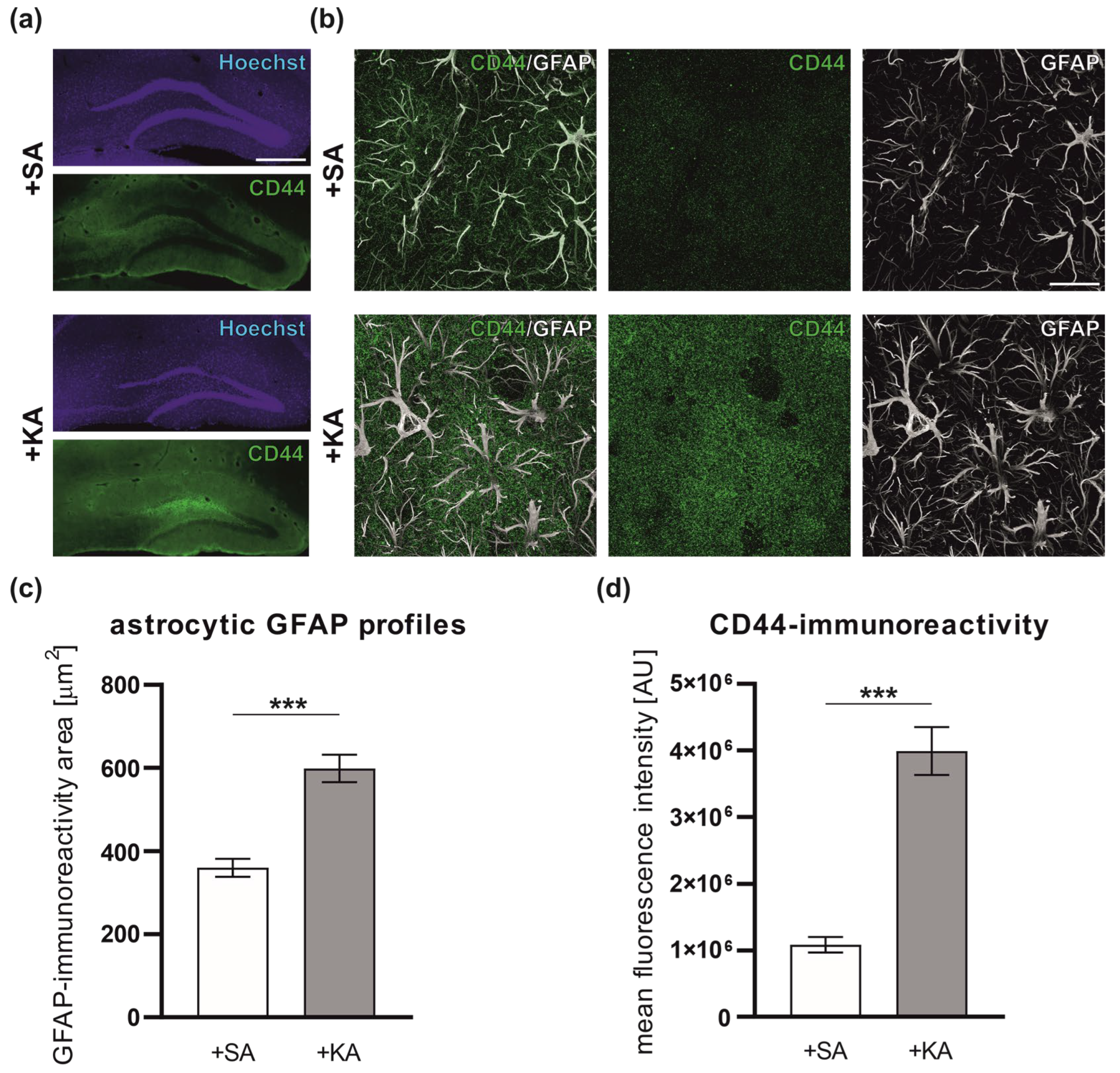
3.1. Increased Level of CD44 in Astrocytes following Kainate-Induced Status Epilepticus
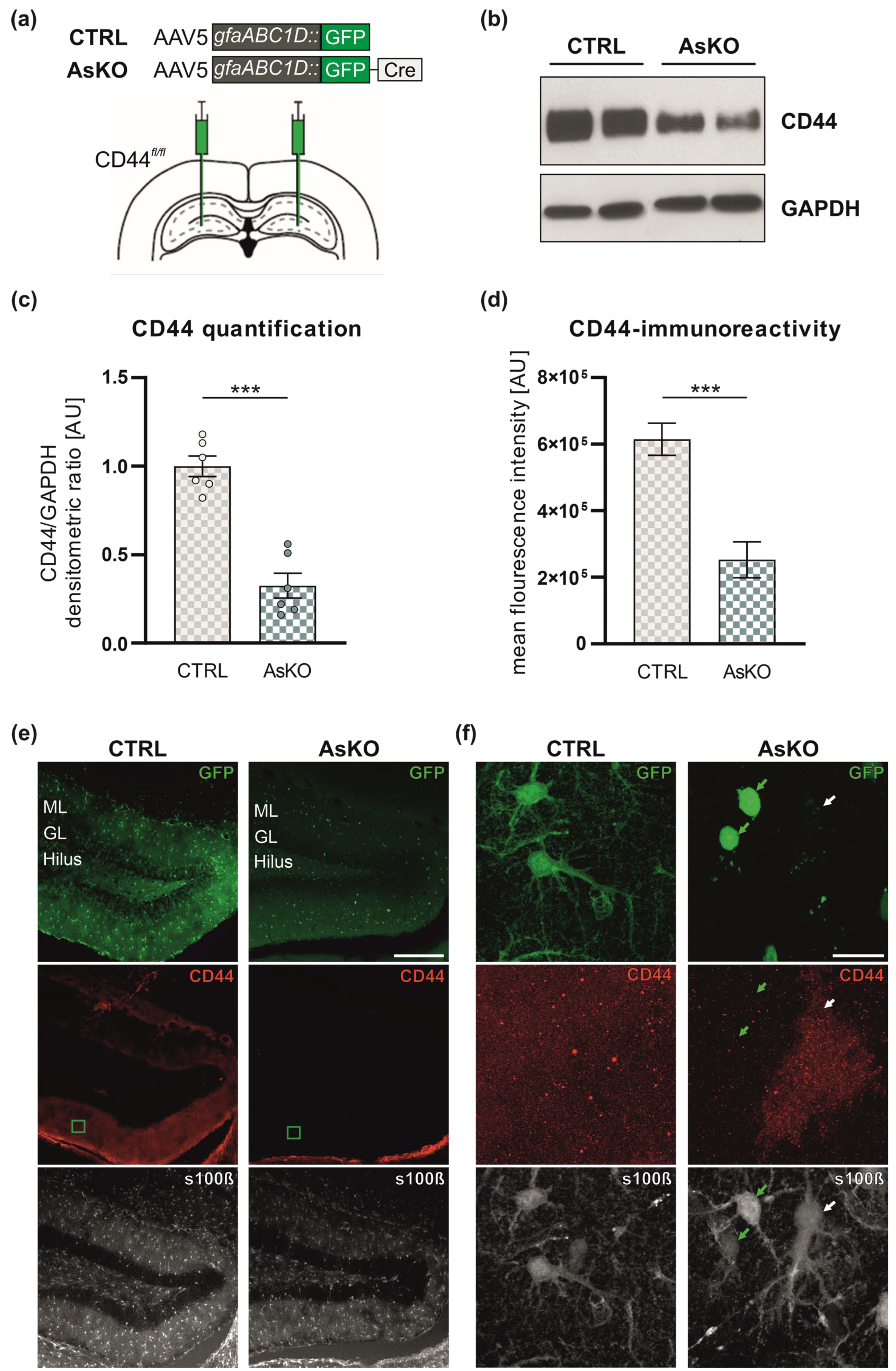
3.2. CD44 Depletion in Astrocytes by AAV5-Mediated Expression of gfaABC1D::GFP-Cre Recombinase in the Hippocampi of CD44fl/fl Mice
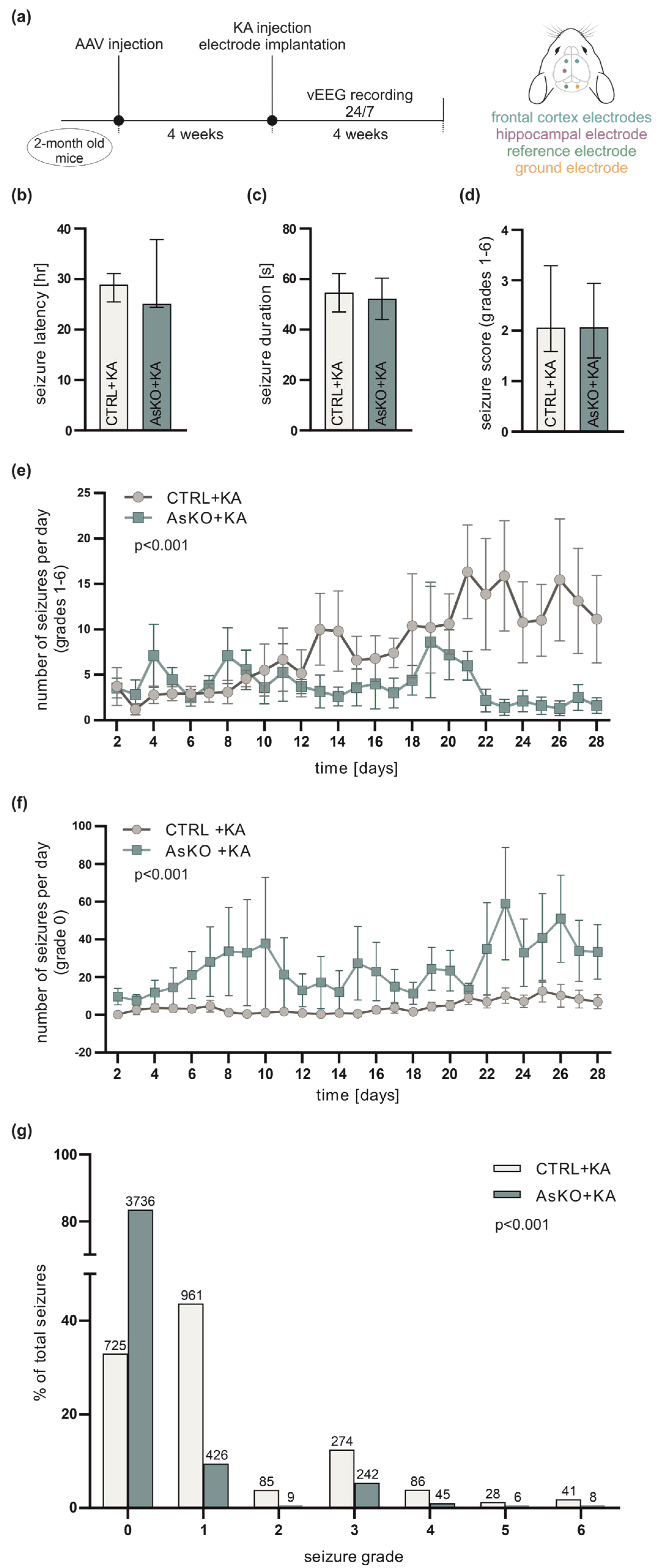
3.3. CD44 Deficiency Reduces KA-Induced Epileptogenesis
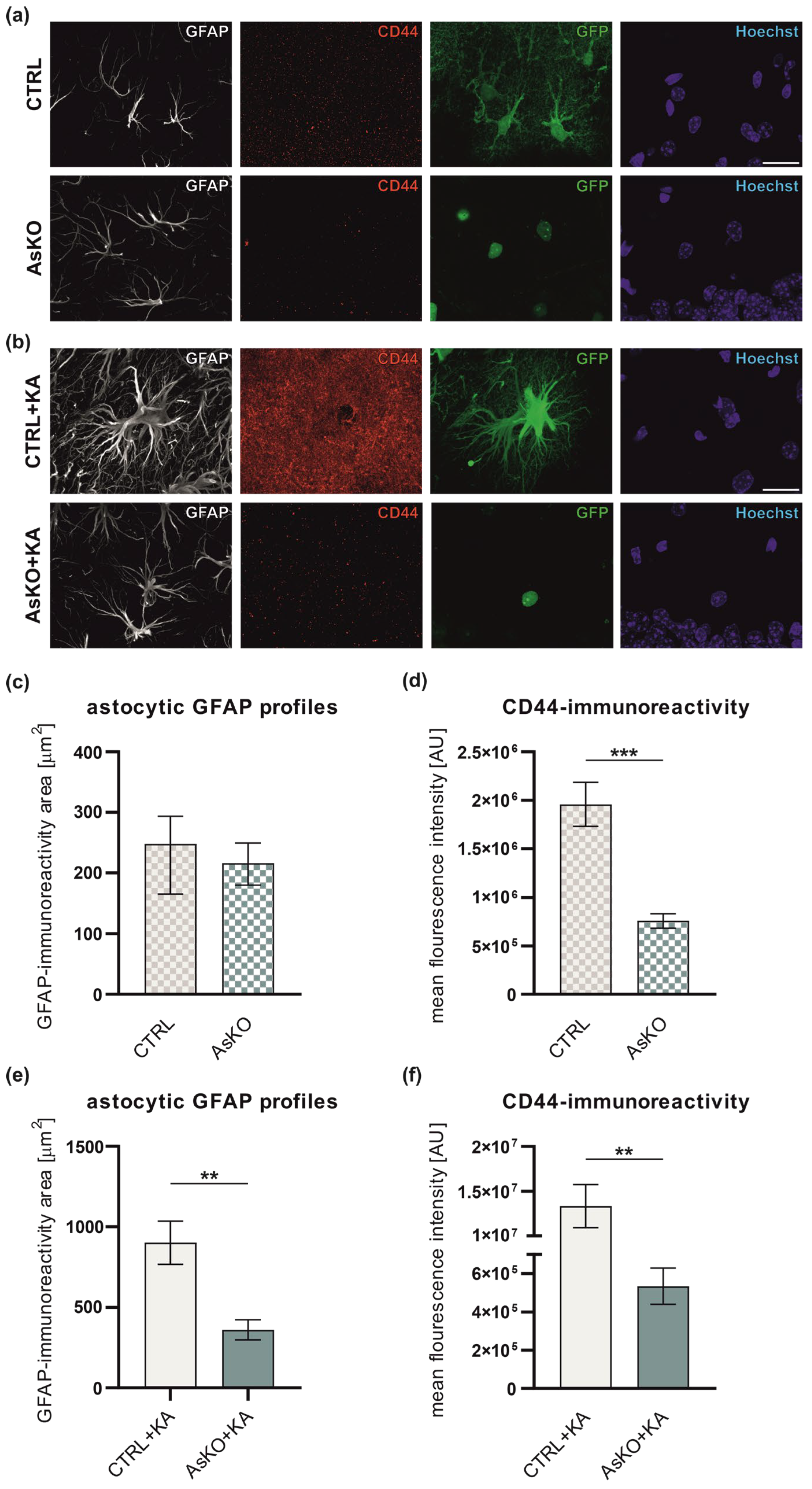
3.4. CD44 Deficiency Reduces Reactive Astrogliosis after Kainate-Induced Status Epilepticus
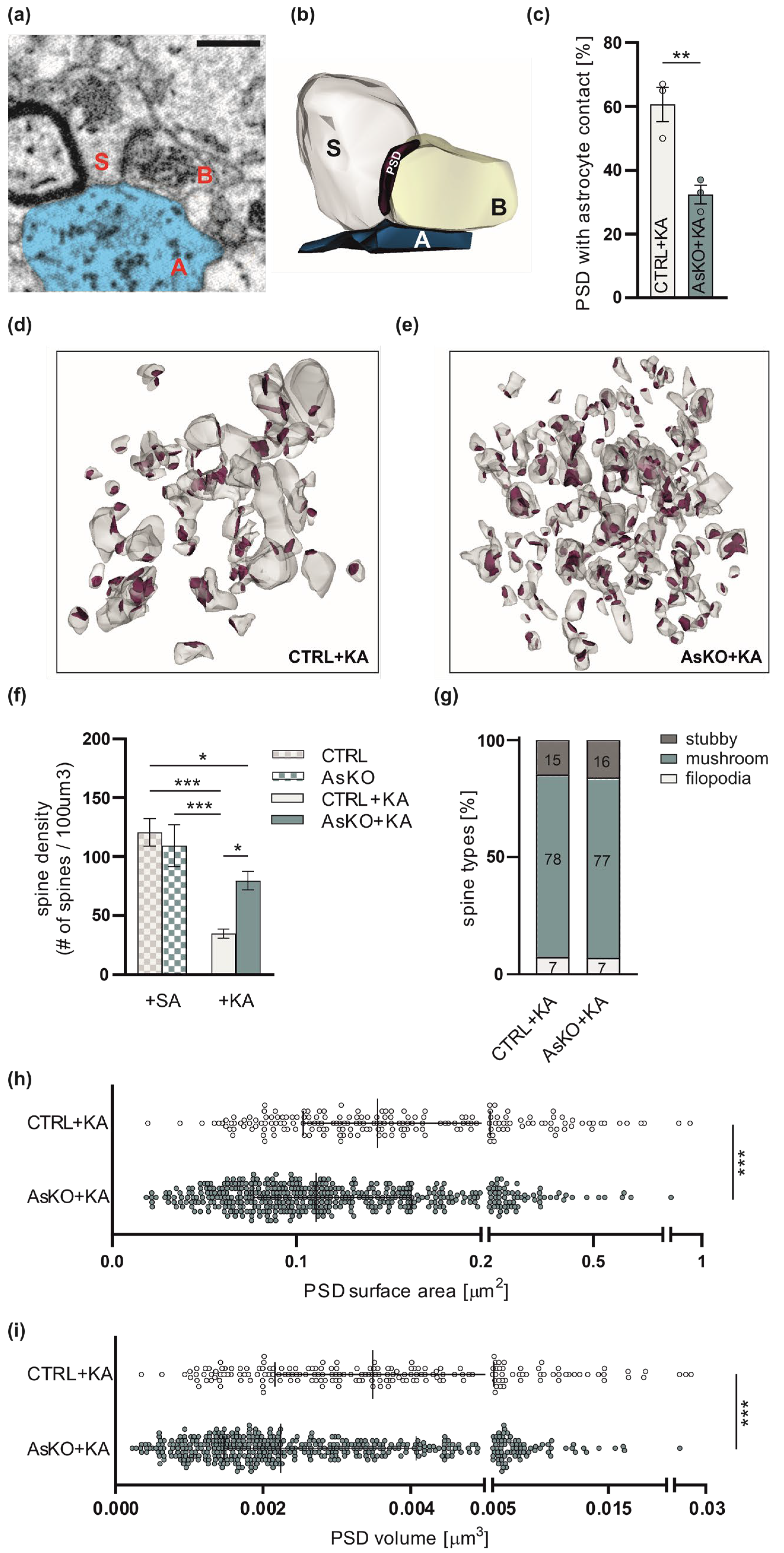
3.5. Ultrastructural Changes in Astrocyte-Synapse Interactions and Dendritic Spines in CD44 AsKO Mice Hippocampi upon Seizures
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Epilepsy. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/epilepsy (accessed on 15 January 2023).
- Chen, B.; Choi, H.; Hirsch, L.J.; Katz, A.; Legge, A.; Buchsbaum, R.; Detyniecki, K. Psychiatric and behavioral side effects of antiepileptic drugs in adults with epilepsy. Epilepsy Behav. 2017, 76, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Tian, N.; Boring, M.; Kobau, R.; Zack, M.M.; Croft, J.B. Active Epilepsy and Seizure Control in Adults—United States, 2013 and 2015. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Hayatdavoudi, P.; Hosseini, M.; Hajali, V.; Hosseini, A.; Rajabian, A. The role of astrocytes in epileptic disorders. Physiol. Rep. 2022, 10, e15239. [Google Scholar] [CrossRef] [PubMed]
- Wetherington, J.; Serrano, G.; Dingledine, R. Astrocytes in the epileptic brain. Neuron 2008, 58, 168–178. [Google Scholar] [CrossRef]
- Myer, D.J.; Gurkoff, G.G.; Lee, S.M.; Hovda, D.A.; Sofroniew, M.V. Essential protective roles of reactive astrocytes in traumatic brain injury. Brain 2006, 129, 2761–2772. [Google Scholar] [CrossRef]
- Chan, F.; Lax, N.Z.; Voss, C.M.; Aldana, B.I.; Whyte, S.; Jenkins, A.; Nicholson, C.; Nichols, S.; Tilley, E.; Powell, Z.; et al. The role of astrocytes in seizure generation: Insights from a novel in vitro seizure model based on mitochondrial dysfunction. Brain 2019, 142, 391–411. [Google Scholar] [CrossRef]
- Gonzalez Otarula, K.A.; Schuele, S. Networks in Temporal Lobe Epilepsy. Neurosurg. Clin. N. Am. 2020, 31, 309–317. [Google Scholar] [CrossRef]
- Isokawa, M. Remodeling dendritic spines of dentate granule cells in temporal lobe epilepsy patients and the rat pilocarpine model. Epilepsia 2000, 41 (Suppl. S6), S14–S17. [Google Scholar] [CrossRef]
- Ben-Ari, Y. Cell death and synaptic reorganizations produced by seizures. Epilepsia 2001, 42 (Suppl. S3), 5–7. [Google Scholar] [CrossRef]
- Magloczky, Z.; Wittner, L.; Borhegyi, Z.; Halasz, P.; Vajda, J.; Czirjak, S.; Freund, T.F. Changes in the distribution and connectivity of interneurons in the epileptic human dentate gyrus. Neuroscience 2000, 96, 7–25. [Google Scholar] [CrossRef]
- Buckmaster, P.S.; Dudek, F.E. Neuron loss, granule cell axon reorganization, and functional changes in the dentate gyrus of epileptic kainate-treated rats. J. Comp. Neurol. 1997, 385, 385–404. [Google Scholar] [CrossRef]
- Rao, M.S.; Hattiangady, B.; Reddy, D.S.; Shetty, A.K. Hippocampal neurodegeneration, spontaneous seizures, and mossy fiber sprouting in the F344 rat model of temporal lobe epilepsy. J. Neurosci. Res. 2006, 83, 1088–1105. [Google Scholar] [CrossRef]
- Bouilleret, V.; Loup, F.; Kiener, T.; Marescaux, C.; Fritschy, J.M. Early loss of interneurons and delayed subunit-specific changes in GABA(A)-receptor expression in a mouse model of mesial temporal lobe epilepsy. Hippocampus 2000, 10, 305–324. [Google Scholar] [CrossRef]
- Borges, K.; Gearing, M.; McDermott, D.L.; Smith, A.B.; Almonte, A.G.; Wainer, B.H.; Dingledine, R. Neuronal and glial pathological changes during epileptogenesis in the mouse pilocarpine model. Exp. Neurol. 2003, 182, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Li, T.; Song, X.; Sun, H.; Liu, J.; Yang, J.; Zhao, W.; Cheng, L.; Chen, H.; Liu, B.; et al. Dynamic alteration of dendrites and dendritic spines in the hippocampus and microglia in mouse brain tissues after kainate-induced status epilepticus. Int. J. Neurosci. 2021, 131, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Flores-Soto, M.; Romero-Guerrero, C.; Vazquez-Hernandez, N.; Tejeda-Martinez, A.; Martin-Amaya-Barajas, F.L.; Orozco-Suarez, S.; Gonzalez-Burgos, I. Pentylenetetrazol-induced seizures in adult rats are associated with plastic changes to the dendritic spines on hippocampal CA1 pyramidal neurons. Behav. Brain Res. 2021, 406, 113198. [Google Scholar] [CrossRef]
- Witcher, M.R.; Park, Y.D.; Lee, M.R.; Sharma, S.; Harris, K.M.; Kirov, S.A. Three-dimensional relationships between perisynaptic astroglia and human hippocampal synapses. Glia 2010, 58, 572–587. [Google Scholar] [CrossRef]
- Perea, G.; Araque, A. GLIA modulates synaptic transmission. Brain Res. Rev. 2010, 63, 93–102. [Google Scholar] [CrossRef]
- Durkee, C.A.; Araque, A. Diversity and Specificity of Astrocyte–neuron Communication. Neuroscience 2019, 396, 73–78. [Google Scholar] [CrossRef]
- Bernardinelli, Y.; Muller, D.; Nikonenko, I. Astrocyte-synapse structural plasticity. Neural Plast. 2014, 2014, 232105. [Google Scholar] [CrossRef]
- Henneberger, C.; Bard, L.; Panatier, A.; Reynolds, J.P.; Kopach, O.; Medvedev, N.I.; Minge, D.; Herde, M.K.; Anders, S.; Kraev, I.; et al. LTP Induction Boosts Glutamate Spillover by Driving Withdrawal of Perisynaptic Astroglia. Neuron 2020, 108, 919–936.e11. [Google Scholar] [CrossRef] [PubMed]
- Lushnikova, I.; Skibo, G.; Muller, D.; Nikonenko, I. Synaptic potentiation induces increased glial coverage of excitatory synapses in CA1 hippocampus. Hippocampus 2009, 19, 753–762. [Google Scholar] [CrossRef]
- Perez-Alvarez, A.; Navarrete, M.; Covelo, A.; Martin, E.D.; Araque, A. Structural and functional plasticity of astrocyte processes and dendritic spine interactions. J. Neurosci. 2014, 34, 12738–12744. [Google Scholar] [CrossRef]
- Roch, C.; Leroy, C.; Nehlig, A.; Namer, I.J. Magnetic resonance imaging in the study of the lithium-pilocarpine model of temporal lobe epilepsy in adult rats. Epilepsia 2002, 43, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Tian, G.F.; Han, X.; Peng, W.; Takano, T.; Ransom, B.; Nedergaard, M. Loss of astrocytic domain organization in the epileptic brain. J. Neurosci. 2008, 28, 3264–3276. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, C.; Smeal, R.M.; Hasenoehrl, M.G.; White, J.A.; Rubio, M.E.; Wilcox, K.S. Ultrastructural and functional changes at the tripartite synapse during epileptogenesis in a model of temporal lobe epilepsy. Exp. Neurol. 2020, 326, 113196. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, J.L.; Dalchau, R.; Fabre, J.W. Biochemical characterisation and localization in brain of a human brain-leucocyte membrane glycoprotein recognised by a monoclonal antibody. J. Neurochem. 1982, 39, 1461–1466. [Google Scholar] [CrossRef]
- Girgrah, N.; Letarte, M.; Becker, L.E.; Cruz, T.F.; Theriault, E.; Moscarello, M.A. Localization of the CD44 glycoprotein to fibrous astrocytes in normal white matter and to reactive astrocytes in active lesions in multiple sclerosis. J. Neuropathol. Exp. Neurol. 1991, 50, 779–792. [Google Scholar] [CrossRef]
- Vogel, H.; Butcher, E.C.; Picker, L.J. H-CAM expression in the human nervous system: Evidence for a role in diverse glial interactions. J. Neurocytol. 1992, 21, 363–373. [Google Scholar] [CrossRef]
- Jones, L.L.; Liu, Z.; Shen, J.; Werner, A.; Kreutzberg, G.W.; Raivich, G. Regulation of the cell adhesion molecule CD44 after nerve transection and direct trauma to the mouse brain. J. Comp. Neurol. 2000, 426, 468–492. [Google Scholar] [CrossRef]
- Roszkowska, M.; Skupien, A.; Wojtowicz, T.; Konopka, A.; Gorlewicz, A.; Kisiel, M.; Bekisz, M.; Ruszczycki, B.; Dolezyczek, H.; Rejmak, E.; et al. CD44: A novel synaptic cell adhesion molecule regulating structural and functional plasticity of dendritic spines. Mol. Biol. Cell. 2016, 27, 4055–4066. [Google Scholar] [CrossRef] [PubMed]
- Skupien, A.; Konopka, A.; Trzaskoma, P.; Labus, J.; Gorlewicz, A.; Swiech, L.; Babraj, M.; Dolezyczek, H.; Figiel, I.; Ponimaskin, E.; et al. CD44 regulates dendrite morphogenesis through Src tyrosine kinase-dependent positioning of the Golgi. J. Cell. Sci. 2014, 127 Pt 23, 5038–5051. [Google Scholar]
- Glezer, I.; Bittencourt, J.C.; Rivest, S. Neuronal expression of Cd36, Cd44, and Cd83 antigen transcripts maps to distinct and specific murine brain circuits. J. Comp. Neurol. 2009, 517, 906–924. [Google Scholar] [CrossRef] [PubMed]
- Kaaijk, P.; Pals, S.T.; Morsink, F.; Bosch, D.A.; Troost, D. Differential expression of CD44 splice variants in the normal human central nervous system. J. Neuroimmunol. 1997, 73, 70–76. [Google Scholar] [CrossRef]
- Naruse, M.; Shibasaki, K.; Yokoyama, S.; Kurachi, M.; Ishizaki, Y. Dynamic changes of CD44 expression from progenitors to subpopulations of astrocytes and neurons in developing cerebellum. PLoS ONE 2013, 8, e53109. [Google Scholar] [CrossRef] [PubMed]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Gilad, E.; Rothman, K.; Peyrollier, K. Hyaluronan-CD44 interaction with IQGAP1 promotes Cdc42 and ERK signaling, leading to actin binding, Elk-1/estrogen receptor transcriptional activation, and ovarian cancer progression. J. Biol. Chem. 2005, 280, 11961–11972. [Google Scholar] [CrossRef]
- Konopka, A.; Zeug, A.; Skupien, A.; Kaza, B.; Mueller, F.; Chwedorowicz, A.; Ponimaskin, E.; Wilczynski, G.M.; Dzwonek, J. Cleavage of Hyaluronan and CD44 Adhesion Molecule Regulate Astrocyte Morphology via Rac1 Signalling. PLoS ONE 2016, 11, e0155053. [Google Scholar] [CrossRef]
- Bausch, S.B. Potential roles for hyaluronan and CD44 in kainic acid-induced mossy fiber sprouting in organotypic hippocampal slice cultures. Neuroscience 2006, 143, 339–350. [Google Scholar] [CrossRef]
- Dzwonek, J.; Wilczynski, G.M. CD44: Molecular interactions, signaling and functions in the nervous system. Front. Cell. Neurosci. 2015, 9, 175. [Google Scholar] [CrossRef]
- Borges, K.; McDermott, D.L.; Dingledine, R. Reciprocal changes of CD44 and GAP-43 expression in the dentate gyrus inner molecular layer after status epilepticus in mice. Exp. Neurol. 2004, 188, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Tooyama, I.; Kawamata, T.; Ikeda, K.; McGeer, P.L. Morphological diversities of CD44 positive astrocytes in the cerebral cortex of normal subjects and patients with Alzheimer’s disease. Brain Res. 1993, 632, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Lipponen, A.; Paananen, J.; Puhakka, N.; Pitkanen, A. Analysis of Post-Traumatic Brain Injury Gene Expression Signature Reveals Tubulins, Nfe2l2, Nfkb, Cd44, and S100a4 as Treatment Targets. Sci. Rep. 2016, 6, 31570. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Shao, H.; Chang, L. The Important Role of Perituberal Tissue in Epileptic Patients with Tuberous Sclerosis Complex by the Transcriptome Analysis. Biomed. Res. Int. 2020, 2020, 4980609. [Google Scholar] [CrossRef] [PubMed]
- Sosunov, A.A.; Wu, X.; Tsankova, N.M.; Guilfoyle, E.; McKhann, G.M., II; Goldman, J.E. Phenotypic heterogeneity and plasticity of isocortical and hippocampal astrocytes in the human brain. J. Neurosci. 2014, 34, 2285–2298. [Google Scholar] [CrossRef] [PubMed]
- Denk, W.; Horstmann, H. Serial block-face scanning electron microscopy to reconstruct three-dimensional tissue nanostructure. PLoS Biol. 2004, 2, e329. [Google Scholar] [CrossRef]
- Shatirishvili, M.; Burk, A.S.; Franz, C.M.; Pace, G.; Kastilan, T.; Breuhahn, K.; Hinterseer, E.; Dierich, A.; Bakiri, L.; Wagner, E.F.; et al. Epidermal-specific deletion of CD44 reveals a function in keratinocytes in response to mechanical stress. Cell Death Dis. 2016, 7, e2461. [Google Scholar] [CrossRef]
- Lee, Y.; Messing, A.; Su, M.; Brenner, M. GFAP promoter elements required for region-specific and astrocyte-specific expression. Glia 2008, 56, 481–493. [Google Scholar] [CrossRef]
- Ohgane, K.; Yoshioka, H. Quantification of Gel Bands by an Image J Macro, Band/Peak Quantification Tool; Protocol IO: San Francisco, CA, USA, 2019. [Google Scholar]
- Bouilleret, V.; Ridoux, V.; Depaulis, A.; Marescaux, C.; Nehlig, A.; Le Gal La Salle, G. Recurrent seizures and hippocampal sclerosis following intrahippocampal kainate injection in adult mice: Electroencephalography, histopathology and synaptic reorganization similar to mesial temporal lobe epilepsy. Neuroscience 1999, 89, 717–729. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Schauwecker, P.E.; Steward, O. Genetic determinants of susceptibility to excitotoxic cell death: Implications for gene targeting approaches. Proc. Natl. Acad. Sci. USA 1997, 94, 4103–4108. [Google Scholar] [CrossRef]
- Ndode-Ekane, X.E.; Pitkanen, A. Urokinase-type plasminogen activator receptor modulates epileptogenesis in mouse model of temporal lobe epilepsy. Mol. Neurobiol. 2013, 47, 914–937. [Google Scholar] [CrossRef] [PubMed]
- Kimelberg, H.K. The problem of astrocyte identity. Neurochem. Int. 2004, 45, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Torres-Ceja, B.; Olsen, M.L. A closer look at astrocyte morphology: Development, heterogeneity, and plasticity at astrocyte leaflets. Curr. Opin. Neurobiol. 2022, 74, 102550. [Google Scholar] [CrossRef]
- Deerinck, T.J.; Bushong, E.A.; Ellisman, M.H.; Thor, A. NCMIR methods for 3D EM: A new protocol for preparation of biological specimens for serial block face scanning electron microscopy. Microscopy 2010, 1, 6–8. [Google Scholar]
- Sosunov, A.A.; Guilfoyle, E.; Wu, X.; McKhann, G.M., II; Goldman, J.E. Phenotypic conversions of “protoplasmic” to “reactive” astrocytes in Alexander disease. J. Neurosci. 2013, 33, 7439–7450. [Google Scholar] [CrossRef] [PubMed]
- Utkina-Sosunova, I.V.; Niatsetskaya, Z.V.; Sosunov, S.A.; Ratner, V.I.; Matsiukevich, D.; Ten, V.S. Nelfinavir inhibits intra-mitochondrial calcium influx and protects brain against hypoxic-ischemic injury in neonatal mice. PLoS ONE 2013, 8, e62448. [Google Scholar] [CrossRef]
- Fiala, J.C.; Harris, K.M. Extending unbiased stereology of brain ultrastructure to three-dimensional volumes. J. Am. Med. Inform. Assoc. 2001, 8, 1–16. [Google Scholar] [CrossRef]
- Ostroff, L.E.; Manzur, M.K.; Cain, C.K.; Ledoux, J.E. Synapses lacking astrocyte appear in the amygdala during consolidation of Pavlovian threat conditioning. J. Comp. Neurol. 2014, 522, 2152–2163. [Google Scholar] [CrossRef]
- Devinsky, O.; Vezzani, A.; Najjar, S.; De Lanerolle, N.C.; Rogawski, M.A. Glia and epilepsy: Excitability and inflammation. Trends Neurosci. 2013, 36, 174–184. [Google Scholar] [CrossRef]
- Stylli, S.S.; Kaye, A.H.; Novak, U. Induction of CD44 expression in stab wounds of the brain: Long term persistence of CD44 expression. J. Clin. Neurosci. 2000, 7, 137–140. [Google Scholar] [CrossRef]
- Tse, K.; Hammond, D.; Simpson, D.; Beynon, R.J.; Beamer, E.; Tymianski, M.; Salter, M.W.; Sills, G.J.; Thippeswamy, T. The impact of postsynaptic density 95 blocking peptide (Tat-NR2B9c) and an iNOS inhibitor (1400W) on proteomic profile of the hippocampus in C57BL/6J mouse model of kainate-induced epileptogenesis. J. Neurosci. Res. 2019, 97, 1378–1392. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Takashima, S.; Becker, L.E. CD44 expression in tuberous sclerosis. Pathobiology 2000, 68, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Pai, B.; Kuo, C.-C.; Chen, Y.-C.; Lin, C.-H.; Chen, C.-C.; Chang, Y.-J.; Hsu, Y.-C.; Huang, Y.-S.; Lin, Y.-C.; Li, H.-M.; et al. High-resolution transcriptomics informs glial pathology in human temporal lobe epilepsy. Acta Neuropathol. Commun. 2022, 10, 149. [Google Scholar] [CrossRef] [PubMed]
- Pannasch, U.; Rouach, N. Emerging role for astroglial networks in information processing: From synapse to behavior. Trends Neurosci. 2013, 36, 405–417. [Google Scholar] [CrossRef]
- Papouin, T.; Dunphy, J.M.; Tolman, M.; Dineley, K.T.; Haydon, P.G. Septal Cholinergic Neuromodulation Tunes the Astrocyte-Dependent Gating of Hippocampal NMDA Receptors to Wakefulness. Neuron 2017, 94, 840–854.e847. [Google Scholar] [CrossRef]
- Garcia-Caceres, C.; Quarta, C.; Varela, L.; Gao, Y.; Gruber, T.; Legutko, B.; Jastroch, M.; Johansson, P.; Ninkovic, J.; Yi, C.X.; et al. Astrocytic Insulin Signaling Couples Brain Glucose Uptake with Nutrient Availability. Cell 2016, 166, 867–880. [Google Scholar] [CrossRef]
- Su, M.; Hu, H.; Lee, Y.; d’Azzo, A.; Messing, A.; Brenner, M. Expression specificity of GFAP transgenes. Neurochem. Res. 2004, 29, 2075–2093. [Google Scholar] [CrossRef]
- Jessberger, S.; Parent, J.M. Epilepsy and Adult Neurogenesis. Cold Spring Harb. Perspect. Biol. 2015, 7, a020677. [Google Scholar] [CrossRef]
- Fukuda, S.; Kato, F.; Tozuka, Y.; Yamaguchi, M.; Miyamoto, Y.; Hisatsune, T. Two distinct subpopulations of nestin-positive cells in adult mouse dentate gyrus. J. Neurosci. 2003, 23, 9357–9366. [Google Scholar] [CrossRef]
- Sierra, A.; Martin-Suarez, S.; Valcarcel-Martin, R.; Pascual-Brazo, J.; Aelvoet, S.A.; Abiega, O.; Deudero, J.J.; Brewster, A.L.; Bernales, I.; Anderson, A.E.; et al. Neuronal hyperactivity accelerates depletion of neural stem cells and impairs hippocampal neurogenesis. Cell Stem Cell 2015, 16, 488–503. [Google Scholar] [CrossRef] [PubMed]
- Bonde, S.; Ekdahl, C.T.; Lindvall, O. Long-term neuronal replacement in adult rat hippocampus after status epilepticus despite chronic inflammation. Eur. J. Neurosci. 2006, 23, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Rezai Amin, S.; Gruszczynski, C.; Guiard, B.P.; Callebert, J.; Launay, J.M.; Louis, F.; Betancur, C.; Vialou, V.; Gautron, S. Viral vector-mediated Cre recombinase expression in substantia nigra induces lesions of the nigrostriatal pathway associated with perturbations of dopamine-related behaviors and hallmarks of programmed cell death. J. Neurochem. 2019, 150, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Ortinski, P.I.; Dong, J.; Mungenast, A.; Yue, C.; Takano, H.; Watson, D.J.; Haydon, P.G.; Coulter, D.A. Selective induction of astrocytic gliosis generates deficits in neuronal inhibition. Nat. Neurosci. 2010, 13, 584–591. [Google Scholar] [CrossRef]
- Wenzel, H.J.; Woolley, C.S.; Robbins, C.A.; Schwartzkroin, P.A. Kainic acid-induced mossy fiber sprouting and synapse formation in the dentate gyrus of rats. Hippocampus 2000, 10, 244–260. [Google Scholar] [CrossRef]
- Badia-Soteras, A.; Llorens-Martin, M.; Jurado-Arjona, J.; Avila, J.; Hernandez, F. Retraction of Astrocyte Leaflets From the Synapse Enhances Fear Memory. Biol. Psychiatry 2022, in press. [Google Scholar] [CrossRef]
- Pure, E.; Cuff, C.A. A crucial role for CD44 in inflammation. Trends Mol. Med. 2001, 7, 213–221. [Google Scholar] [CrossRef]
- Tsukita, S.; Yonemura, S. ERM (ezrin/radixin/moesin) family: From cytoskeleton to signal transduction. Curr. Opin. Cell Biol. 1997, 9, 70–75. [Google Scholar] [CrossRef]
- Robel, S.; Buckingham, S.C.; Boni, J.L.; Campbell, S.L.; Danbolt, N.C.; Riedemann, T.; Sutor, B.; Sontheimer, H. Reactive astrogliosis causes the development of spontaneous seizures. J. Neurosci. 2015, 35, 3330–3345. [Google Scholar] [CrossRef]
- Heja, L.; Barabas, P.; Nyitrai, G.; Kekesi, K.A.; Lasztoczi, B.; Toke, O.; Tarkanyi, G.; Madsen, K.; Schousboe, A.; Dobolyi, A.; et al. Glutamate uptake triggers transporter-mediated GABA release from astrocytes. PLoS ONE 2009, 4, e7153. [Google Scholar] [CrossRef]
- Kovacs, Z.; Skatchkov, S.N.; Veh, R.W.; Szabo, Z.; Nemeth, K.; Szabo, P.T.; Kardos, J.; Heja, L. Critical Role of Astrocytic Polyamine and GABA Metabolism in Epileptogenesis. Front. Cell Neurosci. 2021, 15, 787319. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, Z.; Skatchkov, S.N.; Szabo, Z.; Qahtan, S.; Mendez-Gonzalez, M.P.; Malpica-Nieves, C.J.; Eaton, M.J.; Kardos, J.; Heja, L. Putrescine Intensifies Glu/GABA Exchange Mechanism and Promotes Early Termination of Seizures. Int. J. Mol. Sci. 2022, 23, 8191. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.; Robitaille, R.; Volterra, A. Gliotransmitters travel in time and space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef]
- Halassa, M.M.; Haydon, P.G. Integrated brain circuits: Astrocytic networks modulate neuronal activity and behavior. Annu. Rev. Physiol. 2010, 72, 335–355. [Google Scholar] [CrossRef]
- Gomez-Gonzalo, M.; Losi, G.; Chiavegato, A.; Zonta, M.; Cammarota, M.; Brondi, M.; Vetri, F.; Uva, L.; Pozzan, T.; de Curtis, M.; et al. An excitatory loop with astrocytes contributes to drive neurons to seizure threshold. PLoS Biol. 2010, 8, e1000352. [Google Scholar] [CrossRef]
- Price, B.R.; Johnson, L.A.; Norris, C.M. Reactive Astrocytes: The Nexus of Pathological and Clinical Hallmarks of Alzheimer’s Disease. Ageing Res. Rev. 2021, 68, 101335. [Google Scholar] [CrossRef]
- Zhvania, M.G.; Ksovreli, M.; Japaridze, N.J.; Lordkipanidze, T.G. Ultrastructural changes to rat hippocampus in pentylenetetrazol- and kainic acid-induced status epilepticus: A study using electron microscopy. Micron 2015, 74, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Fiala, J.C.; Spacek, J.; Harris, K.M. Dendritic spine pathology: Cause or consequence of neurological disorders? Brain Res. Brain Res. Rev. 2002, 39, 29–54. [Google Scholar] [CrossRef]
- Chidambaram, S.B.; Rathipriya, A.G.; Bolla, S.R.; Bhat, A.; Ray, B.; Mahalakshmi, A.M.; Manivasagam, T.; Thenmozhi, A.J.; Essa, M.M.; Guillemin, G.J.; et al. Dendritic spines: Revisiting the physiological role. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 92, 161–193. [Google Scholar] [CrossRef]
- Kayser, M.S.; Nolt, M.J.; Dalva, M.B. EphB receptors couple dendritic filopodia motility to synapse formation. Neuron 2008, 59, 56–69. [Google Scholar] [CrossRef]
- Petrak, L.J.; Harris, K.M.; Kirov, S.A. Synaptogenesis on mature hippocampal dendrites occurs via filopodia and immature spines during blocked synaptic transmission. J. Comp. Neurol. 2005, 484, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Toni, N.; Teng, E.M.; Bushong, E.A.; Aimone, J.B.; Zhao, C.; Consiglio, A.; van Praag, H.; Martone, M.E.; Ellisman, M.H.; Gage, F.H. Synapse formation on neurons born in the adult hippocampus. Nat. Neurosci. 2007, 10, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Ziv, N.E.; Smith, S.J. Evidence for a role of dendritic filopodia in synaptogenesis and spine formation. Neuron 1996, 17, 91–102. [Google Scholar] [CrossRef]
- Tonnesen, J.; Nagerl, U.V. Dendritic Spines as Tunable Regulators of Synaptic Signals. Front. Psychiatry 2016, 7, 101. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruk, P.K.; Nader, K.; Skupien-Jaroszek, A.; Wójtowicz, T.; Buszka, A.; Olech-Kochańczyk, G.; Wilczynski, G.M.; Worch, R.; Kalita, K.; Włodarczyk, J.; et al. Astrocytic CD44 Deficiency Reduces the Severity of Kainate-Induced Epilepsy. Cells 2023, 12, 1483. https://doi.org/10.3390/cells12111483
Kruk PK, Nader K, Skupien-Jaroszek A, Wójtowicz T, Buszka A, Olech-Kochańczyk G, Wilczynski GM, Worch R, Kalita K, Włodarczyk J, et al. Astrocytic CD44 Deficiency Reduces the Severity of Kainate-Induced Epilepsy. Cells. 2023; 12(11):1483. https://doi.org/10.3390/cells12111483
Chicago/Turabian StyleKruk, Patrycja K., Karolina Nader, Anna Skupien-Jaroszek, Tomasz Wójtowicz, Anna Buszka, Gabriela Olech-Kochańczyk, Grzegorz M. Wilczynski, Remigiusz Worch, Katarzyna Kalita, Jakub Włodarczyk, and et al. 2023. "Astrocytic CD44 Deficiency Reduces the Severity of Kainate-Induced Epilepsy" Cells 12, no. 11: 1483. https://doi.org/10.3390/cells12111483